

Abstract
Aims: Prostaglandins produced by the action of cyclooxygenases (COX) are important mediators of systemic vasodilatation and inflammation in liver cirrhosis. The aim of this study was to investigate the distribution of COX-1 and COX–2 in postviral cirrhosis.
Methods: The immunohistochemical expression of the constitutive (COX-1) and the inducible (COX-2) isoenzymes was investigated in 15 patients with cirrhosis after hepatitis B and C infection; three normal control livers were also analysed.
Results: COX-2 was absent from normal liver but was highly expressed in cirrhosis, mainly in the inflammatory, sinusoidal, vascular endothelial, and biliary epithelial cells. Low amounts of COX-1 were expressed in both normal and cirrhotic livers, exclusively in sinusoidal and vascular endothelial cells, with no differences seen between normal and cirrhotic livers.
Conclusions: COX-2 is overexpressed in liver cirrhosis, and possibly contributes to prostaglandin overproduction, which may be a major component of the inflammation and hyperdynamic circulation associated with cirrhosis. Because COX-2 is thought to contribute to tumour development, high COX-2 production could be a contributor to hepatocellular carcinoma development in cirrhosis. The finding of COX-2 and not COX-1 upregulation in cirrhosis could provide a possible new role for selective COX-2 inhibitors in reducing inflammation and minimising the occurrence of hepatocellular carcinoma in patients with cirrhosis.
Keywords: liver cirrhosis, viral hepatitis, portal hypertension, prostaglandins, cyclooxygenase
Cyclooxygenase (COX) is the rate limiting enzyme involved in the conversion of arachidonic acid to prostaglandin H2 (PGH2), the precursor of various compounds including PGs, prostacyclin, and thromboxanes, which are important inflammatory mediators.1 Two COX isoforms, COX-1 and COX-2, have been found to share more than 60% identity at the amino acid level. COX-1 is constitutively expressed in many tissues and responsible for various physiological functions, including cytoprotection of the stomach, vasodilatation in the kidney, and the production of a proaggregatory prostanoid, thromboxane A2, by platelets. In contrast, COX-2 is an inducible immediate early gene originally found to be induced by various stimuli such as mitogens and growth factors.2–5 Therefore, COX-2 is responsible for the release of PGs during inflammatory conditions, but COX-1 produces those PGs needed for the maintenance of normal physiological body functions. This has led to the concept that inhibition of COX-2 may explain the therapeutic usefulness of non-steroidal anti-inflammatory drugs (NSAIDs) as anti-inflammatory agents, whereas the inhibition of COX-1 may explain the unwanted renal and gastrointestinal side effects associated with their use. Overexpression of COX-2 has been demonstrated in various chronic inflammatory diseases, such as rheumatoid arthritis, Crohn’s disease, ulcerative colitis, gastritis caused by Helicobacter pylori, and chronic venous leg ulcers.6–9
“The in vivo profile of both cyclooxygenase isoforms in human liver is unknown”
The liver has emerged as the major organ participating in the degradation and elimination of arachidonic acid products of systemic origin.10 PGE2 specifically regulates important liver functions, such as portal blood pressure, glucose homeostasis, delivery of nutrients to liver parenchymal cells, and pathogenesis of liver fibrosis.11 In vitro studies have shown that primary Kupffer cells express only COX-1; however, lipopolysaccharide treated Kupffer cells express both COX-1 and COX-2.12 The in vivo profile of both COX isoforms in human liver is unknown. Therefore, our study aimed to investigate the expression of COX-1 and COX-2 in patients with liver cirrhosis using immunohistochemical staining in liver tissues to determine which isoform could be involved.
