

Abstract
Multiple older studies report that immunoglobulin directed to rough mutant bacteria, such as E. coli J5, provides broad protection against challenge with heterologous strains of Gram-negative bacteria. This protection was initially believed to occur through binding of immunoglobulin to bacterial lipopolysaccharide (LPS). However, hundreds of millions of dollars have been invested in attempting to develop clinically-effective anti-LPS monoclonal antibodies without success, and no study has shown that IgG from this antiserum binds LPS. Identification of the protective mechanism would facilitate development of broadly protective human monoclonal antibodies for treating sepsis. IgG from this antiserum binds 2 bacterial outer membrane proteins: murein lipoprotein (MLP) and peptidoglycan-associated lipoprotein (PAL). Both of these outer membrane proteins are highly conserved, have lipid domains that are anchored in the bacterial membrane, are shed from bacteria in blebs together with LPS, and activate cells through Toll-like receptor 2. Our goal in the current work was to determine if passive immunization directed to MLP and PAL protects mice from Gram-negative sepsis. Neither monoclonal nor polyclonal IgG directed to MLP or PAL conferred survival protection in 3 different models of sepsis: cecal ligation and puncture, an infected burn model, and an infected fibrin clot model mimicking peritonitis. Our results are not supportive of the hypothesis that either anti-MLP or anti-PAL IgG are the protective antibodies in the previously described anti-rough mutant bacterial antisera. These studies suggest that a different mechanism of protection is involved.
INTRODUCTION
In 1968, Chedid and colleagues reported that there was a common epitope on the outer cell membrane of Gram-negative bacteria that was exposed by serum and was bound by antibody in antisera raised to rough mutant bacteria (1). Over the next 2 decades, several investigators attempted to produce immunoglobulin directed to conserved elements in the glycolipid core of lipopolysaccharide. These studies consistently indicated that passively administered antisera raised to vaccines generated from rough mutant bacteria (such as E. coli J5) protected against challenge with heterologous organisms (1–6). Two lines of evidence suggested that the protective element was cross-reactive immunoglobulin. First, fractionation experiments revealed that the protective substance co-purified with immunoglobulin (7,8). Second, absorption experiments indicated that protective activity was removed by absorption with rough mutant bacteria (1,4,9,10). The cross-reactive epitope was assumed to be LPS, although there were no direct data supporting this view. In 1982, a landmark human trial was performed using with polyclonal human antiserum to a vaccine of E. coli J5 in which mortality in those who received the polyclonal antiserum was roughly half of controls (2).
From the start, there has been longstanding controversy regarding the existence of such cross-reactive anti-LPS antibodies. Nevertheless, based upon this concept, in the late 1980s and early 1990s, 2 companies produced monoclonal antibodies apparently directed to lipid A in the endotoxin core and studied their protective efficacy in large clinical trials. Neither of these monoclonal anti-lipid A antibodies proved to be protective (11–15). Although immunoglobulin in antiserum directed to rough mutant bacteria was hypothesized to protect by binding and neutralizing or clearing LPS, we (16) and others (15,17) were unable to find substantial increased binding of the immunoglobulin in polyclonal antiserum to E. coli J5 or the anti-lipid A monoclonal antibodies to LPS. A single monoclonal antibody has been described that reportedly binds to the inner core of LPS and neutralizes its effect (18). Thus, although this antiserum was consistently reported to protect in numerous studies, the mechanism by which this protection is accomplished is unknown.
In the late 1990s our laboratory found that IgG in antiserum to E. coli J5 bound to 2 non-LPS bacterial membrane lipoproteins (19) that were identified to be MLP and PAL (20). Subsequent work indicated that LPS is released in a complex with both MLP and PAL during sepsis (21,22), raising the possibility that the protective IgG could be anti-MLP or anti-PAL.
Both MLP and PAL are highly conserved across Gram-negative bacterial species and exert pro-inflammatory activity through signaling via Toll-like receptor 2 (TLR2) (23,24). MLP is the most abundant lipoprotein in the bacterial outer membrane, has a molecular weight of 7–9kD, and consists of 58 amino acid residues and an N-terminal lipid portion consisting of 3 fatty acids covalently linked to glycerylcysteine (25,26). Approximately two thirds of MLP exist in a free form within the outer membrane, while the remainder are covalently linked to the peptidoglycan macromolecule. MLP may aid in stabilization of the outer membrane via extension of the N-terminal lipid portion into the lipid bilayer (27,28). PAL (19 kDa) also is involved in stabilization of the outer membrane through a strong but non-covalent association with the peptidoglycan layer and interaction of N-terminal fatty acids with the outer membrane (29,30). Purified MLP is synergistic with LPS and causes lethal shock and pro-inflammatory cytokine production in both LPS-responsive and -nonresponsive mice (31). Similarly, PAL induces pro-inflammatory cytokines in vitro and in vivo in mice (22) and also is markedly synergistic with LPS in the induction of pro-inflammatory cytokines (32).
These data suggested that anti-MLP or anti-PAL IgG might protect against multiple strains of Gram-negative bacterial challenge by clearing membrane blebs containing MLP and/or PAL and LPS, by neutralizing the TLR2 agonist signal from either lipoprotein, or possibly by clearing bacteria themselves. We therefore studied the protective effects of passive immunization with antibodies to MLP and PAL in different models of Gram-negative sepsis in mice as assessed by survival, bacteremia, and inflammatory cytokine release.
