

Abstract
The factor catalyzing the first step in the synthesis of the characteristic pentaglycine interpeptide in Staphylococcus aureus peptidoglycan was found to be encoded by the essential gene fmhB. We have analyzed murein composition and structure synthesized when fmhB expression is reduced. The endogenous fmhB promoter was substituted with the xylose regulon from Staphylococcus xylosus, which allowed glucose-controlled repression of fmhB transcription. Repression of fmhB reduced growth and triggered a drastic accumulation of uncrosslinked, unmodified muropeptide monomer precursors at the expense of the oligomeric fraction, leading to a substantial decrease in overall peptidoglycan crosslinking. The composition of the predominant muropeptide was confirmed by MS to be N-acetylglucosamine-(β-1,4)-N-acetylmuramic acid(-l-Ala-d-iGln-l-Lys-d-Ala-d-Ala), proving that FmhB is involved in the attachment of the first glycine to the pentaglycine interpeptide. This interpeptide plays an important role in crosslinking and stability of the S. aureus cell wall, acts as an anchor for cell wall-associated proteins, determinants of pathogenicity, and is essential for the expression of methicillin resistance. Any shortening of the pentaglycine side chain reduces or even abolishes methicillin resistance, as occurred with fmhB repression. Because of its key role FmhB is a potential target for novel antibacterial agents that could control the threat of emerging multiresistant S. aureus.
Keywords: cell wall biosynthesis, murein, glycyl-tRNA, methicillin resistance, drug target
The staphylococcal cell wall plays an important role in infection and pathogenicity. Because of the uniqueness of the peptidoglycan structure and assembly it is one of the preferred targets of antibiotics. The Staphylococcus aureus peptidoglycan consists of linear sugar chains of alternating units of N-acetylglucosamine (GlcNAc) and N-acetylmuramic acid substituted with the pentapeptide l-Ala-d-iGln-l-Lys-d-Ala-d-Ala. Characteristic for S. aureus is the pentaglycine side chain that connects l-Lys of the stem peptide to the d-Ala in position 4 of a neighboring subunit, whereby the d-Ala in position 5 is split off by transpeptidation. The flexible pentaglycine interpeptide allows a peptidoglycan crosslinking degree of up to 90%, thus contributing substantially to cell wall stability (1). In addition, this pentaglycine chain acts as a recipient for staphylococcal surface proteins that are covalently anchored to it by a transpeptidase-like reaction (2). Surface proteins play an important role in adhesion and pathogenicity by interacting with host matrix proteins (3, 4). Staphylococci that have acquired the low-affinity penicillin-binding protein PBP2′ express intrinsic resistance to virtually all β-lactam antibiotics (5). Genetic studies have revealed that inhibition of pentaglycine side-chain formation reduces methicillin resistance without affecting PBP2′ synthesis, resulting in β-lactam hypersusceptibility (6–8). Thus, the interpeptide has several functions related to bacterial growth, cell wall stability, pathogenicity, and antibiotic resistance.
Synthesis of the pentaglycine chain occurs at the membrane-bound lipid II precursor GlcNAc-(β-1,4)-N-acetylmuramic acid(-l-Ala-d-iGln-l-Lys-d-Ala-d-Ala)-pyrophosphoryl-undecaprenol by sequential addition of glycine to the ɛ-amino group of lysine, using glycyl-tRNA as donor, in a ribosome-independent fashion (9). Chain formation depends on proteins of which two, FemA and FemB, have been identified previously. FemA is involved in the incorporation of glycyl residues 2 and 3 (6, 7), and FemB in that of glycyl residues 4 and 5 (10). Although FemA and FemB have a relatively high amino acid sequence identity (40%) and similarity (64%) they are specific and cannot substitute for each other (11). A third factor, FemX, has been postulated to be responsible for the incorporation of the first glycine (12). Because femAB null mutants are barely viable and depend on compensatory mutations for survival (8), femX is predicted to be a lethal target. Interestingly, S. aureus contains one glycyl-tRNA gene for protein biosynthesis and three nonproteinogenic glycyl-tRNA genes that are involved in cell wall biosynthesis (13, 14). It is tempting to speculate that each Fem factor may preferentially recognize one of the three glycyl-tRNA species.
Other FemAB-like factors have been identified in staphylococci, such as Lif in Staphylococcus simulans biovar staphylolyticus (15) and Epr in Staphylococcus capitis (16), which protect these strains from their own glycyl-glycine endopeptidase (17). Genome analysis of a nonpublic database produced three novel femAB-like sequences, fmhA, fmhB, and fmhC (synonym eprh; ref. 18) in S. aureus (19). Whereas inactivation of fmhA and fmhC apparently has no effect on growth and physiology, fmhB was shown to be essential and was postulated to be femX (19). To investigate its function in peptidoglycan biosynthesis we placed fmhB under the control of the xylose regulon of Staphylococcus xylosus (20) and could demonstrate a clear correlation between fmhB repression and accumulation of unsubstituted peptidoglycan monomer precursors, proving the assumption that FmhB has the activity of the hypothetical FemX.
