

Short abstract
RNA interference is implicated in the maintenance of Drosophila telomeres by retrotransposons.
Abstract
Unlike many other organisms, Drosophila maintains its telomeres by the transposition of retrotransposons to chromosome ends. Recent work shows that proteins in the RNA interference pathway specifically regulate the expression of these retrotransposons and frequency of transposition in germline cells, but do not affect retrotransposon expression or telomere function in the soma.
At first glance, the telomeres of the fruit fly Drosophila melanogaster seem very different from those of other organisms. At a second glance, however, the difference is mostly in the size and sequence of the DNA repeats that make up the telomeres. Unlike the simple sequence telomere repeats of most eukaryotes, the Drosophila repeats are made of telomere-specific transposable elements and are three orders of magnitude larger than the repeats found in most eukaryotes (6-12 kb versus 5-9 bp; Figure 1). The Drosophila telomere elements are non-LTR retrotransposons, which transpose by poly(A)+ RNA that is reverse transcribed directly onto the chromosome. Thus, successive transpositions onto the end of the chromosome extend the telomere, as do the simple repeats added by telomerase in other organisms. Nevertheless, both telomerase-associated repeats and retrotransposons appear to serve the same functions in the biology of the cell [1] - such as maintaining the length of chromosome ends, distinguishing true ends from breaks in DNA, preventing fusion of chromosome ends and facilitating meiotic chromosome movements -and there are strong similarities in the basic mechanism of telomere maintenance between Drosophila and other organisms. In both cases, sets of repeats are added to chromosome ends by reverse transcription of an RNA template; in the case of Drosophila this template is the RNA intermediate of transposition. As a variant that accomplishes the same ends by slightly different means, the Drosophila telomere can give us insight into unanswered questions about other telomeres, including our own. It is becoming clear that telomeres have many functions, yet we understand little about how they actually accomplish any of them.
Figure 1.
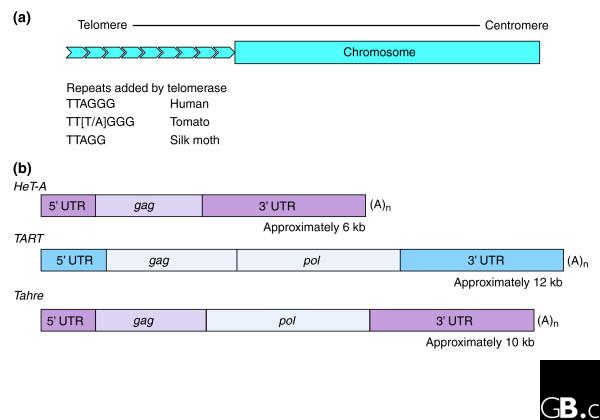
Comparison of telomeres maintained by telomerase with those maintained by the transposition of retrotransposons. (a) Eukaryotic telomeres are composed of long chains of head-to-tail repeats. In many organisms other than Drosophila, the repeats are short simple sequences of around 6 bp and telomeres are maintained by addition of repeats to the ends of the telomeres by the enzyme telomerase. (b) In Drosophila, each repeat is a non-LTR retrotransposon, of which there are three different types, HeT-A, TART, and Tahre, ranging from 6 kb to 12 kb in length. These elements transpose as poly(A)+ RNA and are reverse transcribed onto the end of the chromosome to extend the telomere. The gag and pol are characteristic retroviral genes encoding structural proteins and viral enzymes, respectively. The colors of the segments indicate that HeT-A and Tahre share sequences of 5' UTR, 3' UTR, and gag while TART and Tahre have similar pol sequences but have very different 5' UTR, 3' UTR and gag sequences.
The RNA interference machinery affects telomere elongation in Drosophila
Although the mechanism of transposition of non-LTR transposons is well understood, there is still much to learn about how the addition of retrotransposon elements to the telomere array is regulated. A recent paper by Savitsky et al. [2] provides intriguing evidence that proteins associated with RNA interference (RNAi) may be involved. Earlier work from this same group [3] had shown that mutant alleles of two genes encoding components of the RNAi machinery lead to increased accumulation of RNA from several transposable elements in female germline cells [4]. These two genes, spindle-E (spn-E) and aubergine (aub), respectively encode a DEAD-box helicase [5] and the Aub protein, a member of the Argonaute family that is necessary for assembly of the RNA-induced silencing complex (RISC) [6]. The RNAi machinery is generally thought to have evolved to help cells battle the invasion of parasitic elements, so the increased accumulation of transposable elements in cells with defective RNAi genes was not surprising. It was surprising, however, that one of the transposable elements in this study was HeT-A, a transposable element dedicated to telomere maintenance in Drosophila, not a parasitic invader. Savitsky and colleagues [2] have now extended these studies to show that the RNAi genes are indeed acting on components of the telomere.
