

Abstract
Among flowering plants, females often have little control over the genetic relatedness of pollen deposited on stigmas. Thus, postpollination processes are of primary importance for mate discrimination. The ability to screen and select among male gametes during pollen tube growth within the female tissues of the stigma, style, and ovary is critical to the process of mate choice and reproductive isolation. However, direct evidence of the mechanistic/developmental processes associated with mate choice in flowering plants is sparse. We studied the contribution of postpollination, prefertilization processes to reproductive isolation in two wind-pollinated species of birch (Betula) that commonly hybridize in nature. We exploited the 3-fold difference in ploidy level between these two species to determine the paternity of individual pollen tubes growing within female reproductive tissues. We then tracked their developmental fate in conspecific, heterospecific, and mixed-species crosses. This design allowed a direct comparison of developmental processes responsible for reproductive isolation in both single-species and mixed-species pollinations. Our results have important implications for the population genetic outcome of hybridization dynamics in natural populations. Paternity analysis of progeny from mixed-species pollinations revealed that conspecific pollen tubes sired more than 98% of seedlings. Biased siring success was not the result of differential embryo abortion. We detected strong, early postpollination barriers such as pollen tube incompatibility, slower pollen tube growth, and delayed generative cell mitosis. Conspecific fertilization precedence was mediated by favorable or unfavorable male-female interactions, but there was no evidence for antagonistic male-male interactions.
To distinguish among hypotheses for the origin and maintenance of reproductive isolation, research has focused on traits that contribute to present-day isolation between species. Polyploidy is one mechanism often cited as leading to “instantaneous speciation,” because the F1 hybrids of crosses between taxa of different ploidy levels usually have low fertility (1). Recent molecular and cytogenetic studies have shown that gene flow between diploids and polyploids can be extensive and an important evolutionary force long after genome duplication and divergence have taken place (2). Thus, it is important to understand reproductive processes that constrain gene flow across diploid-polyploid species boundaries. Such processes are likely to share many features with diploid-diploid hybridization, but they also have some unique attributes (3, 4).
Population genetic studies can determine the ultimate consequences of assortative mating at many taxonomic levels, but only developmental studies can examine the underlying mechanisms that contribute to such patterns in nature. Nevertheless, studies of mate discrimination (other than self-incompatibility) have long been hampered by an inability to assign fitnesses to genotypes of individual male and female gametes and zygotes in situ. In plants, differences in rates of development of gamete-bearing structures (gametophytes) and/or zygotes are thought to be critical to mate sorting (5, 6). A definitive study of mate recognition first would localize the stage(s) at which sorting occurs, and then, within each stage, evaluate the relative roles of male-male and male-female interactions. Here we analyze early postpollination barriers between two species of birch. Our results are unique in describing a continuous series of developmental stages that lead to reproductive isolation. These findings are the product of an experimental protocol we devised for definitive identification of the paternity of pollen tubes and zygotes during the entire reproductive process.
Betula occidentalis Hooker (2n = 2x = 28), water birch, frequently hybridizes with B. papyrifera Marsh. (2n = 6x = 84), paper birch, over a broad zone of sympatry in the northern Great Plains and Rocky Mountains of North America (7). These species are wind-pollinated and flower synchronously and thus possess no premating recognition mechanisms. The genus is known to possess gametophytic self-incompatibility, a prefertilization recognition mechanism that also has been implicated in heterospecific incompatibility (8, 9). Based on chromosome numbers, nuclei of paper birch are predicted to have approximately three times the amount of DNA as equivalent nuclei in water birch (10). We exploited this property as a marker of paternity, by directly measuring and comparing DNA content of male gamete nuclei in pollen tubes growing within the female reproductive tissues in mixed-species pollinations. This study directly visualizes interactions among pollen tubes of known paternity within the plant reproductive tract or style.
METHODS
Experimental Pollinations.