MATERIALS AND METHODS
Strains, Plasmids, and Culture Conditions.
All S. aureus strains used in this study are derivatives of strain NCTC8325. S. aureus RN4220 is a phage-less, restriction negative strain transformable by electroporation (21). BB255 (22) was used as template for gene amplifications, and its methicillin-resistant derivative, strain BB270 (22) was used for fmhB promoter replacement. Plasmid pCX15 containing the xylose regulatory system of Staphylocccus xylosus was obtained from K. P. Wieland, University of Tübingen, Tübingen, Germany (20). The Escherichia coli-S. aureus shuttle vector pOX7, temperature sensitive for replication in S. aureus, conferring ampicillin resistance to E. coli and erythromycin resistance to S. aureus, was obtained from K. Dyke, University of Oxford (23). Growth medium was LB (Difco) supplemented with 0.5% glucose or 0.5% xylose where indicated. Transductions in S. aureus were made with generalized transducing phage 80α (24). Transformants and transductants were selected in the presence of 20 μg/ml of erythromycin at 30°C. Strains with chromosomally integrated recombinant temperature-sensitive plasmid were propagated in presence of 10 μg/ml of erythromycin at 42°C to maintain plasmid integration.
Susceptibility Tests.
Minimal inhibitory concentration (MIC) of the glycyl-glycine endopeptidase lysostaphin (Ambi, Trowbridge, U.K.) was determined by microbroth dilution using 104 cells in 100 μl of LB per well as described earlier (7). The MIC of methicillin was measured by Etest (AB Biodisk, Solna, Sweden) (25). When determining MICs in the presence of glucose, the inoculum was prepared from overnight cultures grown on plates containing glucose. Growth was read after 24-h incubation at either 35°C or 42°C.
DNA Manipulations.
Routine DNA manipulations were done according to protocols of Ausubel et al. (26) and Maniatis et al. (27). Sequencing was performed with an ABI 310 automated sequencer (Perkin–Elmer).
Isolation of RNA and Northern Blots.
Overnight cultures of S. aureus in LB were diluted 1:50 in LB plus supplements as indicated and grown to mid-log phase (OD600 = 0.7). The cells were harvested and processed with a FastRNA isolation kit (Bio 101) containing Trizol reagent (GIBCO Life Technologies, Basel) in a FastPrep reciprocating shaker (Bio 101) as described by Cheung et al. (28). Five micrograms of RNA was separated on a 0.8% denaturing gel, transferred to a Biodyne A nylon membrane (Pall) using transfer buffer containing 3 M NaCl, 8 mM NaOH, and 2 mM Na-lauroylsarcosine. Transcripts were detected with a digoxigenin-labeled DNA probe covering fmhB (Fig. 1). Digoxigenin labeling and detection were performed as recommended by Boehringer Mannheim, except for the following modifications: maleic acid buffer contained 0.1 M maleic acid, 3 M NaCl, pH 8, and blocking buffer contained 0.5% blocking reagent.
Figure 1.

Integration of pSR1 into the S. aureus chromosome. Black: fmhB gene; hatched: xylose regulon (xylR and the operator region of xylA); white: fmhB promoter region; bold line: pOX7 plasmid DNA. Relevant restriction sites are indicated as follows: H, HindIII; B, BamHI; K, KpnI; E, EcoRI. Primers used for cloning and PCR controls are indicated by arrows and numbered as in Materials and Methods. The region spanned by the internal DNA probe used for Southern and Northern blots is indicated by a line above fmhB. Not to scale.
Insertional Replacement of the fmhB Promoter.
xylR and the xylA promoter-operator region from S. xylosus located within a 1.7-kb HindIII–BamHI fragment in plasmid pCX15 were subcloned into pUC19 (29). A 630-bp fragment comprising the ribosome binding site and the 5′ region of fmhB (GenBank accession no. AF106849), amplified with the sense primer 1 (5′-GCGGATCCATTGTTAAATAGAAGGAGATATC-3′) and reverse primer 2 (5′-GCGGTACCCCCAGTGATTTTCATTAATTC-3′) from S. aureus BB255, was cloned into the BamHI and KpnI sites downstream of the xylA promoter region. The resulting fusion of the xylose regulon with the 5′ fragment of fmhB was sequenced to ensure fidelity of the PCR. The fusion product was excised by HindIII and EcoRI and subcloned into shuttle vector pOX7. The resulting plasmid pSR1 (Fig. 1) was electroporated into RN4220 from where it was transduced into BB270. To promote integration of the plasmid pSR1 into the chromosome of BB270, transductants grown at 30°C to stationary phase in LB broth containing 20 μg/ml of erythromycin were diluted 1:100 into fresh LB containing 2.5 μg/ml of erythromycin plus xylose, incubated at 40°C for 8 h, then rediluted 1:100 into fresh LB with 2.5 mg/ml of erythromycin plus xylose and incubated overnight at 40°C. Appropriate dilutions were spread on LB-agar plates containing 2.5 μg/ml of erythromycin plus xylose and incubated at 42°C. Colonies were checked for plasmid integration by PCR using sense primer 3 (5′-AAATGAACAATGTGCTATATTACC-3′) and reverse primer 4 (5′-GCCAGCAAACATTAATAGTGC-3′), as well as sense primer 5 (5′-GCGAATTCTTACACAAGCTCTGAATCGAC-3′) and reverse primer 6 (5′-GCCTGCAGATTTAGCTATTTCGGCATGAAG-3′) as indicated in Fig. 1. In the resulting strain SR18 (mec, fmhB∷pSR1) chromosomal integration was verified by Southern blot using a probe covering fmhB from the start codon to the HindIII site (Fig. 1).
