

Abstract
Study design
Spinal reflex excitability study in sensory–motor incomplete spinal cord-injured (SCI) and spinal intact subjects.
Objectives
To investigate the effects of plantar cutaneous afferent excitation on the soleus H-reflex and flexion reflex in both subject groups while seated.
Setting
Rehabilitation Institute of Chicago and City University of New York, USA.
Methods
The flexion reflex in SCI subjects was elicited by non-nociceptive stimulation of the sural nerve. In normal subjects, it was also elicited via innocuous medial arch foot stimulation. In both cases, reflex responses were recorded from the ipsilateral tibialis anterior muscle. Soleus H-reflexes were elicited and recorded via conventional methods. Both reflexes were conditioned by plantar cutaneous afferent stimulation at conditioning test intervals ranging from 3 to 90 ms.
Results
Excitation of plantar cutaneous afferents resulted in facilitation of the soleus H-reflex and late flexion reflex in SCI subjects. In normal subjects, the soleus H-reflex was depressed while the late flexion reflex was absent. The early flexion reflex was irregularly observed in SCI patients, while in normal subjects a bimodal reflex modulation pattern was observed.
Conclusion
The effects of plantar cutaneous afferents change following a lesion to the spinal cord leading to exaggerated activity in both flexors and extensors. This suggests impaired modulation of the spinal inhibitory mechanisms involved in the reflex modulation. Our findings should be considered in programs aimed to restore sensorimotor function and promote recovery in these patients.
Keywords: foot sole, flexion reflex, soleus H-reflex, rehabilitation, sensorimotor integration, SCI
Introduction
Excitation of low-threshold mechanoreceptors or skin afferents induces the classical flexion reflex which involves mutual excitation of flexors and inhibition of extensors, but also an extensor thrust activating mainly the toe extensors.1–2 This suggests that specialized cutaneous pathways act also via ‘private’ neuronal pathways not belonging to the flexion reflex afferent (FRA) system, converging onto common interneurones in the reflex pathways to α-motoneurones,3 with their main functional role to stabilize the foot during the stance phase of the moving limb.4
Experiments in spinalized and decerebrated animals have shown that low-threshold afferents of the foot sole delay or suppress the initiation of swing, inhibit late flexion reflexes, promote stance, and contribute to the correct placement of the foot during walking.5–8 These studies signify that plantar cutaneous afferents interact with neuronal circuits involved in stepping, while their action has been partly associated with the functional recovery of walking in spinalized animals by modifying skin input signalling ground contact.9
In healthy humans, plantar cutaneous afferents modify the swing phase of gait by changing the amplitude of the cutaneous reflex responses,10 while in cases where such information is disrupted, as in patients with sensory polyneuropathy, a large variability in the step-cycle duration has been reported.11 Cutaneous afferents have also segmental short latency reflex effects, as postulated by shifts on the actions of group Ib and presynaptic inhibitory interneurones in man,12,13 verifying further their contribution to the segmental control of movement.
The flexion reflex is known to be under descending control,14,15 and to be modulated by changes in limb posture, site of stimulation, intensity level, electrically induced muscle contraction, excitation of distal cutaneous afferents, and imposed static hip angle changes.16–20 Furthermore, the flexion reflex in human spinal cord injury (SCI) presents a similar neuronal organization with the flexion reflex observed in l-Dopa-treated spinal animals,21–23 supporting the hypothesis that FRA pathways are involved in the spinal stepping generator in humans.
Considerable evidence suggest that plantar cutaneous afferents affect FRA pathways and interact with spinal inhibitory control systems to modify motor output. Accordingly, the present study was undertaken to investigate the effects of skin afferents and especially those arising from the foot sole on flexion and extensor (soleus H-reflex) reflexes in cases where the supraspinal control is either intact or partially disrupted.
Methods
Subjects
Experiments were conducted with local ethics committee approval and the written consent of all subjects prior to testing. Nine spinal-intact (age 22–42 years) and eight subjects with clinically chronic motor and sensory incomplete SCI (American Spinal Cord injury Association (ASIA)) classification C24 ranging from cervical 5 to thoracic 11 spinal segments participated in this study (Table 1). Patients participating in the current tests are the same who participated in previous studies,20,26 and are identified with the same numbers. All tests were conducted with subjects’ seated (hip angle, 120°; knee angle, 160°; ankle angle, 110°) while both feet were supported by footrests.
Table 1.
SCI subjects’ characteristicsa
| Subjects | Gender | Age (years) | Post-injury (months) | Ashworth score | ASIA scale | Lesion level | Sensation | Medication/day |
|---|---|---|---|---|---|---|---|---|
| S1 | M | 23 | 78 | 1 | C | C5 | Intact | – |
| S3 | M | 27 | 26 | 0 | C | C5 | Intact | – |
| S4 | M | 45 | 54 | 2 | C | C6 | Intact | Baclofen 60 mg |
| S5 | M | 20 | 21 | 2 | C | T11 | Intact | Baclofen 20 mg |
| S7 | F | 59 | 26 | 0 | C | T9 | Intact | Baclofen 10 mg |
| S9 | M | 42 | 38 | 0 | C | C8 | Intact | Plavix lipotor 75 mg |
| S10 | M | 50 | 186 | 1 | C | C5 | No sensation of cold/warm | Baclofen 30 mg |
| S11 | M | 50 | 162 | 3 | C | C5 | No sensation of cold/warm | – |
American spinal injury association (ASIA)24 scale C represents sensory and motor incomplete SCI with more than half of muscles below the injury level having a muscle grade of less than 3 out of 5. Spasticity at the ankle was scaled according to the Ashworth score.25 F, female; M, male; C, cervical; T, thoracic
Elicitation and recording protocol of the soleus H-reflex
The H-reflex was evoked by monopolar electrical stimulation of the right posterior tibial nerve at the popliteal fossa using a constant current stimulator (Digitimer Ltd, UK) that was externally triggered by a data acquisition system (1401 Plus running Spike 2 or by an A/D converter running customized LabView instruments). Reflex responses were recorded by a single differential bipolar electrode (Bagnoli 8, DelSys Inc., MA, USA) placed over the soleus muscle, which was secured via adhesive tape. The indifferent electrode was a circular stainless-steel plate (≈3 cm diameter) placed just above the patella.