Experimental crosses were performed on trees from two sites in the Black Hills of South Dakota (paper birch site = Hell Canyon, SD, 43° 44′ N, 103° 55′ W; water birch site = Sunday Canyon, SD, 43° 51′ N, 103° 35′ W). Crosses were performed each day from May 28 to June 3, 1997. Male inflorescences were collected from 15–18 trees (pollen donors) of each species and placed in water until pollen release. Branches from 10 water birch maternal trees and 9 paper birch maternal trees were emasculated and bagged until female flowers became receptive. All inflorescences within each pollination bag were pollinated with one of three pollen mixtures (two replicates/tree). Control, or “single-species,” treatments consisted of a mixture of equal volumes of fresh pollen from at least 12 individuals per species. A pollen competition, or “mixed-species,” treatment consisted of a 50:50 mixture, by volume, of each of the two single-species mixtures. On average, the volume of a paper birch pollen grain is 1.5 times greater than the volume of a water birch pollen grain (8.31 × 103 μm3 vs. 5.56 × 103 μm3, respectively). Thus, the mixed-species treatment consisted of 60% water birch and 40% paper birch pollen grains. Pollen viability of each mixture was assessed 1 week after the last pollination and was > 72% in all three cases (cotton blue method; ref. 11). Pollen was applied by blowing forcefully across a paintbrush dipped in pollen. The method was practiced for consistency beforehand [mean (SD) number of water birch pollen grains per stigma = 16(7), n = 51; paper birch = 22(12), n = 15]. Thus we mimicked both wind pollination and pollen loads likely to be encountered in nature (12).
To examine early pollen tube development, flowers from one inflorescence from each treatment were collected and fixed 45–51 hr after pollination. Flowers were fixed for 24 hr in 3:1 (V1/V2) ethanol-acetic acid and transferred to 75% ethanol for storage at 4°C. Flowers then were dehydrated through an ethanol series, and infiltrated and embedded with glycol methacrylate (JB-4 embedding kit; Polysciences). Embedded flowers were serially sectioned into 5-μm thick ribbons and mounted on microscope slides. Slides were flooded with a solution of 0.25 μg/ml of 4′,6-diamidino-2-phenylindole (DAPI) and 0.1 mg/ml p-phenylenediamine (added to reduce fading) (13) in 0.05 M Tris (pH 7.2) for 1 hr at room temperature in a light-free environment. Microspectrofluorometric measurements of relative DNA levels of DAPI-stained nuclei were performed within 2 hr (for detailed methods, see ref. 14). Such measurements were made with a Zeiss MSP 20 microspectrophotometer with digital microprocessor coupled to a Zeiss Axioskop microscope equipped with epifluorescence (HBO 50 W burner). A UV filter set (model no. 48702) with excitation filter (365 nm, band pass 12 nm), dichroic mirror (FT395), and barrier filter (LP397) was used with a Zeiss Plan Neofluar 40× objective. Additional flowers were collected near the time of fertilization and stained with 0.1% aniline blue in 0.1 N K3PO4 for 5 min, rinsed, and then stained with DAPI, as above.
Pollen Tube Development.
All serial sections containing the two styles of a flower were examined, and each pollen tube was scored for viability, position in the style, and developmental condition of the generative cell. A viable pollen tube contained brightly stained nuclei and cytoplasm and had penetrated the stigmatic surface. Inviable pollen tubes were either burst at the tip, had degenerate nuclei, or were trapped in necrotic stylar tissue. Because variation in pollen quantity and quality can confound conclusions about pollination treatments, we summed counts/style within trees for all analyses described below (15). To estimate differences in pollen tube viability, we used replicated G tests (16) to test for deviations from extrinsic hypotheses based on the relative amount of pollen applied in each treatment. In some comparisons we assumed pollen germination rate to be unaffected by the reduced pollen load size of each donor species in the mixed-species treatment. This expectation is reasonable, because the total pollen load size was held constant for each treatment and given the low numbers and the dispersed spatial distribution of pollen grains observed on stigmas (11).
As a measure of the relative growth rates of pollen tubes, we analyzed the proportion of viable pollen tubes that had reached the bottom fifth of the style by using the Mantel-Haenzsel procedure (16). This procedure tests for homogeneity of log-odds ratios between separate 2×2 tables and is analogous to testing for an interaction effect in a three-way contingency table. If ratios were heterogeneous, we analyzed 2×2 tables separately.
A major regulatory control point in the plant mitotic cell cycle is the G2 to mitosis transition (17). As a measure of the G2-mitosis transition, we scored generative cell nuclei as either premitotic (chromosomes not condensed) or mitotic/postmitotic (prophase of mitosis or later). We used the Mantel-Haenszel procedure to analyze the proportion of generative cell nuclei in mitosis or beyond. For multiple tests within each of the above analyses, P values were adjusted by using the sequential Bonferroni method (18).
Paternity Analysis.
