

Abstract
Mirror neurons, located in the premotor cortex of macaque monkeys, are activated both by the performance and the passive observation of particular goal-directed actions. Although this property would seem to make them the ideal neural substrate for imitation, the puzzling fact is that monkeys simply do not imitate. Indeed, imitation appears to be a uniquely human ability. We are thus left with a fascinating question: if not imitation, what are mirror neurons for? Recent advances in the study of non-human primate social cognition suggest a surprising potential answer.
Introduction
Few discoveries in cognitive neuroscience have captured the imagination of the scientific community in the way that mirror neurons have. First found in area F5 of the macaque premotor cortex [1,2] (Figure 1), and now indirectly inferred to exist in humans (see [3]), mirror neurons have the remarkable property of activating during both the performance and the observation of particular motor actions. For example, mirror neurons activated when a monkey grasps a raisin undergo equivalent activation when the monkey watches a human experimenter do the same thing [1]. This property would seem to make mirror neurons the ideal neural substrate for cognitive capacities that require a non-egocentric representation of action, most notably imitation. However, despite this seemingly perfect fit, all of the available evidence suggests that monkeys cannot imitate.
Figure 1.
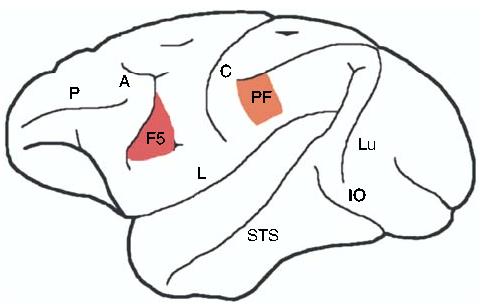
A schematic lateral view of the macaque monkey brain showing approximate regions of mirror neuron activity (adapted from [3]). Mirror neurons were first discovered in area F5 of the premotor cortex [1,2], and have also been reported in area PF of the inferior parietal lobule [23,24]. Labeled structures: A, arcuate sulcus; C, central sulcus; IO, inferior occipital sulcus; L, lateral fissure; Lu, lunate sulcus; P, principal sulcus; STS, superior temporal sulcus.
Indeed, though mirror neurons appear to have evolved at least as early as our most recent common ancestor with Old World monkeys, primatologists have long noted that we humans are unique among primates in our capacity for true imitation (reviewed in [4]). This fact raises two significant questions. First, if not imitation, then what function do mirror neurons serve in non-human primates? Second, how can we account for the emergence of our unique imitative capacity? Here we suggest some potential answers to these questions, motivated by a synthesis of recent results in non-human primate social cognition, human cognitive development and cognitive neuroscience.
Clues from non-human primate social cognition
Although neurophysiological data can tell us much about the properties of mirror neurons, we argue that understanding their core cognitive function will require a broad perspective that incorporates the behavioral level of analysis. Dramatic new developments in the study of non-human primate social cognition seem particularly likely to offer valuable insight.
As recently as ten years ago, the concept of non-human primate social cognition seemed something of an oxymoron. The consensus was that although non-human primates were sophisticated behaviorists, they had no meaningful insight into the mental states of others (see [4]). Recently, however, experimental paradigms that are more thoughtfully matched to non-human primates' competitive social ecology have begun to yield very different results. When competing for limited resources as they do in the wild, non-human primates seem able to reason about other minds in strikingly nuanced ways (reviewed in [5••]).
For example, non-human primates will consider conspecifics' knowledge and perceptual state (i.e. what they can and cannot see and hear) when competing for food. Hare and co-workers [6-8] have demonstrated that chimpanzees (Pan troglodytes) prefer to approach hidden food when a dominant competitor is unable to view the hiding process or is misinformed as to the final location of the food. Work from our own laboratory has demonstrated a similar sensitivity in rhesus macaque monkeys (Macaca mulatta). Macaques take into account both the visual and the auditory awareness of a competitor when attempting to steal food, preferring to steal grapes from an experimenter who cannot see [9••] or hear [10] their approach.
