

Abstract
Operant performance of non-food deprived rats (n=8) was assessed under progressive ratio (PR) and concurrent PR-fixed ratio schedules of food pellet and/or vegetable shortening reinforcement. Post operant baselines, rats were matched and divided into 2 groups based upon the schedule of shortening availability: High restriction binge group (H, 1-hr home cage shortening access each week on Monday, Wednesday, and Friday) and Low restriction (L, 1-hr shortening access daily). Chow and water were continuously available; only access to the shortening was restricted. After 8 weeks, operant performance was reassessed. Lever pressing for shortening increased in the H rats for all schedules, but was either unaffected or decreased in the L rats. Pellet responding under the concurrent schedules increased for both groups. The effects of four dosages of (R)-baclofen (0.3–1.8 mg/kg, i.p.) on operant performance were also assessed. For both groups, 1.0 mg/kg baclofen significantly reduced shortening responding relative to saline for all schedules except one, but had no or minimal effect on pellet responding. This suggests a specific effect of baclofen on responding maintained by fat. These results indicate that intermittent episodes of bingeing on fat can increase the reinforcing efficacy of fat and that GABAB receptor activation can attenuate this effect.
Keywords: Baclofen, Binge eating, Dietary fat, Progressive ratio, Reinforcing efficacy
1. Introduction
Intermittent episodes of behavioral excess characterize several disorders, including substance abuse and the bingeing-related eating disorders. These disorders often share co-morbidity and are difficult to treat. Modeling this kind of behavior in animals would facilitate the elucidation of basic mechanisms and the screening of potential therapeutics. Our laboratory has developed a rat protocol for examining intermittent excessive food intake in which limited access to a preferred food is provided to non-food deprived rats (Corwin, 2004; Corwin et al., 1998; Dimitriou et al., 2000; Thomas et al., 2002). Under this protocol non-food deprived rats given 2 hr of access to vegetable shortening three times per week (high restriction) consume significantly more shortening per period of access than non-food deprived rats provided with daily two-hr access (low restriction). This differential amount of shortening consumption raises the question of whether shortening has become more reinforcing to the high-restriction group than to the low-restriction group. Stated otherwise, does the schedule of access to shortening change the reinforcing efficacy of shortening in non-food deprived rats?
One method of investigating the reinforcing efficacy of a stimulus is to use a progressive ratio schedule of reinforcement. In a progressive ratio schedule the value of the ratio requirement increases with each subsequent reinforcer delivery according to some mathematical formula. The highest ratio that the animal completes is defined as the “breakpoint”, which is used as an index of reinforcer efficacy or reinforcer value, i.e. it serves as a measure of “how hard the animal is willing to work” or “how motivated the animal is” to obtain different reinforcers. Breakpoints can be compared to one another with various independent variable manipulations, and have been used extensively in the study of substance abuse (Richardson and Roberts, 1996; Deroche-Gamonet et al., 2004). Progressive ratio schedules also have been used to assess the reinforcing efficacy of food and have been shown to be sensitive to a variety of independent variable manipulations including reinforcer concentration (Cheeta et al., 1995; Hodos, 1961; Reilly, 1999; Sclafani and Ackroff, 2003), reinforcer magnitude (Spear and Katz, 1991), and the deprivation state of the animal (Cheeta et al., 1995; Hodos and Kalman, 1963).
Progressive ratio schedules in conjunction with agonist and/or antagonist injections have also been used to examine drug self-administration in animals to elucidate the neurochemical system(s) mediating drug mechanism(s) of action. Recent research has indicated that GABAB receptors may be involved in substance abuse disorders. Baclofen, a GABAB agonist, has been shown to reduce cocaine, d- and methamphetamine, apomorphine, nicotine, ethanol and heroin self-administration in animals (Brebner et al., 1999, 2000a,Brebner et al., b, 2005; Campbell et al., 1999; Corrigall et al., 2001; Di Ciano and Everitt, 2003; Loh and Roberts, 1990; Paterson et al., 2004; Ranaldi and Poeggel, 2002; Roberts, 1989; Roberts et al., 1996; Stromberg, 2004). The majority of these studies used progressive ratio schedules of reinforcement alone or in combination with other schedules of reinforcement. Of relevance to the present study is research showing that baclofen reduced progressive ratio responding for cocaine at lower dosages than was required to reduce fixed ratio responding for cocaine or for food (Brebner et al., 2000a).
While the above studies have reported that baclofen reduced drug self-administration, numerous other studies have reported that baclofen had either no significant effect on or increased food intake and/or food-maintained responding whether given either peripherally (Brebner et al., 2000a; Ebenezer, 1995, 1996; Ebenezer and Patel, 2004; Ebenezer and Pringle, 1992; Higgs and Barber, 2004; Shoaib et al., 1998) or centrally (Brebner et al., 2000b; Ebenezer, 1990; Echo et al., 2002; Jonaidi et al., 2002; McFarland and Kalivas, 2001; Minano et al., 1992; Stratford and Kelley, 1997; Ward et al., 2000; Wirtshafter et al., 1993; Znamensky et al., 2001). In contrast, our lab has recently reported that baclofen reduced shortening intake under both high and low restriction access conditions at dosages that either had no effect on or stimulated chow intake (Buda-Levin et al., 2005). While this suggests that consumption of shortening under limited access conditions may be similar in some ways to drug self-administration, the behavioral requirements are quite different in the two types of studies. Furthermore, food intake measurements are confounded by satiation that can occur during the test sessions. Therefore, intake is not a sensitive assay for assessing reinforcing efficacy, nor for assessing potential receptors that may modulate reinforcing efficacy.
In order to assess the effects of the shortening access schedule on reinforcing efficacy, the present study examined operant responding for solid vegetable shortening and for food pellets alone under a progressive ratio schedule, and for shortening and food pellets under concurrent progressive ratio fixed ratio, and concurrent fixed ratio schedules of reinforcement. Pre-binge and post-binge responding for solid vegetable shortening and for food pellets was compared in non-food deprived rats maintained on our high and low restriction protocols (Corwin et al., 1998). In order to compare the effects of baclofen more directly to those reported for drug self-administration, the effects of baclofen were assessed during the post-binge behavioral assessment of operant performance.
2. Materials and methods
2.1. Animals
Eighteen male Sprague Dawley rats, 60 days of age and weighing 275–301 g (Harlan, Indianapolis, IN) at the start of the study, were individually housed in hanging stainless steel wire cages in a temperature- and humidity-controlled environment with a 12:12 light:dark cycle. All rats were maintained on a nutritionally complete commercial laboratory rodent diet (Laboratory Rodent Diet 5001, PMI Feeds, Richmond IN; percent of calories as protein: 28.05%, fat: 12.14%, carbohydrate: 59.81%; 3.3 kcal/g) placed in hanging metal food hoppers at the front of the cage. Tap water was freely available. All procedures were approved by the Pennsylvania State University Institutional Animal Care and Use Committee.