Seeds were collected in late summer of 1997, stored at 4°C (ca. 6% moisture content) for 9 months, and then germinated in a growth chamber (22°C with 12 hr light/day). Seeds were scored for germination 5 weeks after planting. To determine paternity of the progeny of mixed-species crosses, a random sample of seedlings from the mixed-species crosses was subjected to electrophoretic analysis, along with leaves from their mothers and all pollen donors. Because there were no fixed electrophoretic markers that differentiated the paternal species, we performed a multilocus paternity analysis on the progeny. Based on a larger survey, we chose eight enzyme systems (10 loci) that were either highly polymorphic among individuals of both species or had pronounced allele frequency differences between species. These systems were Aat-4, Fe-3, 6-Pgdh-1, 6-Pgdh-2, Per-5, Pgi-2, Pgm, Tpi-2, Ugpp-1, and Ugpp-2.
We performed paternity analysis by using a simple exclusion approach (19): when the paternal allele(s) at a locus could be inferred from the observed progeny and maternal genotypes, then all fathers that lacked the allele(s) were excluded. This process was repeated for each locus until all fathers of one species could be excluded. Fewer than 1% of the progeny were ambiguous. We assigned paternity to these progeny by using the fractional paternity method (20), summing the fractional paternities of all potential fathers within each species.
RESULTS
Development.
The birch flower has two styles, each with a broad stigmatic surface along its length (Fig. 1). At pollination, the ovary is minimally developed and normally contains one ovule primordium per style (Fig. 1). Pollen tube development occurs in two phases. Phase one includes rapid germination, penetration of the stigma, mitosis of the generative cell during or soon after penetration, and rapid pollen tube growth within the transmitting tract to the base of the style. Partial necrosis of stylar/stigmatic cells was observed in most flowers 2 days after pollination. Necrosis proceeded from the tip toward the base, and thus imposed a constraint on pollen tubes to undergo rapid development. At the base of the style, pollen tubes formed a cyst-like structure containing the tube cell nucleus and two sperm cells. Pollen tube “cysts” remained dormant for up to 4 weeks, until the ovary matured and the female gametophyte initiated development. During phase 2, viable pollen tube “cysts” regerminated and grew into the ovary, but ovules were rarely approached by more than one pollen tube.
Figure 1.

Cross-section of birch flower at anthesis. Pollination occurs above the sheathing bract [note pollen grains (Pg) in this region]. Bst, base of style; N, necrotic tissue; Op, ovule primordium; St, stigma/style; Tt, transmitting tract. (Bar = 50 μm.)
As a control for comparisons between conspecific and heterospecific phase 1 pollen tube development, we compared pollen tubes in conspecific styles in the single-species crosses. There were no significant differences between species in the number of viable pollen tubes/conspecific style [mean (SD) = 9.48 (11.67) vs. 12.03 (11.04), for water and paper birch, respectively; P > 0.37, by two-tailed randomization test]; the proportion of viable conspecific pollen tubes/style that had reached the base of the style (mean proportion = 0.098 vs. 0.087 for water and paper birch, respectively; G = 2.15; P > 0.15), or the proportion of generative cell nuclei that had initiated mitosis (mean proportion = 0.71 vs. 0.74 for water and paper birch, respectively; G = 0.66; P > 0.44). Thus, early male gametophyte and gamete development is similar in both species.
DNA Content.
To determine the paternity of each pollen tube, we measured the relative DNA content of the generative cell nucleus or of one sperm cell nucleus within each pollen tube. Relative fluorescence of conspecific 4′,6-diamidino-2-phenylindole (DAPI)-stained generative cell and sperm cell nuclei was used to standardize C values of all other nuclei (Fig. 2). The relative fluorescence of (i) pollen tube generative cell nuclei during prophase and metaphase and (ii) sperm cell nuclei during telophase of inception should reflect the 2C and the 1C DNA content, respectively, because these nuclei contain the haploid chromosome number in these stages of development. Within each species, DNA content of generative and sperm cell nuclei conformed to the predicted 2:1 ratio (Fig. 2).
Figure 2.

DNA content of haploid nuclei. (Inset) Two sperm nuclei at late telophase of inception. A net photometric value for each section of a nucleus was determined by taking an initial reading of one sperm nucleus, and then subtracting background fluorescence of nearby cytoplasm and embedding medium (B). Relative nuclear DNA content was determined by summation of individual fluorescence values of serial sections through the nucleus, less background fluorescence. Filled columns represent sperm nuclei (Sn). Empty columns represent generative cell nuclei (Gn). Error bars represent 1 SD. Pt, pollen tube; Tn, tube nucleus. (Bar = 10 μm.)