MATERIALS AND METHODS
Patients
We studied a total of 15 patients, 12 men and three women, with a mean (SD) age of 42.3 (14.3) years, from the department of tropical medicine, Minia University Hospital, Minia, Egypt. All patients were subjected to thorough clinical examination, routine laboratory investigations (blood picture, urine, and stool), liver function tests, abdominal ultrasonography, upper gastrointestinal endoscopy, sigmoidoscopy, and liver and rectal biopsies. Eleven11 patients had chronic hepatitis C virus (HCV) infection, as shown by serum HCV polymerase chain reaction (PCR) positivity, and four patients had chronic hepatitis B virus (HBV) infection, as shown by serum HBV surface antigen positivity; three patients (two with HCV and one with HBV) had coexistent schistosomiasis. The severity of the liver cirrhosis was graded clinically according to the Child-Pugh classification.13 All patients had a grade B score: this scoring depended on the presence or absence of ascites, total bilirubin, and serum albumin, in addition to the degree of encephalopathy and nutritional status of the patients. Patients received no vasoactive drugs (nitrates, β blockers, or antibiotics). None of the patients showed combined liver disease, diabetes, renal disease, arterial hypertension, congestive heart failure, or severe extrahepatic diseases
Pathological examination
We used 15 liver biopsies from patients with cirrhosis whose expression of nitric oxide synthase has been described previously.14 Informed consent was obtained from all patients whose biopsies were used in our study, after ethical approval by the local committee. Liver sections were stained with haematoxylin and eosin and pathological diagnosis was confirmed by a pathologist (RFTMcM). The presence of active inflammation, cirrhotic nodules, and fibrosis was confirmed. The Ishak necroinflammatory total score15 and the individual interface (A), lobular (C), and portal (D) components were assessed, and the Ishak fibrosis stage was confirmed as either 5 or 6. None of the cases had evidence of active schistosomiasis. Three liver biopsies with normal liver histology, from the histopathology department, Manchester Royal Infirmary, taken during cholecystectomy, were used as controls.
Immunohistochemistry
Paraffin wax embedded liver biopsies were used. Sections (5 μm thick) were cut on to poly-L-lysine coated slides. Sections were dewaxed and antigen retrieval was performed by adding 0.1% trypsin (Sigma; Poole, Dorset, UK) in calcium chloride to the section for one hour at 37°C. The slides were then processed for immunohistochemistry. Before immunolabelling, endogenous peroxidases were quenched by treatment with 0.5% H2O2 in methanol, with subsequent washing in Tris buffered saline (TBS; 0.7% Tris HCl, 0.2% Tris base, 0.02% NaCl, 1% Triton ×100). Non-specific binding of IgG was blocked using normal goat serum diluted 1/50 in 0.1% bovine serum albumin in TBS for one hour. The sections were incubated with 1/200 (COX-2) and 1/500 (COX-1) diluted primary antibodies (polyclonal rabbit anti-COX-2 and anti-COX-1 (Cayman, Ann Arbor, Michigan, USA)) at 4°C overnight, washed, and incubated for a further 60 minutes with biotinylated secondary antibodies (goat antirabbit; Vector Laboratories, Burlingame, California, USA; diluted 1/500). After incubation for a further 60 minutes with the Vectastain ABC kit, the substrate, diaminobenzidine tetrahydrochloride (Sigma), was added for 10 minutes. Positive cells were labelled brown. For the negative control, the primary antibodies were replaced with normal goat serum (the host species used to raise the secondary antibody). Haematoxylin was used as a counterstain to show the nuclei.
RESULTS
COX-2 expression was not seen in the control livers (fig 1A). The expression of COX-2 was greatly upregulated in cirrhotic liver (fig 1B–F), where it was localised in inflammatory cells infiltrating the liver (fig 1B), mainly mononuclear-like cells (fig 1D), vascular endothelial lining cells (fig 1C), Kupffer cells seen in sinusoidal spaces (fig 1E), and the epithelial lining of bile ducts (fig 1F). COX-2 was expressed only in the cytoplasmic compartment of the positive cells. It was completely absent from hepatocytes.
Figure 1 .