2.2. Apparatus
The animals were tested in three identical operant chambers (30×30×30 cm) located in a room adjacent to the vivarium. Each chamber contained a food magazine (Gerbrands model D-1) that delivered 45 mg food pellets (Noyes Precisions Pellets™, PJAI-0045 Research Diets, Inc., New Brunswick, NJ; percent of calories as protein: 25%, fat:11%, carbohydrate:64%; 3.2 kcal/g) into a receptacle located on the right side of the front chamber wall. The nutrient composition of the pellets was based upon the same formulation as the maintenance rodent diet, but the pellets also contained corn syrup (~12% of energy). A solid-fat pump (Razel model A, Razel Scientific Instruments, Inc., Stamford, CN) located outside the operant chamber delivered solid vegetable shortening (Crisco® All-Vegetable shortening, J.M. Smucker Co., Orrville, OH, 9.17 kcal/g) packed into a 30 cc syringe (Becton Dickinson, Franklin Lakes, NJ) through a hole located on the left side of the front chamber wall. A lever (BRS/LVE model CRL-005) located in the middle of the right side of the chamber served as the “food lever”, and an identical lever located in the middle of the front panel of the chamber served as the “shortening lever”. Each lever was present in the chamber only when there was a reinforcement contingency in effect for that lever.
2.3. Operant training
Animals were randomly assigned to two groups and 1-hr experimental training sessions were conducted on Mondays, Wednesdays and Fridays for one group, and on Tuesdays, Thursdays and Saturdays for the other group. During these training sessions only, all rats were food-deprived Mondays through Saturdays by providing approximately 15 g of rodent chow in their home cages 2 hr after the start of the dark cycle. The rats were allowed to eat ad libitum for 24 hr after the last operant session of the week. All rats were trained to lever press under a fixed ratio one (FR1) schedule of food pellet reinforcement, and then exposed to one session each of FR5, FR10, FR15, FR20, FR25, FR30 and FR40 schedules of reinforcement. The sound of the pellet dispenser served as the discriminative stimulus signaling food delivery. Following the training sessions in which food pellets served as the reinforcer, the above training conditions were reinstated but a one-tenth gram (0.1 g) of solid shortening served as the reinforcer for lever pressing. A 5-sec illumination of a light located above the hole where shortening was delivered served as the discriminative stimulus signaling shortening delivery. During this training period, all animals had 1-hr access to shortening either 30 min after the last experimental session of the day or 2 hr prior to the start of the dark cycle on non-session days.
Once the initial operant training was complete, all rats were given free access to the rodent diet and were not food-deprived for the remainder of the study; i.e., the results reported below were obtained under non-food deprived conditions. During this post-training period, all animals had 1-hr access to shortening every day. That is, all of the rats were assigned to the low restriction shortening access protocol at this time. Shortening was provided either after the last experimental session of the day or 2 hr prior to the start of the dark cycle on non-session days. One-hr experimental sessions were conducted on Mondays, Wednesdays and Fridays for half of the rats, and on Tuesdays, Thursdays and Saturdays for the other half. The first experimental session of each day started 1.5 hr prior to the beginning of the dark cycle, the second session started 0.5 hr prior to the beginning of the dark cycle, and the third session of the day started 1 hr after the beginning of the dark cycle. All animals were exposed to the following order of reinforcement schedules and reinforcer types: progressive ratio (PR) for food pellets (PRp), fixed ratio 5 for pellets (FR5p), PR for shortening (PRs), FR 5 for shortening (FR5s), concurrent PR for shortening-FR 5 for food pellets (CPRsFR5p) and concurrent FR5 for shortening-PR for food pellets (CFR5sPRp).
The choice of operant schedules was based upon other reports in which comparable schedules were used when responding was maintained by cocaine and food (Brebner et al., 1999, 2000a,Brebner et al., b; Roberts et al., 1996). Under the PR schedule the first reinforcer was contingent on a single lever press. Thereafter, the number of lever presses required to obtain subsequent reinforcers was incremented through the following exponential progression: 1, 2, 4, 6, 9, 12, 15, 20, 25, 32, 40, 50, 62, 77, 95, 118 (Richardson and Roberts, 1996). This schedule has been used extensively for the study of drug self-administration (Brebner et al., 2000a,b; Mobini et al., 2000; Roberts et al., 1996), and a similar schedule has been used for the study of sucrose-maintained behavior (Brennan et al., 2001). An exponential PR was used because it generates high levels of responding while avoiding confounds associated with high intakes (e.g. satiation) during the session (Richardson and Roberts, 1996). FR, on the other hand, provides a measure more comparable to free feeding (Kissileff, 1970).
The concurrent schedules were included in order to determine the specificity of baclofen effects later in the study. This was important, as baclofen can have motor suppressant effects (McManus and Greenshaw, 1991), which could potentially inhibit responding. A previous study had shown the usefulness of a concurrent schedule in which PR responding was reinforced by cocaine, and FR responding was reinforced with food pellets. Baclofen reduced PR responding for cocaine in that study, while having no effect on FR responding for food pellets (Brebner et al., 2000a). This showed that the rats could still perform an operant response during the time that baclofen reduced responding maintained by cocaine, i.e. the inhibition of responding induced by baclofen was not due to motoric disruption. The use of these schedules, then, was designed to provide a measure of behavioral specificity, and to facilitate comparisons to previously published work.
In the present study, breakpoint was defined as the number of reinforcers earned. Stability criterion for the PR schedules was defined as three consecutive sessions with a breakpoint within a range of three. Stability criterion for the FR schedules was three consecutive sessions with no more than a 20% difference between sessions for the total reinforcers obtained. Stability criterion for the concurrent schedules was based on the PR performance.
2.4. Shortening feeding protocols
After the last session of the operant training period, the animals were matched by body weight, amount of chow consumed in the home cage, amount of shortening consumed in the home cage, number of reinforcers earned under the PRs, and PRp schedules of reinforcement [F(1,16)<1, for all] and divided into two groups. The high restriction group (H) had 1 hr of access to shortening 2 hr prior to the start of the dark cycle in the home cage on Mondays, Wednesdays and Fridays, while the low restriction group (L) had 1 hr of access to shortening in the home cage 2 hr prior to the start of the dark cycle every day. This condition lasted for 8 weeks to ensure reliable binge-type eating by the H group. No operant sessions were conducted during this eight-week exposure to the shortening feeding protocols.
2.5. Operant schedules of reinforcement after exposure to the shortening feeding protocols
At the end of the eight-week home cage shortening condition, all animals were tested under the following order of schedules of reinforcement and type of reinforcer during 1-hr sessions three times a week as stated above: PR for shortening (PRs); concurrent PR for shortening-FR5 for food pellets (CPRs-FR5p), concurrent FR5 for shortening-PR for food pellets (CFR5s-PRp), PR for food pellets (PRp), and concurrent FR1 for shortening-FR1 for pellets (CFR1s-FR1p). The CFR1s-FR1p schedule was included in order to approximate home cage feeding conditions within the operant environment. Performance under each of the schedules was determined to be stable as described above. Testing during this phase of the study was performed under non-food deprived conditions. The H group was tested on Mondays, Wednesdays, and Fridays and the L group was tested on Tuesdays, Thursdays and Saturdays. Both groups were given 1-hr access to shortening in their home cages 30 min after the last operant session of the day. On non-operant testing days, the L group was given 1-hr access to shortening in their home cage at the time they would have received shortening on operant testing days.