MATERIALS AND METHODS
Bacteria
E. coli O18:K1:H7 (E. coli 018K+) was a gift from A. Cross (University of Maryland, Baltimore). E. coli O4:K54:H5 (CP9) was a gift from Tom Russo (SUNY Buffalo).
The E. coli K-12 mutant that lacks murein lipoprotein due to a deletion of the lpo gene, JE5505 (F− lpo his proA argE thi gal lac xyl mtl tsx) and its otherwise identical lpo-positive partner that contains MLP, JE5506 (F− pps his proA argE thi gal lac xyl mtl tsx), were provided by H. Nikaido (University of California, Berkeley) (28). E. coli K-12 1292 and JC7752 (PAL-deficient derivative of 1292) were a gift from J-C Lazzaroni (Universite Claude Bernard, Lyon, France) (33).
Purification of MLP and PAL
MLP
Total membranes were prepared from log phase cultures of E. coli O18K+. Outer membranes were prepared by density ultracentrifugation (200,000g, 4°C, 15 h) using 25% to 60% continuous sucrose density gradients (34), and pelleted from peak fractions by ultracentrifugation (100,000g, 4°C, 60 min). The resultant outer membrane pellet was suspended in 4% Triton X-100, 0.5 mM MgCl2, HEPES 10 mM, pH 7.4, incubated at room temperature for 30 min, and ultracentrifuged (100,000g, 90 min). The pellet was then suspended in 0.5% Triton X-100, 5 mM EDTA, HEPES 10 mM, pH 7.4, incubated at room temperature for 30 min and ultracentrifuged again as above. MLP is recovered in the supernatant of the second Triton X-100 extraction. Non-MLP protein contaminants were removed by size separation using continuous gel electrophoresis on a 12% SDS-PAGE tube gel as previously described (22). Peak fractions were pooled, and contaminating LPS removed by phenol extraction. MLP was pelleted by centrifugation, suspended in 50 mM NaPhos buffer, and dialyzed against 50 mM NaPhos, pH 7.4. This preparation was then used to generate mouse monoclonal and rabbit polyclonal antibodies against MLP.
PAL
PAL was purified from the outer membranes of E. coli O18K+ as previously described (20).
Antibodies
Mouse monoclonal antibodies were prepared against MLP and PAL as described elsewhere (20). Rabbit polyclonal antisera against PAL were elicited by immunizing New Zealand white rabbits with 10 μg PAL purified from E. coli O18, mixed with incomplete Freund’s adjuvant at 3-week intervals for a total of 4 doses. Rabbits were bled 3 weeks after the final dose and antisera prepared (22). Rabbit polyclonal antisera against MLP were prepared in a similar fashion by immunizing rabbits with 100 μg purified MLP from E. coli O18, mixed with incomplete Freund’s adjuvant. Rabbits received 4 doses over a 6-week period, were bled 2 weeks after the final injection, and antisera prepared from the final bleed (QED Bioscience, San Diego, CA, USA). Rabbit antisera to heat-killed E. coli J5 (J5 antisera) were prepared as described (35). Control antisera were generated by injecting rabbits with sterile saline on a schedule similar to the MLP and PAL immunizations. IgG was prepared from antisera as previously described (36,37). Briefly, affinity chromatography was performed by passage of antisera over a protein G-Sepharose 4 fast-flow column (Pharmacia, Piscataway, NJ, USA). Bound IgG was eluted from the column with 0.1 M glycine (pH 2.5). The eluate was immediately neutralized using 1 M Tris-HCl (pH 9.0). Purified IgG was dialyzed extensively against 1× PBS (pH 7.2) and stored at −80°C until use. Quantitation of IgG was determined by ELISA in a manner analogous to that described by Zollinger and Boslego (38).
Immunoblots
Immunoblotting was used to determine the specificity of rabbit anti-MLP and anti-PAL IgG. Purified MLP, PAL, boiled whole bacteria of MLP and PAL deletion mutant strains and their wild type parent strains, and E. coli strains used in the animal sepsis models were used as antigens. All samples were prepared in sample buffer (0.1 M Tris, 2.5% SDS, 11% glycerol, 0.5% β-mercaptoethanol, and trace bromophenol blue, pH 6.8), electrophoresed on a 12% SDS-polyacrylamide gel and transferred to nitrocellulose as previously described (19). Blots were then incubated in Superblock blocking buffer (Pierce Biotechnology, Rockford, IL, USA) at 4°C overnight and then washed in TTBS (150 mM NaCl, 50 mM TRIS, 0.1% Tween-20, pH 7.5). Blots were incubated with either rabbit polyclonal anti-MLP or anti-PAL at 2 μg/mL for 1 h, washed with TTBS, and incubated in HRP-conjugated anti-rabbit IgG antibody (Sigma, St. Louis, MO, USA) diluted 1:5000 in TTBS for 1 h. Blots were then developed using chemiluminescence (PerkinElmer Life Sciences, Boston, MA, USA).
Animal Models
Mice
The Institutional Animal Care and Use Committee and Massachusetts General Hospital (MGH) approved the animal studies. The mice strains used were C3H/HeN and C57BL/6 (Charles River Laboratories, Wilmington, MA, USA, and The Jackson Laboratory, Bar Harbor, ME, USA).