The 1C DNA content of water birch is approximately 50 relative fluorescence units (RFUs), whereas the 1C DNA content of paper birch is approximately 145 RFU. Thus, the 1C DNA value of paper birch (6x = 84) is close to three times the 1C DNA value of water birch (2x = 28; Fig. 2). The relative fluorescence values of paper birch 1C sperm nuclei were virtually nonoverlapping with the relative fluorescence values of water birch 2C generative cell nuclei (P < 0.0001; Fig. 2). Thus, regardless of position in the cell cycle, the paternity of each pollen tube could be reliably assessed in the mixed-pollination styles.
Pollen Tube Viability.
On both maternal species, highly significant pooled G tests indicated that conspecific pollen tube viability was higher than heterospecific pollen tube viability in all cases, although two paper birch females were heterogeneous in the magnitude of departure from expectation (Fig. 3; and see below). We observed two major causes of heterospecific pollen tube inviability: delayed pollen germination and/or pollen tube bursting. On water birch stigmas, paper birch pollen germinated and penetrated the stigma normally, but the pollen tubes often burst within the upper part of the style. Most paper birch trees displayed a strikingly different pattern. On every stigma of seven paper birch trees, heterospecific pollen tubes either failed to exit the pollen grain or were burst at the surface of the stigma. Stigmas on the remaining two paper birch trees displayed the same pattern as water birch mothers, with the only reduction in the heterospecific cohort occurring in the upper part of the style. Henceforth, we refer to paper birch trees as either “heteroincompatible” (HI), indicating trees that possess a strong early viability barrier at the maternal genotype level; or “heterocompatible” (HC), indicating trees that possess only weak early viability barriers.
Figure 3.

The mean number of viable pollen tubes observed per style per tree 48 hr after pollination in (A) water birch (n = 10) and (B) HC (n = 2) and HI (n = 7) paper birch. Inset numbers refer to the relative amount of pollen applied in single-species (pure conspecific or pure heterospecific) or mixed-species (conspecific and heterospecific mixture) crosses. Asterisks indicate significance levels of ratio tests against the expected ratio of conspecific/heterospecific pollen applied in each treatment. ∗, <0.05; ∗∗∗∗, <0.0001.
We tested for interspecific paternal interactions by comparing pollen tube viability in single- vs. mixed-species crosses. On water birch trees the ratio of conspecific/conspecific, or heterospecific/heterospecific, viable pollen tubes in single- vs. mixed-species crosses did not differ significantly from expected (Fig. 3A; P > 0.15 in both cases). However, the seven HI paper birch trees had significantly higher heterospecific pollen tube viability in the mixed-species than in the single-species crosses (Fig. 3B; GPooled = 20.44, P < 0.0001). The two HC paper birch had significantly higher conspecific pollen tube viability in mixed- vs. single-species crosses (Fig. 3B; GPooled = 27.34, P < 0.0001). Other comparisons were nonsignificant (P > 0.71). Thus, HI paper birch styles accepted a low amount of heterospecific pollen in the presence of conspecific pollen, but there was no evidence for antagonistic male-male interactions in water or paper birch styles.
Pollen Tube Growth.
A measure of both the vigor of pollen tubes and their ability to interact favorably with maternal tissue is the proportion of each cohort that reached the base of the style 48 hr after pollination. On water birch mothers, the proportion of viable conspecific pollen tubes that had reached the base of the style did not differ significantly from that of heterospecific pollen tubes (mean = 0.090 vs. 0.120 for paper vs. water birch, respectively; χ2MH = 2.33, P > 0.13). This result did not depend on whether pollen tubes grew alone or in competition (χ2H = 1.39, P > 0.24). Thus, in water birch styles, heterospecific pollen tubes that survived early viability barriers were equally likely to grow to the base of the style as conspecific pollen tubes. In addition, there was no evidence for interspecific male-male interactions.
On HI paper birch mothers, no heterospecific pollen tubes reached the base of the style. On the two HC mothers, few heterospecific pollen tubes reached the base of the style (mean proportion = 0.034 vs. 0.123 for water vs. paper birch, respectively; χ2MH = 5.77, P < 0.10). This result did not depend on whether pollen tubes grew alone or in competition (χ2H = 1.80, P > 0.19). Thus, in most paper birch styles, heterospecific pollen tubes that survived the viability barrier did poorly in terms of growth, regardless of whether pollen tubes grew alone or in competition.
Gamete Development.
We next sought to determine whether there are more subtle indicators of fitness than viability and growth of the male gametophyte. In water birch styles, conspecific generative cell nuclei were 2.9 times more likely than heterospecific generative cell nuclei to have initiated mitosis (Table 1; χ2MH = 51.77, P < 0.0001). This result was the same in both single- and mixed-species crosses (Table 1; χ2H = 2.49, P > 0.12). Thus, in mixed pollinations, conspecific and heterospecific pollen tubes at the same stage of growth rarely contained generative cells at the same stage of the cell cycle (as in Fig. 4A). In most cases, heterospecific pollen tubes contained premitotic generative cell nuclei, whereas conspecific pollen tubes contained postmitotic sperm nuclei in the G1 or later stages of the cell cycle (Fig. 4B).
