

Abstract
We studied the use of song types and their acoustic features in different social contexts in the banded wren (Thryothorus pleurostictus), a resident tropical songbird in which males possess about 20 distinctive song types varying in duration, bandwidth, note composition, and trill structure. We recorded six focal males intensively for four days each while we observed context information such as during versus after dawn chorus, presence of the female, counter-versus solo-singing, location at the edge versus centre of the territory, and proximity to the nest. All males used at least some song types differentially during each of these pairs of alternative contexts. Males also preferentially used the song types they shared with a given neighbour when interacting with that bird. Songs delivered during dawn chorus were significantly longer, wider in bandwidth, often compound (double songs), and more likely to contain a rattle or buzz and an up-sweeping trill, compared to songs delivered after dawn chorus. Similar features were also more commonly observed when birds were engaged in intense male-male interactions and boundary disputes after dawn chorus, especially when countersinging at the edge of the territory. The presence of the female caused the male to deliver song types with narrower whole-song and trill bandwidth and fewer rattles and buzzes, and song-type diversity and fraction of compound songs were higher when the female was present. Thus, in addition to using type matching and variations in song-type switching and diversity to signal different levels of aggressive intention, male banded wrens also select song types based on their acoustic structure in different social contexts.
In approximately 75% of oscine songbird species, male singers possess repertoires of two or more distinct song types acquired by innovation or imitative learning (MacDougall-Shackleton 1997). Some studies have suggested that each song type conveys the same message and that the primary function of song types is to increase stimulus diversity, reduce habituation of the receiver, and/or relieve stress on syringeal muscles (Nottebohm 1972; Hartshorne 1973; Krebs et al. 1978; Searcy et al. 1982; Lambrechts & Dhondt 1988; Searcy 1992). However, studies on a growing number of species indicate that song types are used selectively in different contexts and therefore probably convey different messages (reviewed in Catchpole and Slater 1995).
Context-specific use of song types was first described in the wood warblers (Parulinae), where males were found to possess two acoustically distinct categories of song types apparently specialized for use in intersexual versus intrasexual communication (reviewed in Spector 1992). Type I songs are sung during daylight, relatively slowly, and when near females or by unmated males, suggesting a mate attraction function. Type II songs are sung rapidly in a dawn bout and later in the day during encounters with rival males or while close to territory boundaries, suggesting a territorial function. These associations between song types and contexts are only probabilistic - singers may use Type II songs during male encounters, and some investigators have argued that the different song types all serve a territorial function but encode variation in “mood” or aggressive tendencies (Lein 1978; Morrison & Hardy 1983). Two studies measured the acoustic characteristics of song types used in different contexts, and found that Type II songs were lower in frequency and more complex in structure and song delivery pattern (Spector 1991; Staicer 1996).
An association between certain song types and different social contexts (e.g. presence of female, or aggressive interaction) has also been found in several non-paruline species (Baptista 1978; Smith et al. 1978; Järvi et al. 1980; Schroeder & Wiley 1983; Catchpole 1983; Cozens & Falls 1984; Gaddis 1986; Johnson 1987; Derrickson 1987; Nelson & Croner 1991; Dabelsteen 1992; Luschi & del Seppia 1996; Wiebe & Lein 1999; Duguay & Ritchison 1999). Some level of differential song-type use may turn out to be fairly common in species with repertoires. In none of the studies mentioned above has the acoustic structure of song types used in different contexts been analyzed quantitatively and related in a functional way to the type of information conveyed in the alternative contexts.
Although defining the context in which a communication signal or behaviour occurs is a key to understanding the probable function of the signal (Smith 1965), identifying the correct context is not always straightforward. For example, different contexts such as time of day, stage of breeding, location and presence of male or female conspecifics, can be correlated with each other, confounding the efforts to find the true context. The methods for analyzing these relationships have become increasingly more sophisticated and robust. To test hypotheses of association rigorously, many independent observations of occurrences of the behaviours must be accumulated in an unbiased manner, and both the behaviours and contexts must be carefully defined and classified. Correlations and interactions among context variables can now be identified and statistically controlled with the use of multivariate statistical models (e.g., Byers 1996). Moreover, collection of data needed to relate contexts and signals generally requires serial recordings and repeated observations on known focal individuals, which can result in pooling artefacts. Fortunately, powerful yet conservative statistical methods are now available for dealing with unbalanced and non-independent repeated measures on the same individuals (Littell et al. 1996).
We examined the issue of context-specific song-type use and song structure in the banded wren Thryothorus pleurostictus, a resident tropical songbird with repertoires of around 20 song types per male. Males share about 75% of their song types with adjacent neighbours. Banded wrens are usually immediate variety singers, switching to a new song type after each song, but sometimes repeat a song type several to many times. Prior work has shown that males reduce the diversity of song types used and their switching rate during the course of the morning following dawn chorus, and during countersinging interactions with neighbours (Molles & Vehrencamp 1999). Playback studies have corroborated the more aggressive context of repeat-mode singing (Molles 1999). Banded wren males also type match each other, and playback studies have shown that a type-matching stimulus elicits a stronger aggressive approach response than repertoire matching (singing a shared but not matching type). Moreover, singing an unshared song type seems to be a signal of retreat (Molles & Vehrencamp 2001).
In preliminary observations, we noticed that males delivered more vigorous-sounding songs during the dawn chorus and during aggressive interactions, whereas shorter, weaker-sounding songs seemed to predominate later in the morning. In this study we asked whether specific song types were used in different social contexts with a series of non-exclusive questions: 1) Do males use their song types differentially in different social contexts, such as during/after dawn chorus, edge/centre of territory, presence/absence of the female, proximity to the nest, and when counter/solo singing? 2) If males do show some differential song-type use, do all males associate the same song types with the same contexts? 3) Are there specific structural acoustic features of the song types, such as duration, frequency characteristics, or note and trill structure that underlie the differential use of song types? 4) Do males use a different subset of song types when interacting with different neighbours?
METHODS
Study Site and Subjects
The study was conducted from 5 June to 3 August 2000 in Santa Rosa National Park, Guanacaste, Costa Rica (10° 51’ N, 85° 38′ W). The habitat consists of tropical dry deciduous forest, mixed with occasional patches of regenerating secondary growth. We have been banding and monitoring banded wrens since 1996 in two neighbourhoods separated by 0.5 km. The Santa Rosa population contains unbroken mature forest that supports a stable, high-density population of banded wrens. The Cerco de Piedra population inhabits a strip of forest surrounded by areas of secondary growth. This population exhibits a higher turnover of males and occasional unoccupied areas. See Molles & Vehrencamp (1999) for a more detailed description of these two areas and of their differences in song sharing patterns.
Because banded wren males possess a substantial number of song types, it was necessary to record each bird intensively for long periods of time throughout the day and on multiple days throughout the breeding season to obtain a good sample of their song types, including rarely sung ones, and no empty cells in our factorial context combinations. This objective necessarily resulted in a trade-off with the number of birds we could study. Six mated territorial males, three birds from each site, served as subjects for this study. In each site we selected two males known to have defended the same territory area at least one year prior to the study, and one newcomer male defending a territory in the study area for the first year. We chose males that were at least two territories apart from each other so that the focal birds would have independent sets of neighbours (Fig. 1a, b). One of the subjects disappeared from his territory after the first day of recording. This territory and the resident female were quickly taken over by another male (T) that we had banded two years earlier in a nearby area. Although new to the territory, this bird was known to be older and a resident of the local population, so we continued our monitoring of this territory with the new male and excluded the first day of recording on the prior male. Table 1 summarizes the neighbourhood, age, repertoire size, mean song sharing with neighbours, and sampling information for each of the six subjects. Nesting activity was also monitored for adjacent neighbours and all other territorial males in the two study areas.
Figure 1.