Cecal Ligation and Puncture (CLP)
The CLP model is a polymicrobial peritonitis model of bowel perforation that has been described by Wichterman and others (39,40). C3H/HeN mice were anesthetized with Ketamine HCl (90 mg/kg, Abbott Laboratories, Chicago, IL) and Xylazine (10 mg/kg, Sigma, St. Louis, MO, USA), given a 1-mL subcutaneous normal saline bolus, and a laparotomy was performed. The cecum was ligated with 4-0 silk and both walls were punctured with a 19-gauge needle distal to the ligature. A small amount of fecal material was expressed through each puncture site, the bowel was returned to the abdomen, and the wound was closed. Bupivacaine HCl (Astrazeneca, Wilmington, DE, USA) was placed between the fascia and skin during closure, and Buprenorphine (5 μg/kg) was administered subcutaneously for analgesia. Control (sham) mice underwent an identical operation, but without ligation or puncture of the cecum. In some experiments mice received a dose of Imipenem/Cilastin (Merck, Whitehouse Station, NJ, USA) 0.5 mg in 5% dextrose subcutaneously at 17 h postprocedure, and every 12 h for 60 h postprocedure as indicated.
Burn mouse model of Gram-negative sepsis
We utilized an infected burn model of Gram-negative sepsis in mice that was adapted from Stevens et al. (41) and was analogous to that described for rats (42). C3H/HeN mice were anesthetized with Ketamine (90 mg/kg) and Xylazine (10 mg/kg) and subjected to a 15% total body surface area full-thickness burn by application of heated brass bars (100°C, 15 s). Mice were then inoculated by subcutaneous injection of E. coli O18K+ (10–100 cfu) into the burned area. Control mice received the burn but were inoculated with saline instead of bacteria. In some experiments mice received Ceftriaxone (GlaxoSmithKline, Research Triangle Park, NC, USA) 200 μg intramuscularly 42 h postprocedure.
Peritonitis model of Gram-negative sepsis
A model of sepsis due to peritonitis was adapted from Ahrenholz and Simmons (43). Eight-week-old female C57/BL6 mice were anesthetized with ketamine (90 mg/kg) and xylazine (10 mg/kg), 0.5 mL saline bolus was given subcutaneously and Buprenorphine (5 μg/kg) for analgesia, and laparotomy was performed. A 0.5% fibrin clot containing 2.5 × 105 cfu of E. coli O4 was implanted in the peritoneum and the incision closed in 2 layers using 4.0 vicryl suture. Before closing the peritoneal layer, an additional 0.5 mL normal saline was injected into the peritoneum.
Blood Cultures
Mice were bled either by tail vein or cardiac puncture at 20 to 24 h into syringes that were prerinsed with heparin (1000 units/mL). Blood was diluted with sterile normal saline and plated on either tryptic soy agar or McConky agar plates. Plates were incubated at 37°C and colonies counted the next morning.
Cytokine Assays
Mice were bled at 24 h by cardiac puncture into syringes that were pre-rinsed with heparin (1000 units/mL). Blood was then centrifuged (12,200g, 10 min, 4°C) and the plasma collected and frozen at −80°C until analysis. TNF and IL-6 levels were measured in plasma samples using ELISA (R&D Systems, Minneapolis, MN)
Statistical Analysis
Bacteremia and cytokine levels in mice treated with polyclonal anti-MLP or anti-PAL vs. control were analyzed using the Mann-Whitney test. Survival of treated and control groups were compared using the Kaplan-Meier method and were analyzed using the log rank test. Bars on scatter graphs represent median values.
RESULTS
Passive Immunization with Monoclonal Antibodies Against MLP or PAL in the CLP Model of Sepsis
To determine if monoclonal antibodies that were known to bind MLP and PAL by immunoblotting would improve survival in an animal model of sepsis, mice were injected intravenously with monoclonal anti-MLP, anti-PAL IgG, or control IgG (0.5 mg/mouse). The mice then immediately underwent cecal ligation and puncture that resulted in the development of sepsis. There was no significant difference in survival at 72 h, nor was there a significant difference in the number of mice that developed bacteremia (Table 1).
Table 1.
Survival and bacteremia in mice immunized with monoclonal anti-MLP or monoclonal anti-PAL IgG in the CLP model of sepsis.
| Survival at 72 h | # of mice bacteremica | |
|---|---|---|
| Anti-MLP IgG | 6/17 | 5/16 |
| Control IgG | 5/18 | 4/17 |
| Anti-PAL IgG | 8/25 | 6/19 |
| Control IgG | 6/25 | 6/21 |
Sensitivity of culture ≥ 400 cfu/mL.
Passive Immunization with Polyclonal Antibodies Against MLP or PAL in a Peritonitis Model of Sepsis
Because passive immunization with monoclonal IgG did not improve survival, we next studied the ability of polyclonal IgG to protect, reasoning that IgG with multiple different specificities against released MLP or PAL would have greater protective effect. In addition, we changed the sepsis model to a monobacterial peritonitis model utilizing E. coli O4 to study the antibodies in a model more specific for Gram-negative sepsis and in which the inoculum could be more carefully controlled than with the CLP model. The specificity of the IgG purified from antisera from rabbits immunized with purified MLP or PAL was confirmed by immunoblotting against the strains of E. coli used in the sepsis models. Strong binding for each IgG to its respective antigen was detectable at 2 μg/mL, and binding was not detectable in bacteria with deletion mutants for MLP and PAL (Figure 1).
Figure 1.