2.6. Baclofen testing
The effect of (R)-baclofen (Tocris, Ellisville, MO) pretreatment (0.3, 0.6, 1.0 and 1.8 mg/kg, IP, volume 1 ml/kg, administered 30 min prior to the session) was examined after the performance under each schedule stabilized. All rats received all dosages, with dosages assigned to each rat using a uniform Latin square design. Baclofen was administered on Mondays and Fridays to the H group and Tuesdays and Saturdays to the L group.
2.7. Statistics
SAS v.9.1 (SAS Institute, Cary, NC) was used to analyze the data. Between group t-tests were used to analyze the average 1-hr chow (kcal), 1-hr shortening (kcal), and 1-hr total intake (kcal) during the last 2 weeks of the shortening feeding protocol period. Daily total energy intake was also analyzed using between group t-tests during the last 2 weeks of the shortening feeding protocol. Reinforcers earned during the operant sessions prior to, and after, exposure to the shortening feeding protocols were compared using paired t-tests. Responding during the final three sessions of each operant schedule was averaged for the analyses. The baclofen results were analyzed using two-way (group×-dosage) ANOVA (proc GLM), followed by one-way within group ANOVA and Tukey's HSD to determine the effect of dosage on each outcome variable within each group. Alpha was set at 0.05 for results to be considered significant. If F was <1, then results were not significant and p-values are not reported.
3. Results
3.1. Intake during the shortening feeding protocols
Home cage consumption of both shortening and chow during the 1-hr access period was significantly affected by the schedule of shortening access (Fig. 1A). During this 1-hr period the H group (Monday, Wednesday, Friday access) consumed significantly more energy from shortening (t-test: p <0.01), significantly more energy from chow (t-test: p <0.0001) and significantly more total energy (t-test: p<0.01) than did the L group (daily shortening access). Furthermore, the total daily 24-hr energy intake was also significantly influenced by the schedule of shortening access (Fig. 1B). The H group consumed significantly more energy than the L group on days when shortening was available, but consumed significantly less energy than the L group on days when shortening was unavailable. This overeat/undereat or “sawtooth” pattern of energy intake was similar to that previously reported (Corwin, 2004; Corwin et al., 1998; Dimitriou et al., 2000; Thomas et al., 2002).
Fig. 1.
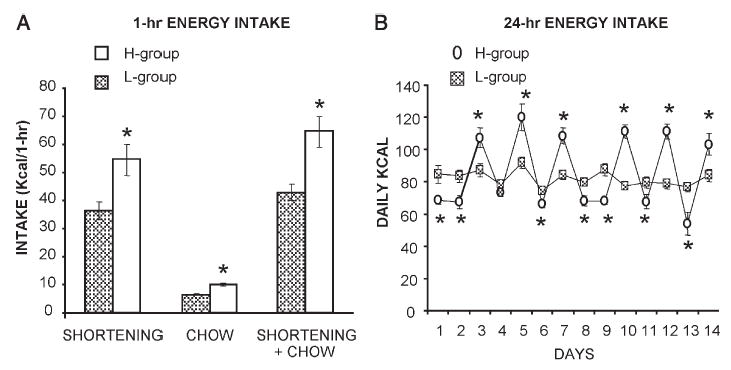
(A) One hr shortening intake, chow intake and total intake averaged for the last 2 weeks of the shortening feeding protocol for non-food deprived rats on the high restriction (H) (1-hr shortening on Mondays, Wednesdays, and Fridays) and low restriction protocols (L) (1-hr shortening daily). * indicates significant differences between the H and L groups. (B) Total (shortening and chow) 24-hr intake (kcal) for the H and L groups during the last 2 weeks of the shortening feeding protocols. * indicates significant differences between the H and L groups.
3.2. Comparison of operant performance before and after exposure to the shortening feeding protocols
There were significant differences in operant performance within each group when behavior prior to and after the shortening feeding protocols was compared (Figs. 2 and 3). In general, responding maintained by shortening increased in the H group, but was unaffected or decreased in the L group. Specifically, the H group earned significantly more shortening deliveries after the feeding protocols than before the feeding protocols under the PRs (t-test: p<0.01) schedule (Fig. 2A) and both concurrent CPRs-FR5p (t-test: p<0.05) and CFR5s-PRp (t-test: p<0.001) schedules (Fig. 3, left panels). In contrast, the L group earned the same number of shortening deliveries under the PRs (t-test: ns) and concurrent CFR5s-PRp (t-test: ns) schedules, but significantly fewer shortening deliveries under the CPRs-FRp (t-test: p<0.001) schedule.
Fig. 2.
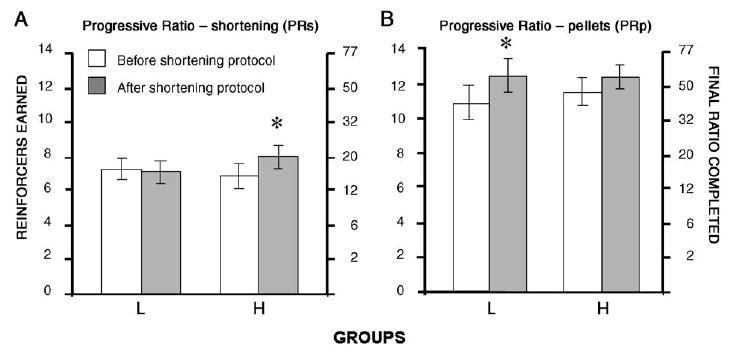
Operant performance (number of reinforcer deliveries and final ratio completed) averaged for the last three sessions under a progressive ratio schedule of reinforcement when either shortening (A) or food pellets (B) served as reinforcers for H and L rats. * indicates significant difference before and after exposure to the shortening feeding protocols.
Fig. 3.
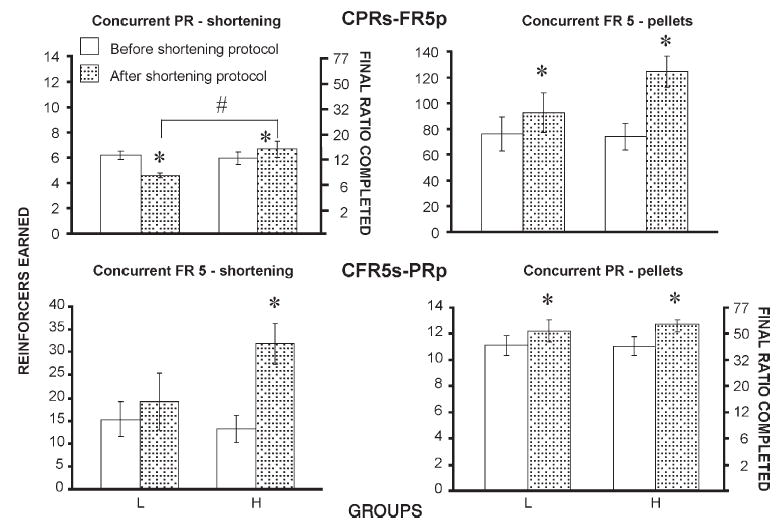
Operant performance (number of reinforcer deliveries and final ratio completed) averaged for the last 3 sessions under concurrent progressive ratio-fixed ratio schedules of reinforcement when both shortening and food pellets served as reinforcers for H and L rats. Top graphs: concurrent PR (shortening)-FR5 (pellets). Bottom graphs: concurrent PR (pellets)-FR5 (shortening). Left graphs: responding for shortening. Right graphs: responding for pellets. * indicates significant difference before and after exposure to the shortening feeding protocols. # indicates significant difference between H and L groups.