Isolation of Peptidoglycan, Preparation, and Fractionation of Muropeptides.
Five milliliters of overnight cultures of strain SR18 or BB270 grown at 42°C was diluted into 500 ml of fresh medium, grown aerobically to the exponential growth phase, and harvested at an OD600 of 0.7. Peptidoglycan was isolated as described (7, 12). Lyophilized murein was enzymatically degraded, and the resulting muropeptides were reduced to their muramitol derivatives and analyzed by reversed-phase HPLC using a Waters 626 system (7, 12). Separated muropeptides were detected at 206 nm and identified by amino acid analysis, MS, or comparison with control samples (30).
Desalting of Muropeptides.
Selected peaks from a muropeptide separation were collected, chromatographed again under the same conditions, and desalted on a Waters 626 HPLC system using a ODS Hypersil column (Knauer, Berlin; particle size 3 μm, column dimensions 4 × 250 mm). Elution was performed at 30°C with a linear gradient from 0.05% trifluoroacetic acid (TFA) in water to 0.035% TFA in 30% CH3CN in 100 min starting 10 min after injection of the sample. The flow rate was 0.5 ml/min. Desalted muropeptides were eluted between 45 and 55 min.
Amino Acid Analysis and MS.
For amino acid analysis, 1 mg of lyophilized murein or desalted muropeptide was hydrolyzed in 6 N HCl at 166°C for 1 h, dried, and subjected to a Biotronic LC 5000 amino acid analyzer. For MS the desalted and lyophilized muropeptide was dissolved in 100 μl of 5% CH3CN containing 0.035% trifluoroacetic acid (TFA). Two microliters of the sample was introduced into a Finnigan LCQ mass spectrometer (Thermoquest, Bremen, Germany) by loop injection. The flow rate of the solvent (0.035% TFA in 50% CH3CN) was 40 μl/min. Data acquisition was performed between 200 and 1,800 Da with scan times in the order of 2–3 s.
RESULTS
Construction of SR18 (mec, fmhB∷pSR1).
Because fmhB could not be inactivated by various gene replacement strategies it was postulated to be essential (19). The fmhB promoter region in the S. aureus chromosome was replaced by the glucose-repressible xylose regulon of S. xylosus. For this purpose a shuttle plasmid, pSR1, temperature sensitive for replication in S. aureus, containing the xylose regulon of S. xylosus, fused to the 5′ end of the fmhB gene, was constructed and integrated into the S. aureus chromosome by homologous recombination. This integration led to replacement of the fmhB promoter with the xylose regulon as shown in Fig. 1. This promoter replacement was done in methicillin-resistant strain BB270 to simultaneously monitor the effects of fmhB repression on cell wall composition as well as expression of methicillin resistance. PCR on chromosomal DNA of strain SR18 with primer pair 3/4 produced a 1,041-bp band, whereas with primer pair 5/6 no product was detected, indicating chromosomal insertion of pSR1 into the fmhB gene (see Fig. 1). To ensure that the plasmid had integrated correctly without rearrangements, and that no free plasmid was present, Southern blots were performed with a probe covering either side of the 3′ end of the fmhB fragment in pSR1. HindIII digests of chromosomal DNA produced a single 3.1-kb band in BB255 and two bands of 2.6 and 9 kb in SR18. No free pSR1 was detectable, confirming the integration as shown in Fig. 1 (data not shown). Strain SR18 was maintained at 42°C in the presence of erythromycin to prevent excision of the plasmid.
The doubling time of SR18 at 42°C increased from 58 to 72 min upon addition of glucose. Addition of xylose, in contrast, had no significant effects and did not enhance growth of SR18 compared with unsupplemented LB medium, suggesting that in the uninduced state, the xylose regulon allowed a similar growth rate as when fmhB was controlled by its endogeneous promoter. The doubling time of the parent BB270 was not affected significantly by the addition of xylose and slightly decreased (15%) by glucose.
The fmhB transcript in BB270, determined by Northern blots, had a length of about 1,600 nt, suggesting that fmhB is a monocistronic gene. Transcription of fmhB from the uninduced xylA promoter in SR18 was about 10-fold higher than that from the endogenous fmhB promoter (data not shown). We assume that overexpression of fmhB is not toxic for the cells, as growth was unaffected. Addition of xylose to strain SR18 increased fmhB expression about 2-fold, whereas glucose reduced it substantially (Fig. 2). Although strongly repressed by glucose, there was residual activity of the xylA promoter, which may explain why SR18 still grew, albeit at a reduced rate, in the presence of glucose.
Figure 2.