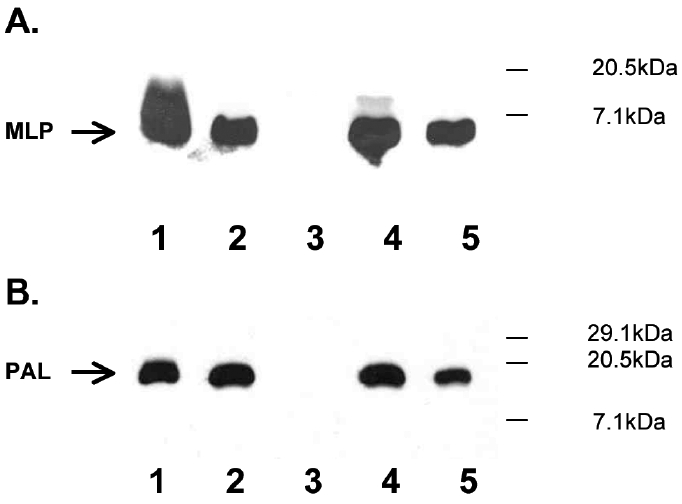
Immunoblot analysis of rabbit anti-MLP and anti-PAL. Purified MLP, PAL, boiled whole bacteria of MLP and PAL deletion mutant strains and their wild-type parent strains, and the E. coli strains used in the animal models were used as antigens. Positions of MLP and PAL bands are indicated by arrows on the left. Molecular mass markers (kDa) are at right. (A). Anti-MLP IgG. Lane 1, purified MLP (50 ng); Lane 2, JE5506 E. coli K12; Lane 3, JE5505, the corresponding E. coli K-12 MLP deletion mutant; Lane 4, E. coli O4; Lane 5, E. coli O18K+. (B). Anti–PAL IgG. Lane 1, purified PAL (50 ng); Lane 2, E. coli K-12 1292; Lane 3, JC7752, the corresponding E. coli K-12 PAL deletion mutant; Lane 4, E. coli O4; Lane 5, E. coli O18K+.
Mice were injected intravenously with polyclonal rabbit anti-MLP, anti-PAL, or control IgG (0.2 mg/mouse) and underwent laparotomy with implantation of a fibrin clot containing 2.5 × 105 cfu of E. coli O4. No significant difference in survival was seen in either the anti-MLP or anti-PAL groups compared with controls (Figure 2). Blood cultures obtained at 24 h after infection in the anti-MLP group showed no significant difference in the median cfu/mL as compared with controls. Similarly, plasma levels of TNF and IL-6 were not significantly different between immunized mice and controls (Figure 3).
Figure 2.

Protective efficacy of rabbit anti-MLP and rabbit anti-PAL in the peritonitis model of sepsis. C57BL/6 mice were injected with 200 μg of rabbit anti-MLP, rabbit anti-PAL, or control rabbit IgG and implanted with a fibrin clot containing 2.5 × 105 colony forming units cfu (cfu) of E. coli O4. n = 24 for anti-MLP and control groups and n = 16 for anti-PAL and control groups; P > 0.05, Mann-Whitney test.
Figure 3.
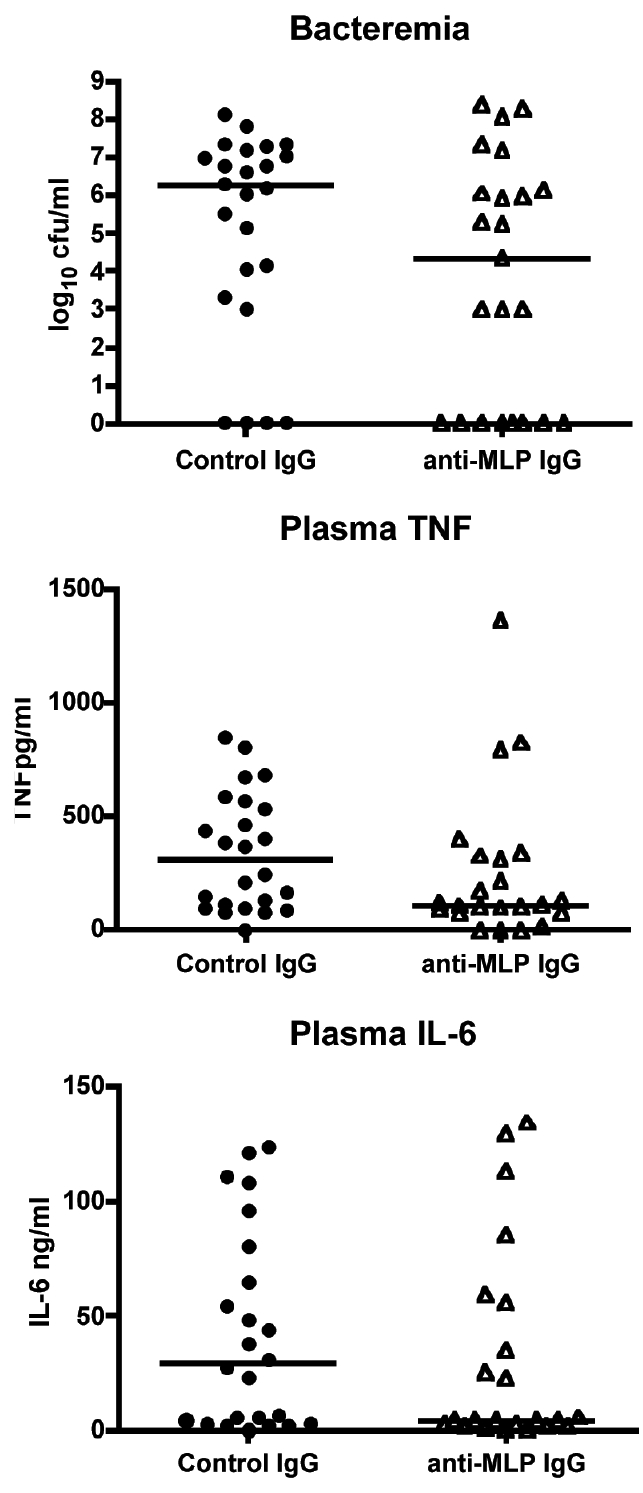
Bacteremia and TNF and IL-6 levels after passive immunization with rabbit anti-MLP in the peritonitis model of sepsis. C57BL/6 mice were injected with 200 μg of rabbit anti-MLP or control rabbit IgG and implanted with a fibrin clot containing 2.5 × 105 cfu E. coli. Mice were bled at 24 h postim-plantation and quantitative blood cultures performed and plasma TNF and IL-6 levels measured by ELISA. n = 24 for anti-MLP and control groups; P > 0.05, Mann-Whitney test.