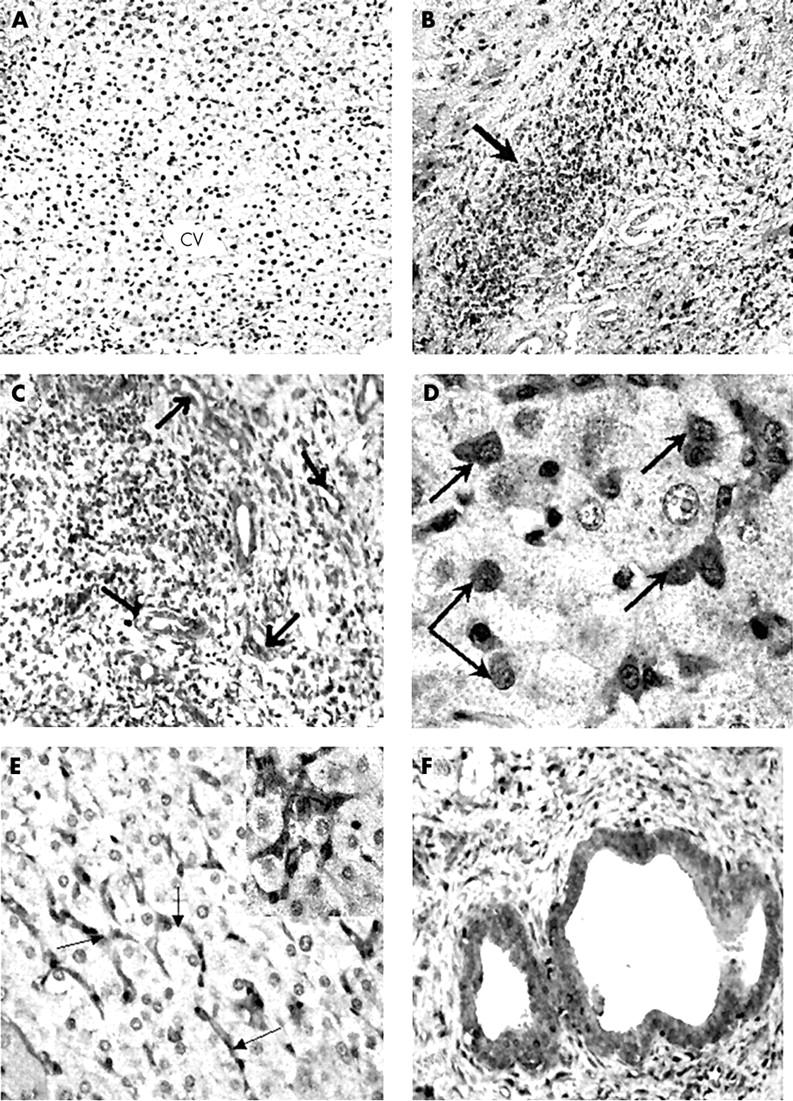
Immunoperoxidase showing COX-2 expression in (A) normal and (B–F) cirrhotic liver samples. (A) Normal human liver showing complete absence of COX-2 immunoreactivity. CV, central (terminal hepatic) vein region; original magnification, ×10. (B) A cirrhotic liver showing infiltration with inflammatory cells, which show dense COX-2 immunoreactivity (arrow); original magnification, ×25. (C) Large number of blood vessels seen in a cirrhotic liver showing COX-2 expression (arrows); original magnification, ×50. (D) Macrophage-like cells (arrows) infiltrating between hepatocytes show high COX-2 immunoreactivity; original magnification, ×100. (E) COX-2 immunoreactivity is seen in sinusoidal cells (arrows) in cirrhotic liver. The COX-2 positive sinusoidal cells are seen surrounding negatively stained hepatocytes; original magnification, ×50. (F) A liver cirrhosis sample showing COX-2 expression in the epithelial lining of bile ducts; original magnification, ×100.
COX-1 was seen in normal (fig 2A) and in cirrhotic livers (fig 2B,C). It was mainly expressed in Kupffer cells and vascular endothelial lining cells (fig 2A,B). There was no significant difference in COX-1 expression between normal and cirrhotic livers.