For each subject at the start of the test, a hand-held monopolar stainless-steel electrode was used as a probe to establish the correct site for stimulating the posterior tibial nerve. This was identified as the one during which the soleus H-reflex was present without the M-wave being present. Having established this site, the probe electrode was replaced by a permanent one (N-10-A, Ambu Inc., Denmark) under constant pressure, and the evoked responses were observed online on the oscilloscope screen. In case the surface electrode did not evoke the same response behavior, this procedure was repeated. After this procedure was completed, the maximal M-wave was evoked and measured as peak to peak amplitude of the unrectified waveform.
The susceptibility of the reflex to inhibition and facilitation depends on the size of the test reflex.27 Accordingly, the stimulus strength was adjusted to give a control H-reflex of 15–30% of the maximal M-wave across subjects tested. This allowed a valid comparison to be made on the amount of facilitation/inhibition brought by excitation of plantar cutaneous afferents on the soleus H-reflex. In addition, the M-wave amplitude was continually monitored during the course of an experiment to ensure stability in the stimulation and recording procedures, while it constituted a within subject factor for accepting the conditioned H-reflexes. In both subject groups, for each H-reflex (control or conditioned) recorded in this study, 20 responses evoked every 5 s were acquired and saved for further analysis.
Elicitation and recording protocol of the flexion reflex
The flexion reflex in SCI subjects was evoked by two disposable pregelled Ag–AgCl electrodes (Ambu Inc., Denmark) positioned across the receptive field of the sural nerve at the right lateral submalleolar region. The sural nerve was stimulated by a 30 ms train of 1-ms pulses delivered at 300 Hz once every 10 s using a constant current stimulator (DS7A, Digitimer, UK). Reflex responses were recorded by a single differential electrode (DE-2.1; DelSys, Boston, MA, USA) placed over the tibialis anterior (TA) muscle following light mechanical abrasion of the skin. At this interstimulus interval, no evidence of habituation in the flexion reflex was encountered.28,29 In every trial (control or conditioned), 20 TA flexion reflexes were evoked and saved for further analysis.
At the beginning of the experiment and for each subject, the stimulus delivered to the sural nerve was set at 0 mA and was increased slowly so as to observe the evoked response on the oscilloscope screen. The stimulus intensity during which the initial EMG TA activity was induced was identified as the reflex threshold (RT). This reflex response was categorized as early if its latency was less than 100 ms, and as late when its latency was beyond 120 ms.20,30 During testing, the sural nerve was stimulated at 1.5 times RT. At this intensity, no limb movement was evoked and subjects reported no pain following sural nerve stimulation, indicating that the test afferent volley included large cutaneous afferents eliciting the non-nociceptive flexion reflex.31 The flexion reflex in spinal intact subjects was elicited by bipolar stimulation of the medial arch and sural nerve of the right foot following identical procedures employed in SCI subjects.
For both reflexes (flexion and H-reflex), the recorded EMG signals were amplified 1000 times and band pass filtered (10 Hz–1 kHz), and subjected to an analog to digital conversion (PCI-MIO 16E, National Instruments Co., Austin, TX, USA; 1401 plus running Spike 2, CED Ltd, Hertfordshire, UK).
Plantar cutaneous afferent conditioning stimulation
Plantar cutaneous afferents stimulation was identical to that previously employed in both normal and SCI subjects.20,32 They were excited via two Ag–AgCl surface electrodes (Ambu Inc., Ølstykke, Denmark) placed transversely across the first and third metatarsals. The conditioning stimulus train consisted of five pulses with an interstimulus interval of 4.8 ms and pulse train duration of 24 ms. The conditioning pulse train preceded the test reflexes (soleus H-reflex and/or TA flexion reflex) at different conditioning test (C-T) intervals, measured as the time between the end of the pulse train delivered to the foot sole and the beginning of the pulse train or single pulse delivered to the sural nerve/medial arch or to the posterior tibial nerve, respectively. The C-T intervals were delivered randomly, and ranged from 3 to 90 ms (3, 6, 9, 15, 30, 60, and 90 ms) for both subject groups, to observe short and long latency effects on both reflexes.
In all subjects, initially the perceptual threshold (PT) that corresponded to the stimulus intensity first perceived by the subject was established. All conditioning stimuli were equivalent to three times PT. At this stimulation intensity no movement of the intrinsic muscles of the foot was elicited and no pain was reported, verifying that the conditioning afferent volley included mainly large cutaneous afferents of the foot sole. In both subject groups, the conditioning effects of plantar cutaneous afferent excitation on the soleus H-reflex and flexion reflex were examined in a different session. Each session lasted no more than 2 h, while at each test at least nine different reflex sequences were recorded.
Data analysis
The digitized EMG signals were full-wave rectified and the size of the flexion reflex (early and/or late) was quantified by calculating the area under the EMG records corresponding to the period between the onset latency and the point in time at which the response fell below 1.5 standard deviation of the baseline EMG activity. The soleus H-reflex and M-wave were also measured as the area under the curve of the full-wave rectified waveform.
For each subject, the 20 conditioned reflex responses (H-reflex, early, and/or late flexion reflex) recorded at each C-T interval were expressed as a percentage of the mean size of the associated control reflex. Then, a one-way analysis of variance (ANOVA) along with post hoc Bonferroni tests was applied to the data to establish significant differences between the control and the conditioned reflexes. This analysis was performed for each subject separately. The mean size of the conditioned reflex from each subject was then grouped based on the C-T interval investigated. A one-way ANOVA with repeated measures was applied to the data so to establish changes in the magnitude of the conditioned reflexes across C-T intervals investigated. Conditioned soleus H-reflexes were only accepted when there was no statistically significant difference between the M-waves recorded under control conditions and following conditioning stimulation. Statistically significant difference was established at 95% of confidence level. The results are presented as mean values and standard error of the mean (SEM).