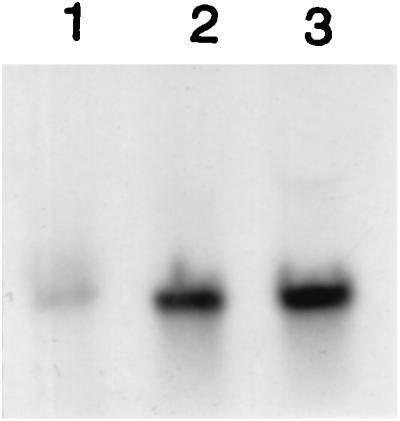
Northern blots of fmhB mRNA. Strain SR18 was grown in LB at 42°C to exponential phase in presence of glucose (lane 1), unsupplemented LB (lane 2), or xylose (lane 3). RNA was probed with the fmhB probe shown in Fig. 1.
Cell Wall Composition of S. aureus SR18.
The sequence similarity of FmhB to FemA and FemB, which are involved in the synthesis of the pentaglycine interpeptide bridge, suggests that FmhB may have a similar function. To confirm this assumption, the amino acid composition of the cell wall peptidoglycan isolated from strain SR18 grown at 42°C in presence of glucose or xylose was compared with that of the parental strain BB270 grown under identical conditions. As shown in Table 1, the amino acid composition of the cell walls of both strains grown in the presence of xylose was in agreement with the composition of S. aureus peptidoglycan; the molar ratio of glutamic acid to glycine was in the range of 1:4 to 1:5. In strain SR18 grown in the presence of glucose, a reduction in the cell wall glycine content in relation to glutamic acid was observed, and the molar ratio of alanine to glutamic acid was slightly increased. These data allow the deduction that repression of fmhB caused these changes in the amino acid composition of the peptidoglycan and that FmhB is involved in cell wall biosynthesis.
Table 1.
Cell wall amino acid composition of BB270 and SR18 grown with fmhB induction (xylose) or repression (glucose)
| Amino acid | BB270 xylose | BB270 glucose | SR18 xylose | SR18 glucose |
|---|---|---|---|---|
| Glu | 1.00 | 1.00 | 1.00 | 1.00 |
| Gly | 4.86 | 3.57 | 3.54 | 2.94 |
| Ala | 2.41 | 1.83 | 1.80 | 2.49 |
| Lys | 0.91 | 0.94 | 0.94 | 1.06 |
| Ser | 0.1 | 0.11 | 0.12 | 0.06 |
Molar masses of amino acids related to glutamic acid.
Muropeptide Analysis of S. aureus SR18.
To characterize the role of FmhB in cell wall biosynthesis in more detail, muropeptides isolated from BB270 and SR18 grown in the presence or absence of glucose were analyzed. The HPLC profile derived from the peptidoglycan of the strain BB270 grown at 42°C revealed the typical muropeptide pattern of a wild-type S. aureus strain with a high amount of oligomeric muropeptides and the highest peak in the dimeric fraction (Fig. 3a). No differences in the cell wall composition of the wild-type strain upon growth in the presence of either glucose or xylose could be observed, and both patterns corresponded to previous results found with this strain grown at 37°C (7, 12). The cell wall of SR18 grown in the presence of xylose (Fig. 3c) revealed the typical bell-shaped muropeptide profile, superimposable to that of strain BB270 (Fig. 3a). A strikingly different muropeptide profile was obtained from SR18 when grown in the presence of glucose (Fig. 3d). The amount of oligomeric muropeptides was reduced and the highest peak was found in the monomeric muropeptide fraction, indicating a reduced overall peptidoglycan crosslinking. Although the relative amounts of the dimeric and trimeric muropeptide fractions were in the same range for BB270 and glucose-repressed SR18, the overall crosslinking degree dropped from approximately 80% to 65% (Table 2), mainly because of the increase of the relative amounts of the monomeric muropeptides at the expense of the oligomeric fraction (Table 2). The lowered crosslinking of the fmhB-repressed strain SR18, caused by the abundance of uncrosslinked murein subunits carrying an unsubstituted stem peptide, also may explain the increased molar ratio of alanine to glutamic acid found in the amino acid cell wall composition of this strain.
Figure 3.

Muropeptide profiles. Isolated peptidoglycan was digested with muramidase and muropeptides were subjected to reversed-phase HPLC. Strain BB270 grown at 42°C (a) with xylose, (b) with glucose. Strain SR18 grown (c) with xylose, (d) with glucose. The muropeptide fractions are indicated as follows: monomers, corresponding to uncrosslinked subunits (retention time 10–45 min); dimers, corresponding to two crosslinked muropeptides (45–85 min); trimers, corresponding to three crosslinked muropeptides (85–105 min); and oligomers, corresponding to three or more crosslinked muropeptides. More highly crosslinked muropeptides in the oligomeric fraction are indicated by numbers.
Table 2.