Savitsky et al. [2] have now further explored the effects of spn-E and aub mutations on telomere maintenance, and find evidence that these two genes have a regulatory role. The mutant phenotypes indicate that these genes are involved in controlling the flow of telomere elements to the ends of chromosomes. The results of the study show that this control is complex, affecting the telomere-specific elements in different ways.
Apart from HeT-A, the two other telomere-specific retro-transposons are TART and Tahre. Savitsky et al. [2] show that the expression of TART is also affected by mutant alleles of spn-E and aub. Tahre may be affected but, because it shares so much of the HeT-A sequence [7], it would be included in the HeT-A results in these studies. The mutations studied affect HeT-A and TART differently. Firstly, although both RNAs were upregulated specifically in germline cells, they had different patterns of accumulation. The increase in TART RNA was detected in the nurse cells in the later stages of oogenesis. In contrast, increased levels of HeT-A transcripts were detected in oocytes even in early stages and were then seen later in nurse cells. Secondly, the effect of mutation on HeT-A expression was less robust than on TART, with all mutant alleles increasing TART expression but some having no effect on HeT-A. For example, several aub alleles increased HeT-A expression in only a fraction of the ovaries, or sometimes only in some ovarioles in a single ovary. Only in one case was HeT-A expression increased to a higher level than TART: in homozygous spn-E1 flies TART abundance is about twice that seen in heterozygotes, whereas HeT-A is over ten times more abundant.
The differences in the response of HeT-A and TART might seem surprising, but they are less so when considered in the light of other differences between these two elements. Indeed, the two retrotransposons are regulated quite differently in non-mutant flies. HeT-A is expressed only in diploid cells, predominantly in S-phase cells, and yields few, if any, antisense transcripts. TART is expressed in both diploid and polyploid cells and produces much more antisense than sense RNA [8]. The effects seen by Savitsky et al. [2] now show that the regulation of HeT-A and TART expression by RNAi is also different.
Increased expression of HeT-A and TART correlates with increased transposition to telomeres in the germline
If spn-E and aub have a role in telomere maintenance, mutant flies with increased RNA expression should also have increased addition of elements to telomere arrays. Testing this possibility is difficult, because wild-type telomeres contain many elements, and there is no good way to detect and quantify new additions. The Georgiev group has, however, developed an elegant system to screen for new additions to a broken chromosome end in the germline [9]. This system is built on several observations. First, mobilization of P elements frequently results in broken chromosome ends that are capable of passing through cell-cycle checkpoints and completing the cell cycle. Second, the yellow gene, which controls cuticle pigmentation, is located near the telomere of the X chromosome with its promoter at the distal end, that is, the end nearest the telomere. Third, the enhancer controlling yellow expression in bristles is in the intron, but enhancers affecting expression in the cuticle are distal to exon 1. Thus cuticle enhancers are nearer to the telomere than bristle enhancers. Flies with a terminally deleted X chromosome broken in the region of the upstream enhancers have mutant (yellow) body color but wild-type bristles because they lack the cuticle enhancers but still have the bristle enhancer in the intron. Finally, transposition of either HeT-A or TART to the broken X chromosome end restores wild-type pigmentation to the aristae (terminal segments of the antenna). Thus, the number of flies with wild-type aristae in the progeny of a fly with the broken X chromosome measures the rate of transposition in the parental germline (Figure 2). The DNA sequence of the newly transposed elements in these progeny can be studied because the elements are attached to the easily identifiable yellow gene.
Figure 2.