Between group differences were observed only under the CPRs-FR5p schedule of reinforcement. Under this schedule the H group earned significantly more shortening deliveries (t-test: p<0.01) than the L group (Fig. 3, top left panel).
Responding maintained by pellets, on the other hand, generally increased in both groups. The H group earned significantly more food pellets under the concurrent, CPRs-FR5p (t-test: p<0.001) and CFR5s-PRp (t-test: p<0.05) schedules (Fig. 3, right panels), and a nearly identical number under the PRp (t-test: p=0.0737) schedule (Fig. 2B). The L group also earned significantly more food pellets under both concurrent CPRs-FR5p (t-test: p<0.05) and CFR5s-PRp (t-test: p<0.05) schedules, as well as the PRp (t-test: p<0.01) schedule.
3.3. Post-binge concurrent FR1 pellets FR1 shortening
There was a significant difference between the H and L groups under the CFR1s-FR1p schedule. The H group earned significantly more (t-test: p<0.01) shortening deliveries than the L group. However, there was no significant difference (t-test: ns) between the groups in the number of pellets earned. The actual number of reinforcer deliveries for each type of reinforcer for each schedule pre- and post-shortening feeding protocols is shown in Table 1.
Table 1.
Reinforcers earned before (pre-binge) and after (post-binge) shortening feeding protocols a
| H group
|
L group
|
|||||||
|---|---|---|---|---|---|---|---|---|
| Food pellet deliveries
|
Shortening deliveries
|
Food pellet deliveries
|
Shortening deliveries
|
|||||
| Schedule | Pre-binge | Post-binge | Pre-binge | Post-binge | Pre-binge | Post-binge | Pre-binge | Post-binge |
| PRs | 6.93 (0.76) | 8.00b (0.68) | 7.26 (0.62) | 7.07 (0.60) | ||||
| CPRsFR5p | 73.93 (10.48) | 124.15 b (12.24) | 5.96 (0.49) | 6.67b, c (0.63) | 75.85 (13.11) | 92.56b (15.24) | 6.19 (0.32) | 4.59b (0.21) |
| CFR5sPRp | 11.04 (0.78) | 12.59 b (0.48) | 13.07 (2.99) | 31.81b (4.50) | 11.11 (0.79) | 12.22b (0.83) | 15.19 (3.88) | 19.15 (6.20) |
| PRp | 11.56 (0.78) | 12.39 (0.67) | 10.85 (0.97) | 12.44 (0.94) | ||||
| CFR1sFR1p | 138.11 (15.38) | 48.56c (6.56) | 102.85 (15.34) | 24.33 (4.33) | ||||
Data are presented as number of deliveries (se).
p<0.05 relative to before exposure to shortening feeding protocols.
p<0.05 H vs. L.
3.4. Post-binge baclofen administration
Baclofen decreased responding in a dose-related manner for all schedules (main effect of dosage p<0.0001 for all schedules; Figs. 4–6). In general, a lower dosage of baclofen was required to significantly reduce responding for shortening than was required to reduce responding for food pellets. For both the H and L groups, 1.0 mg/kg baclofen significantly reduced shortening responding relative to saline (Tukey's HSD p<0.05 for all schedules except CPRs-FR5p in the L group), but had no or minimal effect on pellet responding (Tukey's HSD NS for all schedules except p<0.05 CFR5s-PRp in the H group). For both the H and L groups, 1.8 mg/kg baclofen significantly reduced pellet and shortening responding for all schedules (Tukey's HSD p<0.05 for all).
Fig. 4.
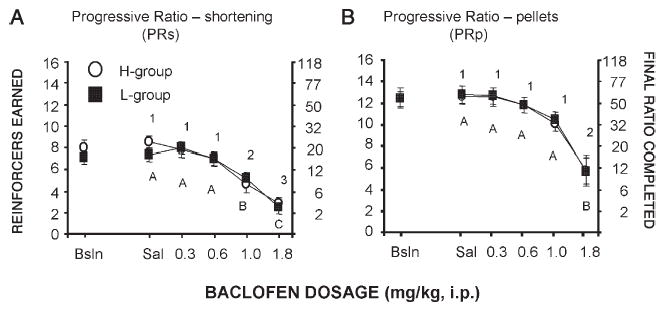
Effects of baclofen on operant performance (number of reinforcer deliveries and final ratio completed) under a progressive ratio schedule of reinforcement when either shortening (A) or food pellets (B) served as reinforcers for H and L rats. Points that have different numbers (H group) or letters (L group) are significantly different from each other; points that share the same numbers (H group) or letters (L group) are not statistically different from each other.
Fig. 6.
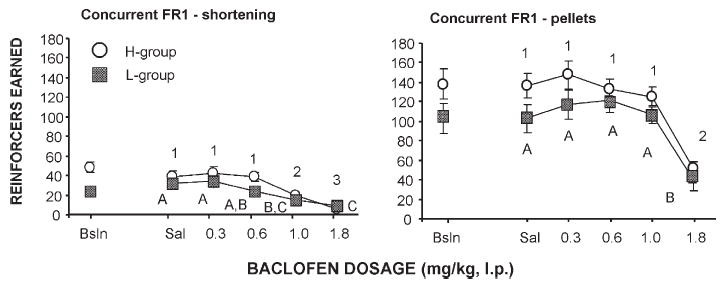
Effects of baclofen on operant performance (number of reinforcer deliveries and final ratio completed) under a concurrent fixed ratio 1 (shortening)-fixed ratio 1 (pellets) schedule of reinforcement for H and L rats. Points that have different numbers (H group) or letters (L group) are significantly different from each other; points that share the same numbers (H group) or letters (L group) are not statistically different from each other.
Responding of the H group was significantly greater than that of the L group during the fixed ratio pellet components of the CPRs-FRp (main effect of group: F(1,80) = 14.42, p<0.001), and CFR1s-FR1p (main effect of group: F(1,80)= 7.88, p<0.01). Main effects did not reveal any other differences between the H and L groups. There were no significant group by dosage interactions for any of the schedules.
For both groups under all schedules, saline responding was not significantly different from the post-binge pre-drug baselines and these baselines did not change during baclofen testing on non-drug days.