Amount of muropeptides and overall crosslinking degree (%) of BB270 and SR18 grown with xylose or glucose
| Strain | Monomer* | Dimer* | Trimer* | Oligomer* | CL† |
|---|---|---|---|---|---|
| BB270 xylose | 4.2 | 7.9 | 7.5 | 80.4 | 81.3 |
| BB270 glucose | 6.7 | 11.2 | 10.0 | 72.1 | 77.2 |
| SR18 xylose | 5.0 | 8.8 | 7.9 | 78.3 | 80.2 |
| SR18 glucose | 19.9 | 11.7 | 9.0 | 59.4 | 65.4 |
Peak areas of the indicated fractions were added and calculated in percent.
The value was calculated as follows: 0.5 × dimer (%) + 67 × trimer (%) + 0.9 × oligomer (%) = CL (crosslinking) (33).
A detailed analysis of the monomeric and dimeric muropeptide fraction SR18 grown in presence of xylose or glucose is depicted in Fig. 4.
Figure 4.

Detailed view of the monomeric and dimeric muropeptide pattern of SR18 grown with fmhB induction (a) and repression (b). Peaks were identified by their retention time upon comparison with standard samples. Peak M1 was analyzed by MS. The peak nomenclature is shown in Table 3.
When strain SR18 was grown in the presence of xylose, the majority of muropeptides contained a pentaglycine side chain represented by the large peak M4 (Fig. 4a, for peak nomenclature see Table 3), and only minor amounts of muropeptides containing no (M1), one (M2), or three (M3) glycine residues were detected. A drastic change in cell wall composition was triggered by glucose (Fig. 4b). The monomer pattern was dominated by the nonsubstituted pentapeptide (M1) (retention time 16 min). A novel peak, which was shown by a pH shift to contain the nonamidated form of M1 plus a second nonidentified muropeptide component showing no pH shift, was detected in the monomeric fraction and designated M11. Concomitantly, the amounts of muropeptides modified by one (M2), three (M3), or five (M4) glycine residues were substantially reduced (Fig. 4b). In addition, the degradation products of these monomeric subunits carrying stem tetrapeptides instead of stem pentapeptides (M2*, M6*) could no longer be detected. Peak M9, which represents the nonamidated form of the pentaglycine-modified muropeptide M4, also was found in reduced amounts, whereas the quantity of the muropeptide species M10, carrying an alanine residue in position one of the interpeptide side chain was unchanged.
Table 3.
Peak nomenclature
| Peak | Structure | Abbreviations |
|---|---|---|
| M1 | M.P. | M. monomer; P. pentapeptide as stem peptide |
| M2 | M.P.G1 | Monomer pentapeptide modified with one glycine |
| M2* | M.T.G2/2 | Tetrapeptide as stem peptide; G glycine residue; 2/2 two glycine residues per two possible attachment sites (Lys in position 3 or Ala in position 4) |
| M3 | M.P.G3 | Monomer pentapeptide modified with three glycines |
| M4 | M.P.G5 | Monomer pentapeptide modified with five glycines |
| M6* | M.T.G6/2 | See M2* |
| M9 | M.Pn.G5 + M.P.A1 | A alanine; n, glutamic acid instead of glutamine in position 2; two muropeptides with identical retention time |
| M10 | M.P.G4A1 | See M4 and M9 |
| M11 | M.Pn. | See M1 plus unknown muropeptide component |
| D1 | D.P.G5 + T.G1 or D.P.G1 + T.G5 | Dimer of M4 and M2 |
| D2* | D.2xT.G11/3 | Dimer of M4 (degradation product) |
| D3 | D.P.G5 + T.G5 | Dimer of M4 |
| D4 | D.P.G5 + T.G3 | Dimer of M3 and M4 |
| D5 | D.Pn.G5 + T.G5 or D.P.G5 + Tn.G5 | Dimer of M4 and M9 |
| D10 | D.P.G5 + T.G4A1 or D.P.G4A1 +T.G5 | Dimer of M4 and M10 |
| T3 | D.P.G5 + 2xT.G5 | Trimer of M4 |
| Proposed structures for novel peaks | ||
| x | M.T.G1/1 | See M2*; Gly at position 4 of alanine |
| y1 | M.P. + M.P. | Dimer of M1 |
| y2 | M.P. + M.P.G1 | Dimer of M1 and M2 |
| y3 | M.P. + M.P.G5 | Dimer of M1 and M2 |
| z2 | M.P. + M.P.G1 + M.P.G5 | |
| z3 | M.P. + M.P.G5 + M.P.G5 | |
Structures are probably degradation products produced by an endopeptidase.
In contrast to the monomeric fraction, the dimer pattern of glucose-grown SR18 was found to be similar to that of a wild-type strain, although the amounts of these muropeptides were strongly reduced. The highest peak (D3) was the same as in the wild type or in S. aureus SR18 grown in the presence of xylose (Fig. 4a) and was composed of two crosslinked monomers carrying pentaglycine side chains (Fig. 4b). It thus can be assumed that the unmodified muropeptide M1 cannot be used efficiently for crosslinking in the peptidoglycan network.