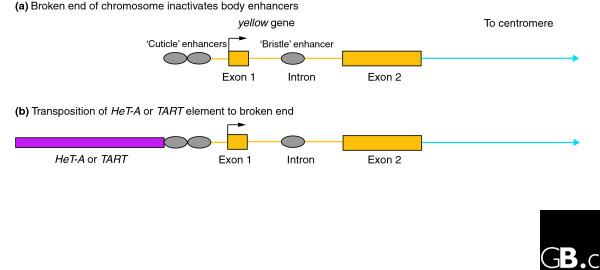
Assay to measure the frequency of transposition onto a chromosome end. Expression of the yellow gene (which controls normal pigmentation) is controlled in different body tissues by enhancers located in different regions of the gene. (a) A break at the end of the chromosome inactivates upstream enhancers that direct expression of the yellow gene in the cuticle, producing flies with a yellow (mutant) body but still producing normal pigmentation in the bristles, as directed from the intronic enhancer. (b) Transposition of HeT-A or TART to the broken end reactivates the upstream enhancer for expression of yellow+ in the aristae (the terminal segments of the antenna), and these structures are pigmented normally.
Using this system, Savitzky et al. [2] clearly show that the increased transcription in the mutant flies is correlated with an increased frequency of transposition of the retrotransposons and consequently in telomere elongation. For non-mutant flies the frequency of transposition onto the broken end was 0.04%. In flies carrying a single copy of either a spn-E or an aub mutation, this rate was increased by between 20- and 100-fold or more, depending on the allele. Surprisingly, more than 95% of the new transpositions were of TART, whereas the majority of transpositions in wild-type flies are of HeT-A [9,10]. Only in homozygous spn-E1 flies, where HeT-A expression is dramatically increased, is HeT-A transposition more frequent than that of TART. Clusters of progeny with identical TART attachments indicated that at least some of the transposition occurred in premeiotic cells. These results add to the increasingly complex picture of regulation of the telomeric retrotransposons.
Telomere length regulation is complex
Savitsky et al. [2] show that the spn-E and aub mutations lead to increased HeT-A and TART expression, which correlates with their increased rate of transposition to chromosome ends. But it is important to note that despite the greatly increased frequency of attachment of HeT-A and TART to broken ends observed in this assay, lines heterozygous for spn-E or aub mutations do not have detectably greater numbers of HeT-A and TART in their genomes [2]. This suggests that there are further levels of regulation of telomere elongation involving additional important players. One explanation might be that the mechanism of elongation studied here could act only on broken chromosome ends. On the other hand, it could be acting on all chromosome ends, including unbroken telomeres. The frequency of retrotransposon addition in mutant flies is low enough for it to take generations to make a significant change in the very long Drosophila telomeres, even though addition is easily detected with the powerful screen of a broken chromosome. Similarly, the lack of an RNAi effect on either expression of HeT-A or TART [2] or telomere fusions [11] in somatic tissues indicates that regulation of telomere length in somatic tissues is independent of RNAi, or at least does not involve the RNAi genes studied here.
Earlier work has shown that mutations in the gene for HP1, a chromatin protein, also increase both the abundance of HeT-A and TART RNA and their frequency of transposition to broken ends [9]. In contrast, loss of one copy of either of the DNA repair genes Ku70 or Ku80 strongly increased transposition to broken ends but did not increase expression of HeT-A [10] (expression of TART was not reported). Taken together, these observations suggest that there may be different pathways of telomere length regulation that may be specific to different cells and different types of telomere defects.
The work of Savitsky et al. [2] shows clearly that products of spn-E and aub are involved in regulating the expression and transposition of HeT-A and TART. The mechanism of this regulation is not yet determined but, because both spn-E and aub are components of the RNAi-based silencing mechanism [4], it is likely that RNAi is involved. This possibility is supported by the evidence that short (26-29 nucleotide) RNAs with HeT-A and TART sequences are found in wild-type flies and flies heterozygous for a spn-E mutant allele, but are absent in flies carrying two spn-E mutant alleles [2]. Short RNAs of this size have been shown to be involved in transcriptional silencing in plants [12], Caenorhabditis elegans [13] and mammals [14], and in genome rearrangements in Tetrahymena [15], suggesting that RNAi is affecting Drosophila telomeres by acting on the chromosome rather than on an RNA transcript.