Results
Effects of plantar cutaneous afferent excitation on the H-reflex in spinal intact and injured subjects
In Figure 1a, the effects on the average (n = 20) soleus H-reflex recorded in one normal and in one SCI subject (S11) following excitation of the ipsilateral plantar cutaneous afferents at 15 and at 60 ms C-T intervals are illustrated. Note that the reflex depression in the normal subject and the reflex facilitation in the SCI subject following excitation of plantar cutaneous afferents were observed with no significant changes in the amplitude of the M-waves signifying stable stimulation and recording conditions.
Figure 1.
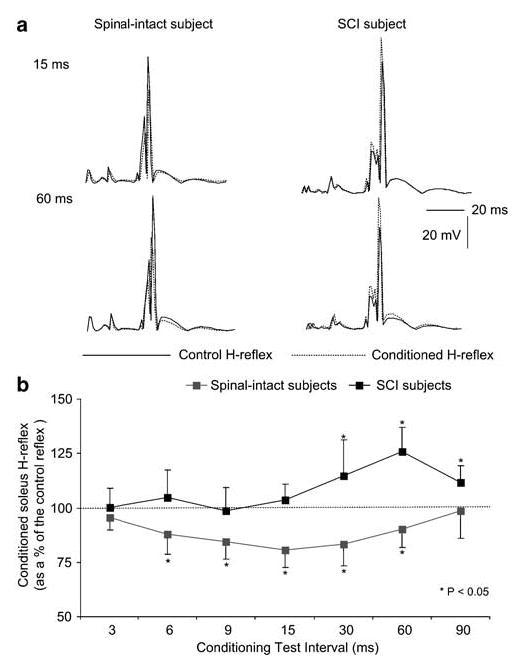
Effects of plantar cutaneous afferent excitation on the soleus H-reflex in normal and SCI subjects. (a) The average soleus H-reflex (n = 20) under control conditions (solid lines) and during conditioning of the reflex (dashed lines) at 15 and 60 ms of conditioning test (C-T) intervals for one normal and one SCI subject is indicated. (b) Pool data showing the effects of plantar cutaneous afferents excitation on the soleus H-reflex in normal and SCI subjects. For each C-T interval tested, the overall average (mean±SEM) size of the conditioned H-reflexes for both subject groups is indicated. Asterisks indicate cases of statistically significant differences between the control and the conditioned reflex sizes (P < 0.05)
In SCI subjects, excitation of plantar cutaneous afferents for C-T intervals ranging from 3 to 15 ms induced no significant effects on the soleus H-reflex (Figure 1b). Conditioning stimulation delivered at longer C-T intervals (30, 60, and 90 ms) resulted, however, in a significant facilitation of the soleus H-reflex (Figure 1b). The amplitude of the conditioned reflex did not differ across the long C-T intervals (30–90 ms) investigated (P = 0.55). In contrast, in normal subjects, excitation of plantar cutaneous afferents induced no significant effects on the soleus H-reflex when plantar sole stimulation was delivered at either 3 or 90 ms C-T intervals (P > 0.05). During the intermediate C-T intervals tested (6–60 ms), the soleus H-reflex was significantly depressed compared to control reflex values (P < 0.05) (Figure 1b).
Effects of plantar cutaneous afferent excitation on the flexion reflex in spinal intact and injured subjects
In SCI subjects, sural nerve stimulation at non-nociceptive levels induced an early flexion reflex (<100 ms latency) in two SCI subjects (S1, S4) only during conditioning of the reflex, and in one subject (S3) under control conditions and following excitation of plantar cutaneous afferents. Given the lack of the reflex appearance under control conditions in subjects 1 and 4, no conditioning effects could be established. In subject 3, the early flexion reflex was facilitated across all C-T intervals tested (3–90 ms) reaching amplitudes as high as 300% of control reflex values (data not shown graphically).
In Figure 2a, the effects of plantar cutaneous afferent excitation at three times PT on the average (n = 20) flexion reflex for two SCI subjects is illustrated. In the EMG recordings of subject 1 (top traces of Figure 2a), it can be seen that the conditioning stimulus resulted in inducing a response at latency less than 100 ms, while the magnitude of the TA responses at longer latencies was significantly increased. The latter was also observed in subject 9, while the early reflex component was absent.
Figure 2.
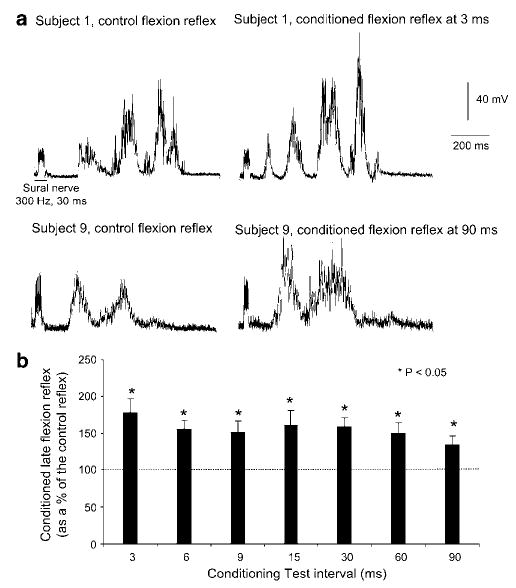
Effects of plantar cutaneous afferent excitation on the TA flexion reflex in SCI subjects. (a) The average (n = 20) flexion reflex recorded under control conditions and following conditioning stimulation delivered at 3 and at 90 ms conditioning intervals for two subjects (S1 and S9) is presented. See text for further description. (b) Pool data (all SCI subjects tested) showing the effects of plantar cutaneous afferent excitation on the late flexion reflex. For each conditioning test interval, the average size of the conditioned late flexion reflexes (as a percentage of the control late flexion reflex) was calculated for all subjects tested. Asterisks indicate cases of statistically significant differences between the control and the conditioned reflex sizes (P < 0.05). Error bars indicate the SEM
The late flexion reflexes (>120 ms) were consistently observed under all conditions across all SCI subjects. Excitation of tactile afferents of the foot sole resulted in a significant facilitation of the late flexion reflex across all subjects (Figure 2b). The late flexion reflex reached an overall amplitude of 180±18 and 135±11% of the associated control reflex values at 3 and at 90 ms, respectively. The amplitude of the conditioned reflex and thus the amount of reflex facilitation did not vary with changes in the C-T interval tested (P = 0.9).