Additional peaks (y and z in Fig. 4b) were detected in minor quantities in the glucose-repressed strain SR18 that were not present in the wild type. Based on the retention time and the typical profile of the muropeptide separation pattern of the wild type it is speculated that these peaks are dimeric (y) and trimeric (z) muropeptides composed of the unsubstituted muropeptide M1 crosslinked to one or two pentaglycine-modified muropeptides (y3 and z3 in Fig. 4b) or one or two monoglycine containing muropeptides (y2 and z2 in Fig. 4b). Correspondingly, it is possible that y1 contains two unsubstituted muropeptides directly crosslinked between the lysine residue in position three of the acceptor muropeptide and the alanine in position four of the donor peptide, although the direct crosslinking of muropeptides without the glycine interpeptide bridge has never been observed in wild-type S. aureus strains. Because these novel peaks were found only in minor quantities, we were not able to analyze their structures by MS or amino acid analysis.
Besides the novel peaks found in the dimeric and trimeric muropeptide fractions, an additional unknown peak x (Fig. 4b) with a lower retention time than the nonsubstituted muropeptide M1 was detected in relatively high abundance in the glucose-repressed strain SR18. We speculate that this peak corresponds to the unmodified muropeptide containing a stem tetrapeptide instead of a stem pentapeptide, which might be derived from formerly crosslinked cell wall material by the action of an endopeptidase. In this case the peak should contain glycine linked to the alanine in position 4 of the stem peptide. The comparatively high amount of this peak may indicate that crosslinked muropeptides composed of one unsubstituted moiety are unstable and preferred substrates for an endopeptidase.
Structure of the Muropeptide M1.
To verify the structure of muropeptide M1 found in the glucose-repressed strain SR18, it was isolated and subjected to amino acid analysis. The following molar mass ratios in relation to glutamic acid (1.0) were found: alanine, 3.01, and lysine, 1.08. No glycine or other amino acids were detected, consistent with the amino acid composition of an unmodified monomeric muropeptide. MS analysis of the whole peptide indicated a molecular mass of 968.4 Da with a smaller component of 765.4. The difference of 203 Da corresponds to the mass of GlcNAc and can be explained by the spontaneous loss of this residue. MS experiments of the fragmentation products obtained from the ion at 968.4 Da and the ion at 765.4 Da were found to be identical with a difference of 203 Da for the GlcNAc, indicating the presence of the typical stem peptide in both molecules (data not shown). Combining the results of amino acid composition and MS the proposed structure, GlcNAc-(β-1,4)-N-acetylmuramic acid(-l-Ala-d-iGln-l-Lys-d-Ala-d-Ala), for the muropeptide M1 was confirmed.
Susceptibility to Methicillin and Lysostaphin.
Methicillin resistance in S. aureus is the result of the acquisition of the foreign PBP2′ and can be expressed only when the pentaglycine interpeptide is present. Resistance is abolished in femAB null mutants that produce a peptidoglycan precursor with only one glycine residue (7). A similar effect was expected to occur in SR18 after fmhB repression. Because this strain had to be maintained at 42°C one must take into account the temperature dependence of methicillin resistance. At 35°C the methicillin-resistant parent BB270 had an MIC of methicillin of 16 μg/ml, which was reduced to 1.5 μg/ml at 42°C. Strain SR18 had a similar MIC of methicillin of 1 μg/ml at 42°C, and in the presence of glucose the MIC was reduced to 0.19 μg/ml. Addition of glucose to the medium had no significant effect on the methicillin MIC in BB270 at either temperature.
Strains with a single glycine residue in the interpeptide, such as femAB mutants, are resistant to the glycyl-glycine endopeptidase lysostaphin. SR18 grown in the presence of glucose has a poorly crosslinked peptidoglycan but still possesses intact pentaglycine side chains. Interestingly, this structural difference rendered strain SR18 more susceptible to lysostaphin, decreasing the MIC more than 10-fold from 0.125 μg/ml to 0.01 μg/ml. Glucose did not affect the susceptibility of BB270 to lysostaphin, which had an MIC of 0.21 μg/ml at 35°C and 0.25 at 42°C.
DISCUSSION
Elucidation of the role of FemA and FemB in interpeptide synthesis (7, 10) led to the postulation of a hypothetical factor FemX responsible for the first step in pentaglycine side-chain formation (12) (Fig. 5). Inactivation of FemX is expected to lead to an accumulation of peptidoglycan precursors. Repression of fmhB caused a decrease in cell wall glycine content and an increase in muropeptides containing no glycine residues at the expense of oligomeric muropeptides. The structure of the accumulating unsubstituted muropeptide M1, confirmed by MS, demonstrated that FmhB corresponds to the postulated FemX needed for the addition of the first glycine to the ɛ-amino group of l-Lys in the muropeptide precursor. Although the specificities of FmhB, FemA and FemB in pentaglycine synthesis are now clear, we cannot rule out involvement of additional factors in the process.
Figure 5.

Pentaglycine side-chain formation and peptidoglycan crosslinking. (Upper) Transpeptidase reaction crosslinking the fifth glycine residue of the acceptor muropeptide to the d-Ala of the donor stem peptide with release of the terminal d-Ala. Direct linkage to the ɛ-amino group of lysine does not occur in S. aureus. (Lower) Synthesis of the pentaglycine side chain at the membrane-bound lipid II intermediate.