Open questions
On their evidence that the regulation of TART is more sensitive to disruptions in the RNAi pathway than is HeT-A regulation, Savitsky et al. [2] suggest that TART is the principal target of RNAi regulation in the germline. This is based on the observation that heterozygous mutations induce much more transposition of TART than of HeT-A. This contrasts with the situation in wild-type flies, however, where HeT-A transpositions to broken ends are much more frequent than TART transpositions [9,10]. In addition, HeT-A is significantly more abundant than TART in the genomes of stocks that have been examined (P.G. DeBaryshe, personal communication). The predominance of HeT-A has always been puzzling, because HeT-A is an exceptional retrotransposon in that it does not encode the Pol protein, which provides enzymatic activities such as reverse transcriptase needed for retrotransposition. TART does encode Pol and the possibility that TART supplies Pol for HeT-A is supported by evidence that HeT-A Gag protein localizes TART Gag to telomeres [16]. The finding that TART is the more sensitive RNAi target opens new avenues to explore the collaboration of HeT-A and TART that is seen in all Drosophila species [17].
One result from the Savitsky et al. study [2] suggests that the collaboration between HeT-A and TART may not be simple. The dramatic increase in HeT-A RNA and the predominance of HeT-A transposition seen in homozygous spn-E1 flies shows a nice correlation between expression and transposition frequency, but it raises questions about the mechanism by which gene dosage can affect the dominance relationship of HeT-A and TART.
One of the unusual features of TART is the production of abundant antisense RNA. This feature is conserved in all Drosophila species, suggesting that the antisense RNA is important for telomere maintenance or regulation [17]. Surprisingly, spn-E and aub mutants did not affect the expression of antisense TART RNA. Because there is a decrease in the levels of short TART RNAs (see above), an increase in TART antisense RNA should be expected. The failure of Sav-itsky et al. [2] to find any effect on the expression of this RNA could indicate that TART antisense RNA makes only a minor contribution to double-stranded RNA production. If so, this would set some limits on antisense functions. Perhaps TART antisense RNA acts in a different pathway of regulation, such as the somatic regulation of telomere length, or in a different aspect of telomere maintenance, such as epigenetic control of the heterochromatic structure of the telomeres.
Transposable elements or telomeres?
HeT-A and TART have split personalities. They have the hallmarks of non-LTR retrotransposons but at the same time these two elements have been dedicated to telomere maintenance throughout the more than 60 million years since the separation of the genus Drosophila [17]. Their regulation by the RNAi machinery could simply be a reflection of their retrotransposon nature. On the other hand, there is recent evidence that RNAi regulates telomere activity in organisms that have telomerase; mutations in the RNAi machinery have been shown to disrupt telomere function in both Schizosaccharomyces pombe [18] and Tetrahymena [19]. Perhaps this is another case where the variant Drosophila telomere is sharing in a general cellular mechanism.
Finally, it is important to keep in mind that, in addition to the effect on transposition onto broken chromosome ends in germline cells demonstrated by Savitsky et al. [2], the RNAi regulation of HeT-A and TART might have other functions yet to be discovered. Mammalian telomerase has other roles, still not completely understood, that are independent of telomere elongation [20]. Once again, studying Drosophila telomeres, apparently so different, might shed light on possibly important features of other eukaryote telomeres.
Acknowledgments
Acknowledgements
This work has been supported by National Institutes of Health Grant GM50315 to M.L.P. We thank M.L Espinàs, D. Huertas and F. Azorín for critical reading of the manuscript.
References
- Pardue ML, DeBaryshe PG. Retrotransposons provide an evolutionary robust non-telomerase mechanism to maintain telomeres. Annu Rev Genet. 2003;37:485–511. doi: 10.1146/annurev.genet.38.072902.093115. [DOI] [PubMed] [Google Scholar]