At an even more abstract level, non-human primates appear to use the intentional structure of behavior to differentiate among actions that have nearly identical surface perceptual features. For example, Lyons and Santos (DE Lyons, LR Santos, unpublished data) have demonstrated that capuchin monkeys (Cebus apella) can discriminate between intentional and incidental human actions in order to infer the location of hidden food. In this study, capuchin monkeys watched as a human actor manipulated an opaque container in a goal-directed manner, looking at it and attempting (unsuccessfully) to reach inside and remove something from its interior. A second actor simultaneously handled another container in a manner that, although highly perceptually similar to that of the first actor, was clearly incidental and not goal-directed. When a screen was momentarily raised so that the subject could quickly steal one of the two objects, capuchins showed a strong preference for the container that the first actor had intended to remove something from over the container that was only incidentally manipulated.
Implications: what are mirror neurons for?
We believe that the behavioral work just described offers significant new insight into the cognitive function of mirror neurons. Specifically, the evident sensitivity of non-human primates to the mental states of others is consistent with a mirror neuron system tuned to extract the goal structure of observed action (as opposed to the lower-level kinematic features of the action). In other words, perhaps mirror neurons enable non-human primates to infer the intentions of other agents.
Our existing neurophysiological understanding of mirror neurons is consistent with this speculation. First, merely witnessing an action is insufficient to activate monkey mirror neurons in the absence of an inferable goal. Although mirror neurons will fire when a monkey observes an agent grasping an object, they will not activate when the same grasping action is mimed with no object present [1]. By contrast, mirror neurons can become active when visual access to an action is eliminated but information about the goal of the action is preserved. In an elegant experiment, Umiltàet al. [11] studied monkeys observing a human hand reaching for and grasping a peanut. This grasping action was either completely visible or partially hidden (the subjects saw only the hand reaching behind an opaque screen where a peanut had been previously placed). Remarkably, the sight of the hand disappearing behind the screen hiding the peanut activated the same mirror neurons that were activated by the sight of the peanut being grasped. Thus, the presence of an inferable goal is a necessary and sufficient condition for mirror neurons to become active, whereas simple body movements alone are ineffective. Even more suggestively, Fogassi et al. [12••] have recently demonstrated that monkeys show distinct patterns of mirror neuron activity when observing identical grasping actions depending on the implied underlying intention (i.e. grasping to eat versus grasping to move).
Rizzolatti and co-workers have previously interpreted these results as suggesting that mirror neurons support the function of ‘action understanding’. However, the historic view that non-human primates lacked awareness of mental states forced this argument to balance on a very fine distinction between what it means to understand the ‘goal’ of an action and what it means to understand the ‘intention’ underlying that action. Thus, although Rizzolatti et al. argued that “monkeys, like humans, can infer the goal of an action”, their position was weakened by the arguably inconsistent caveat that “we are not claiming that… the monkeys understood the intention of the agent of the action” ([13], p. 667). We argue that the new behavioral data resolves this inconsistency, and makes tenable the hypothesis that mirror neurons are the neural substrate supporting the intentional understanding of non-human primates.
Why don't non-human primates imitate?
Ironically, the possibility that mirror neurons support intentional understanding could help to explain why non-human primates lack the seemingly simpler ability to imitate. That is, one of the potential benefits of construing behavior in intentional terms is that doing so might facilitate more concise representation: rather than encoding the complicated visual features of an action sequence, the sequence can be compressed to a simpler description of the underlying intention. Although such ‘intentional compression’ would be advantageous in many ways, as with any computational data compression algorithm the disadvantage is that information might be lost. Thus, if mirror neurons are attuned to the intentional structure of behavior, it is possible that non-human primates might simply be unable to encode lower-level visual details of observed actions. Imitation, which depends crucially on such visual details, would thus be rendered impossible.
Why do humans succeed at imitation when non-human primates fail?