In normal subjects, sural nerve stimulation at non-nociceptive levels induced no early or late flexion reflex. Further, even at higher intensities inducing pain, only in one subject a late flexion reflex response was observed. In this case, this reflex component was present during control conditions but following conditioning stimulation it was only observed at 3 and 6 ms of C-T intervals, being not significantly different from control reflex values (P > 0.05). At the remaining C-T intervals tested, the flexion reflex was not present. Based on our observations, it was apparent that sural nerve stimulation at non-nociceptive or noxious levels did not elicit a flexion reflex (either early or late) in normal subjects.
Non-nociceptive medial arch stimulation of the foot induced irregularly the late flexion reflex across normal subjects, being present in three out of nine. In two subjects, it was observed only under control conditions and it was absent during excitation of plantar cutaneous afferents. In the third subject, the late flexion reflex was depressed equally across C-T intervals, while the early flexion reflex was concurrently facilitated. Data from this subject was not included in the pool data analysis of the early flexion reflex given the different type of flexion reflex expression compared to the rest subjects.
The early flexion reflex (<100 ms latency) was observed uniformly across all normal subjects following non-nociceptive stimulation of the medial arch of the foot. Plantar cutaneous afferent excitation resulted in significant effects on the magnitude of this reflex component. These effects can easily be recognized in Figure 3, where the average full-wave rectified integrated EMG activity of the TA is illustrated.
Figure 3.
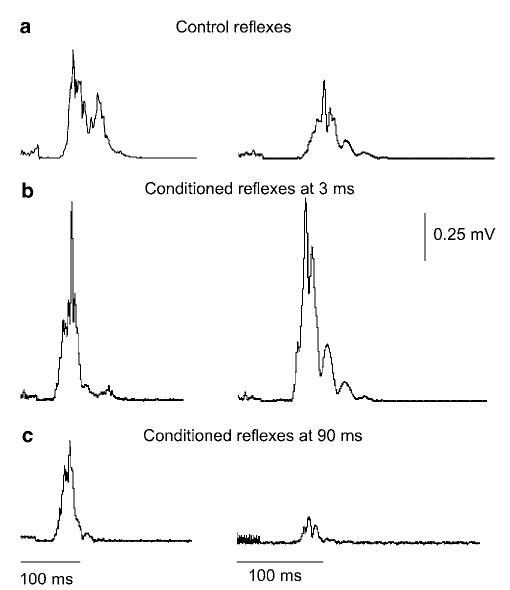
Full-wave rectified early flexion reflex elicited via medial arch stimulation in two normal subjects under control conditions (a) and following excitation of plantar cutaneous afferents at conditioning test (C-T) interval of 3 and 90 ms (b, c). Each waveform constitutes the average of 20 reflex responses elicited every 10 s. For both subjects, the early flexion reflex was facilitated at the C-T interval of 3 ms, while at the longest C-T interval tested (90 ms) the reflex was significantly depressed compared to control reflex values
Excitation of plantar cutaneous afferents induced a significant facilitation of the early flexion reflex at 3 ms (see Figures 3b and 4), while at C-T intervals of 6 and 9 ms the reflex reached its control values. At longer C-T intervals (15–90 ms), the reflex was significantly depressed, reaching overall amplitude of 50±7.6% of control reflex values at the C-T interval of 90 ms (P < 0.05) (Figures 3c and 4). The amplitude of the conditioned early flexion reflexes varied significantly across C-T intervals tested (P < 0.05).
Figure 4.
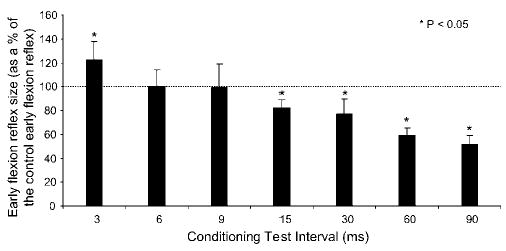
Histogram showing the effects of plantar cutaneous afferents excitation at three times perceptual threshold on the early TA flexion reflex elicited by medial arch stimulation in normal subjects. The average (all subjects tested) amplitude (mean±SEM) of the conditioned reflex is plotted for each conditioning test interval tested. Asterisks identify the conditioning test intervals in which the reflex modulation was significant at 95% confidence level
Discussion
Our current findings strongly support the notion that signals from foot mechanoreceptors participate in spinal interneuronal reflex circuits that have been coupled with human motor control and locomotion. These afferents appear to adjust motoneuronal excitability by their alternative access to reflex pathways involving flexors or extensors. The expression of this sensory afferent feedback changes following a lesion to the spinal cord, enhancing both soleus H-reflex and flexion reflex. Lastly, the flexion reflex in human SCI appears to be different in nature compared to the one observed in spinal intact subjects, probably due to involvement of different neuronal pathways during their expression.
Observations on the expression of the flexion reflex in subjects with intact or injured spinal cord
Non-nociceptive sural nerve stimulation induced a late flexion reflex in all SCI subjects, while the early flexion reflex was irregularly present. This is in line with our previous observations on the incidence of the flexion reflex components in human SCI.19,20 In contrast, non-nociceptive sural nerve stimulation induced no TA reflex responses in any of the spinal intact subjects tested, and even at nociceptive stimulus intensities the late flexion reflex was absent. A flexion reflex in normal subjects was only seen following non-nociceptive stimulation of the medial arch, but it appeared at latencies less than 100 ms.
The early flexion in SCI patients has been associated with the early reflex component observed in spinal intact human subjects and cats.33,34 It has recently been demonstrated that the flexion reflex with a latency of 80 ms (also known as P2 or medium latency response)35 is absent in motor complete SCI patients during assisted walking,36 probably because of a transcortical involvement during its transmission.37 This is in line with our findings, during which the early flexion reflex was observed in normal subjects following non-nociceptive medial arch stimulation and was occasionally observed in SCI subjects. At this point, we should consider the possibility that the absence of the late and early flexion reflex in normal and SCI subjects, respectively, might be related to the methods employed in this study, for example, no tonic preinnervation. Reflexes were tested without a maintained weak muscle contraction, given that the muscle contraction and relaxation phases are problematic in several aspects in human SCI.38,39 Further, the flexion reflex is known to depend not only on the characteristics of the sensory input but also on the state of spinal interneuronal circuitry40 which cannot be controlled through preinnervation.