- Savitsky M, Kwon D, Georgiev P, Kalmykova A, Gvozdev V. Telomere elongation is under the control of the RNAi-based mechanism in the Drosophila germline. Genes Dev. 2006;20:345–354. doi: 10.1101/gad.370206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vagin VV, Klenov MS, Kalmykova AI, Stolyarenko AD, Kotelnikov RN, Gvozdev VA. The RNA interference proteins and the vasa locus are involved in the silencing of retrotransposons in the female germline of Drosophila melanogaster. RNA Biol. 2004;1:54–58. [PubMed] [Google Scholar]
- Kennerdell JR, Yamaguchi S, Carthew RW. RNAi is activated during Drosophila oocyte maturation in a manner dependent on aubergine and spindle-E. Genes Dev. 2002;16:1884–1889. doi: 10.1101/gad.990802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gillispie DE, Berg CA. Homeless is required for RNA localization in Drosophila oogenesis and encodes a new member of the DE-H family of the RNA-dependent ATPases. Genes Dev. 1995;9:2495–2508. doi: 10.1101/gad.9.20.2495. [DOI] [PubMed] [Google Scholar]
- Tomari Y, Du T, Haley B, Schwarz DS, Bennett R, Cook HA, Koppetsch BS, Theurkauf WE, Zamore PD. RISC assembly defects in the Drosophila RNAi mutant armitage. Cell. 2004;116:831–841. doi: 10.1016/S0092-8674(04)00218-1. [DOI] [PubMed] [Google Scholar]
- Abad JP, De Pablos B, Osoegawa K, De Jong PJ, Martin-Gallardo A, Villasante A. TAHRE, a novel telomeric retrotransposon from Drosophila melanogaster, reveals the origin of Drosophila telomeres. Mol Biol Evol. 2004;21:1620–1624. doi: 10.1093/molbev/msh180. [DOI] [PubMed] [Google Scholar]
- George JA, Pardue ML. The promoter of the heterochromatic Drosophila telomeric retrotransposon, HeT-A, is active when moved into euchromatic locations. Genetics. 2003;163:625–635. doi: 10.1093/genetics/163.2.625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Savitsky M, Chravchuk O, Melnikova L, Georgiev P. Heterochromatin protein 1 is involved in control of telomere elongation in Drosophila melanogaster. Mol Cell Biol. 2002;22:3204–3218. doi: 10.1128/MCB.22.9.3204-3218.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melnikova L, Biessmann H, Georgiev P. The Ku protein complex is involved in length regulation of Drosophila telomeres. Genetics. 2005;170:221–235. doi: 10.1534/genetics.104.034538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perrini B, Piacentini L, Fanti L, Altieri F, Chichiarelli S, Berloco M, Turano C, Ferraro A, Pimpinelli S. HP1 controls telomere capping, telomere elongation, and telomere silencing by two different mechanisms in Drosophila. Mol Cell. 2004;15:467–476. doi: 10.1016/j.molcel.2004.06.036. [DOI] [PubMed] [Google Scholar]
- Hamilton A, Voinnet O, Chappell L, Baulcombe D. Two classes of short interfering RNA in RNA silencing. EMBO J. 2002;21:4671–4679. doi: 10.1093/emboj/cdf464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Groishok A. RNAi mechanisms in Caenorhabditis elegans. FEBS Lett. 2005;579:5932–5939. doi: 10.1016/j.febslet.2005.08.001. [DOI] [PubMed] [Google Scholar]
- Bayne EH, Allshire RC. RNA-directed transcriptional gene silencing in mammals. Trends Genet. 2005;21:370–373. doi: 10.1016/j.tig.2005.05.007. [DOI] [PubMed] [Google Scholar]
- Mochizuki K, Fine NA, Fujisawa T, Gorovsky MA. Analysis of a piwi-related gene implicates small RNAs in genome rearrangements in Tetrahymena. Cell. 2002;110:689–699. doi: 10.1016/S0092-8674(02)00909-1. [DOI] [PubMed] [Google Scholar]
- Rashkova S, Karam SE, Kellum R, Pardue ML. Gag proteins of the two Drosophila telomeric retrotransposons are targeted to chromosome ends. J Cell Biol. 2002;159:397–402. doi: 10.1083/jcb.200205039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casacuberta E, Pardue ML. HeT-A and TART, two Drosophila retrotransposons with a bona fide role in chromosome structure for more than 60 million years. Cytogenet Genome Res. 2005;110:152–159. doi: 10.1159/000084947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hall IM, Noma K, Grewal SI. RNA interference machinery regulates chromosome dynamics during mitosis and meiosis in fission yeast. Proc Natl Acad Sci USA. 2003;100:193–198. doi: 10.1073/pnas.232688099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mochizuki K, Gorovsky MA. A Dicer-like protein in Tetrahymena has distinct functions in genome rearrangement, chromosome segregation and meiotic prophase. Genes Dev. 2005;19:77–89. doi: 10.1101/gad.1265105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li S, Rosenberg JE, Donjacour AA, Botchkina IL, Hom YK, Cunha GR, Blackburn EH. Rapid inhibition of cancer cell growth induced by lentiviral delivery and expression of mutant-template telomerase RNA and anti-telomerase short interfering RNA. Cancer Res. 2004;64:4833–4840. doi: 10.1158/0008-5472.CAN-04-0953. [DOI] [PubMed] [Google Scholar]