Transcription of fmhB in SR18 was not completely abolished by the addition of glucose, leading to an intermediate phenotype with regard to the muropeptide composition. Although the amount of the unsubstituted muropeptide M1 was substantially increased, few muropeptides containing one, three, and five glycine residues were still present. Although the oligomeric muropeptide fraction was greatly reduced, the relative amounts of the dimeric and trimeric muropeptides were not significantly changed, showing that the unsubstituted muropeptide was crosslinked into dimeric and trimeric muropeptides but could not be efficiently incorporated into the more highly crosslinked staphylococcal peptidoglycan. The muropeptides containing a pentaglycine interpeptide are better substrates for the PBPs and consequently were incorporated into highly crosslinked peptidoglycan, whereas the unsubstituted muropeptides were found mainly in the monomeric or dimeric muropeptide fraction. Novel peaks found in the glucose-repressed strain in minor quantities suggested the existence of crosslinked muropeptides containing one unsubstituted moiety. In the crosslinking reaction, the bond between the C-terminal d-Ala-d-Ala of the monomeric murein subunit, the donor peptide, is cleaved and the energy is used to transfer this monomeric subunit to the amino group of the terminal glycine residue of the interpeptide side chain of a nascent murein subunit, the acceptor peptide, releasing the alanine residue in position five of the donor. Newly synthesized unmodified murein subunits therefore can still function as donor peptides and be incorporated into the pre-existing murein network (Fig. 5) but cause a dead-end structure. The resulting structure missing a glycine side chain would require a direct linkage of the next donor to the ɛ-amino group of the lysine residue of the acceptor peptide, a reaction that has not been observed in wild-type staphylococci. It has been shown that cell surface proteins in staphylococci are covalently linked to the pentaglycine interpeptide (31). Direct crosslinking of muropeptides as it is known in Gram-negative bacteria thus would exclude anchorage of staphylococcal surface proteins. Hence it becomes clear that FmhB is essential for cell growth as well as pathogenicity.
Although femAB null mutants with mono-glycine side chains are highly resistant to the glycyl-glycine endopeptidase lysostaphin (7), the increased susceptibility to lysostaphin upon fmhB repression was likely the result of the few remaining complete pentaglycine crossbridges, because no direct crosslinking between unsubstituted stem peptides was observed. PBP2′-mediated methicillin resistance, which depends on the biosynthesis of the pentaglycine side chain (8), was reduced accordingly in the fmhB-repressed strain, showing that pentaglycine-substituted precursors in sufficient amounts are required for expression of methicillin resistance.
Signature-tagged mutagenesis in a murine model of bacteriaemia revealed femA and femB as well as a novel factor with significant homologies to femB to be important in virulence (32), of which the latter may be fmhB. The essentiality of fmhB, its key role in the pentaglycine side- chain formation, the requirement of the pentaglycine side chain for covalent linkage of cell wall-associated protein and for expression of methicillin resistance, suggest FmhB as a potential target for antibacterial strategies that may be extended to other pathogenic bacteria with interpeptide bridges such as streptococci and enterococci.
Acknowledgments
We gratefully acknowledge the technical assistance of K. Servan and M. Loriguillo. We thank D. Immler for assistance with MS and W. Schröder for assistance with amino acid analysis. Research was supported by Swiss National Science Foundation Grant 31-52239.97 to B.B.-B.
ABBREVIATIONS
- PBP
penicillin-binding protein
- MIC
minimal inhibitory concentration
Footnotes
This paper was submitted directly (Track II) to the Proceedings Office.
References
- 1.Labischinski H. Med Microbiol Immunol. 1992;181:241–265. doi: 10.1007/BF00198846. [DOI] [PubMed] [Google Scholar]
- 2.Schneewind O, Fowler A, Faull K F. Science. 1995;268:103–106. doi: 10.1126/science.7701329. [DOI] [PubMed] [Google Scholar]
- 3.Mempel M, Schmidt T, Weidinger S, Schnopp C, Foster T, Ring J, Abeck D. J Invest Dermatol. 1998;111:452–456. doi: 10.1046/j.1523-1747.1998.00293.x. [DOI] [PubMed] [Google Scholar]
