

Abstract
The neurobiology of female sexual behavior has largely focused on mechanisms of hormone action on nerve cells and how these effects translate into the display of copulatory motor patterns. Of equal importance, though less studied, are some of the consequences of engaging in sexual behavior, including the rewarding properties of sexual interactions and how sexual experience alters copulatory efficiency. This review summarizes the effects of sexual experience on reward processes and copulation in female Syrian hamsters. Neural correlates of these sexual interactions include long-term cellular changes in dopamine transmission and postsynaptic signaling pathways related to neuronal plasticity (e.g., dendritic spine formation). Taken together, these studies suggest that sexual experience enhances the reinforcing properties of sexual behavior, which has the coincident outcome of increasing copulatory efficiency in a way that can increase reproductive success.
Keywords: Copulation, sensitization, dopamine, nucleus accumbens, signaling, plasticity
1. Introduction
“Why do animals mate?” is a simple question that lies at the heart of the neurobiology of female sexual behavior. No behavioral question has a simple answer, as there are both proximate and distal causes and consequences of behavior that raise their own questions and have their own neurobiological answers. Perhaps the most common answer to that question is “To produce offspring”. This may be an answer in the context of a distal consequence of behavior, but even so, such an answer is undoubtedly incorrect [2]. Agmo [2] cites data from Swedes indicating that only about 0.1% of (presumably) heterosexual copulations produce children. Even among species such as rats, in which a high percentage of matings may result in offspring, such a correlation does not imply that pregnancy is an expected consequence of copulation.
One answer to the question of why animals mate is a straightforward view of female sexual behavior as a ‘reflexive’ response to a fluctuating reproductive physiology combined with stimuli from a reproductively competent male. Such investigations of the neurobiology of female sexual behavior were based on the observation that a sequence of ovarian hormone exposure formed a requisite physiological condition for females to respond sexually to a mounting male [70]. For rodents, several days of estradiol exposure are followed by a more transient surge of progesterone that coordinates ovulation and sexual responsiveness in naturally cycling females [22]. The ensuing logic was that identifying brain regions containing receptors for estradiol and progesterone would provide focal points for detailing neural pathways regulating female sexual behavior [70]. Further, the actions of these steroid hormones on nerve cells would offer insight into the cellular and molecular mechanisms mediating the expression of female sexual responsiveness [71]. There is no doubt that this programmatic approach to the study of female sexual behavior has been highly successful, and the details of this neurobiology in terms of circuitry, neurochemistry and gene expression are well established [e.g., 6,71].
Still, there is another aspect regulating the neurobiology of sexual behavior that is concerned with the immediate and long-term consequences of sexual interactions, i.e., the motivational control of sexual behavior and experiential effects on neural plasticity underlying this system. This neurobiology has been reviewed for males, primarily male rats [2]. The goal of this presentation is to examine such plastic changes in females, focusing on our work with female Syrian hamsters. From this work, it is apparent that while the distal consequences of sexual behavior may be towards reproduction, the proximal rationale is to activate motivational systems, which in fact, drive the behavior.
2. Effects of experience on patterns of female sexual behavior
Two species that offer a nice contrast on how social ecology contributes to patterns of sexual behavior are Norway rats and Syrian hamsters. Both species live in burrow systems. Within those burrows, rats have complex social structures consisting of multiple generations of males and females [3], whereas adult hamsters (both male and female) live separately in individual burrows [26].
The social system of rats lends itself to multiple males and females mating simultaneously [51]. Despite this apparently chaotic scheme, female rats are able to control the pattern of sexual interactions with individual males including deciding which male will contribute an ejaculation during this multiple male mating process [51]. Thus, the female rats are active participants in mating and provide an effective means of controlling the pattern of sexual interactions, including mate selection.
The solicitation component of female sexual behavior in rats provides the clearest evidence of the way in which females can control ongoing sexual interactions with males. When male rats approach an estrous female, the female will respond with a stiff-legged locomotor pattern in which she will hop in place (i.e., hopping) or propel herself (i.e., darting) away from the male [20,49]. These solicitations, combined with running from the male, prevent the male from mounting the female until she stops and permits a copulatory contact [49]. It is interesting that females will permit males to mount again sooner following a mount without intromission than if the female received an intromission [20,50]. This regulation of the male’s copulatory behavior by female rats is termed ‘pacing’ and has clear implications for progestation and fertility [20,21]. Paced mating behavior in female rats is hypothesized to be under the control of nucleus accumbens dopamine [4,28,29,32,33,58,84]. On the surface, the complex pattern of pacing by female rats suggests a behavior that could be modified by experience. The limited available data, however, suggest otherwise [19] and the prevailing conclusion [20] is that “...pacing is a stable and inborn component of sexual responsiveness in the female rat” (p. 482).
Given their solitary existence, female hamsters have a very different mating pattern, the information for which has been derived from laboratory studies [e.g., 46], rather than from naturalistic observations. Female (as well as male) hamsters insert occlusions in the main tunnel leading into the burrow system [26]. A female hamster actively recruits males to the burrow by opening that occlusion and laying down a vaginal scent trail leading to the burrow entrance in anticipation of the onset of her behavioral estrus [46]. It is both unknown whether there is mate selection by female hamsters for males or how such mate selection could be accomplished in the wild. Once the male is sequestered in the burrow, the male and female reside together until the female attains the estrous state and mating is initiated [46]. After mating the male is evicted from the female’s burrow [46].
The immobility of the female hamster’s sexual posture contrasts with the overtly active interchange with males during sexual behavior in female rats. Female hamsters quickly assume a rigid stance accompanying lordosis, a posture that can be maintained for upwards of 95% of a 10 min test [15]. While the female is maintaining this position, the male will mount and/or mount with intromission apparently at his own pace. The seemingly obvious conclusion drawn from these observations was that female hamsters, unlike female rats, do not pace the male’s sexual interactions.