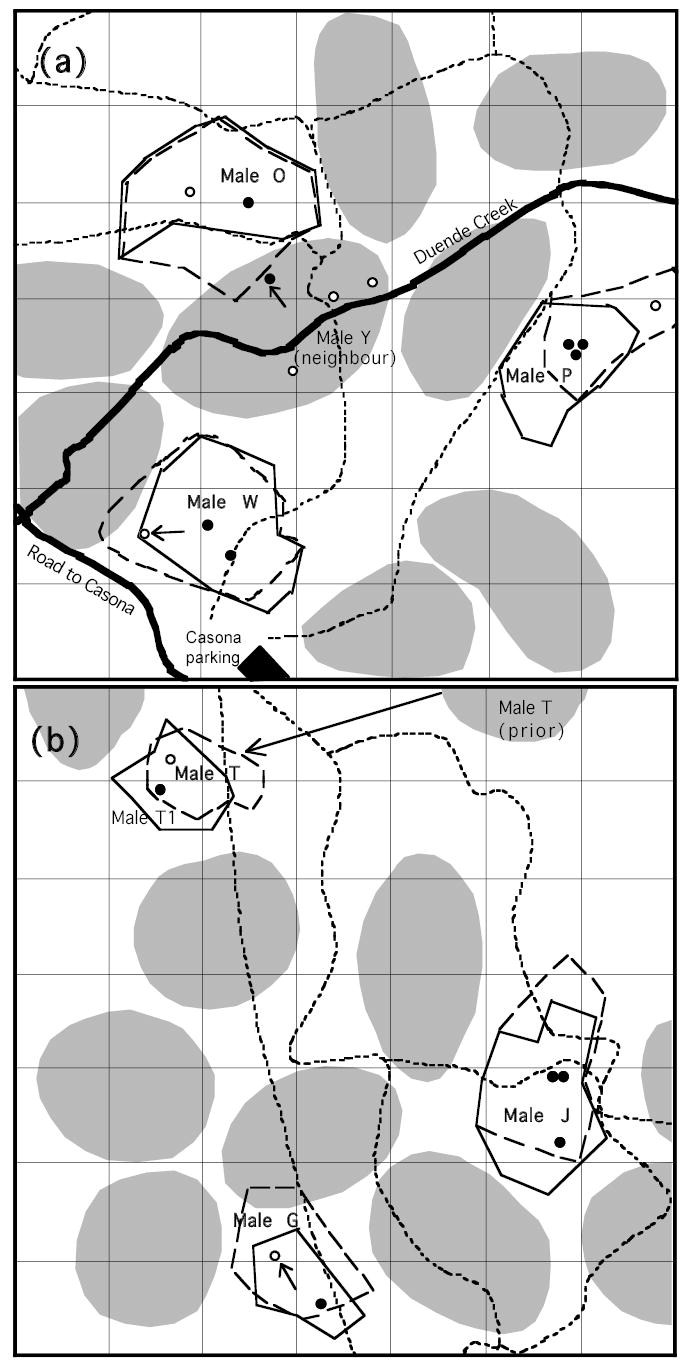
Maps of study areas showing first and last days’ territory boundaries and sequential nest site locations for the six focal males in the Santa Rosa population (a) and the Cerco de Piedra population (b). The solid black outlines shows the initial territory area determined from focal bird activity on the first day of observation, and black dots show nest locations in the same period. The dashed outlines and white dots show the territory boundaries and nest locations toward the end of the season. The approximate locations of neighbour territories are shown in grey ovals. The four nest sites for Male O’s neighbour Y are also shown in colors associated with male O’s boundary positions; male Y’s boundary nest caused the boundary shift between these two males. Arrows indicate the nests that caused boundary shifts and the direction of the shift. Male T1 occupied Male T’s territory on the first recording day. Each square is 50 m on a side. Trails are drawn with thin dashed lines, and streams, roads and buildings are also indicated.
Table 1.
Summary of focal male characteristics (means given with ±SE)
| Male: | O | W | P | G | T | J |
|---|---|---|---|---|---|---|
| Site | SR | SR | SR | CP | CP | CP |
| Territory size (ha) | 0.633 | 0.516 | 0.328 | 0.283 | 0.199 | 0.445 |
| Year on territory | 5 | 2 | 1 | 2 | 0 | 1 |
| Presumed age | old | old | young | old | old | young |
| Repertoire size (single song types only) | 24 | 26 | 23 | 17 | 25 | 20 |
| Fraction single song types shared w/neighbours | 0.740 | 0.769 | 0.667 | 0.824 | 0.733 | 0.725 |
| Mean song duration (sec) (single song types only) | 3.06 ±0.162 | 3.07 ±0.157 | 3.01 ±0.180 | 3.21 ±0.236 | 3.26 ±0.160 | 2.82 ±0.195 |
| Mean song bandwidth (Hz) (single song types only) | 5215 ±436 | 5144 ±319 | 4968 ±421 | 5023 ±451 | 5145 ±339 | 4503 ±377 |
| % rattle-buzz song types | 37.5 | 42.3 | 36.4 | 41.2 | 37.5 | 42.9 |
| No. compound song types | 6 | 8 | 5 | 9 | 12 | 4 |
| Dawn chorus song rate (single songs/min) | 4.88 ±0.82 | 5.95 ±0.39 | 4.77 ±1.32 | 4.09 ±1.00 | 5.87 ±1.62 | 5.28 ±0.55 |
| Total songs recorded | 1744 | 1307 | 1081 | 1734 | 1001 | 1317 |
Focal Male Monitoring
Each focal individual was recorded for four days during the season (three days for male T). Successive recording days were separated by 10 to 20 days. We tried to record each male during each stage of breeding (nest construction/egg laying stage, incubation stage, and nestling/fledging stage), but we never found a nest with eggs for one male and two other males were unable to reach the nestling stage. Observations began at 0500 and ended at 1300 hours, with two 1-hour breaks for the observer interspersed at different times on subsequent observation days so that all time periods between 0500 and 1300 hours were equally covered. During the 1–2 hour observation sessions, the vocal behaviour of the focal males was continuously recorded on a Tascam DA-P1 digital tape recorder and an Audio-Technica AT4071 directional microphone with foam windscreen. At the beginning and end of each recording session we corroborated the identity of the focal bird by its colour bands. We noted specific variables related to each observation day such as changes in the nest location, stage of nesting, and territory borders. Nesting activity was also monitored between the intensive recording days. During each recording session, we followed the focal bird as closely as possible without disturbing it while it moved through its territory. Movements around the territory and other behaviours observed with the aid of binoculars were recorded instantaneously by quietly announcing them into the microphone.
For each song delivered by the focal bird, the following context variables were noted:
Location on the territory – Singing from the edge versus centre of the territory. The bird had to be within 5 m of the boundary to qualify as edge, and neighbour ID at that border was also noted. See below for method of determining boundaries.
Distance from the nest – singing close to versus far from nest. Bird had to be within a radius of 4 m around the nest to qualify as close, and singing beyond 4 m was classified as far. Observations on birds with no known nest site were classified as unknown and omitted from analyses involving nest distance.
Female – singing with female present versus absent. Female had to be either seen or heard to be classified as present. All cases where the female was not detected were classified as female absent. Although females sing much less often than males and may have been present but undetected, we found that they usually made some type of sound that signalled to both their mate and the observer that they were close by.
Singing state – countersinging with neighbours versus solo singing. Countersinging was noted if a song by the focal bird was preceded or followed within 2–3 sec by the song of a neighbour. If no other birds were singing or there was no coordination of singing, songs were classified as solo singing. Cases in which we were unsure of the classification were scored as uncertain and omitted from analyses involving singing state.
Time of day. Recording start and end times were noted so that exact times for all songs could be computed from the tape’s time log.
Interactions. We noted all close encounters and physical fights between the focal male and his neighbours, as well as any other interactions with conspecific intruders, other bird species, and nest predators.
Territory and Nest Site Locations
On every focal observation day we assessed the vocally-defended territory boundaries of the focal male by flagging the peripheral song posts. During the later portion of dawn chorus, banded wren males typically approach each side of their territory and sing from favorite high perches while facing and directing their songs toward the closest neighbour. These peripheral perch positions generally defined the actively defended area on a given day, although any vocal activity beyond these perches later in the day expanded our estimate of the territory boundaries. Boundaries with some neighbours were more actively contested and easier to define than others, and this boundary activity often shifted to different neighbours during the season. Undefended but occasionally used areas occurred throughout the study area as defended areas changed. We mapped the initial territory boundaries for each male after the first and last observation days (Fig. 1). Cartesian coordinates of 10 to 12 boundary points per territory were measured using a Garmin GPS 100 Survey II meter with a GA28 external antenna over a 5–10 min period per point to maximize location accuracy. Nest locations were similarly measured for all nesting attempts of our focal birds and their neighbours. Territory areas were measured for the initial (I) and later (L) sets of territory boundaries and the common overlap area (O) for each male using the polygon area routine in Matlab (Math Works, Inc.) and percent change in territory location during the season was computed as [1 − (O/I + O/L)/2] * 100.