Passive Immunization with Anti-PAL Polyclonal Antibodies in a Burn Model of Sepsis without and with Antibiotics
We also studied the protective efficacy of antibodies directed against PAL in a mouse burn model of sepsis (42). Mice were injected intravenously with rabbit anti-PAL IgG or control IgG (0.4 mg/mouse) and then underwent inoculation of a full skin thickness burn with 10–100 E. coli O18. No difference in survival was observed (Table 2). Because the rapid development of overwhelming bacteremia in the models may have masked any potential effect of passive immunization, and because antibiotics lead to lysis of bacteria with increased release of bacterial wall components, we studied the effect of antibiotics on survival in mice that had been passively immunized with polyclonal anti-PAL prior to inducing sepsis. Mice were injected intravenously with rabbit anti-PAL or control IgG (0.4 mg/mouse) and then underwent inoculation of the full skin thickness burn with E. coli O18 as above. All animals received a dose of ceftriaxone at 40 to 42 h post inoculation. Treatment of infected animals with ceftriaxone did not improve survival or decrease the development of bacteremia in either the anti-PAL immunized or control groups (Table 2). Similar results were obtained when mice were treated with imipenem in addition to polyclonal anti-PAL in the CLP model (data not shown).
Table 2.
Survival and bacteremia in mice immunized with rabbit anti-PAL IgG in the burn model of sepsis, with and without ceftriaxone
| No antibiotics | Survival at 72 h | # of mice bacteremica |
|---|---|---|
| Anti-PAL IgG | 2/9 | 2/9 |
| Control IgG | 1/9 | 2/9 |
| Ceftriaxone | ||
| Anti-PAL IgG | 5/18 | 8/18 |
| Control IgG | 6/18 | 9/18 |
Sensitivity of culture ≥ 100 cfu/mL.
DISCUSSION
The major finding of these studies is that passive immunization against MLP and PAL, 2 highly conserved TLR2 agonists that are released from the wall of Gram-negative bacteria into the bloodstream, did not confer protection against mortality in several different mouse models of sepsis. Other markers of infection and inflammation such as bacteremia and inflammatory cytokine levels also were not affected significantly by passive immunization.
Attempts to use a passive immunization strategy directed to another outer membrane component, the TLR4 agonist LPS, have been reported previously. IgG directed to the O-chain of LPS is highly protective in many models (1,6,44–46). However, it has not been possible to develop an IgG preparation that binds to LPS from most clinically relevant strains of Gram-negative bacteria (35). Indeed, antisera directed to E. coli J5 was initially developed with the hope that it would bind to common “core” epitopes that are present in most LPS molecules. While animal studies and subsequently a clinical trial with human polyclonal antisera to E. coli J5 improved survival rates in patients with suspected Gram-negative shock (2), it was difficult to find direct evidence of anti-core antibodies in binding experiments, and further studies using monoclonal antibodies directed to lipid A present in the core ultimately failed in clinical trials (11–15). Subsequent work done in our laboratory identified non-LPS outer membrane components that were bound by J5 antisera, namely outer membrane protein A, PAL, and MLP (20). Both MLP and PAL cause inflammation and contribute to the virulence of Gram-negative infections, and bacteria lacking MLP or PAL have decreased invasiveness, lethality, and ability to induce pro-inflammatory cytokine release (22,47). It therefore seemed reasonable that antibodies present in the J5 antisera directed toward these outer membrane proteins might constitute the protective factor, particularly in light of recent reports that MLP and PAL are released into the bloodstream in membrane complexes with LPS and are synergistic with LPS in the induction of inflammatory cytokines. Our studies were directed at determining if monoclonal and polyclonal antibodies against these components of the bacterial outer membrane would improve survival in models of sepsis. Unfortunately these antibodies did not have a significant impact on survival in any of the models studied.