But what about humans? What accounts for our unique imitative capacity? If mirror neurons in non-human primates encode intentions, and if that function suppresses imitative behavior as proposed above, then why is imitation in humans not similarly suppressed? One solution to this conundrum may be that the human mirror system is qualitatively different from that of other primates: whereas non-human primate mirror neurons highlight intentions, perhaps the human mirror system extracts a low-level kinematic description of movement—something that would serve as an ideal basis for imitation. This could account for the finding that the human mirror system appears to resonate to arbitrary body movements, such as mimed actions and intransitive gestures [14-18], in a way that monkey mirror neurons do not [3].
In our view, however, a more theoretically compelling alternative is that the human mirror system encodes intentions in a manner broadly consistent with other primates, but that it does so in a flexible fashion that enables multiple levels of intentional granularity. That is, if the human mirror system were capable of extracting not only high-level goals (‘do x’) but also more subtle, subsidiary goals (‘do x in manner y’), this could account for our ability to reproduce not only the overall results of observed actions but also the specific means that were used to achieve them.
Behavioral evidence bearing on this theory comes from the phenomenon of ‘overimitation’, demonstrated most recently in a comparative study of imitation in chimpanzees and human children [19•]. In one condition of this experiment, both groups watched an experimenter remove a prize from a simple ‘puzzle box’ (a transparent cube divided into two compartments) by performing a series of causally irrelevant actions followed by a series of necessary actions (see Figure 2). When chimpanzees were allowed to play with the puzzle, they tended to ignore the irrelevant actions and do only what was necessary to obtain the prize. In other words, they reproduced the observed end state but not the specific actions that led up to it. By contrast, children showed an overwhelming tendency to imitate all of the modeled actions — despite the fact that some were clearly causally irrelevant — thereby reproducing not only the goal of attaining the prize but also the subsidiary goal of doing so in a particular manner (see also [20-22]).
Figure 2.

Diagram showing one sequence of causally irrelevant and necessary actions used by an adult demonstrator to remove a reward from the puzzle object in Horner and Whiten [19•]. (a) During the causally irrelevant actions (yellow background), a bolt on top of the puzzle object was (i) hit with the tool then (ii) pushed aside, revealing a hole leading to an empty upper compartment. (iii) The tool was inserted into this hole. The demonstrator then immediately continued on to the actions that were actually necessary for retrieving the reward (blue background). (iv) An opening on the front of the box was uncovered by lifting a door, (v) then the tool was inserted into this opening and (vi) used to extract the reward. (b) When the walls of the puzzle box were transparent such that the irrelevance of steps i–iii was obvious (see photo), chimpanzees tended to reproduce only the causally necessary actions. By contrast, children showed a strong tendency to precisely imitate the entire demonstrated sequence. Puzzle object diagrams and photo reproduced with permission.
Although connections to the mirror system are clearly speculative at this point, these results may indicate that the human mirror system extracts finer-grained goals than non-human primate mirror neurons do, thereby driving imitation in a species-unique way. Indeed, it may be that extracting goals that encompass both the end state and the manner of action is the default for the human mirror system. This conjecture is consistent with recent work in our own laboratory (DE Lyons, AG Young, FC Keil, unpublished data), demonstrating that young children who watch an adult modeling actions on a novel object lose the ability to determine the causal relevancy of those actions. It is as though children automatically extract the particular manner in which the adult achieves his goal as an intentional aspect of the demonstration, implicitly revising their causal understanding of the manipulated object in order to make sense of irrelevant actions. A mirror system tuned to ‘manner-of-action’ level goals is a plausible neural basis for these behavioral results, as well as for the broader human propensity for imitation. As more detailed results from the human mirror system become available, it will be very exciting to see how they accord with these behaviorally-motivated hypotheses.
Conclusions
As we have described here, one of the most exciting aspects of research on mirror neurons is the degree to which it coheres with recent advances in the domains of non-human primate social cognition and human cognitive development. By combining the unique insights of cognitive neuroscience with the methodological and theoretical tools of these behavioral research traditions, we are beginning to obtain a unified account of primate social cognitive abilities — one that does full justice both to the uniqueness of human capacities and to their continuity with our evolutionary relatives. We hope that our theory of mirror neurons' cognitive function will serve as a useful point of departure for continuing interdisciplinary investigations into the primate mind.