Statistical Analyses
We constructed three kinds of databases for the analyses presented here. The primary database, called the SongEvent file, consisted of an entry for each song extracted from the original tape recordings. Compound songs (two or more discrete song types sung together), incomplete songs (generally missing the last half of the song), and unidentifiable songs by the focal bird were included as song events. Entries associated with each song included: focal bird ID, time of day, song type if known, complete or incomplete, compound or single, location on the territory, distance from the nest, female presence, number of singing neighbours, singing state, neighbour ID if the focal male was countersinging or at a boundary, song types sung by neighbours if discernable, and occurrence of song-type matching and song overlapping. Since singing behaviour, male movements, and interactions with neighbours and mates are fundamentally different in the first hour of the day compared to later (Burt & Vehrencamp 2005), we split time of day into two periods: during dawn chorus (before 0600 hr) and after dawn chorus (after 0600 hr).
Using spectrograms generated with Syrinx sound analysis software (J. M. Burt: http://syrinxpc.com) with settings FFT size = 512 and Blackman window type, we characterized the acoustic features of each song type by measuring of song duration, whole-song bandwidth (maximum frequency minus minimum frequency), trill duration, trill bandwidth, and trill note rate, of three examples of each males’ song types and entered an average of these values in the file for each complete song event of that type; compound songs were also measured in this way. Because of the known production constraint trade-off between note rate and bandwidth of trilled songs (Podos 1997), we derived an index of trill vigour by multiplying the standardized scores of these two parameters. Each identified song was scored as possessing or lacking a rattle or buzz in the introduction. These two elements are broadband, harsh-sounding, approximately 1-sec long notes, clearly different from the other tonal frequency-modulated notes in the songs (see Fig. 2 for examples). Finally, the terminal trill of each song was classified into one of three shape categories based on the direction of the frequency sweep: up-modulated, down-modulated, hook-shaped (chevron) notes, and flat-component notes (see Fig. 2 for examples).
Figure 2.
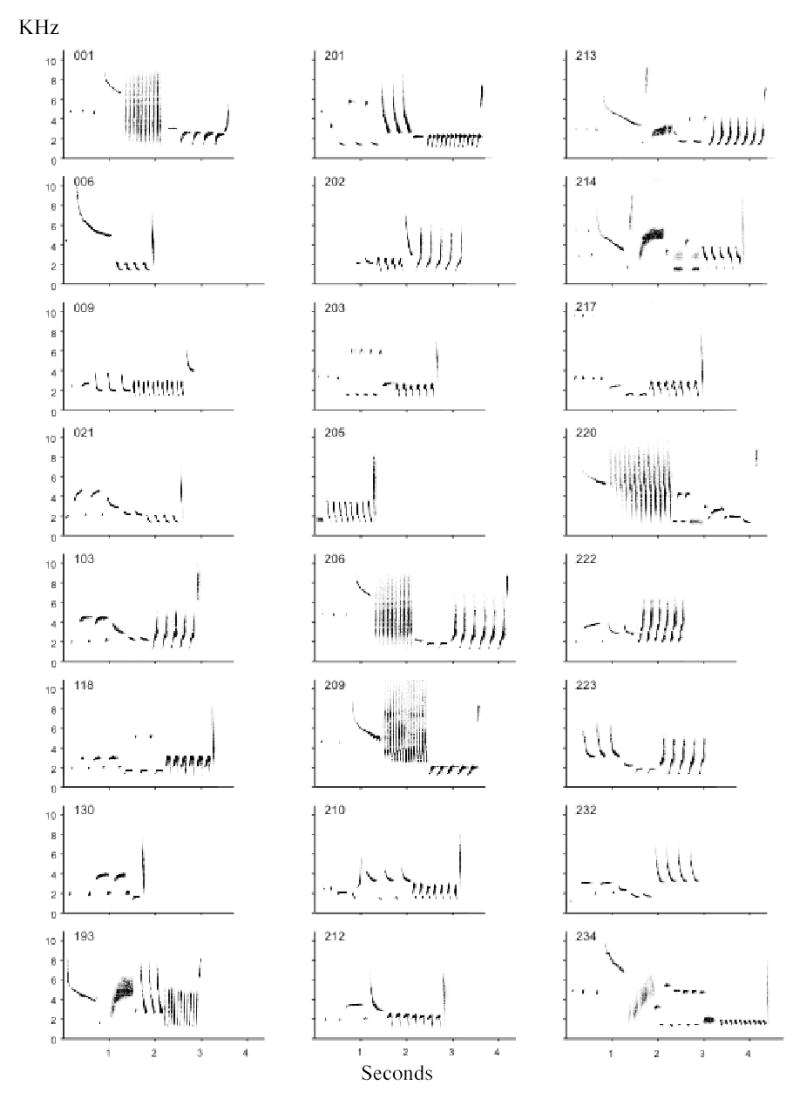
Single-song repertoire of focal bird O, compound songs not shown. Spectrogram of each song type identified by number. Rattles can be seen in song types 1, 206, 209, and 220, buzzes in song types 193, 213, 214, and 234. Song types with up-sweeping trill notes include 103, 201, 202, 206, 213, 222, 223; down-sweeping trills include 6, 9, 21,193, 205, 214, 232, 234; hook-note trills include 118, 203, 210, 212, 217; flat trills include 1 and 209; types 130 and 220 have no trill.
For analyses involving song type, a second database called the SongType file was generated from the SongEvent file by splitting the compound songs into their respective single types. All unidentified song types were eliminated from this file, as well as the song measurements. The contextual information was retained. A third database was generated based on bouts of singing. Initially, sequences of songs were segregated into bouts if there was a time gap of 2.0 or more minutes between successive song events. These bouts were further subdivided if: the focal bird moved between centre and edge locations or to different boundaries; the male moved close to or away from the nest; the male shifted between countersinging and solo singing; or the female appeared or left. Thus each bout was assigned to one level for each of the context variables, including whether it occurred during or after dawn chorus. Bouts shorter than three songs were not considered. For each bout we computed song rate, song types per song delivered, and song-type switches per songs in the bout minus 1. Compound songs were treated as two separate songs for the latter two measures.
Earlier studies of context-dependent song-type usage employed contingency table analysis to evaluate the association between song types and social context categories. Such analyses contain three inherent problems: 1) counts of songs or bouts delivered in a temporal series are not independent events; 2) counts from individual males were often pooled; and 3) each level of social context may be confounded by other contexts (e.g., male encounters could be positively or negatively associated with female presence, time of day, etc.). All of our analyses dealt with problem 3 by simultaneously including up to four categorical social context variables and all interaction terms between them in a multivariate model. Manual stepwise removal of non-significant interaction terms (alpha value greater than 0.05) was then undertaken to find the model containing the maximum number of significant terms for each dependent variable. Note that all of our context variables contained just two levels apiece, and each level of any given variable could co-occur with either level of all other variables. This categorical method of defining social contexts enabled us to quantify any interactions among contexts. Either song events or bouts were the observation units in all of these analyses.
Our method of dealing with problems 1 and 2 depended on whether the dependent variable was nominal or continuous. For analyses involving continuous dependent variables such as song duration, bandwidth, and the bout variables, we ran repeated measures ANOVA using the PROC MIXED procedure of SAS/STAT v8.2 (SAS Institute, Inc.; Littell et al. 1996). This procedure corrects for bird differences with the declaration of bird ID as the repeated measures subject. The non-independence of sequential song events was corrected for by specifying a one-lag autoregressive covariance structure AR(1). When no explicit time variable is included in the model, subsequent temporal events must be ordered in sequential rows of the dataset, and PROC MIXED removes the autocorrelated effect of the prior song or bout. We re-ran the final model with different covariance structures, such as variance components (VC) and compound symmetry (CS), to verify that the autoregressive covariance structure gave the best fit as measured using the Aikaike’s Information Criterion (AIC) provided by PROC MIXED (it always did). We avoided including two strongly collinear terms in the same model. For example, location on the territory and distance from the nest were highly confounded – only three birds had nests that were close enough to their territory boundary to sing occasionally both close to the nest and at the territory edge. We therefore first built models using the independent context variables time of day, singing state, female, location on the territory, and their interactions, and then ran a second series of analyses substituting distance from the nest for location on the territory to determine if distance from the nest had any additional effects. Repeated measures ANOVA provides a very conservative test, with the denominator degrees of freedom for the F test of each term in the final model reflecting the number of subjects. Specific contrasts were tested for significance in a similarly conservative way using a t-test of the least-squares means difference. We found that most of the birds must show an effect in the same direction for it to be significant.
Three of our acoustic measurement variables that possessed just two levels (presence/absence of rattle or buzz, complete/incomplete, and compound/single) could also be analyzed with the same rigor as the continuous variables using the GLIMMIX macro in SAS (Littell et al. 1996). This shell program specifies a logit link function for modelling such binomial data. Analyses for these variables therefore corrected for interaction effects, repeated measures effects, and autocorrelated events.