3.5. Body weight
Body weights did not differ between the groups at any time during the study, but did increase steadily throughout the study duration (main effect of time, F(2,48)=43.50, p<0.0001; no main effect of group; no time×group interaction). Initial body weights for the H and L groups were 491±13 and 490±8, respectively; terminal body weights were 591±16 and 610±13, respectively.
4. Discussion
The results of the present study demonstrate that 1-hr access to shortening was sufficient to generate bingeing behavior in the highly restricted group (H group) relative to the less restricted group (L). These results are consistent with the results reported previously (Corwin, 2004; Corwin et al., 1998; Dimitriou et al., 2000; Thomas et al., 2002) in which access to shortening was provided on the same schedules of access for 2-hr periods. The intake of the H group, relative to the L group, fit the definition of bingeing: consumption of a greater quantity of a given substance in the same time period than would normally be consumed under similar circumstances (DSM-IV).
The results of the present study also demonstrate that operant methodology used to assess the reinforcing efficacy of drugs can also be used to assess the reinforcing efficacy of solid fat, even in non-food deprived rats. This is an important advance, because solid fat is more highly preferred than oil in rats (Lucas et al., 1989), and because most of the fatty foods that promote excessive consumption and craving in humans are solids. Solid fats, therefore, are potentially powerful reinforcers that could be used in models of craving, loss of control, relapse, etc. The present results are unique because comparisons were made by manipulating the shortening access feeding protocol, rather than by manipulating deprivation state, reinforcer concentration, or reinforcer magnitude as has been done by others (Cheeta et al., 1995; Hodos, 1961; Hodos and Kalman, 1963; Reilly, 1999; Sclafani and Ackroff, 2003). This suggests that the manner in which fat is consumed and/or its availability can influence its reinforcing properties.
Under the three operant conditions (PRs, CPRs-FR5p, and CFR5s-PRp) in which shortening was available both before and after exposure to the shortening access feeding protocols, the H group earned significantly more shortening reinforcers post-binge than pre-binge, while the L group either earned the same or significantly fewer shortening reinforcers. The H group also consumed significantly more shortening post-binge than the L group in the home cage condition as noted above. Furthermore, the H group earned significantly more shortening reinforcers under the CFR1-FR1p schedules than the L group lending support to the notion that low fixed ratio schedule requirements are comparable to consummatory behaviors outside of the context of an experimenter defined operant. The H group also earned significantly more shortening reinforcers than the L group post-binge under the CPRs-FR5p schedule due to a combination of a decrease in the number of reinforcers earned by the L group and an increase in the number of reinforcers earned by the H group. The differences in operant responding are particularly striking, since neither group was food-deprived; the only difference between the two groups was the home cage shortening access feeding protocol.
Aside from the concurrent PR-shortening/FR5-food pellet and the concurrent FR1-shortening/FR1-food pellet schedules, no other between group differences were observed. The lack of between group differences in post-binge progressive ratio responding may have been due to the particular progressive ratio schedule that was used. Hodos and Kalman (1963) showed that altering the response cost of PR schedules could change the shape of the response functions obtained. In the present experiment the PR values were those used to examine drug self-administration. It is possible that a different PR schedule might have shown more consistent breakpoint differences for shortening between the two groups.
For both groups under the CFR5s-PRp and CPRs-FR5p schedules the number of food pellet reinforcers earned post-binge was significantly greater than pre-binge. The relatively consistent increase in earned pellet reinforcers post-binge from pre-binge for both groups likely reflected age and weight increases over the course of the study. Both groups weighed significantly more at the end of the study relative to their starting weights; however, the weights of the two groups were not statistically different at any point. This increase in responding for pellets makes the decrease in responding under the CPRs-FR5p schedule for shortening by the L group even more striking. Despite the increase in body weight and in the reinforcing efficacy of the food pellets, the reinforcing efficacy of shortening for the L group did not increase.
The high level of responding for pellets relative to shortening was surprising and may be due to the different energy densities (3.2 kcal/g for the pellets, 9.17 kcal/g for the shortening) as well as to the fact that the pellets were only available during the 1-hr operant sessions. That is, the pellets were only available under a high restriction access schedule, and were never provided in the home cage. Furthermore, the addition of corn syrup likely made the pellets quite palatable to the rats. It is unlikely that the shortening intake per se promoted high responding for the pellets, as others have shown that adult rats maintained on a high-fat diet do not consume more sugar solution than do rats maintained on a high carbohydrate diet (Reed and Friedman, 1990). However, whether limited access to fat would enhance responding for other palatable foods to which access is limited is not known.
Baclofen significantly decreased shortening responding at a lower dosage (1.0 mg/kg) for all schedules than was required to significantly decrease pellet responding (1.8 mg/kg), indicating that the effects of the 1.0 mg/kg dosage of baclofen on shortening-maintained responding were not due to generalized behavior disruption. While the different levels of baseline responding between the shortening and the pellets may have influenced the baclofen effects, there are reasons to believe that this may not have been the case. Under the concurrent schedules, for instance, there were differential amounts of responding between schedules for the same reinforcer, yet the dosage of baclofen required to significantly decrease responding for each of the reinforcers was the same. For example, during the CPRs-FR5p schedule the total number of responses for pellets under the FR5 was 2 to 2.5 times greater than the total number of responses for pellets under the PR schedule of the CFR5s-PRp schedule. Similarly, the total number of responses for shortening under the FR5 schedule during the CFR5s-PRp was 2 to 2.5 times greater than the total number of responses for shortening under the PR schedule of the CPRs-FR5p schedule. Nonetheless, baclofen reduced responding for shortening at a lower dosage than was required to reduce responding for pellets. This selective effect of baclofen was similar to that observed in cocaine self-administration studies (e.g., Brebner et al., 2000a; Roberts et al., 1996) with comparable schedule arrangements. In those studies baclofen significantly decreased the breakpoint for cocaine-maintained responding at a dosage that had no significant effect on FR5 or PR food-maintained responding. Although baclofen had differential effects on responding maintained by shortening vs. food pellets in the present study, the effects of baclofen were similar in the H and the L groups. The fact that the H and L rats were equally sensitive to the effects of baclofen on shortening-maintained operant responding is consistent with a recent report in which baclofen had comparable effects on shortening intake in H and L rats under home cage feeding conditions (Buda-Levin et al., 2005).
What is inconsistent between this study and previous reports is the effect of baclofen on food-maintained responding. In the present report, baclofen reduced responding maintained by food, whereas in most previous reports baclofen either had no effect on, or stimulated, food intake and food-maintained responding at low to moderate dosages (Brebner et al., 2000a,b; Ebenezer, 1990, 1995, 1996; Ebenezer and Patel, 2004; Ebenezer and Pringle, 1992; Echo et al., 2002; Higgs and Barber, 2004; Jonaidi et al., 2002; McFarland and Kalivas, 2001; Minano et al., 1992; Shoaib et al., 1998; Stratford and Kelley, 1997; Ward et al., 2000; Wirtshafter et al., 1993; Znamensky et al., 2001). Baclofen did not reduce food intake, or responding for food, in these studies regardless of the environmental conditions used. In some of the studies, the rats were food-deprived in some form, from 16 hr to 80% free-feeding weight, while in other studies the rats were non-food deprived. In some studies the experimenter-defined operant behavior varied from lever pressing to nose poking to runway performance, while in other studies the defined operant was eating and licking a particular food substance. Additionally, the schedule of reinforcement in these studies varied from progressive ratio to fixed ratios of one to five, to runway length, to simply eating and licking. Relatively high dosages of baclofen were generally required to reduce food intake or food-maintained responding, suggesting that these reductions may have been due to non-specific motoric disruptions (Bungo et al., 2003; Paterson et al., 2004; Zarrindast et al., 1989).