Despite the appearance of immobility, female hamsters are in fact quite active participants in mating interactions with males [46]. Noble [62] first noted that female hamsters make active perineal movements in response to perivaginal tactile stimulation from a male hamster, with the female moving her perineum in the direction of the stimulation. The female moves her vagina in the direction of the point of contact of the male’s thrusts to facilitate intravaginal insertion by the male [62]. In fact, application of a topical anesthetic to the female hamster’s perineum dramatically reduces the male hamster’s ability to achieve penile insertion [63].
Taken together, female rats and hamsters differ in the way in which they look to regulate copulation. The difference between female rats and hamsters lies in the ability of these animals to regulate mounting by the male. Female rats can determine whether or not a male will actually mount. Female hamsters do not control the frequency of mounts by the male, but can influence whether or not the male will successfully intromit on a particular mounting attempt. As such, pacing in rats can be readily observed, whereas it is very difficult to quantify perineal movements in female hamsters during mating. As a solution, we took an indirect approach to measuring the female hamster’s role in regulating intromission by the male. We reasoned that if the number of mounts a female hamster receives is determined by the male, but mounts culminating in intromission are limited by the female’s behavior, then the percentage of mounts that include intromission (in the literature termed ‘hit rate’) is actually a measure strongly dependent on the female’s behavior.
To test this proposition, we examined female hamsters that were sexually naive or females that had previously received 6 weekly, 10 minute sexual interactions with males [8]. We then let each female mate with a sexually naive male hamster and recorded the copulatory behavior. Naive males paired with sexually experienced females had a higher hit rate (greater percentage of mounts with intromission) than did naive males tested with naive females (Fig. 1). Further, the same difference in hit rate was observed whether the females were tested 1 or 6 weeks after their last sexual experience test, suggesting a stable learned response.
Figure 1.
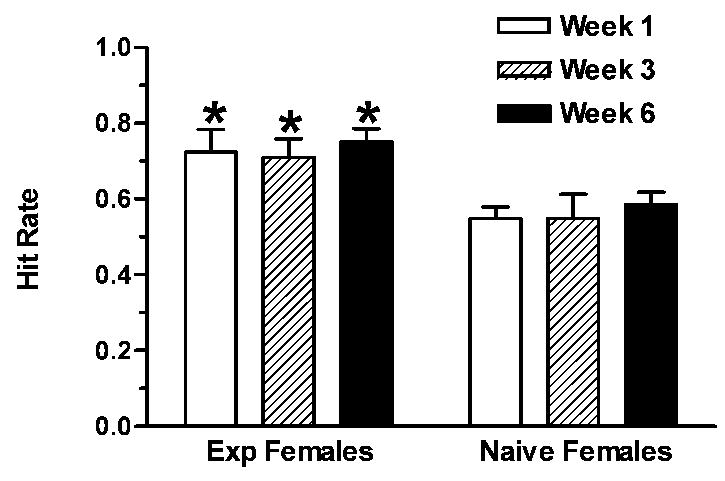
Female hamsters were tested for sexual behavior with a sexually naïve male either 1, 3 or 6 weeks after their last experience test. The hit rate (proportion of mounts culminating in intromission) of the males was higher when mated with sexually-experienced (Exp Females) compared to naïve females (*p<0.05) and this effect of experience persisted even with a 6 week hiatus in behavioral testing. Data from [8].
An additional experiment implicated dopamine in the effects of the female’s sexual experience on the male’s copulatory performance [8]. The dopamine neurotoxin, 6-hydroxydopamine, was injected into the basal forebrain, including the nucleus accumbens, of female hamsters prior to receipt of sexual experience. Naive males tested with these females did not show the elevated hit rate characteristic of mating with experienced females (Figure 2). The impact of the dopamine neurotoxin on sexual interactions was specific to the increment in hit rate associated with sexual experience, as there was no effect of these lesions on the behavior of inexperienced male-female pairings.
Figure 2.
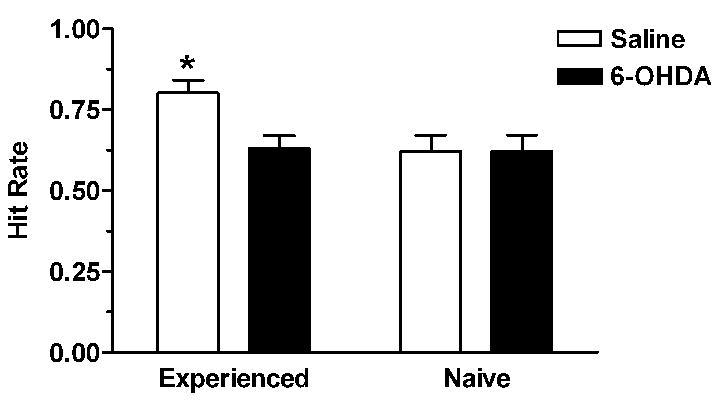
Infusions of the dopamine neurotoxin, 6-hydroxydopamine (6-OHDA), into the region of the nucleus accumbens prior to sexual experience eliminated the effect of the female hamster’s sexual experience on the copulatory efficiency (hit rate) of naïve males (*p<0.05 relative to lesioned females). Data from [8].
3. Sexual experience has rewarding consequences in females
Repeated sexual interactions with males also produce long-term behavioral consequences for the female in the context of reward. Conditioned place preference [14] has been a useful approach to uncovering reinforcing components of sexual behavior. In this paradigm, repeated sexual interactions with a male are associated with one compartment of a multi-compartment chamber. On matched occasions the female is placed alone in a similar but distinctive compartment. Prior to and following these conditioning trials, the female is offered the opportunity to explore the apparatus (in the absence of a male) to determine the relative amount of time the female spends in the compartment associated with copulation. Copulation with the male is operationally defined as reinforcing if the female spends significantly more time in the compartment associated with mating after the sexual behavior trials than before conditioning.