Figure 2 .
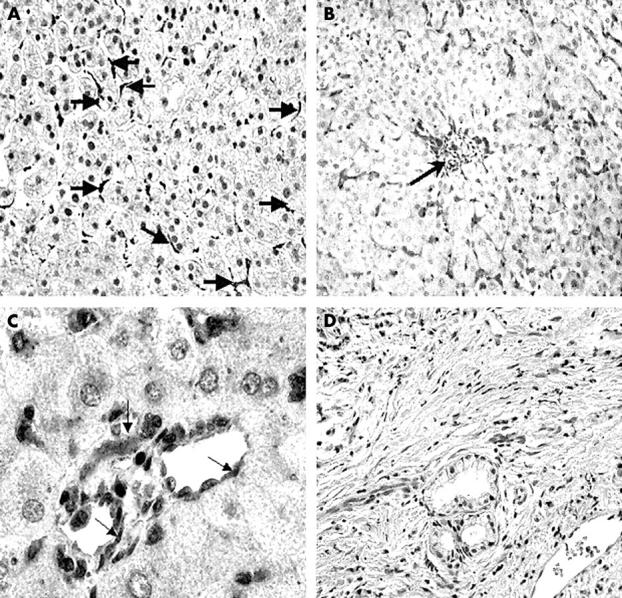
Immunoperoxidase staining showing COX-1 distribution in (A) normal liver and (B,C) human cirrhotic liver. (A) COX-1 is seen in normal liver, localised in the sinusoidal cells (arrows); original magnification, ×50. (B) In cirrhotic liver, COX-1 immunoreactivity is seen in the perivenular region (arrow) and in sinusoidal cells extending from the terminal hepatic vein; original magnification, ×25. (C) COX-1 is localised to the endothelial lining of two blood vessels seen in cirrhotic liver (arrows), with no expression in hepatocytes; original magnification, ×100. (D) A negative control in which the primary antibodies (COX-1 and COX-2) were omitted from the staining procedure, showing complete absence of staining and indicating the high specificity of the antibody used in our study; original magnification, ×50.
The negative control, in which the primary antibody was replaced by normal serum from the host species used to raise the secondary antibody, showed complete absence of staining (fig 2D), indicating the high specificity of the antibodies used.
DISCUSSION
In our study, we have shown that COX-1 is expressed both in normal and cirrhotic livers. In contrast, COX-2 was not seen in normal liver, but showed de novo synthesis and pronounced upregulation in liver cirrhosis. This induction of COX-2 may be the result of active inflammation in cirrhosis, secondary to hepatitis. Interestingly, high COX-2 expression has been reported to be highly correlated with the degree of inflammation16 and the development of fibrosis.17 This is reasonable, considering the fundamental action of COX-2 as a mediator of inflammation. In liver cirrhosis, the induction of COX-2 is probably multifactorial. The ischaemic environment in liver cirrhosis18 could be one of the inducers of COX-2 because there is evidence that ischaemia and hypoxia can induce COX-2.19 Moreover, COX-2 may be induced in the liver by growth factors.2–5 Endotoxins are also major inducers of COX-2. Dinchuk et al showed that COX-2 mediates endotoxin induced liver injury in COX-2 deficient mice.20 There is direct interaction between Kupffer cells and endotoxins that are removed from the circulation primarily by Kupffer cells, which subsequently become activated and increase prostaglandin synthesis.21–23 This may imply a role for endotoxins in the induction of COX-2 in cirrhosis.
Many of the known biological effects of PGs are mediated through their interaction with specific receptors. PGs are the key mediators of cell signalling between Kupffer cells and hepatocytes.24,25 They act on receptors on hepatocytes, increasing triglyceride synthesis and accumulation in liver. This was confirmed by the finding that COX inhibition reduces hepatic lipid accumulation.26 In a study of rat liver, Suzuki-Yamamoto et al demonstrated COX-1 staining in hepatic endothelial cells,27 whereas Yasojima et al revealed COX-1 and COX-2 expression by measuring both mRNA and protein,28 with more COX-1 than COX-2 in human livers from patients with brain diseases, including Alzheimer’s disease.