For analyses of nominal dependent variables with three or more levels, such as trill type or song type, we used the nominal logistic regression procedure from JMP 5.0 (SAS Institute, Inc.). Song type was examined separately for each male, which avoided pooling problems, and rare song types given fewer than 10 times were omitted from the analysis. To examine trill type (four levels) for all males pooled, we added bird ID and the interaction between bird ID and all of the other context terms to the model, and declared these terms containing bird ID as random effects. This type of model produces a random block design that approximates the repeated measures ANOVA for continuous variables. During the stepwise elimination process, both the fixed effect term and its random equivalent containing bird ID were removed. There is no statistical method of temporal autocorrelation control with nominal logistic regression. Because this species primarily sings with immediate variety, each song reflects an independent choice by the singer, and even repetitions of the same song represent strategic signalling behaviour (Molles 1999), so we did not lump repeated song types into one event. We report the P-values from the Wald χ2 tests for each context term in the final model.
Statview 5.0 (SAS Institute, Inc.) was used for all other basic statistical analyses. Simple contingency table analyses were conducted as a descriptive technique to identify under- and over-represented song types in different contexts (rare song types removed). The Phi coefficient from a 2 by 2 contingency table analysis of two context variables is a useful measure of the degree of association between the variables that can be interpreted like a correlation coefficient (0 ≤ Phi ≤ 1.0). All continuous variables used in parametric analyses were transformed as needed to produce normally-distributed residuals.
RESULTS
The Composition of a Bird’s Repertoire
The single-song repertoires of the six focal birds averaged 22.5 songs (range 17 to 26, Table 1). Figure 2 illustrates all of the single song types used by one of the birds. Each song type contains a unique combination of introductory notes, middle notes or syllables (usually repeated a few times), a terminal trill, and occasionally a tick note at the end. The same element may be found in several different song types. For example, types 1, 206, and 209 have the same introductory note, types 234 and 203 have the same middle notes, and 103 and 223 have the same trill. Complete songs range from 1.32 to 5.85 sec in duration (all birds). In addition, each bird has several favourite song types that it prefers to combine into compound songs. Types 1, 6, 21 and 193 are frequently combined with other types. Compound songs are significantly longer than single songs (up to 8.21 sec).
Context and Differential Song-type Use
Although any level of each of our five context variables could co-occur in principle with either level of any other context variable, there were some associations among the context variables as shown in Table 2. For example, during the dawn chorus birds were more likely to countersing at the edge of their territory far from the nest, and females were more likely to be absent. Not surprisingly, females were more likely to be present when the male was singing near the nest. Distance from the nest and location on the territory were very strongly confounded, as mentioned in the methods section.
Table 2.
Association matrix for context variables1
| During dawn chorus | Countersinging | Female present | Territory edge | |
|---|---|---|---|---|
| Countersinging | 0.209 (+) | |||
| Female present | 0.073 (−) | 0.082 (+) | ||
| Territory edge | 0.087 (+) | 0.051 (+) | 0.104 (−) | |
| Close to nest | 0.141 (−) | 0.060 (−) | 0.258 (+) | 0.392 (−) |
Table shows Phi coefficient of association (effect size) for each 2 by 2 contingency table comparing pairs of dichotomous contexts. Values of Phi can range from 0 (no association) to 1 (perfect association). A plus (+) or minus (−) indicates whether the observed counts of the listed levels of the variables were greater or less than expected, respectively.
To determine whether birds differentially used their song types as a function of context (question 1) we first performed separate nominal logistic regression analyses on each bird using the SongType database. The initial models included the four main effects (time of day, location on the territory, female presence, and singing state), as well as the pairwise interaction terms between each of these main effects; song type was the dependent variable. Results are shown in Table 3. All males exhibited highly significant differential song-type use in all contexts, usually in the main effect, although sometimes in an interaction with another context. An examination of the individual contingency tables for each bird and context indicated that only a few song types were strongly over- or underused in each context. Nearly all song types were delivered at some time in the two alternative levels of each context, so there was no evidence for exclusive use of subsets of songs types in any context.
Table 3.
Results of nominal logistic regression of song types versus all context main effects and two-way interactions conducted on each bird separately1
| Male
|
||||||
|---|---|---|---|---|---|---|
| Terms | O | W | P | G | T | J |
| Time of day | <0.001 | <0.001 | <0.001 | 0.947 | <0.001 | 0.622 |
| Singing state | 0.056 | <0.001 | 0.028 | 0.100 | 0.021 | <0.001 |
| Location on territory | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.103 |
| Female presence | <0.001 | 0.001 | 0.001 | <0.001 | <0.001 | 0.121 |
| Time*Territory | --- | --- | --- | --- | --- | 0.003 |
| Time*Female | --- | --- | --- | --- | --- | 0.022 |
| Singstate*Female | 0.002 | --- | --- | --- | --- | --- |
| Territory*Female | --- | --- | --- | 0.011 | --- | --- |
| df | 22 | 25 | 18 | 16 | 24 | 18 |
| N | 1685 | 1299 | 910 | 1590 | 908 | 1145 |
Table shows P values of variables in the final model, significant terms indicated in bold.
We next examined whether different birds tended to prefer the same song types in the same contexts (question 2). Eliminating song types that were given by fewer than three birds, a nominal logistic regression analysis was run on all birds pooled. The model included main effects, bird ID, and the interaction terms between bird ID and each main effect. The main effects were all highly significant (time: χ232 = 122.1, P < 0.0001; territory: χ232 = 62.5, P < 0.0001; female: χ232 = 47.6, P = 0.0019; and singing state: χ232 = 66.6, P < 0.0001). Examining the contingency table analyses for these shared song types, only a few types were consistently over- or underused relative to expected values in a given context by all or most of the males. Types 203 and 206 were consistently overused during dawn chorus, type 210 was consistently overused at the edge of the territory, type 202 was consistently overused during countersinging, and type 130 was strongly overused by birds solo-singing from the centre of their territories. Males preferentially sang types 1, 6, 118, and 212 when the female was present, while consistently underusing types 217, 220, and 307 in this same context. Other song types were preferentially used in different contexts by the individual males, or used according to expectation.
Context and Quantitative Song Features
Given that song types are used differentially, we asked whether the structural acoustic features of these songs were associated with different contexts (question 3). Some of the acoustic features were correlated with each other (Table 4). Each quantitative song measure from the SongEvent database, and the three bout-based song delivery pattern variables, served as the dependent variable in separate repeated measures analyses. The final models, after removal of non-significant terms, are summarized in Table 5. Trill note rate was not significantly associated with any context and is therefore not included in the table. The derived trill index was highly correlated with one of its components, trill bandwidth, and yielded a very similar final model, so the redundant results for trill index are also not shown in the table.
Table 4.
Correlation matrix of continuous song variables
| Song duration | Song bandwidth | Trill duration | Trill bandwidth | Trill rate | Trill index | |
|---|---|---|---|---|---|---|
| Song bandwidth | 0.544*** | |||||
| Trill duration | 0.291*** | 0.118 | ||||
| Trill bandwidth | 0.158* | 0.139 | 0.411*** | |||
| Trill rate | 0.078 | 0.063 | −0.039 | −0.089 | ||
| Trill index | 0.167* | 0.142 | 0.235** | 0.576*** | 0.763*** | |
| Rattle-buzz | 0.470*** | 0.528*** | 0.168* | 0.093 | −0.017 | 0.047 |
Table shows Pearson correlation coefficients, N =179;
* P < 0.0001,
P < 0.01,
P < 0.05
Table 5.