The clear (though perhaps unsurprising) outcome of these studies in female rats [e.g., 65,69] and hamsters [56] is that sexual interactions are reinforcing. The stimulus requirements for this conditioning to occur were not as obvious. For neither rats nor hamsters is the simple display of lordosis during mating tests sufficient to effect conditioned place preference. As noted, female rats have a preferred rate of sexual contacts with the mounting male that has neuroendocrine consequences related to progestation and fertility. Permitting female rats to pace at their preferred interval is necessary for the acquisition of a conditioned place preference, as sexual interactions in which the female does not pace do not produce conditioning [25,27,34,67,68]. The temporal pattern here is important, though not necessarily the control of pacing, as regulating pacing by removing and introducing a male at the female’s preferred interval will also lead to place preference conditioning [34].
Female hamsters do not have a temporal requirement for mating [42], though they also exhibit conditioned place preference to mating [56]. One way in which the importance of sexual contacts by the male for place preference conditioning was tested in female hamsters was to compare the effectiveness of normal sexual interactions with sexual interactions in which intravaginal intromission by the male was prevented by occluding the female’s vagina [39]. Here, place preference conditioning was apparent regardless of whether or not the female received vaginal stimulation during the sexual behavior conditioning trials. This experimental outcome appears to violate the observation that similar vaginal occlusion prevents the elevation in accumbens dopamine during sexual interactions with a male [40]. However, the females were sexually naive in that microdialysis study. It would appear that the multitude of sensory properties accrued during sexual experience, for example during the conditioning trials of a place preference paradigm [39], broadens the sensory stimuli contributing to sexual reward from the restricted role of vaginal stimulation in sexually naive females [40].
There has been little investigation of neurotransmitter systems mediating place preference conditioning to sexual interactions. In one study, antagonizing opioid neurotransmission by treating female rats with naloxone prior to sexual interactions eliminated place preference conditioning [68]. Conversely, several studies using dopamine receptor antagonists have produced mixed results. Pretreating female hamsters with dopamine D2 receptor antagonists [57] prevented the acquisition of a conditioned place preference to sexual interactions (Fig. 3). A similar study in rats produced no effect [30].
Figure 3.
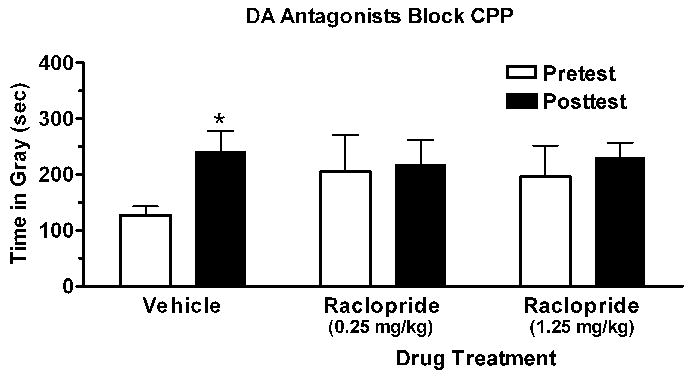
Repeated pairings of copulation with the gray compartment in a conditioned place preference (CPP) apparatus resulted in the female hamsters spending more time in that compartment in the absence of copulation (*p<0.05). This effect was blocked by systemic administration of the dopamine (DA) D2 receptor antagonist, raclopride, prior to each sexual behavior test. Data from [57].
4. Neurotransmitter and cellular plasticity following sexual experience in females
There is a rich tradition of research into mechanisms of dopamine signaling as they relate to components of motivated behaviors and drug abuse [e.g., 60]. Borrowing from that literature, we explored the possibility that sexual experience could affect dopamine neurotransmission in the mesolimbic pathway and that plasticity in that system was the basis for the behavioral consequences of sexual experience, e.g., changes in copulatory efficiency and reward. Within the mesolimbic dopamine system there is both evidence for activation during female sexual interactions, as well as long term effects on structural and neurochemical plasticity. Initial microdialysis experiments demonstrated that extracellular dopamine levels in the nucleus accumbens of females were elevated during mating [55,58]. For female rats, dopamine release was particularly sensitive to paced mating interactions with males [4,33,58], and for (at least sexually naive) female hamsters, dopamine elevations depended on vaginal stimulation received during mating [40]. In a follow-up experiment we took a slightly different approach, this time measuring extracellular dopamine in the nucleus accumbens during mating in sexually naive female hamsters or in females that had sexual experience prior to the microdialysis test [38]. Sexual experience produced an exaggerated increase in extracellular dopamine that persisted throughout the sexual interaction with a male, compared with the dopamine levels in sexually naive females (Figure 4). Perhaps the increased dopamine response in sexually experienced females reflects the enriched array of mating associated stimuli to which female hamsters become responsive as a result of that experience.
Figure 4.
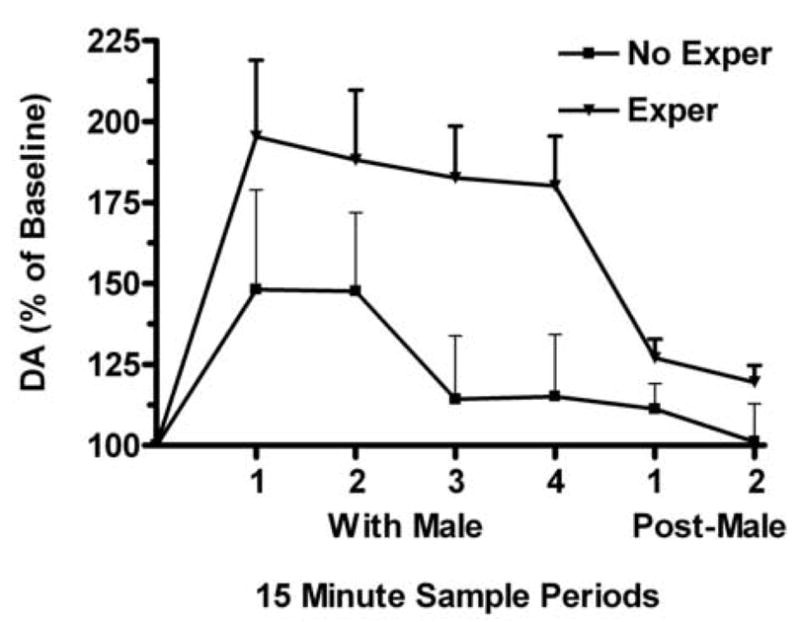
Sexually experienced (Exper) or inexperienced female (No Exper) hamsters were implanted with microdialysis probes in the nucleus accumbens and the females were placed with a male for 1 hr. Samples were taken every 15 min during the 1 hr sexual behavior test. Dopamine levels in the microdialysis samples were elevated significantly during the first two sampling periods in No Exper females. Across all sampling periods, dopamine levels were significantly higher in Exper females compared with No Exper females. Data from [38].