Following l-Dopa administration in spinal cats, FRA excitation at non-nociceptive levels induces long-lasting synchronized discharges of flexors, while short latency flexion reflexes are depressed.23 The late flexion reflex in SCI man has shown similar neuronal characteristics to that observed in spinal cats,21,22 which is believed to utilize interneuronal circuits involved in spinal stepping and locomotion.41 Based on our findings, it is apparent that the flexion reflex responses in normal and SCI subjects have a different reflex organization mediated through different neuronal pathways.15
Modulation of spinal reflexes following excitation of plantar cutaneous afferents in subjects with intact or injured spinal cord
Excitation of plantar cutaneous afferents influenced significantly FRA pathways. In SCI subjects, a significant facilitation of the late flexion reflex for C-T intervals ranging from 3 to 90 ms was observed (Figure 2b). In neurologically intact subjects, the early flexion reflex elicited by non-nociceptive stimulation of the medial arch of the foot resulted in early facilitation followed by a significant depression at the longer C-T intervals tested (Figures 3 and 4). Sural nerve, medial arch, and plantar cutaneous afferents were all stimulated at low intensities exciting mainly low-threshold muscle and cutaneous afferents. In addition, all stimulation sites had equivalent distances to spinal segmental levels associated with the flexion reflex. Therefore, we can assume that the conduction time of the testing and conditioning afferent volleys was similar. Given the long latency of both flexion reflex components (>60 ms) and the long duration of the C-T intervals tested, it is likely that the reflex modulation was mediated through long latency spinal reflex circuits.
Identical cutaneous input modulated differently the flexion reflex in normal and SCI subjects. Nonetheless, the late flexion reflex in motor complete SCI patients evoked following non-nociceptive sural nerve stimulation is inhibited when cutaneous afferents of a major knee extensor muscle are activated,19 suggesting that cutaneous inputs affect FRA pathways differently depending on their location.
Many studies have demonstrated that cutaneous afferents can influence flexion reflex expression and motor activity during human walking. In particular, sural nerve reflex responses are reported to display a phase-dependent modulation pattern contributing to activation of distal leg muscles during walking, and to reversal of cutaneous-induced reflex responses.35,42–44 Further, it has recently been demonstrated that plantar cutaneous afferents interact with hip proprioceptors modulating the amplitude of both soleus H-reflex and late TA flexion reflex in the same SCI subjects participating in this study.20,26 Based on our findings, it is apparent that plantar cutaneous input results in augmentation of soleus H-reflex and TA flexion excitability in SCI subjects. The augmented reflex excitability, however, ceases when plantar cutaneous input interacts with hip-mediated sensory feedback.20,26 Thus, it is possible to suggest that in standing SCI patients, interaction of plantar cutaneous afferents and hip-mediated input may account as a mechanism in normalizing the exaggerated cutaneous transmission observed in this study. However, it is clear that further research is needed on the sensorimotor integration of these signals in SCI patients during standing and assisted walking.
It is important to note at this point that our study was conducted with subjects seated and that reorganization of the flexion reflex during assisted walking in SCI patients (or even in normal subjects) might result in different response patterns. For example, excitation of the medial plantar nerve innervating the plantar surface of the foot in spinalized cats during walking have mostly inhibitory effects on lumbar motoneurones,9 while after complete removal of cutaneous signals from the foot sole, the incorrect placement of the foot is associated with coactivation of ankle flexors and extensors.8 To conclude, signals from the foot sole participate in complex multisegmental spinal interneuronal circuits affecting motoneuronal excitability of distal flexors and extensors.
Excitation of plantar cutaneous afferents resulted in facilitation of the soleus H-reflex in SCI subjects only during the long C-T intervals tested (30–90 ms), while in normal subjects identical afferent input depressed soleus H-reflex for C-T intervals ranging from 6 to 60 ms (Figure 1b). These findings are in line with our previous observation, where plantar afferent excitation in seated spinal intact subjects induced a significant soleus H-reflex depression at C-T intervals of 0–9 ms.32 Nonetheless, a different modulation pattern for the soleus H-reflex has also been reported. Stimulation of sural and saphenous nerves (purely cutaneous) induces an early inhibition followed by a substantial reflex facilitation in normal subjects.45 No changes on the soleus H-reflex following excitation of plantar cutaneous afferents have also been reported.12,46 However, we used a pulse train to excite plantar cutaneous afferents while in the above studies a single pulse was used.
Non-nociceptive sural nerve stimulation facilitates the ipsilateral TA H-reflex and decreases the heteronymous Ia facilitation exerted from the quadriceps onto soleus α-motoneurones.21 These findings suggest that FRA induce presynaptic inhibition of Ia afferent terminals, and that plantar cutaneous afferents have spinal oligosynaptic connections with the interneurones intercalated in the FRA pathway. In this respect, it has been demonstrated that plantar cutaneous afferents can adjust motoneuronal excitability by modulating the level of both presynaptic and postsynaptic inhibition. Plantar cutaneous afferents depress the actions of Ib reflex pathways to motoneurones,12 increase the reciprocal inhibition from TA to soleus α-motoneurones,46 and modulate transmission in presynaptic inhibitory pathways of both flexor and extensor group Ia afferent fibres in man.13,47 Thus, there is substantial evidence to suggest that signals from the cutaneous afferents of the foot sole interact with spinal inhibitory control systems contributing to the control of movement and posture in humans. The latter is further supported by studies reporting that when signals from the plantar cutaneous afferents are disrupted, other sensory mechanisms cannot compensate the reduced plantar sensitivity during standing,48 and that cutaneous afferents from the foot sole play a regulatory mechanism during standing by adjusting small in amplitude body sways.49
Additionally, excitation of plantar cutaneous afferents during assisted walking in SCI patients modulates soleus H-reflex excitability to a level similar to that observed in neurologically intact subjects.50 This finding supports the idea that sensory afferent feedback from the plantar soles contributes to the motor recovery of SCI patients by normalizing the impaired spinal mechanisms acting on Ia afferents or on α-motoneurones. The latter may be driven through interaction with body loading sensory feedback51 being absent when patients are at rest. In this respect, load applied to the foot sole activating both load mechanoreceptors and skin afferents decrease soleus H-reflex and TA flexion reflex excitability in motor complete SCI subjects.52,53 These findings suggest that the reflex modulation pattern changes in SCI subjects when concurrent input from load receptors is present, and that the mechanisms involved in reflex modulation are spinal in origin.