- 4.Foster T J, Mcdevitt D. FEMS Microbiol Lett. 1994;118:199–205. doi: 10.1111/j.1574-6968.1994.tb06828.x. [DOI] [PubMed] [Google Scholar]
- 5.Song M D, Wachi M, Doi M, Ishino F, Matsuhashi M. FEBS Lett. 1987;221:167–171. doi: 10.1016/0014-5793(87)80373-3. [DOI] [PubMed] [Google Scholar]
- 6.Maidhof H, Reinicke B, Blümel P, Berger-Bächi B, Labischinski H. J Bacteriol. 1991;173:3507–3513. doi: 10.1128/jb.173.11.3507-3513.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Strandén A M, Ehlert K, Labischinski H, Berger-Bächi B. J Bacteriol. 1997;179:9–16. doi: 10.1128/jb.179.1.9-16.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ling B D, Berger-Bächi B. Antimicrob Agents Chemother. 1998;42:936–938. doi: 10.1128/aac.42.4.936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kamiryo T, Matsuhashi M. J Biol Chem. 1972;247:6306–6311. [PubMed] [Google Scholar]
- 10.Henze U, Sidow T, Wecke J, Labischinski H, Berger-Bächi B. J Bacteriol. 1993;175:1612–1620. doi: 10.1128/jb.175.6.1612-1620.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ehlert K, Schröder W, Labischinski H. J Bacteriol. 1997;179:7573–7576. doi: 10.1128/jb.179.23.7573-7576.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kopp U, Roos M, Wecke J, Labischinski H. Microb Drug Resist. 1996;2:29–41. doi: 10.1089/mdr.1996.2.29. [DOI] [PubMed] [Google Scholar]
- 13.Roberts J R, Lovinger G G, Tamura T, Strominger J L. J Biol Chem. 1974;249:4781–4786. [PubMed] [Google Scholar]
- 14.Green C J, Vold B S. J Bacteriol. 1993;175:5091–5096. doi: 10.1128/jb.175.16.5091-5096.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Schindler C A, Schuhardt V T. Biochim Biophys Acta. 1965;97:242–250. doi: 10.1016/0304-4165(65)90088-7. [DOI] [PubMed] [Google Scholar]
- 16.Sugai M, Fujiwara T, Ohta K, Komatsuzawa H, Ohara M, Suginaka H. J Bacteriol. 1997;179:4311–4318. doi: 10.1128/jb.179.13.4311-4318.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Thumm G, Götz F. Mol Microbiol. 1997;23:1251–1265. doi: 10.1046/j.1365-2958.1997.2911657.x. [DOI] [PubMed] [Google Scholar]
- 18.Sugai M, Fujiwara T, Komatsuzawa H, Suginaka H. Gene. 1998;224:67–75. doi: 10.1016/s0378-1119(98)00508-3. [DOI] [PubMed] [Google Scholar]
- 19.Tschierske M, Mori C, Rohrer S, Ehlert K, Shaw K J, Berger-Bächi B. FEMS Microbiol Lett. 1999;171:97–102. doi: 10.1111/j.1574-6968.1999.tb13417.x. [DOI] [PubMed] [Google Scholar]
- 20.Wieland K-P, Wieland B, Götz F. Gene. 1995;158:91–96. doi: 10.1016/0378-1119(95)00137-u. [DOI] [PubMed] [Google Scholar]
- 21.Kreiswirth B N, Löfdahl S, Bentley M J, O’Reilly M, Schlievert P M, Bergdoll M S, Novick R P. Nature (London) 1983;305:680–685. doi: 10.1038/305709a0. [DOI] [PubMed] [Google Scholar]
- 22.Berger-Bächi B. J Bacteriol. 1983;154:479–487. doi: 10.1128/jb.154.1.479-487.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.O’Connell C, Pattee P A, Foster T J. J Gen Microbiol. 1993;139:1449–1460. doi: 10.1099/00221287-139-7-1449. [DOI] [PubMed] [Google Scholar]
- 24.Berger-Bächi B, Kohler M L. FEMS Microbiol Lett. 1983;20:305–309. [Google Scholar]
- 25.Baker C N, Stocker S A, Culver D H, Thornsberry C. J Clin Microbiol. 1991;29:533–538. doi: 10.1128/jcm.29.3.533-538.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ausubel F M, Brent R, Kingston R E, Moore D D, Seidman J G, Smith J A, Struhl K. Current Protocols in Molecular Biology. New York: Wiley; 1987. [Google Scholar]
- 27.Maniatis T, Fritsch E F, Sambrook J E. Molecular Cloning: A Laboratory Manual. Plainview, NY: Cold Spring Harbor Lab. Press; 1982. [Google Scholar]
- 28.Cheung A L, Eberhardt K, Fischetti V A. Anal Biochem. 1994;222:511–514. doi: 10.1006/abio.1994.1528. [DOI] [PubMed] [Google Scholar]
- 29.Yanisch-Perron C, Vieira J, Messing J. Gene. 1985;33:103–119. doi: 10.1016/0378-1119(85)90120-9. [DOI] [PubMed] [Google Scholar]
- 30.Roos M, Pittenauer E, Schmid E, Beyer M, Reinike B, Allmaier G, Labischinski H. J Chromatogr B. 1998;705:183–192. doi: 10.1016/s0378-4347(97)00506-9. [DOI] [PubMed] [Google Scholar]
- 31.Ton-That H, Labischinski H, Berger-Bächi B, Schneewind O. J Biol Chem. 1998;273:29143–29149. doi: 10.1074/jbc.273.44.29143. [DOI] [PubMed] [Google Scholar]
- 32.Mei J M, Nourbakhsh F, Ford C W, Holden D W. Mol Microbiol. 1997;26:399–407. doi: 10.1046/j.1365-2958.1997.5911966.x. [DOI] [PubMed] [Google Scholar]
- 33.Fordham W D, Gilvarg C. J Biol Chem. 1974;259:2478–2482. [PubMed] [Google Scholar]