The elevation in dopamine release in experienced female hamsters is reminiscent of the effects of repeated exposure of animals to drugs of abuse [75]. In this literature, the heightened level of dopamine in response to a fixed dose of drug is termed “sensitization” [75]. Drug sensitization is accompanied by a variety of cellular responses thought to enhance synaptic efficacy and information flow through the mesolimbic pathway [74].
One entry point into the mechanism through which behavioral experience could alter neuronal plasticity is at the level of synapses. An indirect approach to this question has been taken by measuring dendritic changes in striatal (including the nucleus accumbens) neurons in response to drug administration or following behavioral experience. Repeated administration of a variety of abused substances with different pharmacological profiles will increase dendritic length and/or spine density in the terminal dendritic branches of medium spiny neurons [13,23,44,45,64,76,77,78]. Far fewer examples exist for behavioral experience producing comparable effects on dendrites, though induction of salt appetite [79], male sexual behavior [24] and female sexual behavior [59] will alter dendritic morphology in medium spiny neurons of the nucleus accumbens.
Sexual experience in female hamsters had a differential impact on dendritic spine density [59] depending on the region examined (Fig. 5). In this experiment, female hamsters were given our basic paradigm of 6 weeks of sexual experience or remained sexually naive [38]. On the 7th week, all females were given an estradiol and progesterone priming regimen and sacrificed about 4 hr after progesterone injection. Brains were processed for Golgi staining and 240 μm slices analyzed. Spines were counted from terminal dendritic branches of pyramidal neurons in the medial prefrontal cortex, medium spiny neurons of the nucleus accumbens (shell and core combined), or medium spiny neurons of the dorsal caudate. Within the medium spiny neurons of the nucleus accumbens, dendritic spine density (normalized to 10 μm of dendritic length) was higher in sexually experienced, than in sexually naive, females. The converse was found in the apical dendrites of layer V neurons of the prefrontal cortex. There were no group differences in spine density in caudate medium spiny neurons. We interpret these differences in spine density as reflecting plasticity in excitatory neurotransmission on dopaminergic-responsive neurons [37].
Figure 5.
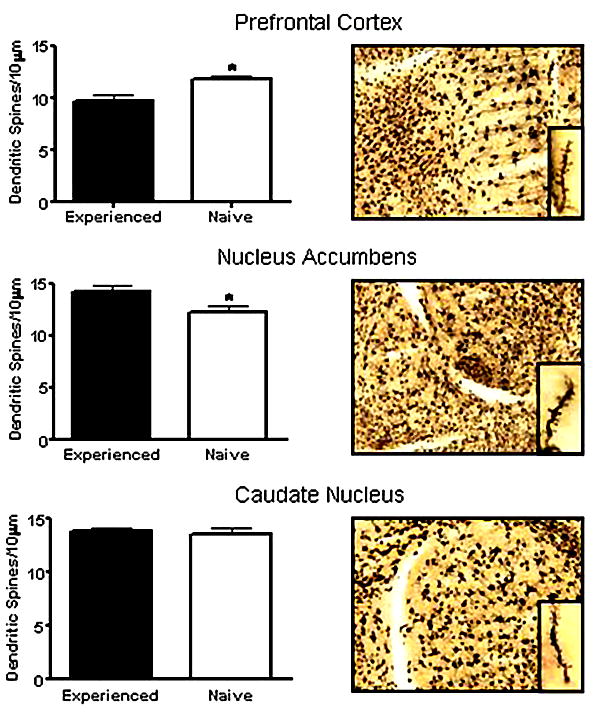
Spine densities (normalized per 10 μm) were measured in the terminal dendrites of neurons (examples of Golgi staining are presented in the right panel) from the prefrontal cortex, nucleus accumbens or caudate nucleus of sexually experienced (n=7) or naïve (n=5) female hamsters. In the prefrontal cortex there was a significant reduction in spine density with experience (*p <0.02), with a corresponding significant increase in spine density in the nucleus accumbens (*p < 0.04).
If we take plasticity in dendritic spines as a distal cellular marker of sexual experience, we can hypothesize a cascade of cellular events that are triggered by repeated sexual interactions. In other words, the focus should be on two of the classes of responses illustrated by treatment with drugs of abuse [36], i.e., an exaggerated response to sexual behavior and altered cellular responses in the absence of sexual behavior. The proposed signaling events are depicted in Fig. 6. This proposal is neither novel nor radical, as dendritic plasticity arising from stimuli as diverse as steroid hormones [54], drugs of abuse [61], or long term potentiation [1] all involve the illustrated events. It is because these pathways are so well represented in varied examples of neural plasticity that it seems likely that as the gaps are filled in the same will be true of sexual behavior effects on the nucleus accumbens.
Figure 6.