Table 1.
Proportion of viable generative cell nuclei at mitosis or beyond, in single-species (separate styles) and mixed-species (same style) crosses
| Maternal species | Single species
|
Mixed species
|
||
|---|---|---|---|---|
| Con ♂ | Het ♂ | Con ♂ | Het ♂ | |
| Water (2x) | 0.706 | 0.470 | 0.799 | 0.551 |
| (379) | (204) | (256) | (91) | |
| Paper (6x)* | 0.743 | 0.455 | 0.815 | 0.487 |
| (433) | (25) | (285) | (29) | |
Con, conspecific; Het, heterospecific. Sample size in parentheses. *Excluding one individual; see text.
Figure 4.

Pollen competition in water birch (2x) styles. (A) Sperm cell nuclei in water birch (2x) and paper birch (6x) pollen tubes at equivalent stages of development. (B) Undivided generative cell nucleus in paper birch (6x) pollen tube growing near water birch (2x) pollen tube containing sperm cell nuclei. (Bar = 10 μm.)
Delayed mitosis was even more pronounced when diploid water birch pollen tubes grew in polyploid paper birch styles (Table 1). In these trees, mitosis of conspecific generative cell nuclei was 4.7 times as likely to have been initiated as mitosis of heterospecific generative cell nuclei (χ2MH = 27.52, P < 0.0001). This result was the same in both single- and mixed-species crosses (Table 1; χ2H = 0.10, P > 0.76). On one HC paper birch mother, mitosis of heterospecific generative cells was not significantly different from mitosis of conspecific generative cells (single-species G = 3.34, P > 0.10; mixed-species G = 4.67, P > 0.10). In general, mitosis of heterospecific generative cells was significantly delayed relative to mitosis of conspecific generative cells in paper birch styles.
Seed Viability and Paternity.
To examine the outcome of pollen competition, we compared relative conspecific success (RCS) in single- vs. mixed-species crosses (Table 2). Paternity analysis of mixed-species seedlings indicated RCS was either equal to or significantly higher than expected, compared with RCS in the control (Table 2). In HC paper birch, heterospecific siring success was particularly high in the control (29.2%), but virtually absent in the mixed-species crosses (RCS >99%). Thus, conspecific pollen preferentially sired seeds even when control crosses indicated weak barriers.
Table 2.
Siring success in conspecific (Con), heterospecific (Het), and mixed-species (Mix) treatments (backtransformed mean/tree; n = # of progeny scored)
| Maternal species | RCS
|
G | P | ||||
|---|---|---|---|---|---|---|---|
| Proportion of seeds germinated
| |||||||
| Con | Het | Mix | Single species | Mixed species | |||
| Water birch | 0.180 | 0.015 | 0.134 | 0.923 | 0.982 | 8.95 | 0.005 |
| n = 1,693 | n = 1,650 | n = 1,673 | n = 128 | ||||
| HC paper | 0.470 | 0.292 | 0.436 | 0.617 | 0.992 | 28.22 | <0.0001 |
| birch | n = 378 | n = 211 | n = 457 | n = 32 | |||
| HI paper | 0.172 | 0.006 | 0.197 | 0.966 | 0.982 | 1.07 | 0.300 |
| birch | n = 1,317 | n = 1,299 | n = 1,442 | n = 112 | |||
G tests compare RCS in single-species versus mixed-species crosses. In the single-species crosses, RCS was calculated as % conspecific germination/(% conspecific germination + % heterospecific germination). In the mixed-species crosses RCS was calculated as the proportion of seedlings sired by conspecific fathers, as determined by paternity analysis. A value of 0.5 would indicate equivalent success.
The lack of heterospecific siring success in the mixed-species crosses cannot be attributed to preferential embryo abortion. If heterospecific fertilizations had resulted in aborted embryos in the mixed-species crosses, then mixed-species siring success would have been reduced relative to the conspecific control. Although water birch mixed-species seed germination was slightly lower than that of the conspecific control (13.4% vs. 18%, respectively; Table 2), the difference was not significant in this comparison or in the same two comparisons on paper birch mothers (two-tailed t test on arcsine transformed data; P > 0.35 in all three cases; Table 2).