There are several possible reasons for these negative results. First, it is possible that the models we utilized were not ideally suited to identify any therapeutic intervention for clinically relevant sepsis. Much effort has been put into developing various animal models to study the pathophysiological process of sepsis, and the models we utilized are frequently used by academic and pharmaceutical laboratories to assess protective efficacy of agents in the treatment of sepsis. However, all 3 of the models utilized were of acute sepsis, and the majority of animals that died did so within 48 h of the intervention that initiated sepsis. Addition of antibiotics did not improve outcomes in our experiments. It may be that these acute models do not accurately reflect the pathophysiology that occurs in septic patients, and that the effects of anti-MLP and anti-PAL antibodies may be different in more chronic models of sepsis. For example, it is possible that bacterial outer membrane proteins such as MLP and PAL are cleared to tissues such as the liver, lung, and kidney and induce secondary local inflammation that results in organ failure at later time points. This pathophysiology, which may be a major reason for endorgan failure and ultimate mortality in the intensive care unit, would not be detected in the models that we utilized. Second, although IgG in E. coli J5 binds predominantly to MLP and PAL in most species of Enterobacteriacae (20–21), there are likely additional outer membrane components that have pro-inflammatory activity and that could cause pathology in these acute mouse models. It is possible that the effects of these other pro-inflammatory molecules, and the potential for synergy between them, could outweigh any protective effect of a single passively administered antibody. Third, our laboratory has shown that MLP and PAL are released from bacteria into the bloodstream during sepsis as part of complexes containing LPS and perhaps other lipoproteins as well (21–22). This clustering of cell wall components may effectively shield PAL and/or MLP such that potentially protective immunoglobulins would be unable to bind to their epitopes due to steric hindrance in vivo.
Despite these possibilities, the most logical and direct conclusion of our studies is that the IgG in antiserum to E. coli J5 that is predominantly directed to bacterial wall lipoproteins MLP and PAL is not responsible for the broad protection that is described. Accordingly, the search for the protective antibody in this antiserum remains an unsolved challenge.
ACKNOWLEDGMENTS
This work was supported by NIH grants RO1-GM59694, RO1-AI058106, and 2T32AI007061-26.
Footnotes
Online address: http://www.molmed.org
REFERENCES
- 1.Chedid L, Parant M, Boyer F. A proposed mechanism for natural immunity to enterobacterial pathogens. J Immunol. 1968;100:292–301. [PubMed] [Google Scholar]
- 2.Ziegler EJ, McCutchan JA, Fierer J, Glauser MP, Sadoff JC, Douglas H, et al. Treatment of gram-negative bacteremia and shock with human antiserum to a mutant Escherichia coli. N Engl J Med. 1982;307:1225–30. doi: 10.1056/NEJM198211113072001. [DOI] [PubMed] [Google Scholar]
- 3.Braude AI, Douglas H, Davis CE. Treatment and prevention of intravascular coagulation with antiserum to endotoxin. J Infect Dis. 1973;128:S157–64. doi: 10.1093/infdis/128.supplement_1.s157. [DOI] [PubMed] [Google Scholar]
- 4.McCabe WR. Immunization with R mutants of S. minnesota. I Protection against challenge with heterologous gram-negative bacilli. J Immunol. 1972;108:601–10. [PubMed] [Google Scholar]
- 5.McCabe WR, Bruins SC, Craven DE, Johns M. Cross-reactive antigens: Their potential for Immunization-induced immunity to gram-negative bacteria. J Infect Dis. 1977;136:S161–6. doi: 10.1093/infdis/136.supplement.s161. [DOI] [PubMed] [Google Scholar]
- 6.McCabe WR, Kreger BE, Johns M. Type-specific and Cross-reactive antibodies in Gram-negative bacteremia. N Engl J Med. 1972;287:261–7. doi: 10.1056/NEJM197208102870601. [DOI] [PubMed] [Google Scholar]
- 7.McCabe WR, DeMaria A, Berberich H, Johns MA. Immunization with rough mutants of Salmonella minnesota: protective activity of IgM and IgG antibody to the R595 (Re Chemotype) mutant. J Infect Dis. 1988;158:291–300. doi: 10.1093/infdis/158.2.291. [DOI] [PubMed] [Google Scholar]
- 8.Ziegler EJ, McCutchan JA, Douglas H, Braude AI. Prevention of lethal Pseudomonas bacteremia with epimerase-deficient E. coli antiserum. Trans Assoc Am Phys. 1975;88:101–8. [PubMed] [Google Scholar]
- 9.McCabe WR. Immunization with R mutants of S. minnesota. I Protection against challenge with heterologous gram-negative bacilli. J Immunol. 1972;108:601–10. [PubMed] [Google Scholar]
- 10.Johns MA, Skehill A, McCabe WR. Immunization with rough mutants of Salmonella minnesota. IV Protection by antisera to O and rough antigens against endotoxin. J Infect Dis. 1983;147:57–67. doi: 10.1093/infdis/147.1.57. [DOI] [PubMed] [Google Scholar]