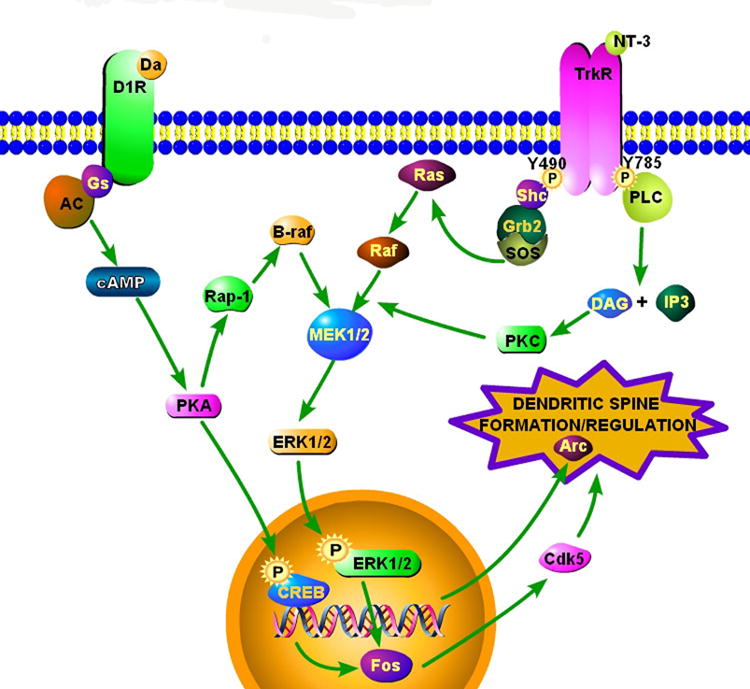
Schematic diagram of some signaling pathways that could mediate long-term changes in cellular plasticity as a function of sexual experience. Our microarray analyses [7] pointed to several nodes in these pathways that were altered by prior sexual experience. Neural signals affecting long-term plasticity could arise from the activation of dopamine D1 receptors, tyrosine kinase receptors (TrkR) or inositol triphosphate (IP3). (Potential signaling from ionotropic glutamate receptors is omitted here.) Activation of these receptors trips a cascade of signaling mechanisms that converge on gene transcription via CREB or MAP kinase mediated processes [31]. For example, G-protein coupled receptors, such as dopamine receptors, can phosphorylate CREB through cAMP formation and PKA activity [47] or indirectly phosphorylate ERK through activation of the MAP kinase pathway [48]. Stimulation of Trk receptors, for example by neurotrophin-3 (NT-3) or BDNF (not shown), or IP3 activity will also stimulate MAP kinase pathways, though through PKC activation [72,82]. What are illustrated as individual pathways and have been isolated in cell systems, are actually highly integrated molecular pathways in intact neurons [35]. In turn, nuclear transcription factors are expressed (e.g., c-Fos and ΔFosB [43,53,73]) that regulate molecular mediators of dendritic plasticity (e.g., cyclin-dependent kinase 5; Cdk5, or activity-regulated cytoskeleton-associated protein; Arc) [5,11,41,81]. This figure was made using Pathway Builder v.2.0 (Protein Lounge, San Diego).
The discovery approach, utilizing gene microarrays [7], along with experimental approaches have begun to validate altered activity or protein expression at several points in these pathways resulting from sexual experience. Transcription factors represent one set of molecular events that can impact dendritic structure leading to long-term plasticity [5,17,52]. Both c-Fos and FosB staining were examined in response to sexual experience and mating in female Syrian hamsters. Following sexual interactions with a male, c-Fos staining was elevated in the core of the nucleus accumbens, a response that was magnified in sexually-experienced females (Fig. 7) [9]. FosB staining was not detectibly affected by a sexual interaction, though the levels of staining were higher in the core of the nucleus accumbens in sexually-experienced female hamsters compared with naïve females (Fig. 8). Neither c-Fos nor FosB were affected by sexual behavior or sexual experience in either the shell of the nucleus accumbens or dorsal striatum in these females. In our experiments, changes in c-Fos and FosB occur in parallel, both regionally and as a function of experience, though in other studies alterations in these proteins do not always covary [e.g., 12].
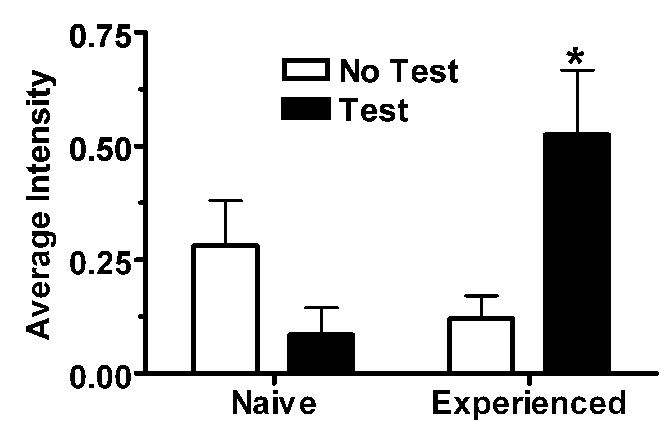
Figure 7.
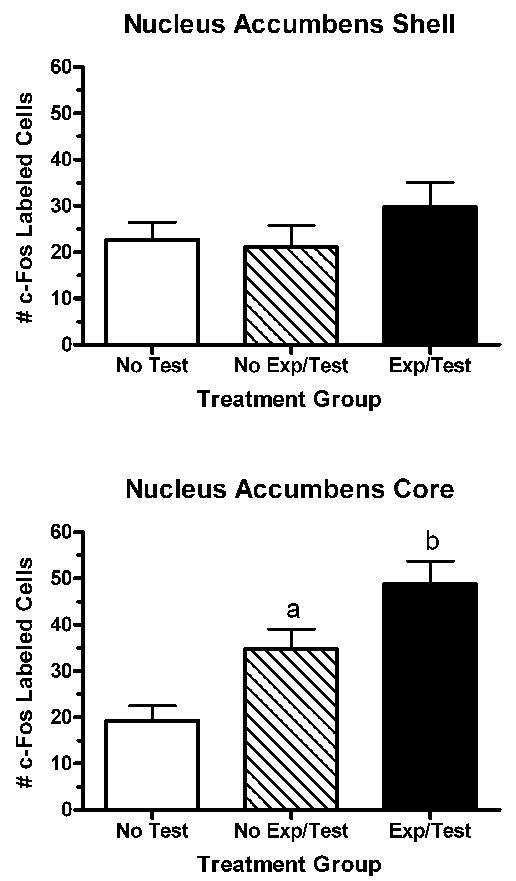
Sexual behavior testing (Test) significantly increases c-Fos staining (ap<0.05 vs. No Test) in the core of the nucleus accumbens of female hamsters, an effect that is magnified in sexually experienced (Exp) females (bp<0.05 vs. no Exp/Test). These effects were not observed in the shell of the nucleus accumbens. Data from [9].