Potential mechanisms that contributed to the soleus H-reflex and flexion reflex modulation following excitation of tactile plantar cutaneous afferents include changes in the amount of presynaptic inhibition,13 interaction of cutaneous afferents with Ib inhibitory interneurons,12 and segmental interneuronal effects. However, based on the experimental protocol employed, differentiation between post- and presynaptic mechanisms cannot be fully ascribed. To summarize, activation of tactile plantar cutaneous afferents increase soleus H-reflex and flexion reflex excitability only in SCI patients, suggesting that sensory input from cutaneous mechanoreceptors of the foot sole modulate differently spinal reflex excitability in SCI patients.
At this point we should consider the possibility that cutaneous input in relaxed seated SCI patients might contribute to the reflexive enhanced leg muscle activity (or muscle spasms), observed frequently in the clinic especially in motor incomplete SCI patients.54,55 All SCI subjects participating in this study displayed both extensor and flexor spasms, which were triggered not only by changes in body posture (from sit to supine)56 but also by cutaneous stimuli of different nature while seated. Given the multijoint involvement of these spasms and the multisegmental interneuronal organization of the late flexion reflex, it is likely that spasms are mediated by organized interneuronal circuits, acting through polysynaptic spinal reflex pathways, that are involved in locomotion.57 Thus, when SCI patients are at rest, spasms may be regarded as an exaggerated fractional expression of the locomotion cycle. In this line, during assisted walking (in human motor incomplete SCI), they might be reorganized when sensory feedback from load receptors and plantar cutaneous afferents are present. It is clear, however, that the above statements warrant further investigation.
Conclusions
This study has shown that plantar cutaneous afferent input modulates soleus H-reflex and flexion reflex differently in neurologically intact and SCI subjects. The flexion reflex appeared to be different following a lesion to the spinal cord, engaging possibly different neuronal pathways. Although the mechanisms account for the reflex modulation observed in this study cannot be fully ascribed, it is apparent that signals from the cutaneous afferents of the plantar sole interact (and may be transmitted) with organized spinal interneuronal circuits involving several segmental levels of the partly isolated human spinal cord. These interneuronal circuits might be involved with stepping and locomotion. It is clear that further research is needed on the sensorimotor integration and its relative contribution to the final common pathway in human SCI.
Acknowledgments
I express my gratitude to the patients of the Rehabilitation Institute of Chicago for participating on many experimental sessions, acknowledge the continuous support of the staff of the Sensory Motor Performance Program, and thank Dr E Kay for her help during this work.
References
- 1.Sherrington CS. On innervation of antagonistic muscles: sixth note. Proc R Soc London Ser Biol. 1900;66:66–67. [Google Scholar]
- 2.Sherrington CS. Qualitative difference of spinal reflex corresponding with qualitative difference of cutaneous stimulus. J Physiol (London) 1903;30:39–46. doi: 10.1113/jphysiol.1903.sp000980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kniffki KD, Schomburg ED, Steffens H. Convergence in segmental reflex pathways from fine muscle afferents and cutaneous or group II muscle afferents to α-motoneurones. Brain Res. 1981;218:342–346. doi: 10.1016/0006-8993(81)91312-3. [DOI] [PubMed] [Google Scholar]
- 4.Engberg I. Reflexes to foot muscles in the cat. Acta Physiol Scand (Suppl) 1964;62:1–64. [PubMed] [Google Scholar]
- 5.Duysens J, Pearson KG. The role of cutaneous afferents from the distal hindlimb in the regulation of the step cycle of thalamic cats. Exp Brain Res. 1976;24:245–255. doi: 10.1007/BF00235013. [DOI] [PubMed] [Google Scholar]
- 6.Duysens J. Reflex control of locomotion as revealed by stimulation of cutaneous afferents in spontaneously walking premammillary cats. J Neurophysiol. 1977;40:737–751. doi: 10.1152/jn.1977.40.4.737. [DOI] [PubMed] [Google Scholar]
- 7.Conway BA, Scott DT, Riddell JS. The effects of plantar nerve stimulation on long latency flexion reflexes in the acute spinal cat. In: Taylor A, Gladden MN, Durbada R (eds). Alpha and Gamma Motor Systems Plenum Press: New York 1995, pp 593–595.
- 8.Bouyer LJG, Rossignol S. Contribution of cutaneous inputs from the hindpaw to the control of locomotion. II. Spinal cats J Neurophysiol. 2003;90:3640–3653. doi: 10.1152/jn.00497.2003. [DOI] [PubMed] [Google Scholar]
- 9.Côte M-P, Gossard J-P. Step training-dependent plasticity in spinal cutaneous pathways. J Neurosci. 2004;24:11317–11327. doi: 10.1523/JNEUROSCI.1486-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bastiaanse CM, Duysens J, Dietz V. Modulation of cutaneous reflexes by load receptor input during human walking. Exp Brain Res. 2000;135:189–198. doi: 10.1007/s002210000511. [DOI] [PubMed] [Google Scholar]
- 11.van Wezel BMH, van Engelen BMG, Gabreëls FJM, Gabreëls-Festen AAWM, Duysens J. Aβ fibers mediate cutaneous reflexes during human walking. J Neurophysiol. 2000;83:2980–2986. doi: 10.1152/jn.2000.83.5.2980. [DOI] [PubMed] [Google Scholar]
- 12.Pierrot-Deseilligny E, Bergego C, Katz R, Morin C. Cutaneous depression of Ib reflex pathways to motoneurons in man. Exp Brain Res. 1981;42:351–361. doi: 10.1007/BF00237500. [DOI] [PubMed] [Google Scholar]
- 13.Iles JF. Evidence for cutaneous and corticospinal modulation of presynaptic inhibition of Ia afferents from the human lower limb. J Physiol (London) 1996;491:197–207. doi: 10.1113/jphysiol.1996.sp021207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Baldissera F, Hultborn H, Illert M. Integration in spinal neuronal systems. In: Handbook of Physiology, The Nervous System 1981, American Physiological Society: Bethesda, pp 509–595.