Acknowledgements
DE Lyons was supported by a National Defense Science and Engineering Graduate Fellowship sponsored by the Department of Defense High Performance Computing Modernization Program. LR Santos was supported by Yale University. FC Keil was supported by Yale University and by National Institutes of Health grant R37 HD023922.
References and recommended reading
Papers of particular interest, published within the annual period of review, have been highlighted as:
• of special interest
•• of outstanding interest
- 1.Gallese V, Fadiga L, Fogassi L, Rizzolatti G. Action recognition in the premotor cortex. Brain. 1996;119:593–609. doi: 10.1093/brain/119.2.593. [DOI] [PubMed] [Google Scholar]
- 2.Rizzolatti G, Fadiga L, Fogassi L, Gallese V. Premotor cortex and the recognition of motor actions. Brain Res Cogn Brain Res. 1996;3:131–141. doi: 10.1016/0926-6410(95)00038-0. [DOI] [PubMed] [Google Scholar]
- 3.Rizzolatti G, Craighero L. The mirror neuron system. Annu Rev Neurosci. 2004;27:169–192. doi: 10.1146/annurev.neuro.27.070203.144230. [DOI] [PubMed] [Google Scholar]
- 4.Tomasello M, Call J. Primate cognition. Oxford University Press; Oxford: 1997. [Google Scholar]
- 5••.Lyons DE, Santos LR. Ecology, domain specificity, and the origins of theory of mind: is competition the catalyst? Philosophy Compass. in press. This theoretical review synthesizes new findings on non-human primate social cognition to argue against the conclusion that these animals have no understanding of others' mental states. New experimental evidence motivates the claim that within the ecologically salient context of social competition, non-human primates can reason about other minds in surprisingly subtle ways. [Google Scholar]
- 6.Hare B, Call J, Agnetta B, Tomasello M. Chimpanzees know what conspecifics do and do not see. Anim Behav. 2000;59:771–785. doi: 10.1006/anbe.1999.1377. [DOI] [PubMed] [Google Scholar]
- 7.Hare B, Call J, Tomasello M. Do chimpanzees know what conspecifics know? Anim Behav. 2001;61:139–151. doi: 10.1006/anbe.2000.1518. [DOI] [PubMed] [Google Scholar]
- 8.Tomasello M, Call J, Hare B. Chimpanzees understand psychological states: the question is which ones and to what extent. Trends Cogn Sci. 2003;7:153–156. doi: 10.1016/s1364-6613(03)00035-4. [DOI] [PubMed] [Google Scholar]
- 9••.Flombaum JI, Santos LR. Rhesus monkeys attribute perceptions to others. Curr Biol. 2005;15:447–452. doi: 10.1016/j.cub.2004.12.076. This study provides the first evidence that rhesus macaque monkeys are sensitive to the perceptual state of other agents. A competition paradigm is used to demonstrate that macaques prefer to steal grapes from an experimenter who is unable to see their approach as opposed to an experimenter with full visual awareness. [DOI] [PubMed] [Google Scholar]
- 10.Santos LR, Nissen AG, Ferrugia J. Do rhesus monkeys (Macaca mulatta) know what competitors can and cannot hear? Animal Behaviour. in press. [Google Scholar]
- 11.Umiltà MA, Kohler E, Gallese V, Fogassi L, Fadiga L, Kaysers C, Rizzolatti G. I know what you are doing: a neurophysiological study. Neuron. 2001;31:155–165. doi: 10.1016/s0896-6273(01)00337-3. [DOI] [PubMed] [Google Scholar]
- 12••.Fogassi L, Ferrari PF, Gesierich B, Rozzi S, Chersi F, Rizzolatti G. Parietal lobe: from action organization to intention understanding. Science. 2005;308:662–667. doi: 10.1126/science.1106138. This elegant study demonstrates that monkeys show distinct patterns of mirror neuron activity when observing identical grasping actions depending on the implied underlying intention. These results provide an important neurophysiological counterpart to new behavioral data indicating that monkeys are sensitive to the intentional structure of action. [DOI] [PubMed] [Google Scholar]