The reductions in both shortening and pellet-maintained responding at the highest (R)-baclofen dosage in the present study (1.8 mg/kg) suggest that this dosage induced a generalized suppression of behavior. Although dosages higher than 1.8 mg/kg reportedly did not induce behavioral suppression in other studies, the racemate was generally used (or the chiral form was not reported). Witczuk et al. (1980) determined that the pharmacological activity of the (R)-enantiomer was 4.2–9.2-fold greater than the (S) form and 1.4 to 1.9-fold greater than the racemate. Since baclofen is a muscle relaxant, it would not be surprising for behavioral disruption to be induced by a lower dosage of the active (R)-isomer than would be required of the racemate. Surprisingly, our lab has recently reported that 1.8 mg/kg (R)-baclofen either had no effect on or significantly increased chow intake in non-food deprived rats given a choice between shortening and powdered chow in their home cages (Buda-Levin et al., 2005). The reductions in operant responding at 1.8 mg/kg in the present study may reflect non-specific behavioral suppressant effects that were not seen under the relatively lower response cost of home cage eating in the previous report. The possible non-specific effects would also have been more apparent in the present study, since there was less time for recovery to occur; the test sessions were only 1 hr in length in the present study, whereas in our home cage study the test sessions were 2-hr long.
The distribution of responding throughout the session also supports a baclofen-induced non-specific disruption of behavior at the 1.8 mg/kg dosage. In general, a major portion of the responding for shortening and/or pellets under all of the schedules occurred in the first 10 min of the session when saline and lower dosages of baclofen were administered. Furthermore, the least amount of responding occurred during the last 10 min of the session. When 1.8 mg/kg of baclofen was administered, the distribution of responding shifted such that the least amount of responding occurred in the first 10 min and then responding gradually increased throughout the session. For example, under the CPRs-FR5p schedule of reinforcement approximately 35%–40% of the total session responses for both pellets and shortening occurred in the first 10 min of the session, and approximately 5%–8% of the total session responses occurred in the last 10 min for both groups when saline or lower dosages of baclofen were administered. Pretreatment with 1.8 mg/kg of (R)-baclofen decreased responding for both pellets and shortening to 0%–12% of the total sessions responses during the first 10 min. However, responding during the last 10 min of the session rebounded to approximately 30%–65% of the total session responses for both pellets and shortening. This shift in the response distributions is consistent with the idea that the 1.8 mg/kg dosage reduced responding due to non-specific effects.
The pattern of responding in the present study is different from that reported by Brebner et al. (2000a). In the Brebner et al. (2000a) study, only the food lever was available during the first 5 min of the experimental session, after which both the food- and cocaine levers were available and responding was reinforced under a concurrent PR-cocaine/FR1-food schedule of reinforcement. When the 3.2 mg/kg dosage of racemic baclofen was administered, the animals responded at high rates on the food lever during the first 5 min. However, when the cocaine lever was made available, animals switched from the food lever and responded on the cocaine lever for the next hr. They then resumed responding on the food lever. Two factors may account for the differences between the present results and those reported by Brebner et al. (2000a). The first is that the animals in the Brebner et al. (2000a) study were 16-hr. food-deprived, while the animals in the current study were not food-deprived. The second factor may be that food-maintained responding in the Brebner et al. (2000a) study was recorded for the first 2 hr of the experimental sessions, while the sessions in the current study were conducted for only 1 hr.
In summary, several new findings are reported. First, solid vegetable shortening can serve as a reinforcer in non-food deprived rats under several different schedules of reinforcement. Second, exposure to a schedule of relatively highly restricted shortening access in non-food deprived rats increases the reinforcing efficacy of the shortening, as indicated by increased progressive ratio responding. Third, baclofen reduces responding for shortening at a lower dosage (1.0 mg/kg) than what is required to reduce responding for food pellets. This series of investigations indicates the usefulness of the limited access protocol for enhancing the reinforcing efficacy of a natural reinforcer, i.e. dietary fat. Furthermore, these findings extend previous results obtained with baclofen in the home cage to effects of baclofen on fat-maintained operant responding. Taken together, these results support the idea that conditions of limited access (such as occurs with “forbidden” foods or abused substances) may enhance the reinforcing efficacy of the target commodity, and that activation of GABAB receptors can attenuate this effect.
Fig. 5.
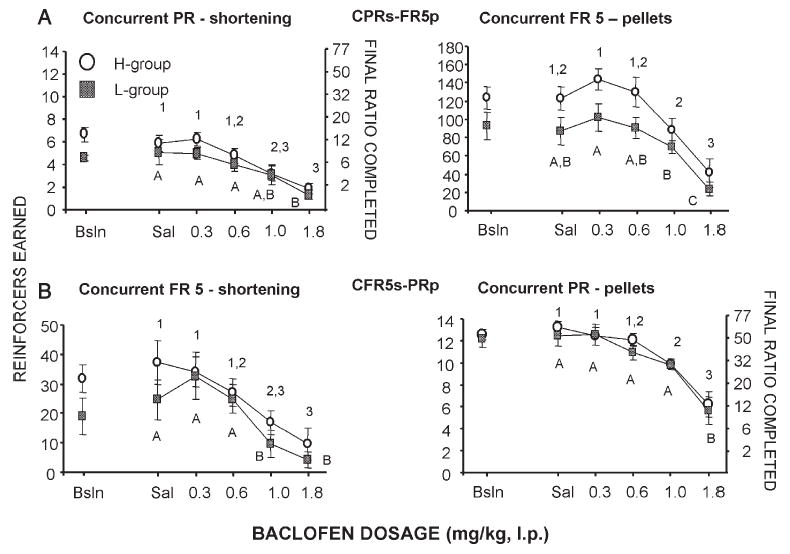
Effects of baclofen on operant performance (number of reinforcer deliveries and final ratio completed) under concurrent progressive ratio-fixed ratio schedules of reinforcement when both shortening and food pellets served as reinforcers for H and L rats. Top graphs: concurrent PR (shortening)-FR5 (pellets). Bottom graphs: concurrent PR (pellets)-FR5 (shortening). Left graphs: responding for shortening. Right graphs: responding for pellets. Points that have different numbers (H group) or letters (L group) are significantly different from each other; points that share the same numbers (H group) or letters (L group) are not statistically different from each other.
Acknowledgments
The authors gratefully acknowledge the support provided from NIH RO1 MH067943 (RLC).