- 11.Greenman RL, Schein RMH, Martin MA, Wenzel RP, MacIntyre NR, Emmanuel G, et al. A controlled clinical trial of E5 murine monoclonal IgM antibody to endotoxin in the treatment of gram-negative sepsis. J Amer Med Assoc. 1991;266:1097–102. [PubMed] [Google Scholar]
- 12.Bone RC, Balk RA, Fein AM, Perl TM, Wenzel RP, Reines HD, et al. A second large controlled clinical study of E5, a monoclonal antibody to endotoxin: Results of a prospective, multicenter, randomized, controlled trial. Crit Care Med. 1995;23:994–1006. doi: 10.1097/00003246-199506000-00003. [DOI] [PubMed] [Google Scholar]
- 13.Ziegler EJ, Fisher CJ, Jr, Sprung CL, Straube RC, Sadoff JC, Foulke GE, et al. Treatment of gram-negative bacteremia and septic shock with HA-1A human monoclonal antibody against endotoxin: a randomized, double-blind, placebo-controlled trial. N Engl J Med. 1991;324:429–36. doi: 10.1056/NEJM199102143240701. [DOI] [PubMed] [Google Scholar]
- 14.McCloskey RV, Straube RC, Sanders C, Smith SM, Smith CR Chess Trial Study Group. Treatment of septic shock with human monoclonal antibody HA-1A. A randomized, double-blind, placebo-controlled trial. Ann Intern Med. 1994;121:1–5. doi: 10.7326/0003-4819-121-1-199407010-00001. [DOI] [PubMed] [Google Scholar]
- 15.Cross AS. Antiendotoxin antibodies: A dead end? Ann Intern Med. 1994;121:58–9. doi: 10.7326/0003-4819-121-1-199407010-00011. [DOI] [PubMed] [Google Scholar]
- 16.Warren HS, Amato SF, Fitting C, Black KM, Loiselle PM, Pasternack MS, et al. Assessment of ability of murine and human anti-lipid A monoclonal antibodies to bind and neutralize lipopolysaccharide. J Exp Med. 1993;177:89–97. doi: 10.1084/jem.177.1.89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Heumann D, Baumgartner J-D, Jacot-Guillarmod H, Glauser MP. Antibodies to core lipopolysaccharide determinants: Absence of cross-reactivity with heterologous lipopolysaccharides. J Infect Dis. 1991;163:762–8. doi: 10.1093/infdis/163.4.762. [DOI] [PubMed] [Google Scholar]
- 18.Di Padova F, Brade H, Barclay GR, Poxton IR, Liehl E, Schuetze E, et al. A broadly cross-protective monoclonal antibody binding to Escherichia coli and Salmonella lipopolysaccharides. Infect Immun. 1993;61:3863–72. doi: 10.1128/iai.61.9.3863-3872.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hellman J, Zanzot EM, Loiselle PM, Amato SF, Black KM, Ge Y, et al. Antiserum against Escherichia coli J5 contains antibodies reactive with outer membrane proteins of heterologous Gram-negative bacteria. J Infect Dis. 1997;176:1260–8. doi: 10.1086/514121. [DOI] [PubMed] [Google Scholar]
- 20.Hellman J, Loiselle PM, Tehan MM, Allaire JE, Boyle LA, Kurnick JT, et al. Outer membrane protein A, peptidoglycan-associated lipoprotein, and murein lipoprotein are released by Escherichia coli bacteria into serum. Infect Immun. 2000;68:2566–72. doi: 10.1128/iai.68.5.2566-2572.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hellman J, Loiselle PM, Zanzot EM, Tehan MM, Boyle LA, Kurnick JT, et al. Release of Gram-negative outer membrane proteins into human serum and septic rat blood and their interactions with immunoglobulin in antiserum to Escherichia coli J5. J Infect Dis. 2000;181:1034–43. doi: 10.1086/315302. [DOI] [PubMed] [Google Scholar]
- 22.Hellman J, Roberts JDJ, Tehan MM, Allaire JE, Warren HS. Bacterial peptidoglycan-associated lipoprotein is released into the bloodstream in gram-negative sepsis and causes inflammation and death in mice. J Biol Chem. 2002;277(16):14274–80. doi: 10.1074/jbc.M109696200. [DOI] [PubMed] [Google Scholar]
- 23.Aliprantis AO, Yang RB, Mark MR, Suggett S, Devaux B, Radolf JD, et al. Cell activation and apoptosis by bacterial lipoproteins through toll-like receptor-2. Science. 1999;285:736–9. doi: 10.1126/science.285.5428.736. [DOI] [PubMed] [Google Scholar]
- 24.Lien E, Sellati TJ, Yoshimura A, Flo TH, Rawadi G, Finberg RW, et al. Toll-like receptor 2 functions as a pattern recognition receptor for diverse bacterial products. J Biol Chem. 1999;274:33419–25. doi: 10.1074/jbc.274.47.33419. [DOI] [PubMed] [Google Scholar]
- 25.Braun V. Covalent lipoprotein from the outer membrane of Escherichia coli. Biochim Biophys Acta. 1975;415:335–77. doi: 10.1016/0304-4157(75)90013-1. [DOI] [PubMed] [Google Scholar]
- 26.Inouye S, Takeishi K, Lee N, DeMartini M, Hiroshima A, Inouye M. Lipoprotein from the outer membrane of Escherichia coli: purification, parachrystallization, and some properties of its free form. J Bacteriology. 1976;127:555. doi: 10.1128/jb.127.1.555-563.1976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Suzuki H, Nishimura Y, Yasuda S, Nishimura A, Yamada M, Hirota Y. Murein-lipoprotein of Escherichia coli: a protein involved in the stabilization of bacterial cell envelope. Mol Gen Genet. 1978;167:1–9. doi: 10.1007/BF00270315. [DOI] [PubMed] [Google Scholar]