“This induction of COX-2 may be the result of active inflammation in cirrhosis, secondary to hepatitis”
There is thought to be a link between hepatitis and liver cirrhosis and the development of hepatocellular carcinoma (HCC) because liver cirrhosis is seen in up to 90% of patients with HCC.29 High COX-2 expression was found in various types of carcinoma including HCC.30,31 Kondo et al looked at the expression of COX-2 in HCC and non-tumorous tissue by immunohistochemistry using the same antibody as that used in our study.16 Expression was greatest in established cirrhosis compared with normal and non-cirrhotic liver, and was also greater than in dysplastic nodules and HCC. It was also suggested that COX-2 could play a role in the relapse of HCC. Morinaga et al have shown COX-2 overexpression in non-tumorous liver compared with HCC and demonstrated a correlation with the histological activity index, transaminase values, and proliferative activity,32 suggesting that COX-2 is related to the background necroinflammatory and regenerative activity. It has been suggested that COX is a carcinogenic agent and COX inhibitors (NSAIDs) were found to have anti-tumour activities.33,34 In an animal model, selective COX-2 inhibitors prevented carcinogenesis by the induction of apoptosis in tumour cells.35,36 Moreover, PGs have a vasodilatory action37 and COX-2 facilitates angiogenesis via the enhanced release of angiogenic growth factors, such as vascular endothelial growth factor,38 which was found to be increased in cirrhosis.39 Therefore, COX-2 may play a role in the vasodilatation and angiogenesis associated with hepatocellular disease. Thus, in liver cirrhosis, COX-2 could contribute to the pathogenesis of HCC by increasing necroinflammatory activity and promoting proliferation,32 enhancing angiogenesis,39 and inhibiting apoptosis.40–42
Take home messages
The expression of COX-2 is increased in liver cirrhosis, and possibly contributes to prostaglandin overproduction—which may be a major component of the inflammation and hyperdynamic circulation associated with cirrhosis
COX-2 is thought to contribute to tumour development, so that high COX-2 production might be important in the development of hepatocellular carcinoma (HCC) in cirrhosis
Because COX-2 but not COX-1 is upregulated in cirrhosis, selective COX-2 inhibitors might be useful in reducing inflammation and minimising the occurrence of HCC in patients with cirrhosis
In human liver cirrhosis and carbon tetrachloride (CCl4) induced liver cirrhosis in rats, there is increased renal synthesis of vasodilator PGs, which counteract the actions of endogenous vasoconstrictors such as angiotensin II, norepinephrine, and antidiuretic hormone on the renal vascular and tubular systems.43 Therefore, administration of NSAIDs in cirrhosis could induce renal failure by inhibiting renal COX and blocking PG synthesis. Interestingly, NSAIDs suppressed cirrhosis and subsequent malignant transformation in an animal model.44 However, these drugs are not recommended in patients with liver cirrhosis because of the renal side effects. This limitation could be overcome by the recent findings concerning selective COX-2 inhibitors and their possible use in some human diseases.2 Recently, it has been shown that selective COX-2 inhibitors did not impair renal function in a rat model of liver cirrhosis.45 This suggests that effective treatment by selective COX-2 inhibitors may be possible and confirms that the maintenance of renal function is attributed mainly to PGs derived from COX-1.45 Therefore, the fact that the main source of COX in liver cirrhosis is COX-2, with little contribution from COX-1, suggests that the use of selective COX-2 inhibitors in these patients may help to reduce inflammation and provide a potential preventive measure for malignant transformation.
Supplementary Material
Acknowledgments
We thank Minia University for financial support.