References
- Brebner K, Froestl W, Andrews M, Phelan R, Roberts DCS. The GABAB agonist CGP 44532 decreases cocaine self-administration in rats: demonstration using a progressive ratio and a discrete trials procedure. Neuropharmacology. 1999;38:1797–804. doi: 10.1016/s0028-3908(99)00094-5. [DOI] [PubMed] [Google Scholar]
- Brebner K, Phelan R, Roberts DCS. Effect of baclofen on cocaine self administration in rats reinforced under fixed-ratio 1 and progressive ratio schedules. Psychopharmacology. 2000a;148:314–21. doi: 10.1007/s002130050056. [DOI] [PubMed] [Google Scholar]
- Brebner K, Phelan R, Roberts DCS. Intra-VTA baclofen attenuates cocaine self-administration on a progressive ratio schedule of reinforcement. Pharmacol Biochem Behav. 2000b;66:857–62. doi: 10.1016/s0091-3057(00)00286-0. [DOI] [PubMed] [Google Scholar]
- Brebner K, Ahn S, Phillips AG. Attenuation of d-amphetamine self-administration by baclofen in the rat: behavioral and neurochemical correlates. Psychopharmacology. 2005;177(4):409–17. doi: 10.1007/s00213-004-1968-6. [DOI] [PubMed] [Google Scholar]
- Brennan K, Roberts DCS, Anisman H, Merali Z. Individual differences in sucrose consumption in the rat: motivational and neurochemical correlates of hedonia. Psychopharmacology. 2001;157:269–76. doi: 10.1007/s002130100805. [DOI] [PubMed] [Google Scholar]
- Buda-Levin A, Wojnicki FHE, Corwin RL. Baclofen reduces fat intake under binge-type conditions. Physiol Behav. 2005;86:176–84. doi: 10.1016/j.physbeh.2005.07.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bungo T, Izumi T, Kawamura K, Takagi T, Ueda H, Furuse M. Intracerebro-ventricular injection of muscimol, baclofen or nipecotic acid stimulates food intake in layer-type, but not meat-type, chicks. Brain Res. 2003;993:235–8. doi: 10.1016/j.brainres.2003.09.017. [DOI] [PubMed] [Google Scholar]
- Campbell UC, Lac ST, Carroll ME. Effects of baclofen on maintenance and reinstatement of intravenous cocaine self-administration in rats. Psycho-pharmacology. 1999;143:209–14. doi: 10.1007/s002130050937. [DOI] [PubMed] [Google Scholar]
- Cheeta S, Brooks S, Willner P. Effects of reinforcer sweetness and the D2/D3 antagonist raclopride on progressive ratio operant performance. Behav Pharmacol. 1995;6(2):127–32. [PubMed] [Google Scholar]
- Corrigall WA, Coen KM, Zhang J, Adamson KL. GABA mechanisms in the pedunculopontine tegmental nucleus influence particular aspects of nicotine self-administration selectively in the rat. Psychopharmacology. 2001;158 (2):190–7. doi: 10.1007/s002130100869. [DOI] [PubMed] [Google Scholar]
- Corwin RL. Binge-type eating induced by limited access in rats does not require energy restriction on the previous day. Appetite. 2004;42:139–42. doi: 10.1016/j.appet.2003.08.010. [DOI] [PubMed] [Google Scholar]
- Corwin RL, Wojnicki FHE, Fisher JO, Dimitriou SG, Rice HB, Young MA. Limited access to a dietary fat option affects ingestive behavior but not body composition in male rats. Physiol Behav. 1998;65:545–53. doi: 10.1016/s0031-9384(98)00201-7. [DOI] [PubMed] [Google Scholar]
- Di Ciano P, Everitt BJ. The GABA(B) receptor agonist baclofen attenuates cocaine- and heroin-seeking behavior by rats. Neuropsychopharmacology. 2003;28:510–8. doi: 10.1038/sj.npp.1300088. [DOI] [PubMed] [Google Scholar]
- Deroche-Gamonet V, Belin D, Piazza V. Evidence for addiction-like behavior in the rat. Science. 2004;305:1014–7. doi: 10.1126/science.1099020. [DOI] [PubMed] [Google Scholar]
- Dimitriou SG, Rice HB, Corwin RL. Effects of limited access to a fat option on food intake and body composition in female rats. Int J Eat Disord. 2000;28:436–45. doi: 10.1002/1098-108x(200012)28:4<436::aid-eat12>3.0.co;2-p. [DOI] [PubMed] [Google Scholar]
- Ebenezer IS. The effect of intracerebroventricular administration of baclofen on food intake in rats. Neuroreport. 1990;1:73–6. doi: 10.1097/00001756-199009000-00020. [DOI] [PubMed] [Google Scholar]
- Ebenezer IS. Intraperitoneal administration of baclofen increases consumption of both solid and liquid diets in rats. Eur J Pharmacol. 1995;273:183–5. doi: 10.1016/0014-2999(94)00707-e. [DOI] [PubMed] [Google Scholar]
- Ebenezer IS. Baclofen pretreatment attenuates the suppressant effect of intraperitoneal administration of cholecystokinin (CCK) on food intake in rats. Brain Res Bull. 1996;41:269–71. doi: 10.1016/s0361-9230(96)00188-8. [DOI] [PubMed] [Google Scholar]
- Ebenezer IS, Patel SM. Effects of the GABAB receptor agonists baclofen and 3-aminopropylphosphinic acid (3-APA) on food intake in rats. Methods Find Exp Clin Pharmacol. 2004;26:627–30. doi: 10.1358/mf.2004.26.8.863728. [DOI] [PubMed] [Google Scholar]
- Ebenezer IS, Pringle AK. The effect of systemic administration of baclofen on food intake in rats. Neuropharmacology. 1992;31:39–42. doi: 10.1016/0028-3908(92)90158-l. [DOI] [PubMed] [Google Scholar]
- Echo JA, Lamonte N, Ackerman TF, Bodnar RJ. Alterations in food intake elicited by GABA and opioid agonists and antagonists administered into the ventral tegmental region of rats. Physiol Behav. 2002;76:107–16. doi: 10.1016/s0031-9384(02)00690-x. [DOI] [PubMed] [Google Scholar]
- Higgs S, Barber DJ. Effects of baclofen on feeding behavior examined in the runway. Prog Neuropsychopharmacol Biol Psychiatry. 2004;28:405–8. doi: 10.1016/j.pnpbp.2003.11.003. [DOI] [PubMed] [Google Scholar]
- Hodos W. Progressive ratio as a measure of reward strength. Science. 1961;34:943–4. doi: 10.1126/science.134.3483.943. [DOI] [PubMed] [Google Scholar]
- Hodos W, Kalman G. Effects of increment size and reinforcer volume on progressive ratio performance. J Exp Anal Behav. 1963;6(3):387–92. doi: 10.1901/jeab.1963.6-387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jonaidi H, Babapour V, Denbow DM. GABAergic control of food intake in the meat-type chickens. Physiol Behav. 2002;76:465–8. doi: 10.1016/s0031-9384(02)00692-3. [DOI] [PubMed] [Google Scholar]