Final mixed models for each song variable, showing P-values from F or Wald χ2 test (trill type only) for each main effect and interaction term in the model. Significant terms highlighted in bold, direction of effect shown in parentheses. Trill abbreviations Dn=Down, Hk=Hook. For all models, numerator df = 1 and denominator df = 5. Total sample size of bouts or songs shown in the bottom row. Terms eliminated during stepwise removal procedure indicated by “---”.
| Term in model | Song duration | Song bandwidth | Trill duration | Trill bandwidth | Rattle or Buzz | Compound | Complete | Trill type | Song rate | Switch rate | Type diversity |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Time of day(DuringDC/After) | 0.002 (D>A) | 0.001 (D>A) | --- | 0.776 | 0.004 (D>A) | 0.001 (D>A) | 0.002 (D>A) | 0.015 (D-Up) | 0.033 (D>A) | 0.003 (D>A) | --- |
| Singing state (Countersing/Solo) | 0.006 (Cs>S) | 0.055 (Cs>S) | 0.041 (Cs>S) | --- | 0.530 | 0.025 (Cs>S) | 0.009 (Cs>S) | 0.020 (Cs-Up) | --- | 0.004 (Cs>S) | --- |
| Territory location (Edge/Centre) | 0.006 (E>C) | 0.030 (E>C) | 0.011 (E>C) | 0.499 | --- | 0.049 (E>C) | --- | 0.024 (E-Dn) | --- | --- | --- |
| Female presence (Present/Absent) | --- | 0.044 (P<Ab) | --- | 0.036 (P<Ab) | 0.017 (P<Ab) | 0.041 (P>Ab) | --- | 0.177 | 0.030 (P<Ab) | --- | 0.043 (P>Ab) |
| Time*Singstate | --- | --- | --- | --- | 0.041 (Aft:Cs>S) | --- | --- | 0.012 Aft:Cs-Up | --- | --- | --- |
| Time*Territory | --- | --- | --- | 0.021 (Aft:E>C) | --- | --- | --- | --- | --- | --- | --- |
| Time*Female | --- | --- | --- | 0.138 | --- | --- | --- | 0.033 (P-Hk) | --- | --- | --- |
| Territory*Singstate | 0.018 (E:Cs>S) | 0.024 (E:Cs>S) | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| Territory*Female | --- | --- | --- | 0.548 | --- | --- | --- | --- | --- | --- | --- |
| Time*Terr*Female | --- | --- | --- | 0.014 (P<Ab) | --- | --- | --- | --- | --- | --- | --- |
| Total sample n | 7098 | 7098 | 7098 | 8184 | 7098 | 7098 | 7098 | 6876 | 708 | 708 | 708 |
Songs delivered during versus after dawn chorus showed the strongest structural differences. During dawn chorus songs were significantly longer, greater in bandwidth, more likely to contain a rattle in the introductory part and an upsweeping J-note trill at the end, more likely to be compound songs, and delivered as complete songs, compared to songs given after dawn chorus (Figs. 3 and 4). The longer duration of dawn songs was only slightly affected by the prevalence of compound and complete songs. When compound and incomplete songs were omitted from the analysis, the relationship between time of day and duration was still very significant (P = 0.0113 versus P = 0.0019), meaning that longer single song types were also more commonly selected during dawn chorus. Although many songs with dawn chorus characteristics were also given after dawn chorus, song types that were shorter and lacked rattles and buzzes (e.g., types 1, 6, 9, 21, and 130), as well as incomplete songs, predominated later in the morning. Song rate and switching rate were also higher during dawn chorus, corroborating prior results (Molles & Vehrencamp 1999).
Figure 3.
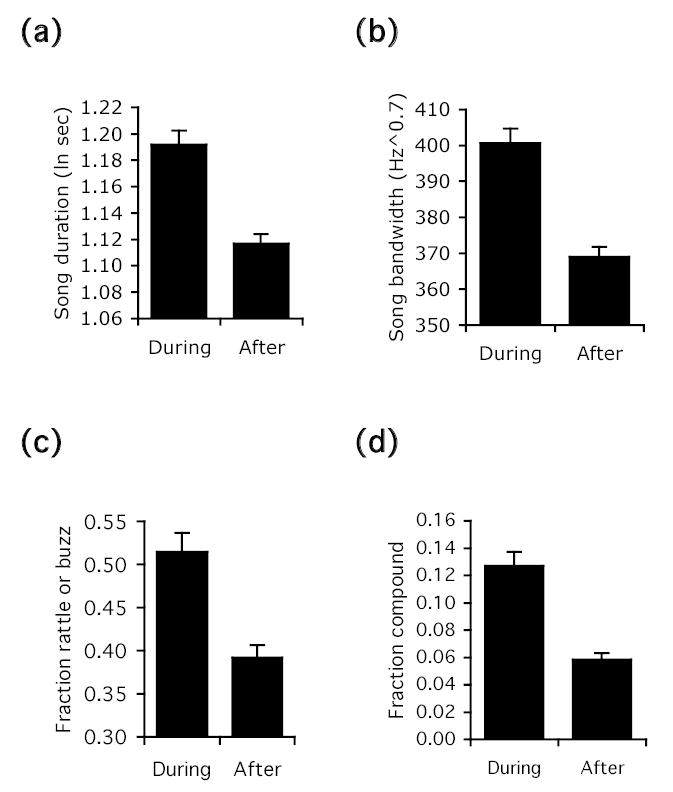
Significant main effects of time of day on a) song duration, b) song bandwidth, c) fraction of songs with rattle or buzz, and d) fraction of compound songs. Bars show least-squares means and standard errors (corrected for bird ID and other effects in the model) for the period during dawn chorus (before 0600) and after dawn chorus (after 0600).
Figure 4.
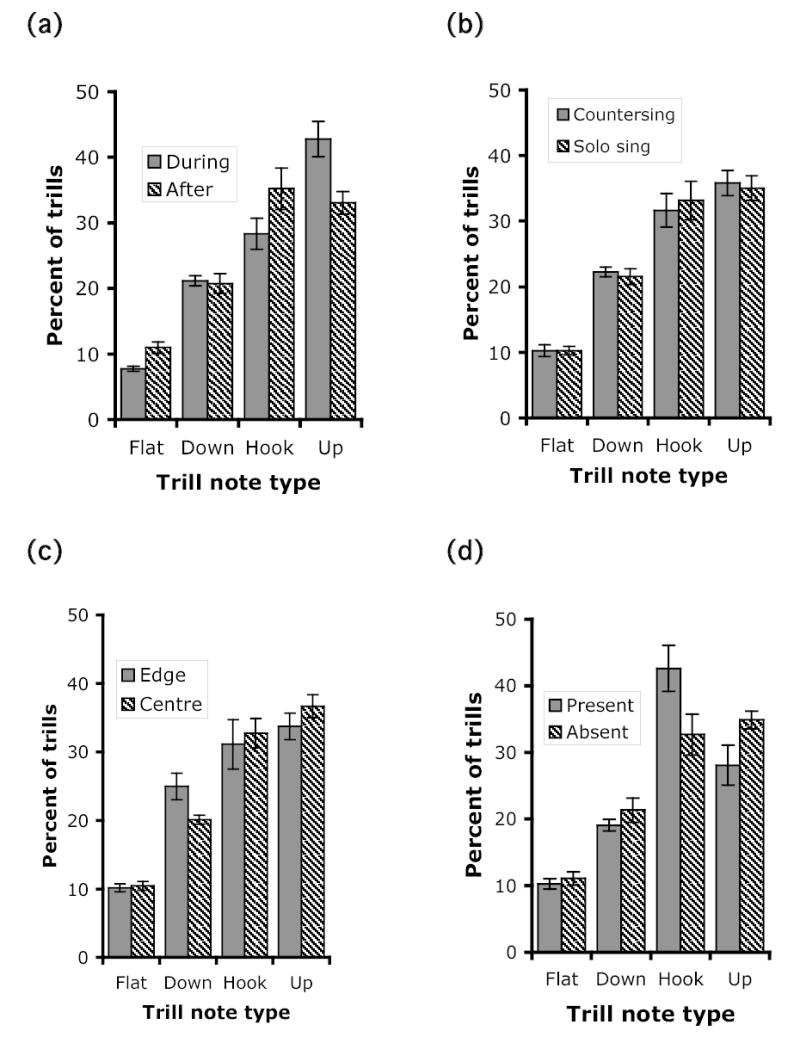
Main effects of each of the four contexts on the percentage of flat-, down-, hook- and up-note trills delivered. Bars show the mean ± SE of the percent of songs of each trill type for the six focal males. a) during versus after dawn chorus, b) countersinging versus solo singing, c) singing from the territory edge versus centre, and d) female present versus absent (after dawn chorus). Results of nominal logistic regression analysis of trill type presented in Table 5.
When birds were countersinging (as opposed to solo singing) and singing at the edge (as opposed to the centre) of their territories, they used song types with structural characteristics similar to those used during the dawn chorus, e.g., longer, broader bandwidth, complete, compound, and delivered with a higher switching rate. These two contexts also exhibited a few interaction effects in the same direction (Fig. 5). Rattle/buzz song types were preferentially used during countersinging primarily after dawn chorus, and birds countersinging at the edge of their territories used especially long and broadband song types. Trill characteristics were more important for the singing state and territory contexts. Trill duration was longer during both countersinging and singing at the territory edge. Up- and down-sweeping trills were more prevalent than hook- or flat-note trills. After dawn chorus, song types with broader bandwidth trills were preferentially used at the territory edge. The consistent association of acoustic features such as long and broadband songs and trills, presence of rattle/buzzes, and high switching rate with more aggressive contexts involving vocal interactions and closer encounters with neighbours suggests that these features convey a stronger threat message.
Figure 5.