- 28.Hirota N, Suzuki H, Nishimura Y, Yasuda S. On the process of cellular division in Escherichia coli: A mutant of E. coli lacking a murein-lipoprotein. Proc Natl Acad Sci. 1977;74:1417–20. doi: 10.1073/pnas.74.4.1417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Mizuno T. A novel peptidoglycan-associated lipoprotein (PAL) found in the outer membrane of Proteus mirabilis and other gram-negative bacteria. J Biochem. 1981;89:1039–49. [PubMed] [Google Scholar]
- 30.Cascales E, Bernadac A, Gavioli M, Lazzaroni JC, Lloubes R. Pal lipoprotein of Escherichia coli plays a major role in outer membrane integrity. J Bacteriol. 2002;184:754–9. doi: 10.1128/JB.184.3.754-759.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Zhang H, Peterson JW, Niesel DW, Klimpel GR. Bacterial lipoprotein and lipopolysaccharide act synergistically to induce lethal shock and proinflammatory cytokine production. J Immunol. 1997;159:4868–78. [PubMed] [Google Scholar]
- 32.Liang MD, Bagchi A, Warren HS, Tehan MM, Trigilio JA, Beasley-Topliffe LK, et al. Bacterial peptidoglycan-associated lipoprotein: a naturally occurring toll-like receptor 2 agonist that is shed into serum and has synergy with lipopolysaccharide. J Infect Dis. 2005;191:939–48. doi: 10.1086/427815. [DOI] [PubMed] [Google Scholar]
- 33.Lazzaroni J-C, Portalier R. The excC gene of Escherichia coli K-12 required for cell envelope integrity encodes the peptidoglycan-associated lipoprotein (PAL) Mol Microbiol. 1992;6:735–42. doi: 10.1111/j.1365-2958.1992.tb01523.x. [DOI] [PubMed] [Google Scholar]
- 34.Nikaido H. Isolation of Outer Membranes. Methods of Enzymology. 1994;235:225–34. doi: 10.1016/0076-6879(94)35143-0. [DOI] [PubMed] [Google Scholar]
- 35.Siber GR, Kania SA, Warren HS. Cross-reactivity of rabbit antibodies to lipopolysaccharides of Escherichia coli J5 and other gram-negative bacteria. J Infect Dis. 1985;152:954–64. doi: 10.1093/infdis/152.5.954. [DOI] [PubMed] [Google Scholar]
- 36.Warren HS, Glennon M, de Deckker FA, Tello D. Role of normal serum in the binding of lipopolysaccharide to IgG fractions from rabbit antisera to Escherichia coli J5 and other gram-negative bacteria. J Infect Dis. 1991;163:1256–66. doi: 10.1093/infdis/163.6.1256. [DOI] [PubMed] [Google Scholar]
- 37.Ge Y, Ezzell RM, Tompkins RG, Warren HS. Cellular distribution of endotoxin after injection of chemically purified lipopolysaccharide differs from that after injection of live bacteria. J Infect Dis. 1994;169:95–104. doi: 10.1093/infdis/169.1.95. [DOI] [PubMed] [Google Scholar]
- 38.Zollinger WD, Boslego JW. A general approach to standardization of the solid-phase radioimmunoassay for quantitation of class-specific antibodies. J Immunol Methods. 1981;46:129–40. doi: 10.1016/0022-1759(81)90130-7. [DOI] [PubMed] [Google Scholar]
- 39.Wichtermann K, Baus A, Chaudry I. Sepsis and septic shock–A review of laboratory models and a proposal. J Surg Res. 1980;28:189. doi: 10.1016/0022-4804(80)90037-2. [DOI] [PubMed] [Google Scholar]
- 40.Ebong S, Call D, Nemzek J, Bolgos G, Newcomb D, Remick D. Immunopathologic alterations in murine models of sepsis of increasing severity. Infect Immun. 1999;67:6603–10. doi: 10.1128/iai.67.12.6603-6610.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Stevens EJ, Ryan CM, Friedberg JS, Barnhill RL, Yarmush ML, Tompkins RG. A quantitative model of invasive Pseudomonas infection in burn injury. J Burn Care Rehab. 1994;15:232–5. doi: 10.1097/00004630-199405000-00005. [DOI] [PubMed] [Google Scholar]
- 42.Busch NA, Zanzot EM, Loiselle PM, Carter EA, Allaire JE, Yarmush ML, et al. A model of infected burn wounds using Escherichia coli O18:K1:H7 for the study of gram-negative bacteremia and sepsis. Infect Immun. 2000;68:3349–51. doi: 10.1128/iai.68.6.3349-3351.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Ahrenholz DH, Simmons RL. Fibrin in peritonitis. I Beneficial and adverse effects of fibrin in experimental E coli peritonitis. Surgery. 1980;88:41–7. [PubMed] [Google Scholar]
- 44.Tate WJ, Douglas HL, Braude AI. Protection against lethality of E. coli endotoxins with “O” antiserum. ANN NY Acad Sci. 1966;133:746–62. doi: 10.1111/j.1749-6632.1966.tb52403.x. [DOI] [PubMed] [Google Scholar]
- 45.Chen T-Y, Warren HS, Greene E, Black KM, Frostell CG, Robinson DR, et al. Protective effects of anti-O polysaccharide and anti-lipid A monoclonal antibodies on pulmonary hemodynamics. J Appl Physiol. 1993;74:423–7. doi: 10.1152/jappl.1993.74.1.423. [DOI] [PubMed] [Google Scholar]
- 46.Kim KS, Kang JH, Cross AS, Kaufman B, Zollinger W, Sadoff J. Functional activities of monoclonal antibodies to the O side chain of Escherichia coli lipopolysaccharides in vitro and in vivo. J Infect Dis. 1988;157:47–53. doi: 10.1093/infdis/157.1.47. [DOI] [PubMed] [Google Scholar]
- 47.Sha J, Fadl AA, Klimpel GR, Niesel DW, Popov VL, Chopra AK. The two murein lipoproteins of Salmonella enterica serovar Typhimurium contribute to the virulence of the organism. Infect Immun. 2004;72:3987–4003. doi: 10.1128/IAI.72.7.3987-4003.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]