The strong pollen tube viability barrier that we measured does not completely explain the high conspecific fertilization precedence. In the mixed-species crosses, conspecific pollen always had >98% siring success (Table 2), which was always significantly higher than the proportion of conspecific pollen tubes present in the mixed-species treatment styles after 48 hr (P < 0.0001 in all three cases). Processes other than pollen germination or early pollen tube incompatibility also must contribute to the lack of heterospecific fertilizations.
DISCUSSION
Genetic analysis of natural hybrid zones reveals that F1 hybrids can act as a bottleneck in the development of hybrid swarms (9). Understanding the developmental barriers to the formation of F1 hybrids offers key insights into forces that shape the genetic variation available for the processes of introgression and hybrid race, or species, formation. Birches and other highly outcrossing perennials have long stood as a paradigm for the strength of extrinsic selection in maintaining species boundaries (21–27). Baker (28) suggested that interspecific pollen competition might reinforce weak genetic barriers in such groups. An important finding of this study is that, regardless of the strength of other internal barriers, pollen competition strongly increases assortative mating. This conclusion is reflected by the higher relative siring success of conspecific gametes in mixed-species than in single-species crosses. Furthermore, because overall siring success in the mixed-species crosses was equal to that found in the conspecific control, the lack of F1 hybrids is not the result of preferential abortion of hybrid zygotes. Rather, heterospecific fertilization rarely occurs. Thus, in mixed-species crosses, postzygotic processes had little effect on reproductive isolation.
Table 3 summarizes stages in which prezygotic barriers were observed. Within diploid water birch styles, pollen tube incompatibility reduced the heterospecific pollen tube cohort by more than 50% (relative to that in native styles). Although the surviving heterospecific pollen tubes showed little evidence of an average growth deficiency, the cell cycle of generative cells within many pollen tubes was aberrant. It is likely that abnormal cell cycle behavior serves as an early indicator of reduced fertilization potential, because coordinate regulation of male and female gamete development has been observed in all plant species studied to date (29). Observations of phase two pollen tube development (J.H.W., unpublished data) suggest that incompatibility, variable pollen tube growth rates, and defective pollen tube guidance in the ovary also contribute to low heterospecific siring success.
Table 3.
Early postpollination barriers in birch styles
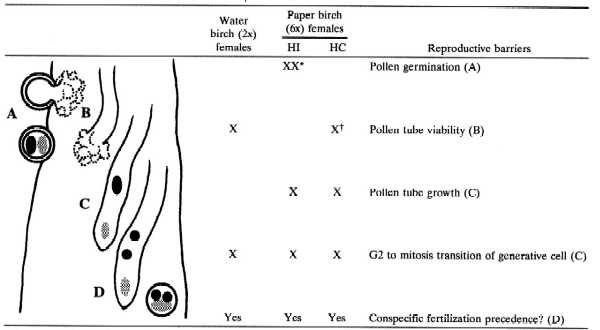 |
Heterospecific germination increased in presence of conspecific pollen: A, B, and C indicate abnormal development; D indicates normal development.
Conspecific viability increased in presence of heterospecific pollen.
On polyploid paper birch, flowers on seven of nine trees exhibited a strong HI barrier at the stigmatic surface (Table 3). The few heterospecific pollen tubes that entered stigmas exhibited reduced growth and delayed generative cell mitosis. Thus, in HI trees, strong early postpollination barriers were reinforced by later barriers, which resulted in a lack of heterospecific siring success in both single- and mixed-species crosses (Table 3). In contrast, two HC paper birch trees had about 29% heterospecific siring success in the control crosses. In these trees, the heterospecific pollen cohort germinated normally and experienced only a small reduction as a result of pollen tube incompatibility within the style. However, heterospecific pollen tubes that survived the viability barrier exhibited reduced growth and delayed generative cell mitosis. The delayed cell cycle may be a result of delayed pollen germination, because generative cell mitosis has been correlated with the duration of pollen tube growth (30). Late pollen germination may explain why in these two trees, heterospecific siring success was relatively high in the control crosses, but < 1% in the mixed-species crosses. Delayed pollen germination and stigma penetration, or slow pollen tube growth and aberrant generative cell mitosis, indicate poor competitive ability, but not the loss of fertilization potential.
In styles of both species, delayed division of heterospecific generative cells was not caused by a paternal genome size effect or a paternal genetic effect, because paper birch generative cell nuclei were equally likely to have initiated mitosis as water birch generative cell nuclei in their native styles. This finding suggests that the male gametic cell cycle is either negatively regulated in heterospecific styles or positively regulated in conspecific styles. Because generative cell mitosis often fails to occur in pollen tubes grown in vitro (ref. 31 and unpublished observations), the latter possibility seems more likely. The potential for rapid divergent evolution and reproductive isolation as a consequence of heterochronic perturbations to the developmental synchrony of the male and female gametic cell cycles is as yet unexplored (29, 32).