- 15.Schomburg ED. Spinal sensorimotor systems and their supraspinal control. Neurosci Res. 1990;7:265–340. doi: 10.1016/0168-0102(90)90008-3. [DOI] [PubMed] [Google Scholar]
- 16.Hagbarth KE. Excitatory and inhibitory skin areas for flexor and extensor motoneurones. Acta Physiol Scand (Suppl) 1952;94:1–57. [PubMed] [Google Scholar]
- 17.Andersen OK, Sonnenborg FA, Arendt-Nielsen L. Modular organization of human leg withdrawal reflexes elicited by electrical stimulation of the foot sole. Muscle Nerve. 1999;22:1520–1530. doi: 10.1002/(sici)1097-4598(199911)22:11<1520::aid-mus6>3.0.co;2-v. [DOI] [PubMed] [Google Scholar]
- 18.Baxendale RH, Ferrell WR. The effect of knee joint discharge on transmission in flexion reflex pathways in decerebrate cats. J Physiol (London) 1981;315:231–242. doi: 10.1113/jphysiol.1981.sp013744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Knikou M, Conway BA. Effects of electrically induced muscle contraction on flexion reflex in human spinal cord injury. Spinal Cord. 2005;43:640–648. doi: 10.1038/sj.sc.3101772. [DOI] [PubMed] [Google Scholar]
- 20.Knikou M, Kay E, Rymer WZ. Modulation of flexion reflex induced by hip angle changes in human spinal cord injury. Exp Brain Res. 2006;168:577–586. doi: 10.1007/s00221-005-0112-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Roby-Brami A, Bussel B. Effects of FRA stimulation on the soleus H-reflex in patients with a complete spinal cord lesion: evidence for presynaptic inhibition of Ia transmission. Exp Brain Res. 1990;81:593–601. doi: 10.1007/BF02423509. [DOI] [PubMed] [Google Scholar]
- 22.Roby-Brami A, Bussel B. Inhibitory effects on flexor reflexes in patients with a complete spinal cord lesion. Exp Brain Res. 1992;90:201–208. doi: 10.1007/BF00229272. [DOI] [PubMed] [Google Scholar]
- 23.Jankowska E, Jukes MGM, Lund S, Lundberg A. The effect of DOPA on the spinal cord. 6. Half centre organization of interneurones transmitting effects from the flexor reflex afferents. Acta Physiol Scand. 1967;70:389–402. doi: 10.1111/j.1748-1716.1967.tb03637.x. [DOI] [PubMed] [Google Scholar]
- 24.Maynard FM, et al. International standards for neurological and functional classification of spinal cord injury. Spinal Cord. 1997;5:266–274. doi: 10.1038/sj.sc.3100432. [DOI] [PubMed] [Google Scholar]
- 25.Ashworth B. Preliminary trial of carisoprodol in multiple sclerosis. Practioner. 1964;192:540–542. [PubMed] [Google Scholar]
- 26.Knikou M. Effects of hip joint angle changes on intersegmental spinal coupling in human spinal cord injury. Exp Brain Res. 2005;167:381–393. doi: 10.1007/s00221-005-0046-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Crone C, Hultborn H, Mazieres L, Nielsen J, Pierrot-Deseilligny E. Sensitivity of monosynaptic test reflexes to facilitation and inhibition as a function of the test reflex size: a study in man and the cat. Exp Brain Res. 1990;81:35–45. doi: 10.1007/BF00230098. [DOI] [PubMed] [Google Scholar]
- 28.Fuhrer MJ. Interstimulus interval effects on habituation of flexor withdrawal activity mediated by the functionally transected human spinal cord. Arch Phys Med Rehabil. 1976;57:577–582. [PubMed] [Google Scholar]
- 29.Andrews BJ, Baxendale RH, Granat MH, Nicol DJ. Long latency flexion reflexes in man may limit restoration of gait in FES in SCI man. J Physiol (London) 1990;420:63. [Google Scholar]
- 30.Roby-Brami A, Bussel B. Long latency spinal reflex in man after flexor reflex afferent stimulation. Brain. 1987;110:707–725. doi: 10.1093/brain/110.3.707. [DOI] [PubMed] [Google Scholar]
- 31.Sherrington CS. Flexion-reflex of the limb, crossed extension-reflex and reflex stepping and standing. J Physiol (London) 1910;40:28–121. doi: 10.1113/jphysiol.1910.sp001362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Knikou M, Rymer WZ. Effects of changes in hip joint angle on H-reflex excitability in humans(erratum in Exp Brain Res 2002; 144: 558) Exp Brain Res. 2002;143:149–159. doi: 10.1007/s00221-001-0978-4. [DOI] [PubMed] [Google Scholar]
- 33.Meinck HM, Kuster S, Benecke R, Conrad B. The flexor reflex-influence of stimulus parameters on the reflex response. Electroenceph Clin Neurophysiol. 1985;61:287–298. doi: 10.1016/0013-4694(85)91095-8. [DOI] [PubMed] [Google Scholar]
- 34.Anden NE, Jukes MG, Lundberg A, Vyklicky L. The effect of Dopa on the spinal cord. 1. Influence on transmission from primary afferents. Acta Physiol Scand. 1966;67:373–386. doi: 10.1111/j.1748-1716.1966.tb03324.x. [DOI] [PubMed] [Google Scholar]
- 35.Duysens J, Tax AAM, Trippel M, Dietz V. Increased amplitude of cutaneous reflexes during human running as compared to standing. Brain Res. 1993;613:230–238. doi: 10.1016/0006-8993(93)90903-z. [DOI] [PubMed] [Google Scholar]
- 36.Dyhre-Poulsen P, Dy C, van der Brand R, Duysens J, Harkema SJ. Cutaneous Reflexes from the Foot During Assisted Stepping in Humans with Clinically Complete Spinal Cord Injury Program No. 515.7, Society for Neuroscience: Washington DC2005.