Figure 8.
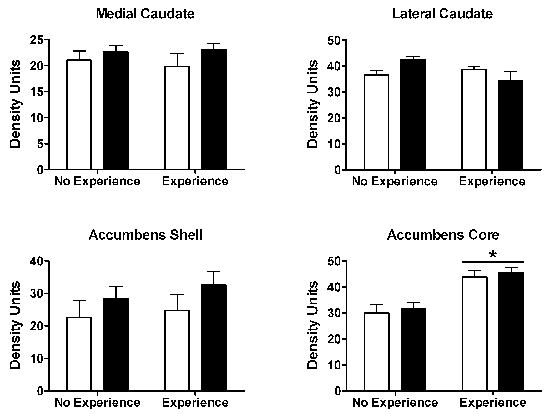
Female hamsters received our standard paradigm of 6 weekly, 10 min sexual behavior tests or were hormone primed, but not tested. On the 7th week, these groups were subdivided, so that half the animals were given a sex behavior test or remained in their home cages. Animals were sacrificed 90 min after testing or the equivalent for non-tested females. Frozen 40 μm brain sections were stained using an antibody (Santa Cruz) that recognizes members of the FosB family. Prior sexual experience, independent of recent testing (dark bars), elevated staining for FosB in the core of the nucleus accumbens (*p<0.05) vs. No Experience females). No effects of either sexual experience or sexual behavior testing were seen in the shell of the nucleus accumbens or in the caudate nucleus.
Fos proteins can be activated through several signaling pathways, including MAP kinase [18]. ERK is a downstream kinase in this pathway and we examined the regulation of ERK following sexual behavior (Fig. 9). In Western blots, total ERK 2 levels were not affected by either sexual behavior or sexual experience. In contrast, pERK 2 was elevated in the nucleus accumbens following sexual behavior, but only in females with prior sexual experience.
Figure 9.
Levels of ERK1/2 were measured by Western blot from punches of the nucleus accumbens and caudate nucleus of female hamsters. Tissue punches (2 mm diameter) from the nucleus accumbens (both core and shell) and caudate nucleus were taken from 2 mm slices encompassing the nucleus accumbens (n = 4/treatment group). The tissue homogenates were subdivided to blot for total ERK1/2 and for pERK1/2. ERK1 and ERK 2 (both total and phosphorylated) were analyzed independently from each Western blot. Sexual behavior testing elevated levels of pERK2 only in the nucleus accumbens of sexually-experienced females (*p<0.05). No significant differences were found for pERK1 or for total ERK1 or ERK2 in either the nucleus accumbens or in the caudate.
Entry into the MAP kinase pathway can come from several sources, including glutamate receptor activation [1], G-protein coupled receptors (e.g., dopamine receptors) [83], inositol triphosphate pathways [66], and through growth factor receptors [16]. Sexual experience effects on these pathways have been implicated through microarray analyses [7], but have not really been examined directly. One mechanism that in fact is regulated by sexual experience is dopamine receptor coupling to adenylate cyclase [10]. Homogenates from the nucleus accumbens were taken from sexually experienced or inexperienced female hamsters. These homogenates were stimulated with dopamine and cAMP accumulation measured (Fig. 10). Dopamine stimulated cAMP accumulation in all treatment groups, with greater stimulation in homogenates from the sexually experienced females. These actions of dopamine were determined to be D1 receptor mediated. Though one component of plasticity following sexual experience is presynaptic (i.e., increased dopamine efflux during sexual interactions), it is just as clear that there are postsynaptic modifications that are not simply reflections of increased synaptic dopamine levels.
Fig 10.
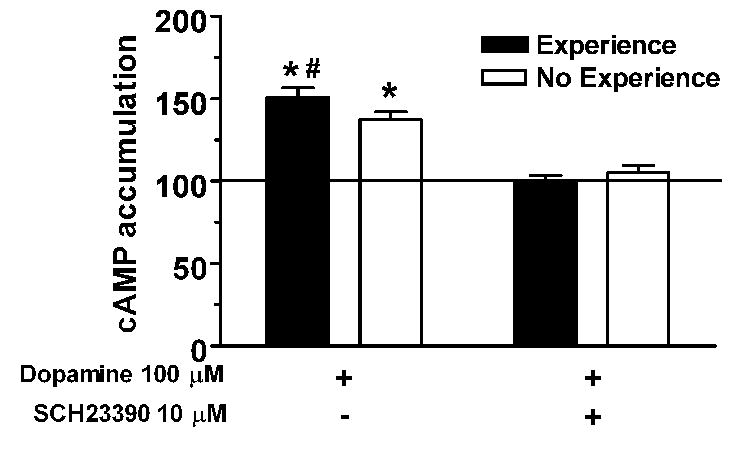
Homogenates from the nucleus accumbens of female hamsters receiving either sexual experience or no experience were measured for cAMP accumulation following dopamine stimulation (data are % no-dopamine baseline). Dopamine elevated cAMP production (* p<0.05 vs. baseline), with a greater effect in membranes from sexually experienced females (# vs. No Experience females). The increase in dopamine stimulated cAMP accumulation was eliminated by concurrent treatment with SCH23390 indicating that dopamine was acting via dopamine D1 receptors. Data from [10].