This study directly visualizes and identifies the paternity of pollen tubes interacting within the same style. Furthermore, we performed the experimental crosses in a natural population under natural pollen loads. Our results are thus a test of whether interspecific pollen competition in natural populations of birch occurs as a result of male-male interaction, male-female interaction, or both. With minor exception, we found little evidence for male-male interaction (Table 2). Both positive (e.g., mentor effects; ref. 33) and negative (e.g., interference, allelopathy, resource preemption; ref. 11) male-male interactions have been observed in vitro, or inferred in vivo from mixed-species pollinations. Such experiments, however, often use high pollen loads to increase the probability of detecting interactions or to mimic animal pollination. By mixing pollen and by applying pollen at one point in time, we maximized the opportunity for interactions in normal pollen loads. If such interactions exist in wind-pollinated birch styles, they appear to be of small effect.
Our results instead point to the overwhelming importance of male-female interactions in determining the success of heterospecific fertilization. European birches are known to possess gametophytic self-incompatibility (GSI), and the GSI system also has been implicated in heterospecific incompatibility (8, 34, 35). Flowers from trees of both species in this study had delayed heterospecific pollen germination and/or pollen tubes burst at the tip, which are both typical of an incompatibility reaction (8). When the polyploid species was the mother, the reaction was particularly strong and early in HI trees, whereas in HC paper birch and in all of the water birch mothers, the reaction was weak and occurred later. Polymorphism for heteroincompatibility has been noted in other wide polyploid crosses (36, 37) and in crosses between species with unilateral incompatibility (34, 38). Such polymorphism constitutes a breakdown of otherwise strong bidirectional genetic barriers and provides one avenue for asymmetric introgression.
Paper and water birch often occur in sympatry, and pollination occurs at the same time in both species. Thus, pollen competition often must limit crossfertilization between these species in nature. As a result, F1 hybrid formation has more likely occurred via pollen swamping of isolated flowering trees (i.e., by parental seed dispersal across local or geographic contact zones) than by pollen dispersal. Our data show that such hybrids are far more likely to be formed on HC paper birch females than on the more common HI paper birch or water birch females. Given these constraints and the strength of reproductive barriers, F1 hybrids will likely contain a subset of the genetic variation present in the parent species. Prezygotic selection on the many genes expressed during gametophytic and gametic development offers one explanation for nonrandom nuclear marker introgression across plant species boundaries (39).
Acknowledgments
We thank J. Hamrick, R. Price, D. Promislow, R. Wyatt, and two anonymous reviewers for comments on an earlier version of the manuscript, J. Hamrick for assistance with gel electrophoresis, and M. Grant for statistical advice. This work was supported by National Science Foundation Grant DEB-9703853 (M.L.A.); National Science Foundation Grant IBN-9696013 and grants-in-aid of research from Carl Ziess, Compaq computer, Lasergraphics, and Leica Instruments (W.E.F.), and National Science Foundation/U.S. Department of Agriculture/Department of Energy Training Grant BIR-9220329 (J.H.W.).