- 13.Rizzolatti G, Fogassi L, Gallese V. Neurophysiological mechanisms underlying the understanding of imitation of action. Nat Rev Neurosci. 2001;2:661–670. doi: 10.1038/35090060. [DOI] [PubMed] [Google Scholar]
- 14.Buccino G, Binkofski F, Fink GR, Fadiga L, Fogassi L, Gallese V, Seitz RJ, Zilles K, Rizzolatti G, Freund HJ. Action observation activates premotor and parietal areas in a somatotopic manner: an fMRI study. Eur J Neurosci. 2001;13:400–404. [PubMed] [Google Scholar]
- 15.Craighero L, Bello A, Fadiga L, Rizzolatti G. Hand action preparation influences the responses to hand pictures. Neuropsychologia. 2002;40:492–502. doi: 10.1016/s0028-3932(01)00134-8. [DOI] [PubMed] [Google Scholar]
- 16.Fadiga L, Fogassi L, Pavesi G, Rizzolatti G. Motor facilitation during action observation: a magnetic stimulation study. J Neurophysiol. 1995;73:2608–2611. doi: 10.1152/jn.1995.73.6.2608. [DOI] [PubMed] [Google Scholar]
- 17.Kilner JM, Paulignan Y, Blakemore S-J. An interference effect of observed biological movement on action. Curr Biol. 2003;13:522–525. doi: 10.1016/s0960-9822(03)00165-9. [DOI] [PubMed] [Google Scholar]
- 18.Blakemore S-J, Frith C. The role of motor contagion in the prediction of action. Neuropsychologia. 2005;43:260–267. doi: 10.1016/j.neuropsychologia.2004.11.012. [DOI] [PubMed] [Google Scholar]
- 19•.Horner V, Whiten A. Causal knowledge and imitation/emulation switching in chimpanzees and children. Anim Cogn. 2005;8:164–181. doi: 10.1007/s10071-004-0239-6. In this comparative study of imitation, children and age-matched chimpanzees observed an experimenter obtaining a prize from inside a novel puzzle object using a sequence of causally relevant and irrelevant actions. When the puzzle object was transparent, such that the causal structure of the experimenter's actions was obvious, chimpanzees tended to reproduce only the causally necessary actions when interacting with the puzzle themselves, whereas children imitated the entire modeled sequence. [DOI] [PubMed] [Google Scholar]
- 20.Nagell K, Olguin RS, Tomasello M. Processes of social learning in the tool use of chimpanzees and human children. J Comp Psychol. 1993;107:174–186. doi: 10.1037/0735-7036.107.2.174. [DOI] [PubMed] [Google Scholar]
- 21.Whiten A, Custance DM, Gomez J-C, Teixidor P, Bard KA. Imitative learning of artificial fruit processing in children (Homo sapiens) and chimpanzees (Pan troglodytes) J Comp Psychol. 1996;110:3–14. doi: 10.1037/0735-7036.110.1.3. [DOI] [PubMed] [Google Scholar]
- 22.Want SC, Harris PL. How do children ape? Applying concepts from the study of non-human primates to the developmental study of ‘imitation’ in children. Dev Sci. 2002;5:1–41. [Google Scholar]
- 23.Fogassi L, Gallese V, Fadiga L, Rizzolatti G. Neurons responding to the sight of goal-directed hand/arm movements in the parietal area PF (7b) of the macaque monkey. Soc Neurosci Abstr. 1998;24:154. [Google Scholar]
- 24.Gallese V, Fogassi L, Fadiga L, Rizzolatti G. Action representation and the inferior parietal lobule. In: Prinz W, Hommel B, editors. Attention and Performance XIX. Common Mechanisms in Perception and Action. Oxford University Press; 2002. pp. 334–355. [Google Scholar]