5. Summary and conclusion
One hypothesis of mesolimbic dopamine function is that this pathway is sensitive to the conditioned properties associated with naturally occurring behaviors in a way that optimizes the functional consequences of those behaviors [80]. From this framework we can conceive of a pattern of behavior in which vaginal stimulation received by females during copulation stimulates dopamine neurotransmission. Though initially this response is unconditioned [55], with experience females learn to produce subtle perineal movements that increase the likelihood of receiving vaginal stimulation from the mounting male [8]. In turn, there is greater dopamine activation, which feeds forward to maintain the behavioral responding. Because receipt of vaginal stimulation through intromissions from the mounting male (preceding ejaculation by the male) is necessary for induction of the progestational state accompanying fertilization (and therefore successful pregnancy) [42], this behavioral regulation would have the indirect effect of increasing copulatory efficiency leading to reproductive success. The answer to the question of “Why do females mate?” is to receive stimulation that has rewarding consequences in the form of forebrain dopamine activity. These ‘pleasurable’ components of sexual behavior have the unanticipated (from the female’s point of view), though highly adaptive, consequences of successful pregnancy and the birth of offspring.
Acknowledgments
We would like to thank a number of people who have contributed importantly to this research including Dr. Katherine Bradley, Alma Haas, Margaret Joppa, Dr. Jess Kohlert, Richard Rowe and Dr. Val Watts. Special thanks to Paul Mermelstein for his advice and sustained interest in our work. This review is based on a talk given at the 2006 Workshop on Steroid Hormones and Brain Function, Breckenridge, Co. We are grateful to The National Science Foundation (IBN-9412543 and IBN-9723876) and National Institutes of Health (DA13680) for their support of this research.
References
- 1.Adams JP, Roberson ED, English JD, Selcher JC, Sweatt JD. MAPK regulation of gene expression in the central nervous system. Acta Neurobiol Exp (Wars) 2000;60:377–394. doi: 10.55782/ane-2000-1357. [DOI] [PubMed] [Google Scholar]
- 2.Ågmo A. Sexual motivation - an inquiry into events determining the occurrence of sexual behavior. Behav Brain Res. 1999;105:129–150. doi: 10.1016/s0166-4328(99)00088-1. [DOI] [PubMed] [Google Scholar]
- 3.Barnett SA. The Rat: A Study in Behaviour. Aldine; Chicago: 1963. [Google Scholar]
- 4.Becker JB, Rudick CN, Jenkins WJ. The role of dopamine in the nucleus accumbens and striatum during sexual behavior in the female rat. J Neurosci. 2001;21:3236–3241. doi: 10.1523/JNEUROSCI.21-09-03236.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bibb JA. Role of Cdk5 in neuronal signaling, plasticity, and drug abuse. Neuro-Signals. 2003;12:191–199. doi: 10.1159/000074620. [DOI] [PubMed] [Google Scholar]
- 6.Blaustein JD, Erskine MS. Feminine sexual behavior: Cellular integration of hormonal and afferent information in the rodent forebrain. In: Pfaff DW, Arnold AP, Etgen AM, Fahrbach SE, Rubin RT, editors. Hormones Brain and Behavior. Vol. 1. Academic Press; Amsterdam: 2002. pp. 139–214. [Google Scholar]
- 7.Bradley KC, Boulware MB, Jiang H, Doerge RW, Meisel RL, Mermelstein PG. Sexual experience generates distinct patterns of gene expression within the nucleus accumbens and dorsal striatum of female Syrian hamsters. Genes Brain Behav. 2005;4:31–44. doi: 10.1111/j.1601-183X.2004.00093.x. [DOI] [PubMed] [Google Scholar]
- 8.Bradley KC, Haas AR, Meisel RL. 6-Hydroxydopamine lesions in female hamsters (Mesocricetus auratus) abolish the sensitized effects of sexual experience on copulatory interactions with males. Behav Neurosci. 2005;119:224–232. doi: 10.1037/0735-7044.119.1.224. [DOI] [PubMed] [Google Scholar]
- 9.Bradley KC, Meisel RL. Sexual behavior induction of c-Fos in the nucleus accumbens and amphetamine-stimulated locomotor activity are sensitized by previous sexual experience in female Syrian hamsters. J Neurosci. 2001;21:2123–2130. doi: 10.1523/JNEUROSCI.21-06-02123.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bradley KC, Mullins AJ, Meisel RL, Watts VJ. Sexual experience alters dopamine D1 receptor mediated cyclic AMP production in the nucleus accumbens of female Syrian hamsters. Synapse. 2004;53:20–27. doi: 10.1002/syn.20030. [DOI] [PubMed] [Google Scholar]
- 11.Bramham CR, Messaoudi E. BDNF function in adult synaptic plasticity: The synaptic consolidation hypothesis. Prog Neurobiol. 2005;76:99–125. doi: 10.1016/j.pneurobio.2005.06.003. [DOI] [PubMed] [Google Scholar]
- 12.Brenhouse HC, Stellar JR. c-Fos and ΔFosB expression are differentially altered in distinct subregions of the nucleus accumbens shell in cocaine-sensitized rats. Neuroscience. 2006;137:773–780. doi: 10.1016/j.neuroscience.2005.09.039. [DOI] [PubMed] [Google Scholar]