ABBREVIATIONS
- HI
heteroincompatible
- HC
heterocompatible
- RCS
relative conspecific success
References
- 1.Coyne J A. Nature (London) 1992;355:511–515. doi: 10.1038/355511a0. [DOI] [PubMed] [Google Scholar]
- 2.Soltis D E, Soltis P S. Crit Rev Plant Sci. 1993;12:243–273. [Google Scholar]
- 3.Muntzing A. Hereditas. 1933;18:33–55. [Google Scholar]
- 4.Levin D A. In: Evolutionary Biology. Hecht M H, Steere W C, Wallace B, editors. Vol. 11. New York: Appleton Century Crofts; 1978. pp. 185–317. [Google Scholar]
- 5.Buchholz J T. Bot Gaz. 1922;73:249–286. [Google Scholar]
- 6.Willson M F, Burley N. Mate Choice in Plants: Tactics, Mechanisms, and Consequences. Princeton: Princeton Univ. Press; 1983. [Google Scholar]
- 7.Dugle J R. Can J Bot. 1966;44:929–1007. [Google Scholar]
- 8.Hagman M. Proc R Soc London B. 1975;188:313–326. [Google Scholar]
- 9.Arnold M L. Natural Hybridization and Evolution. Oxford: Oxford Univ. Press; 1997. [Google Scholar]
- 10.Grant W F. Chromosoma. 1969;26:326–336. [Google Scholar]
- 11.Kearns C A, Inouye D W. Techniques for Pollination Biologists. Niwot, CO: Univ. Press of Colorado; 1993. [Google Scholar]
- 12.Dahl A E, Fredrikson M. Am J Bot. 1996;83:895–902. [Google Scholar]
- 13.Florjin R J, Slats J, Tanke H J, Raap A J. Cytometry. 1995;19:177–182. doi: 10.1002/cyto.990190213. [DOI] [PubMed] [Google Scholar]
- 14.Carmichael J S, Friedman W E. Plant Cell. 1995;7:1975–1988. doi: 10.1105/tpc.7.12.1975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Waser N M, Price M V. Ecology. 1991;72:171–179. [Google Scholar]
- 16.Sokal R R, Rohlf F J. Biometry. New York: Freeman; 1995. [Google Scholar]
- 17.Chasan R. Plant Cell. 1995;7:1–3. doi: 10.1105/tpc.7.11.1737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Rice W R. Evolution. 1989;43:223–225. doi: 10.1111/j.1558-5646.1989.tb04220.x. [DOI] [PubMed] [Google Scholar]
- 19.Hamrick J L, Schnabel A. In: Population Genetics in Forestry: Lecture Notes in Biomathematics. Gregorius H-R, editor. Vol. 60. New York: Springer; 1985. pp. 50–70. [Google Scholar]
- 20.Devlin B, Roeder K, Ellstrand N C. Theor Appl Genet. 1988;76:369–380. doi: 10.1007/BF00265336. [DOI] [PubMed] [Google Scholar]
- 21.Lotsy J P. Genetica. 1925;7:487–506. [Google Scholar]
- 22.Muller C H. Evolution. 1952;6:147–161. [Google Scholar]
- 23.Stebbins G L. Science. 1966;152:1463–1469. doi: 10.1126/science.152.3728.1463. [DOI] [PubMed] [Google Scholar]
- 24.Van Valen L. Taxon. 1976;2/3:233–239. [Google Scholar]
- 25.Grant V. Plant Speciation. New York: Columbia Univ. Press; 1981. [Google Scholar]
- 26.Templeton A R. In: Speciation and its Consequences. Otte D, Endler J A, editors. Sunderland, MA: Sinauer; 1989. pp. 3–27. [Google Scholar]
- 27.Ellstrand N C, Whitkus R, Rieseberg L H. Proc Natl Acad Sci USA. 1996;93:5090–5093. doi: 10.1073/pnas.93.10.5090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Baker H G. Biol Rev. 1951;26:302–337. [Google Scholar]
- 29.Friedman W E. Development (Cambridge, UK) 1999;126:1065–1075. doi: 10.1242/dev.126.5.1065. [DOI] [PubMed] [Google Scholar]
- 30.Brewbaker J L, Majumder S K. Am J Bot. 1961;48:457–464. [Google Scholar]
- 31.Tanaka I. Sex Plant Reprod. 1997;10:1–7. [Google Scholar]
- 32.Zhang X S, O’Neill S D. Plant Cell. 1993;5:403–418. doi: 10.1105/tpc.5.4.403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Stettler R F, Ager A A. In: Cellular Interactions. Linskens H F, Heslop-Harrison J, editors. Berlin: Springer; 1984. pp. 608–623. [Google Scholar]
- 34.Murfett J, Strabala T J, Zurek D M, Mou B, Beecher B, McClure B A. Plant Cell. 1996;8:943–958. doi: 10.1105/tpc.8.6.943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Newbigin E, Anderson M, Clarke A. Plant Cell. 1993;5:1315–1324. doi: 10.1105/tpc.5.10.1315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Lefol E, Fleury A, Darmency H. Sex Plant Reprod. 1996;9:189–196. [Google Scholar]
- 37.Jalani B S, Moss J P. Euphytica. 1980;29:571–579. [Google Scholar]
- 38.Mutschler M A, Liedl B E. In: Genetic Control of Self-Incompatibility and Reproductive Development in Flowering Plants. Williams E G, Clarke A E, Knox R B, editors. Dordrecht, The Netherlands: Kluwer; 1994. pp. 164–188. [Google Scholar]
- 39.Howard D J, Preszler R W, Williams J, Fenchel S, Boecklen W J. Evolution. 1997;51:747–755. doi: 10.1111/j.1558-5646.1997.tb03658.x. [DOI] [PubMed] [Google Scholar]