Abbreviations
COX, cyclooxygenases
HCC, hepatocellular carcinoma
NSAID, non-steroidal anti-inflammatory drug
PG, prostaglandin
TBS, Tris buffered saline
This study was presented to the British Society of Gastroenterology at Birmingham in March 2002 and has been published in abstract form (Gut 2002;50suppl II:A31).
Sadly, Dr N A Mohammed has died since this paper was written.
REFERENCES
- 1.Appleton I, Tomlinson A, Willoughby D A. Induction of cyclooxygenase and nitric oxide synthase in inflammation. Adv Pharmacol 1996;35:27–78. [DOI] [PubMed] [Google Scholar]
- 2.Willoughby DA, Tomlinson A. Inducible enzymes in inflammatory response. In: Willoughby DA, Tomlinson A, eds. Progress in inflammation research. Basel, Boston, Berlin: Birkhäuser, 1999:1–30.
- 3.Jones DA, Carlton DP, Mcintyre TM, et al. Molecular cloning of human prostaglandin endoperoxide synthase type II and demonstration of expression in response to cytokines. J Biol Chem 1993;268:9049–54. [PubMed] [Google Scholar]
- 4.Hamasaki Y, Kitzler J, Hardman R, et al. Phorbol ester and epidermal growth factor enhance the expression of two inducible prostaglandin H synthase genes in rat tracheal epithelial cells. Arch Biochem Biophys 1993;304:226–34. [DOI] [PubMed] [Google Scholar]
- 5.DuBois RN, Awad J, Morrow J, et al. Regulation of eicosanoid production and mitogenesis in rat intestinal epithelial cells by transforming growth factor alpha and phorbol ester. J Clin Invest 1994;93:493–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kang RY, Freire–Moar J, Sigal E, et al. Expression of cyclooxygenase-2 in human and an animal model of rheumatoid arthritis. Br J Rheumatol 1996;35:711–18. [DOI] [PubMed] [Google Scholar]
- 7.Singer II, Kawka DW, Schloemann S, et al. Cyclooxygenase-2 is induced in colonic epithelial cells in inflammatory bowel disease. Gastroenterology 1998;115:297–306. [DOI] [PubMed] [Google Scholar]
- 8.Sawaoka H, Kawano S, Tsuji S, et al. Cyclooxygenase-2 inhibitors suppress the growth of gastric cancer xenografts via induction of apoptosis in nude mice. Am J Physiol 1998;274:G1061–7. [DOI] [PubMed] [Google Scholar]
- 9.Abd-El-Aleem SA, Ferguson MWJ, Appleton I, et al. Expression of cyclooxygenase isoforms in normal human skin and chronic venous ulcers. J Pathol 2001;195:616–23. [DOI] [PubMed] [Google Scholar]
- 10.Quiroga J, Prieto J. Liver cytoprotection by prostaglandins. Pharmacol Ther 1993;58:67–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Wisse E, Knook DL, Fraser R. Cells of hepatic sinusoids, Vol. 7. Leiden, The Netherlands: The Kupffer Cell Foundation, 1999.