- Kissileff HR. Free feeding in normal and “recovered lateral” rats monitored by a pellet-detecting eatometer. Physiol Behav. 1970;5:163–73. doi: 10.1016/0031-9384(70)90060-0. [DOI] [PubMed] [Google Scholar]
- Loh EA, Roberts DCS. Break-points on a progressive ratio schedule reinforced by intravenous cocaine increase following depletion of forebrain serotonin. Psychopharmacology. 1990;101:262–6. doi: 10.1007/BF02244137. [DOI] [PubMed] [Google Scholar]
- Lucas F, Ackroff K, Sclavani A. Dietary fat-induced hyperphagia in rats as a function of fat type and physical form. Physiol Behav. 1989;45:937–46. doi: 10.1016/0031-9384(89)90218-7. [DOI] [PubMed] [Google Scholar]
- McFarland K, Kalivas PW. The circuitry mediating cocaine-induced reinstatement of drug-seeking behavior. J Neurosci. 2001;21:8655–63. doi: 10.1523/JNEUROSCI.21-21-08655.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McManus DJ, Greenshaw AJ. Differential effects of chronic antidepressants in behavioural tests of beta-adrenergic and GABAB receptor function. Psychopharmacology (Berl) 1991;103:204–8. doi: 10.1007/BF02244204. [DOI] [PubMed] [Google Scholar]
- Minano FJ, Meneres Sancho MS, Sancibrian M, Salinas P, Myers RD. GABA-A receptors in the amygdala: role in feeding in fasted and satiated rats. Brain Res. 1992;586:104–10. doi: 10.1016/0006-8993(92)91377-q. [DOI] [PubMed] [Google Scholar]
- Mobini S, Chiang TJ, Ho MY, Bradshaw CM, Szabadi E. Comparison of the effects of clozapione, haloperidol, chlorpromazine, and d-amphetamine on performance on time-constrained progressive ratio schedule on a locomotor behaviour in the rat. Psychopharmacology. 2000;152:47–54. doi: 10.1007/s002130000486. [DOI] [PubMed] [Google Scholar]
- Paterson NE, Froestl W, Markou A. The GABA-B receptor agonists baclofen and CGP44532 decreased nicotine self-administration in the rat. Psychopharmacology. 2004;72:179–86. doi: 10.1007/s00213-003-1637-1. [DOI] [PubMed] [Google Scholar]
- Ranaldi R, Poeggel K. Baclofen decreases methamphetamine self-administration in rats. Neuroreport. 2002;13:1107–10. doi: 10.1097/00001756-200207020-00007. [DOI] [PubMed] [Google Scholar]
- Reed DR, Friedman MI. Diet composition alters the acceptance of fat by rats. Appetite. 1990;14:219–30. doi: 10.1016/0195-6663(90)90089-q. [DOI] [PubMed] [Google Scholar]
- Reilly S. Reinforcement value of gustatory stimuli determined by progressive ratio performance. Pharmacol Biochem Behav. 1999;63(2):301–11. doi: 10.1016/s0091-3057(99)00009-x. [DOI] [PubMed] [Google Scholar]
- Richardson NR, Roberts DCS. Progressive ratio schedules in drug self-administration studies in rats: a method to evaluate reinforcing efficacy. J Neurosci Methods. 1996;66:1–11. doi: 10.1016/0165-0270(95)00153-0. [DOI] [PubMed] [Google Scholar]
- Roberts DCS. Breaking points on a progressive ratio schedule reinforced by intravenous apomorphine increase daily following 6-hydroxydopamine lesions of the nucleus accumbens. Pharmacol Biochem Behav. 1989;32:43–7. doi: 10.1016/0091-3057(89)90208-6. [DOI] [PubMed] [Google Scholar]
- Roberts DCS, Andrews MM, Vickers GJ. Baclofen attenuates the reinforcing effects of cocaine in rats. Neuropsychopharmacology. 1996;15(4):417–23. doi: 10.1016/0893-133X(96)00002-4. [DOI] [PubMed] [Google Scholar]
- Sclafani A, Ackroff K. Reinforcement value of sucrose measured by progressive ratio operant licking in the rat. Physiol Behav. 2003;79:663–70. doi: 10.1016/s0031-9384(03)00143-4. [DOI] [PubMed] [Google Scholar]
- Shoaib M, Swanner LS, Beyer CE, Goldberg SR, Schindler CW. The GABAB agonist baclofen modifies cocaine self-administration in rats. Behav Pharm. 1998;9:195–206. [PubMed] [Google Scholar]
- Spear DJ, Katz JL. Cocaine and food as reinforcers: effects of reinforcer, magnitude and response requirement under second-order fixed-ratio and progressive-ratio schedules. J Exp Anal Behav. 1991;56(2):261–75. doi: 10.1901/jeab.1991.56-261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stratford TR, Kelley AE. GABA in the nucleus accumbens shell participates in the central regulation of feeding behavior. J Neurosci. 1997;17:4434–40. doi: 10.1523/JNEUROSCI.17-11-04434.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stromberg MF. The effect of baclofen alone and in combination with naltrexone on ethanol consumption in the rat. Pharmacol Biochem Behav. 2004;78:743–50. doi: 10.1016/j.pbb.2004.05.006. [DOI] [PubMed] [Google Scholar]
- Thomas MA, Rice HB, Weinstock D, Corwin RL. Effects of aging on food intake and body composition in rats. Physiol Behav. 2002;76:487–500. doi: 10.1016/s0031-9384(02)00800-4. [DOI] [PubMed] [Google Scholar]
- Ward BO, Somerville EM, Clifton PG. Intraaccumbens baclofen selectively enhances feeding behavior in the rat. Physiol Behav. 2000;68:463–8. doi: 10.1016/s0031-9384(99)00197-3. [DOI] [PubMed] [Google Scholar]
- Wirtshafter D, Straford TR, Pitzer MR. Studies on the behavioral activation produced by stimulation of GABA-B receptors in the median raphe nucleus. Behav Brain Res. 1993;59:83–93. doi: 10.1016/0166-4328(93)90154-i. [DOI] [PubMed] [Google Scholar]
- Witczuk B, Khaunina RA, Kupryszewski G. 3-(p-chlorophenyl)-4-aminobuanoic acid — resolution into enantiomers and pharmacological activity. Pol J Pharmacol Phar. 1980;32(2):187–96. [Mar–Apr] [PubMed] [Google Scholar]
- Zarrindast MR, Hosseini-Nia T, Allah-Maddadi S. Food intake suppressant effect of baclofen in rats. Gen Pharmacol. 1989;20:701–3. doi: 10.1016/0306-3623(89)90110-9. [DOI] [PubMed] [Google Scholar]
- Znamensky V, Echo JA, Lamonte N, Christian G, Ragnauth A, Bodnar RJ. Gamma-aminobutyric acid receptor subtype antagonists differentially alter opioid-induced feeding in the shell region of the nucleus accumbens in rats. Brain Res. 2001;906:84–91. doi: 10.1016/s0006-8993(01)02558-6. [DOI] [PubMed] [Google Scholar]