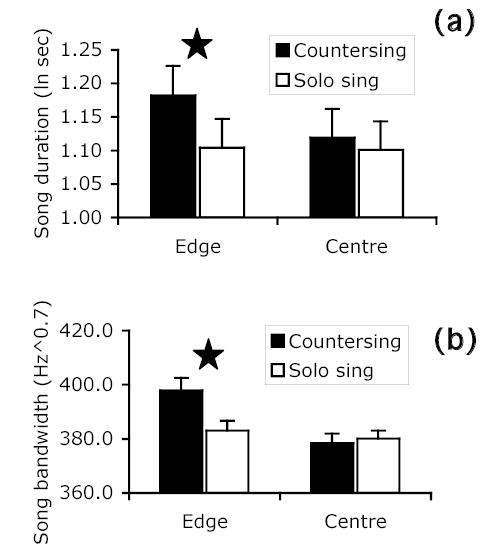
Interaction effects between singing state and location on territory (least-squares means ± SE) for a) song duration and b) song bandwidth. Star indicates significant post hoc contrast between least-squares means (P < 0.05).
Songs delivered in the presence of the female are nearly opposite in acoustic structure to those described above. Female-context songs have a narrower song and trill bandwidth and they are less likely to contain a rattle/buzz. Song rate is slower when the female is present. There was one complex three-way interaction effect involving females and the variable trill bandwidth, in which female presence caused males to narrow their bandwidth when singing at the territory centre after dawn chorus, and when singing at the territory edge during dawn chorus. Finally, males preferentially delivered compound songs and sang with higher song-type diversity in the presence of females (Table 5 and Fig. 6).
Figure 6.
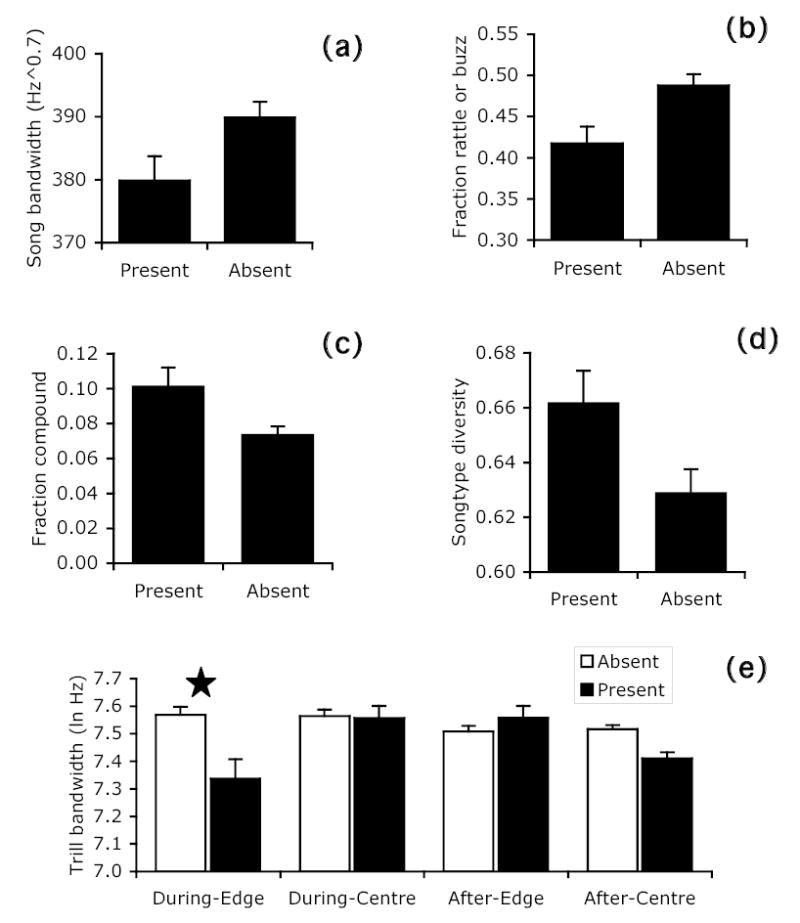
Significant effects of female presence on acoustic parameters in different contexts (least squares means ± SE) for a) song bandwidth, b) fraction rattle or buzz songs, c) fraction compound songs, d) song-type diversity, and e) trill bandwidth (star shows significant pairwise effect).
During all stages of breeding, banded wren males occasionally sing close to the nest. When we substituted distance to the nest instead of territory location into the analyses presented in Table 5, we found that songs close to the nest had similar characteristics to songs delivered in the centre (versus edge) of the territory, i.e., shorter, narrower in bandwidth, less likely to contain a rattle or buzz, and less likely to be complete and compound.
Seasonal effects
Although birds delivered nearly all of their song types on each of the observation days, simple contingency table analyses of song type versus day for each separate male were highly significant (P < 0.0001), indicating that the proportional use of song types differed on at least some days. One possible cause of daily differences was stage of breeding. Our dataset was not adequate for a thorough investigation of breeding stage effects, owing not only to the high nest predation rate that caused the birds to continually initiate new nests, but also to the small sample of birds and recording days per bird. Nevertheless, we attempted two sets of analyses, one that compared differences between the building stage (including nest construction and courtship behaviours) and the incubation stage for five birds, and another that compared building, incubation, and nestling stages for three birds. These repeated measures models contained the nesting stage variable and time of day as a continuous covariate. Song type diversity decreased from building to incubation (F1,4 = 13.6, P = 0.021) and then increased again during the nestling phase (F2,4 = 7.17, P = 0.048). Song types with longer trill duration were used during the incubation stage (F1,4 = 19.3, P < 0.012). Males did not show a reduction in song rate during incubation or nestling feeding, and some males even began constructing nests during these stages. The only conspicuous effect of breeding stage that we have noticed in our larger sample of early morning focal recordings of all study site males is a 1- to 2-day period just before females begin laying when males drop out of the dawn chorus and skulk in low bushes with the female. None of the samples in the current study included such a day.
Another potential source of daily differences in song-type use is differential interactions with neighbours. Territory boundaries sometimes shifted dramatically between recording days (Fig. 1 shows the first and last days’ boundaries for the focal males, along with the nest locations associated with the different territory positions). All birds changed their boundaries to some degree, with a mean change in territory area of 12.4 % (40% if we include male T’s move to a completely new territory). Three of the birds (O, W, and G) shifted their boundaries in response to the selection of a new nest site (by them or a neighbour) close to or beyond the territory boundary. The boundary shift resulted in the new nest being more centrally located in the territory, but impinged on a neighbouring bird’s territory. Physical fights and chases also occurred on most of these days. The other three birds also experienced more intense interactions and close boundary approaches with one of their neighbours on some days compared to other days. We asked whether any song variables differed between disputed versus stable boundary days using all six focal birds. Analytical models included the boundary stability variable and time of day as a continuous covariate. The only consistent difference was a higher song rate on disputed boundary days (F1,3 = 16.0, P = 0.028). There were non-significant trends for songs to be broader in bandwidth, compound, and to contain a rattle/buzz on dispute days. Birds clearly interacted and countersang more with the disputed boundary neighbour while reducing interactions with other neighbours on these days of heightened conflict.
Song-type Use with Different Neighbours
Given the high song-type sharing between adjacent neighbours (Table 1), and the use of type matching as an aggressive threat, we hypothesized that birds would use a different subset of song types when interacting with different neighbours, and in particular, that they would preferentially use the song types they shared with that neighbour (question 4). The frequency of use of a song type within each bird’s repertoire increased as a function of the number of neighbours sharing the song type (Random coefficient model (Littell et al. 1996): F1,5 = 10.48, P = 0.023; effect size Pearson r = 0.267, N = 135, birds pooled). We could only quantitatively examine neighbour-specific song-type use in four birds (males O, W, G, and J) that interacted extensively with neighbours whose repertories were well known. Bouts of exclusive interaction with each of the two neighbours, either via countersinging or solo singing at that neighbour’s boundary, were identified for these four males, and the relative use of each song type was tallied in a contingency table (P < 0.0001 for all four birds). We then looked at the subset of song types that were uniquely shared with just one of the two neighbours and determined whether or not the sharing neighbour received more than the expected number of occurrences of those song types. All four focal birds preferentially used most or all of the song types shared with a single neighbour more often than expected when interacting with that neighbour (male W: 9 of the 10 uniquely shared types; male G: 7 of 7 types; male J: 6 of 8 types; male O: 6 of 8 types) (Z = 4.20, P < 0.0001, test of observed estimate of 28/33 = 0.85 versus expected 0.5 with correction for over-dispersion among males (McCullagh & Nelder 1989, eq. 4.22)).