- 12.Dieter P, Scheibe R, Jakobsson P, et al. Functional coupling of cyclooxygenase 1 and 2 to discrete prostanoid synthases in liver macrophages. Biochem Biophys Res Commun 2000;276:488–92. [DOI] [PubMed] [Google Scholar]
- 13.Infante-Rivard C, Esnaola S, Villeneuve JP. Clinical and statistical validity of conventional prognostic factors in predicting short term survival among cirrhotics. Hepatology 1987;7:660–4. [DOI] [PubMed] [Google Scholar]
- 14.Mohammed NA, Abd El-Aleem S, Appleton I, et al. Expression of nitric oxide synthase isoforms in human liver cirrhosis. J Pathol 2003;200:647–55. [DOI] [PubMed] [Google Scholar]
- 15.Ishak K, Baptista A, Bianchi L, et al. Histological grading and staging of chronic hepatitis. J Hepatol 1995;22:696–9. [DOI] [PubMed] [Google Scholar]
- 16.Kondo M, Yamamoto H, Nagano H, et al. Increased expression of COX-2 in nontumour liver tissue is associated with shorter disease-free survival in patients with hepatocellular carcinoma. Clin Cancer Res 1999;5:4005–12. [PubMed] [Google Scholar]
- 17.Cheng J, Imanishi H, Iijima H, et al. Expression of cyclooxygenase 2 and cytosolic phospholipase A2 in the liver tissue of patients with chronic hepatitis and liver cirrhosis. Hepatol Res 2002;23:185–95. [DOI] [PubMed] [Google Scholar]
- 18.Morales-Ruiz M, Jimenez W, Perez-Sala D, et al. Increased nitric oxide synthase expression in arterial vessels of cirrhotic rats with ascites. Hepatology 1996;24:1481–6. [DOI] [PubMed] [Google Scholar]
- 19.Schmedtje JF Jr, Ji YS, Liu WL, et al. Hypoxia induced cyclooxygenase-2 via the NF-kappa B p65 transcription factors in human vascular endothelial cells. J Biol Chem 1997;272:601–8. [DOI] [PubMed] [Google Scholar]
- 20.Dinchuk JE, Car BD, Focht RJ, et al. Renal abnormalities and an altered inflammatory response in mice lacking cyclooxygenase 2. Nature 1995;378:406–9. [DOI] [PubMed] [Google Scholar]
- 21.Puschel GP, Kirchner C, Schroder A, et al. Glycogenolytic and antiglycogenolytic prostaglandin E2 actions in rat hepatocytes are mediated via different signalling pathways. Eur J Biochem 1993;218:1083–9. [DOI] [PubMed] [Google Scholar]
- 22.Qu W, Savier E, Thurman RG. Stimulation of monooxygenation and conjugation following liver transplantation in the rat: involvement of Kupffer cells. Mol Pharmacol 1992;41:1149–54. [PubMed] [Google Scholar]
- 23.Qu W, Zhong Z, Goto M, et al. Kupffer cell prostaglandin E2 stimulates parenchymal cell O2 consumption: alcohol and cell–cell communication. Am J Physiol 1996;270:G574–80. [DOI] [PubMed] [Google Scholar]
- 24.Coleman RA, Kennedy I, Humphrey PP, et al. Prostanoids and their receptors. In: Hansch C, Sammes PG, Taylor JB, et al, eds. Comprehensive medicinal chemistry. Oxford: Pergamon, 1989:643–714.
- 25.Funk C, Furci L, Fitzgerald GA, et al. Cloning and expression of a cDNA for human prostaglandin E receptor EP1 subtype. J Biol Chem 1993;268:26767–72. [PubMed] [Google Scholar]
- 26.Enomoto N, Ikejima K, Yamashina S, et al. Kupffer cell-derived prostaglandin E2 is involved in alcohol-induced fat accumulation in rat liver. Am J Physiol 2000;279:G100–6. [DOI] [PubMed] [Google Scholar]
- 27.Suzuki-Yamamoto T, Yokoi H, Tsuroro Y, et al. Identification of prostaglandin F-producing cells in the liver. Histochem Cell Biol 1999;112:451–6. [DOI] [PubMed] [Google Scholar]
- 28.Yasojima K, Schwab C, McGeer EG, et al. Distribution of cyclooxygenase-1 and cyclooxygenase-2 mRNAs and proteins in human brains and peripheral organs. Brain Research 1999;830:226–36. [DOI] [PubMed] [Google Scholar]