- 37.Nielsen J, Petersen N, Fedirchuk B. Evidence suggesting a transcortical pathway from cutaneous foot afferents to tibialis anterior motoneurones in man. J Physiol (London) 1997;501:473–484. doi: 10.1111/j.1469-7793.1997.473bn.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Gerrits HL, De Haan A, Hopman MTE, Van Der Woude LHV, Jones DA, Sargeant AJ. Contractile properties of the quadriceps muscle in individuals with spinal cord injury. Muscle Nerve. 1999;22:1249–1256. doi: 10.1002/(sici)1097-4598(199909)22:9<1249::aid-mus13>3.0.co;2-n. [DOI] [PubMed] [Google Scholar]
- 39.Thomas CK, Broton JG, Calancie B. Motor unit forces and recruitment patterns after cervical spinal cord injury. Muscle Nerve. 1997;20:212–220. doi: 10.1002/(sici)1097-4598(199702)20:2<212::aid-mus12>3.0.co;2-4. [DOI] [PubMed] [Google Scholar]
- 40.McCrea DA. Can sense be made of spinal interneuron circuits? Behav Brain Sci. 1992;15:633–643. [Google Scholar]
- 41.Lundberg A. Multisensory control of spinal reflex pathways. In: Granit R, Pomeiano O (eds). Reflex Control of Posture and Movement. Prog Brain Res Elsevier: Amsterdam 1979, pp 11–28. [DOI] [PubMed]
- 42.Duysens J, Tax AA, Trippel M, Dietz V. Phase-dependent reversal of reflexly induced movements during human gait. Exp Brain Res. 1992;90:404–414. doi: 10.1007/BF00227255. [DOI] [PubMed] [Google Scholar]
- 43.Zehr EP, Stein RB, Komiyama T. Function of sural nerve reflexes during human walking. J Physiol (London) 1998;507:305–314. doi: 10.1111/j.1469-7793.1998.305bu.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Spaich EG, Arendt-Nielsen L, Andersen OK. Modulation of lower limb withdrawal reflexes during gait: a topographical study. J Neurophysiol. 2004;91:258–266. doi: 10.1152/jn.00360.2003. [DOI] [PubMed] [Google Scholar]
- 45.Delwaide PJ, Crenna P, Fleron MH. Cutaneous nerve stimulation and motoneuronal excitability: I, soleus and tibialis anterior excitability after ipsilateral and contralateral sural nerve stimulation. J Neurol Neurosurg Psychiatry. 1981;44:699–707. doi: 10.1136/jnnp.44.8.699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Rossi A, Mazzocchio R. Cutaneous control of group I pathways from ankle flexors to extensors in man. Exp Brain Res. 1988;73:8–14. doi: 10.1007/BF00279655. [DOI] [PubMed] [Google Scholar]
- 47.Nakashima K, Rothwell JC, Day BL, Thompson PD, Marsden CD. Cutaneous effects on presynaptic inhibition of flexor Ia afferents in the human forearm. J Physiol (London) 1990;426:369–380. doi: 10.1113/jphysiol.1990.sp018143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Meyer PF, Oddsson LIE, De Luca CJ. The role of plantar cutaneous sensation in unperturbed stance. Exp Brain Res. 2004;156:505–512. doi: 10.1007/s00221-003-1804-y. [DOI] [PubMed] [Google Scholar]
- 49.Kavounoudias A, Roll R, Roll J-P. Foot sole and ankle muscle inputs contribute jointly to human erect posture regulation. J Physiol (London) 2001;532:869–878. doi: 10.1111/j.1469-7793.2001.0869e.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Fung J, Barbeau H. Effects of conditioning cutaneomuscular stimulation on the soleus H-reflex in normal and spastic paretic subjects during walking and standing. J Neurophysiol. 1994;72:2090–2104. doi: 10.1152/jn.1994.72.5.2090. [DOI] [PubMed] [Google Scholar]
- 51.Harkema SJ, Hurley SL, Patel UK, Requejo PS, Dobkin BH, Edgerton VR. Human lumbosacral spinal cord interprets loading during stepping. J Neurophysiol. 1997;77:797–811. doi: 10.1152/jn.1997.77.2.797. [DOI] [PubMed] [Google Scholar]
- 52.Knikou M, Conway BA. Modulation of soleus H-reflex following ipsilateral mechanical loading of the sole of the foot in normal and complete spinal cord injured human subjects. Neurosci Lett. 2001;303:107–110. doi: 10.1016/s0304-3940(01)01718-9. [DOI] [PubMed] [Google Scholar]
- 53.Knikou M. Proprioceptive modulation of flexion withdrawal reflex in spinal cord injured subjects. PhD Thesis, Bioengineering Unit, University of Strathclyde, Scotland, UK, 1999.
- 54.Rudomin P, Schmidt RF. Presynaptic inhibition in the vertebrate spinal cord revisited. Exp Brain Res. 1999;129:1–37. doi: 10.1007/s002210050933. [DOI] [PubMed] [Google Scholar]
- 55.Kuhn R. Functional capacity of the isolated human spinal cord. Brain. 1950;73:1–51. doi: 10.1093/brain/73.1.1. [DOI] [PubMed] [Google Scholar]
- 56.Little JW, Mickelson P, Umlauf R, Britell C. Lower extremity manifestations of spasticity in chronic spinal cord injury. Am J Phys Med Rehab. 1989;68:26–32. doi: 10.1097/00002060-198902000-00009. [DOI] [PubMed] [Google Scholar]
- 57.Steldt RE, Schmit BD. Modulation of coordinated muscle activity during imposed sinusoidal hip movements in human spinal cord injury. J Neurophysiol. 2004;92:673–685. doi: 10.1152/jn.00677.2003. [DOI] [PubMed] [Google Scholar]