DISCUSSION
In this study we showed that male banded wrens differentially use the song types in their repertoires depending on the social context. The social contexts that most affected male singing behaviour and song-type choice were time of day (during versus after dawn chorus), location on the territory (edge, centre, and close to the nest), presence of the female, countersinging versus solo singing, and vocal interaction with specific neighbours. Stage of nesting had relatively little effect on song-type choices. Acoustic features of song types were also associated with these different contexts.
Song types delivered during dawn chorus were longer, wider in bandwidth, and more likely to contain a rattle or buzz and up-sweeping trill notes. Dawn songs were also presented at a higher song rate and with a higher song-type switching rate, and were more likely to be delivered complete and in compound form, compared to the average song delivered later in the morning. These results corroborated our initial impression of more vigorous-sounding songs given at this time of day. Dawn songs are loud and can be heard two territories away. The up-sweeping trills characteristic of many dawn songs are especially loud (casual sound level measurements estimated as 95 dB at 1m for the trill), which may facilitate long-distance communication. Several lines of evidence suggest that dawn singing is a competitive display among neighbouring males. 1) Males approach and sing toward each neighbour at some point during the chorus. 2) Immediate and slightly delayed type-matching of neighbours’ songs occurs commonly during the chorus. Matching rates are significantly higher than expected given the observed levels of song-type sharing among the males. Moreover, males alternately address different neighbours over short time periods with directed type matches, a behaviour that justifies the designation of dawn chorus in the banded wren as a true communication network (Burt & Vehrencamp 2005). 3) The intensity of interaction and type matching during dawn chorus is strongest between adjacent males undergoing a territory boundary dispute, and reflects the intensity of countersinging and aggressive encounters later in the morning (Burt & Vehrencamp 2005; unpublished data). We therefore interpret the greater duration, bandwidth, and complexity of dawn songs as costly, moderately aggressive male-directed acoustic signals that may provide information about male quality, health, and/or motivation. However, we can’t rule out the possibility that females eavesdrop on male interactions and use this information in making later mate-choice decisions, even though they are still on their nests during the dawn chorus.
The characteristics of song types and singing patterns given during countersinging interactions with neighbours after dawn chorus were very similar to the characteristics of dawn chorus songs, although the specific song type preferences are somewhat different. This similarity between dawn chorus and later morning countersinging song characteristics lends further support to the idea that dawn chorus singing is largely directed at rival males. A parallel argument has been made for wood warblers, which use their Type II song during both rival male interactions and dawn chorus (Staicer et al. 1996). In the banded wren, songs delivered during countersinging interactions as well as dawn chorus are significantly more likely to contain a rattle or buzz. These song elements are acoustically similar to the broadband ticking, grunting, and screaming calls the wrens give in alarm, intruder, and chasing contexts. Incorporation of these aggressive call elements into song types may give them a stronger threat salience. A similar argument was made for willow warblers, in which the aggression-associated song type contains initial notes that are similar in structure to call notes given in agonistic contexts (Järvi et al. 1980). Finally, the prevalence of broadband down-sweeping trills given at the territory edge is consistent with Morton’s (1977) motivational-structural rules for more aggressive sounds. Males spent a great deal of their time after dawn chorus solo singing from the central region of their territories and/or close to their nests. These songs were shorter, narrower in bandwidth and simpler in structure. Moreover, they were often delivered incompletely, at a slower rate, and in repeat mode. The song types used in this context rarely occurred during dawn chorus and intense countersinging (and when they did, were often part of a compound song). This manner of singing may function to advertise the presence and location of the male to the mate. Further evidence for the use of shorter song types to signal retreat or non-aggression was recently obtained in an interactive playback of overlapping versus alternating song, in which males selected shorter song types, produced fewer compound songs, and stayed further from the speaker during the overlapping treatment (Hall et al., submitted).
Females approached their mates after emerging from their sleeping or breeding nests in the morning, and when males were engaged in highly aggressive countersinging interactions at a territorial boundary. Mated males and females also interacted within close proximity to the nest. The presence of the female causes an immediate shift in the song type usage of the singing male, with types 118, 218, 06, 09, and 21 given more often than expected (Fig. 2). Acoustic characteristics of these song types included narrower whole-song bandwidth, narrower trill bandwith, hook-note trills, and lack of rattles and buzzes. These song types thus possess largely opposite features to the song types used in aggressive contexts, and could signal a level of appeasement. It is also possible that these song types are similar in structure to the songs females sing and allow the male and female to match each other. The appearance of the female between 0530 and 0540 hours in the morning clearly brings a halt to the male’s dawn chorus interactions with neighbouring males, and the male and female may sing together in an uncoordinated duet for 10–15 min. Female termination of dawn chorus singing has likewise been noted in several other species (Mace 1986; Slagsvold et al. 1994; Slagsvold 1996; Welling et al. 1997).
Singing in the presence of females is also associated with more compound songs and greater song-type diversity. These presentation characteristics are consistent with our finding of higher song-type diversity during the building/courtship phase of nesting. When the new male T took over the territory and mate of the prior male on the second day of recording, he was engaged in both boundary defence and courtship, and delivered many double and triple songs that resulted in high measures of song-type diversity. We have also noted bouts of singing in which the male approaches his fertile female and sings softly with especially high diversity. This manner of singing to females is consistent with the observed female preferences for males with complex songs, high song-type diversity, and large repertoire size in many species (Searcy 1996; MacDougal-Shackleton 1997).
Context-specific use of different trill note structures (broadband up-sweeping and down-sweeping trills in aggressive male contexts, narrowband hook-note trills in the female context) may have further significance from a production constraint point of view. Sound production mechanisms in the banded wren are probably very similar to those described for northern cardinals (Cardinalis cardinalis), a species with similar trilled songs (Suthers & Goller 1997). Broadband up-sweeping and down-sweeping trill notes require a seamless shift from one side of the syrinx to the other coupled with minibreaths between notes, a process involving extremely fine-tuned motor coordination (see song type 222, Fig. 2). Narrowband trills are probably produced from one side of the syrinx (still with minibreaths) and may be somewhat easier to produce. Minibreaths enable the bird to make longer trills before running out of air. Rattles and buzzes involve note repetition rates too rapid to permit minibreaths and are therefore produced with a single, pulsatile expiration. The duration of such elements is thus limited and may constrain the bird’s ability to continue singing a long song (Suthers & Goller 1997). The acoustic features of wren songs used during the dawn chorus and aggressive interactions are therefore more difficult and costly to produce and may be selected as honest indicators of a male’s age, experience, and/or stamina (Vehrencamp 2000; Gil & Gahr 2002).
The preferential, but by no means exclusive, use of different song types in different social contexts in the banded wren appears to be qualitatively similar to the differential song type use found in a variety of other species with medium-sized repertoires (see citations in the introduction). The association we found between acoustic structure and aggressive context is also similar to a pattern described in two wood warbler species (Spector 1991; Staicer 1996). Selection of song types based on their acoustic structure comes on top of song-type choice based on neighbour matching, as well as different short-term patterns of switching and song-type diversity. The banded wren may possess a moderately large repertoire because it uses such a large variety of singing strategies. With respect to male-male communication, a singer requires a set of shared and unshared song types for use with each neighbour, and a subset of song types within each of these categories that vary in duration, frequency structure, and note type. These options give males the ability to signal different degrees of aggressive intentions ranging from disinterest to sustained vocal sparing and from retreat to approach. With respect to female-directed singing, non-aggressive song types are required to coordinate breeding activities and joint territory defence, and a sufficiently large repertoire is needed to court females with high-diversity displays. This use of song in a large range of contexts may be partly responsible for the evolution of immediate-variety singing in this species.
Non-technical Summary
Song types and their structural features are associated with specific contexts in the banded wren. 2005. Trillo, P.A. and Vehrencamp, S. L.
Earlier work on songbird communication suggested that the primary function of different song types in a repertoire was to increase stimulus diversity, reduce habituation of the receiver, and/or relieve stress on syringeal muscles. However, studies on a growing number of species indicate that song types are used selectively in different contexts and therefore probably convey different messages. This study is a unique attempt to examine song-type use in a tropical resident songbird, the banded wren Thryothorus pleurostictus, with a moderately large repertoire of around 20 song types per male. We investigated: (1) the importance of context in male choice of song types, and (2) whether these choices were related to song acoustic features. Males do use song types differentially depending on the context. In addition, songs delivered during dawn chorus are on average longer, of greater pitch range, and more likely to contain a rattle or buzz, compared to songs delivered later. Similar song features are also observed when males engage in territorial boundary disputes. Different song types and acoustic features are associated with presence of the mate. The acoustic structure of a song type is therefore an important component of its message.