- 29.Okuda K. Hepatocellular carcinoma: recent progress. Hepatology 1992;15:948–63. [DOI] [PubMed] [Google Scholar]
- 30.Koga H, Sakisaka S, Ohishi M, et al. Expression of cyclooxygenase-2 in human hepatocellular carcinoma: relevance to tumor dedifferentiation. Hepatology 1999;26:688–96. [DOI] [PubMed] [Google Scholar]
- 31.Shiota G, Okubo M, Noumi T, et al. Cyclooxygenase-2 expression in hepatocellular carcinoma. Hepatogastroenterology 1999;46:407–12. [PubMed] [Google Scholar]
- 32.Morinaga S, Yamamoto Y, Noguchi Y, et al. Cyclooxygenase-2 mRNA is up-regulated in cirrhotic or chronic hepatitis liver adjacent to hepatocellular carcinoma. J Gastroenterol Hepatol 2002;17:1110–16. [DOI] [PubMed] [Google Scholar]
- 33.Oshima M, Dinchuk JE, Kargman SL, et al. Suppression of intestinal polyposis in Apc delta716 knockout mice by inhibition of cyclooxygenase 2 (COX-2). Cell 1996;87:803–9. [DOI] [PubMed] [Google Scholar]
- 34.Taketo MM. Cyclooxygenase-2 inhibitors in tumorigenesis (part II). J Natl Cancer Inst 1998;90:1609–20. [DOI] [PubMed] [Google Scholar]
- 35.Vane J. Suppression of intestinal polyposis by inhibition of COX-2 in Apc knockout mice. Jpn J Cancer Res 1997;88:1244–5. [PubMed] [Google Scholar]
- 36.Okami J, Yamamoto H, Fujiwara Y, et al. Overexpression of cyclooxygenase-2 in carcinoma of the pancreas. Clin Cancer Res 1999;5:2018–24. [PubMed] [Google Scholar]
- 37.Groszmann RJ. Hyperdynamic state in chronic liver diseases. J Hepatol 1993;17 (suppl 2):S38–40. [DOI] [PubMed] [Google Scholar]
- 38.Tsujii M, Kawano S, Tsuji S, et al. Cyclooxygenase regulates angiogenesis induced by colon cancer cells. Cell 1998;93:705–16. [DOI] [PubMed] [Google Scholar]
- 39.El-Assal ON, Yamanoi A, Yamaguchi M, et al. Clinical significance of microvessel density and vascular endothelial growth factor expression in hepatocellular carcinoma and surrounding liver: possible involvement of vascular endothelial growth factors in angiogenesis of cirrhotic liver. Hepatology 1998;27:1554–62. [DOI] [PubMed] [Google Scholar]
- 40.Hara A, Yoshimi N, Niwa M, et al. Apoptosis induced by NS-398, a selective cyclooxygenase-2 inhibitor, in human colorectal cancer cell lines. Jpn J Cancer Res 1997;88:600–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Liu X, Yao S, Kirschenbaum A, et al. NS398, a selective cyclooxygenase-2 inhibitor, induces apoptosis and downregulates bcl-2 expression in LNCaP cells. Cancer Res 1998;58:4245–9. [PubMed] [Google Scholar]
- 42.Denda A, Endoh T, Kitayama W, et al. Inhibition by pyroxicam of oxidative damage, liver cirrhosis and development of enzyme-altered nodules caused by a choline-deficient, L-amino acid-defined diet in rats. Carcinogenesis 1997;18:1921–30. [DOI] [PubMed] [Google Scholar]
- 43.Sola J, Campos J, Arroyo V, et al. Longitudinal study of renal prostaglandin excretion in cirrhotic rats: relationship with the renin–aldosterone system. Clin Sci 1988;75:263–9. [DOI] [PubMed] [Google Scholar]
- 44.Arroyo V, Gines P, Rimola A, et al. Renal function abnormalities, prostaglandins, and effects of nonsteroidal anti-inflammatory drugs in cirrhosis with ascites. An overview with emphasis on pathogenesis. Am J Med 1986;81 (2B):104–22. [DOI] [PubMed] [Google Scholar]
- 45.Bosch-Marce M, Claria J, Titos E, et al. Selective inhibition of cyclooxygenase 2 spares renal function and prostaglandin synthesis in cirrhotic rats with ascites. Gastroenterology 1999;116:1167–75. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.