Acknowledgments
Logistical support for our long-term study has been provided by the staff of the Area Conservación Guanacaste, in particular Director of Research Roger Blanco. Laura Molles and Frank Joyce provided invaluable background information on the Banded Wrens in Santa Rosa. We also thank John Burt, Peter Ajtai and Sebastian Jurado for assistance with the field work. Jeff Lucas and Francoise Vermeylen provided invaluable statistical advice. Several anonymous reviewers provided comments that greatly improved the manuscript. This study was supported by NIMH grant R01-MH60461.
References
- Baptista LF. Territorial, courtship, and duet songs of the Cuban grassquit (Tiaris canora) Journal für Ornithologie. 1978;119:91–101. [Google Scholar]
- Burt, J. M. & Vehrencamp, S. L. 2005. Dawn chorus as an interactive communication network. In: Animal Communication Networks (Ed. by P.K. McGregor), pp. 320–343. Cambridge: Cambridge University Press.
- Byers BE. Messages encoded in the songs of chestnut-sided warblers. Animal Behaviour. 1996;52:691–701. [Google Scholar]
- Catchpole CK. Variation in the song of the Great Reed Warbler Acrocephalus arundinaceus in relation to mate attraction and territorial defence. Animal Behaviour. 1983;31:1217–1225. [Google Scholar]
- Catchpole, C.K. & Slater, P.J.B. 1995. Bird Song: Biological Themes and Variations. Cambridge: Cambridge University Press.
- Cozens SE, Falls JB. Structure and use of song in the Yellow-headed Blackbird (Xanthocephalus zanthocephalus) Zeitschrift für Tierpsychologie. 1984;66:227–241. [Google Scholar]
- Dabelsteen, T. 1992. Interactive playback: a finely tuned response. In: Playback and Studies of Animal Communication (Ed. by McGregor, P. K.), pp. 97–110. New York: Plenum Press.
- Derrickson KC. Behavioral correlates of song types of the northern mockingbird (Mimus polyglottos) Ethology. 1987;74:21–32. [Google Scholar]
- Duguay JP, Ritchison G. A contextual analysis of singing behavior in male tufted titmice. Journal of Field Ornithology. 1999;69:85–94. [Google Scholar]
- Gaddis PK. Differential usage of song types by Plain, Bridled, and Tufted Titmice. Ornis Scandinavica. 1986;14:16–23. [Google Scholar]
- Gil D, Gahr M. The honesty of bird song: multiple constraints for multiple traits. Trends in Ecology & Evolution, 2002;17:133–141. [Google Scholar]
- Hartshorne, C. 1973. Born to sing Bloomington, IN: Indiana University Press.
- Järvi T, Radesater T, Jacobsson S. The song of the Willow Warbler Phylloscopus trochilus with special reference to singing behaviour in agonistic situations. Ornis Scandinavica. 1980;11:236–242. [Google Scholar]
- Johnson LS. Pattern of song type use for territorial defence in the Plain Titmouse Parus inornatus. Ornis Scandinavica. 1987;18:24–32. [Google Scholar]
- Krebs JR, Ashcroft R, Webber M. Song repertoires and territory defence in the great tit. Nature. 1978;271:539–542. [Google Scholar]
- Lambrechts MM, Dhondt AA. The anti-exhaustion hypothesis: a new hypothesis to explain song performance and song switching in the great tit. Animal Behaviour. 1988;36:327–334. [Google Scholar]
- Lein MR. Song variation in a populations of chestnut-sided warblers (Dendroica pensylvanica): its nature and suggested significance. Canadian Journal of Zoology. 1978;56:1226–1283. [Google Scholar]
- Littell, R. C., Milliken, G. A., Stroup, W. W. & Wolfinger, R. D. 1996. SAS System for Mixed Models Cary, N.C.: SAS Institute, Inc.
- Luschi P, del Seppia C. Song-type function during territorial encounters in male Cetti’s Warblers Cettia cetti. Ibis. 1996;138:479–484. [Google Scholar]
- MacDougall-Shackleton SA. Sexual selection and the evolution of song repertoires. Current Ornithology. 1997;14:81–124. [Google Scholar]
- Mace RH. The importance of female behaviour in the dawn chorus. Animal Behaviour. 1986;34:621–622. [Google Scholar]
- McCullagh, P. & Nelder, J. A. 1989. Generalized Linear Models, 2nd ed. Boca Raton, F.L.: Chapman & Hall/CRC.
- Molles, L. E. 1999. Use of song-type repertoires by banded wrens. La Jolla: University of California, San Diego.
- Molles LE, Vehrencamp SL. Repertoire size, repertoire overlap, and singing modes in the banded wren, Thryothorus pleurostictus. Auk. 1999;116:677–689. [Google Scholar]
- Molles LE, Vehrencamp SL. Songbird cheaters pay a retaliation cost: evidence for auditory conventional signals. Proceedings of the Royal Society of London, Series B. 2001;268:2013–2019. doi: 10.1098/rspb.2001.1757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morrison ML, Hardy JW. Vocalizations of the Black-throated Gray Warbler. Wilson Bulletin. 1983;95:640–463. [Google Scholar]
- Morton ES. On the occurrence and significance of motivation-structural rules in some bird and mammal sounds. American Naturalist. 1977;111:855–869. [Google Scholar]
- Nelson DA, Croner LJ. Song categories and their functions in the field sparrow (Spizella pusilla) Auk. 1991;108:42–52. [Google Scholar]
- Nottebohm F. The origins of vocal learning. American Naturalist. 1972;106:116–140. [Google Scholar]
- Podos J. A performance constraint on the evolution of trilled vocalizations in a songbird family (Passeriformes: Emberizidae) Evolution. 1997;51:537–551. doi: 10.1111/j.1558-5646.1997.tb02441.x. [DOI] [PubMed] [Google Scholar]
- Schroeder DJ, Wiley RH. Communication with repertoires of shared song themes in Tufted Titmice. Animal Behaviour. 1983;31:1128–1138. [Google Scholar]
- Searcy WA. Song repertoire and mate choice in birds. American Zoologist. 1992;32:71–80. [Google Scholar]
- Searcy WA, Searcy MH, Marler P. The response of swamp sparrows to acoustically distinct song types. Behaviour. 1982;80:70–83. [Google Scholar]
- Slagsvold T. Dawn and dusk singing of male American Robins in relation to female behavior. Wilson Bulletin. 1996;108:507–515. [Google Scholar]
- Slagsvold T, Dale S, Saetre GP. Dawn singing in the great tit (Parus major): mate attracting, mate guarding, or territorial defence? Behaviour. 1994;131:115–138. [Google Scholar]
- Smith WJ. Message, meaning, and context in ethology. American Naturalist. 1965;99:703–716. [Google Scholar]
- Smith WD, Pawlukiewicz J, Smith ST. Kinds of activities correlated with singing patterns of the yellow-throated vireo. Animal Behaviour. 1978;26:862–884. [Google Scholar]
- Spector DA. The singing behaviour of yellow warblers. Behaviour. 1991;117:36–51. [Google Scholar]
- Spector, D. A. 1992. Wood-warbler song systems: A review of Paruline singing behaviors. In: Current Ornithology (Ed. by Power, D. M.), pp. 199–238. New York: Plenum Press.
- Staicer CA. Acoustic features of song categories of the Adelaide’s Warbler (Dendroica adelaidae) Auk. 1996;113:771–783. [Google Scholar]
- Staicer, C.A., Spector, D.A. & Horn, A.G. 1996. The dawn chorus and other diel patterns of acoustic signalling. In Ecology and Evolution of Acoustic Communication in Birds (Ed. by
- Suthers RA, Goller F. Motor correlates of vocal diversity in songbirds. Current Ornithology. 1997;14:235–288. [Google Scholar]
- Vehrencamp, S. L. 2000. Handicap, index, and conventional signal elements of bird song. In: Animal Signals: Signalling and Signal Design in Animal Communication (Ed. by Espmark, Y., Amundsen, T. & Rosenqvist, G.), pp. 277–300. Trondheim: Tapir Publishers.
- Vehrencamp SL. Is song-type matching a conventional signal of aggressive intentions? Proceedings of the Royal Society of London, Series B. 2001;268:1637–1642. doi: 10.1098/rspb.2001.1714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Welling P, Koivula K, Orell M. Dawn chorus and female behaviour in the Willow Tit Parus montanus. Ibis. 1997;139:1–3. [Google Scholar]
- Wiebe MO, Lein MR. Use of song types by Mountain Chickadees (Poecile gambeli) Wilson Bulletin. 1999;111:368–375. [Google Scholar]