- 13.Brown RW, Kolb B. Nicotine sensitization increases dendritic length and spine density in the nucleus accumbens and cingulate cortex. Brain Res. 2001;899:94–100. doi: 10.1016/s0006-8993(01)02201-6. [DOI] [PubMed] [Google Scholar]
- 14.Carr GD, Fibiger HC, Phillips AG. Conditioned place preference as a measure of drug reward. In: Leibman JM, Cooper SJ, editors. The Neuropharmacological Basis of Reward. Clarendon Press; Oxford: 1989. pp. 264–319. [Google Scholar]
- 15.Carter CS. Postcopulatory sexual receptivity in the female hamster: the role of the ovary and the adrenal. Horm Behav. 1972;3:261–265. doi: 10.1016/0018-506x(72)90039-6. [DOI] [PubMed] [Google Scholar]
- 16.Chao MV. Neurotrophins and their receptors: A convergence point for many signaling pathways. Nat Rev Neurosci. 2003;4:299–309. doi: 10.1038/nrn1078. [DOI] [PubMed] [Google Scholar]
- 17.Cheung ZH, Fu AKY, Ip NY. Synaptic roles of Cdk5; Implications in higher cognitive function and neurodegenerative diseases. Neuron. 2006;50:13–18. doi: 10.1016/j.neuron.2006.02.024. [DOI] [PubMed] [Google Scholar]
- 18.Davis RJ. Transcriptional regulation by MAP kinases. Mol Reprod Dev. 1995;42:459–467. doi: 10.1002/mrd.1080420414. [DOI] [PubMed] [Google Scholar]
- 19.Erskine MS. Effects of paced coital stimulation on estrus duration in intact cycling rats and ovariectomized and ovariectomized-adrenalectomized hormone-primed rats. Behav Neurosci. 1985;99:151–161. doi: 10.1037//0735-7044.99.1.151. [DOI] [PubMed] [Google Scholar]
- 20.Erskine MS. Solicitation behavior in the estrous female rat: A review. Horm Behav. 1989;23:473–502. doi: 10.1016/0018-506x(89)90037-8. [DOI] [PubMed] [Google Scholar]
- 21.Erskine MS, Kornberg E, Cherry JA. Paced copulation in rats: Effects of intromission frequency and duration on luteal activation and estrus length. Physiol Behav. 1989;45:33–39. doi: 10.1016/0031-9384(89)90163-7. [DOI] [PubMed] [Google Scholar]
- 22.Feder HH. Estrous cyclicity in mammals. In: Adler NT, editor. Neuroendocrinology of Reproduction. Plenum Press; New York: pp. 279–348. [Google Scholar]
- 23.Ferrario CR, Gorny G, Crombag HS, Li Y, Kolb B, Robinson TE. Neural and behavioral plasticity associated with the transition from controlled to escalated cocaine use. Biol Psychiatry. 2005;58:751–759. doi: 10.1016/j.biopsych.2005.04.046. [DOI] [PubMed] [Google Scholar]
- 24.Fiorino DF, Kolb BS. Society for Neuroscience. New Orleans, LA, 2003 Abstract Viewer and Itinerary Planner; Washington, DC: 2003. Sexual experience leads to long-lasting morphological changes in male rat prefrontal cortex, parietal cortex, and nucleus accumbens neurons. [Google Scholar]
- 25.Gans S, Erskine MS. Effects of neonatal testosterone treatment on pacing behaviors and development of a conditioned place preference. Horm Behav. 2003;44:354–364. doi: 10.1016/s0018-506x(03)00157-0. [DOI] [PubMed] [Google Scholar]
- 26.Gattermann R, Fritzsche P, Neumann K, Kayser A, Abiad M, Yaku R. Notes on the current distribution and the ecology of wild golden hamsters (Mesocricetus auratus) J Zool Lond. 2001;254:359–365. [Google Scholar]
- 27.González-Florez O, Camacho FJ, Dominguez-Salazar E, Ramírez-Orduna JM, Beyer C, Paredes RG. Progestins and place preference conditioning after paced mating. Horm Behav. 2004;46:151–157. doi: 10.1016/j.yhbeh.2004.02.006. [DOI] [PubMed] [Google Scholar]
- 28.Guarraci FA, Megroz AB, Clark AS. Effects of ibotenic acid lesions of the nucleus accumbens on paced mating behavior in the female rat. Behav Neurosci. 2002;116:568–576. doi: 10.1037//0735-7044.116.4.568. [DOI] [PubMed] [Google Scholar]
- 29.Guarraci FA, Megroz AB, Clark AS. Paced mating behavior in the female rat following lesions of three regions responsive to vaginocervical stimulation. Brain Res. 2004;999:40–52. doi: 10.1016/j.brainres.2003.10.056. [DOI] [PubMed] [Google Scholar]
- 30.Horsman PG, Paredes RG. Dopamine antagonists do not block conditioned place preference induced by paced mating behavior in female rats. Behav Neurosci. 2004;118:356–364. doi: 10.1037/0735-7044.118.2.356. [DOI] [PubMed] [Google Scholar]
- 31.Hyman SE, Malenka RC. Addiction and the brain: The neurobiology of compulsion and its persistence. Nat Rev Neurosci. 2001;2:695–703. doi: 10.1038/35094560. [DOI] [PubMed] [Google Scholar]
- 32.Jenkins WJ, Becker JB. Role of the striatum and nucleus accumbens in paced copulatory behavior in the female rat. Behav Brain Res. 2001;121:119–128. doi: 10.1016/s0166-4328(00)00394-6. [DOI] [PubMed] [Google Scholar]
- 33.Jenkins WJ, Becker JB. Dynamic increases in dopamine during paced copulation in the female rat. Eur J Neurosci. 2003;18:1997–2001. doi: 10.1046/j.1460-9568.2003.02923.x. [DOI] [PubMed] [Google Scholar]
- 34.Jenkins WJ, Becker JB. Female rats develop conditioned place preference for sex at their preferred interval. Horm Behav. 2003;43:503–507. doi: 10.1016/s0018-506x(03)00031-x. [DOI] [PubMed] [Google Scholar]
- 35.Ji Y, Pang PT, Feng L, Lu B. Cyclic AMP controls BDNF-induced TrkB phosphorylation and dendritic spine formation in mature hippocampal neurons. Nat Neruosci. 2005;8:164–172. doi: 10.1038/nn1381. [DOI] [PubMed] [Google Scholar]
- 36.Kalivas PW, Toda S, Bowers MS, Baker DA, Ghasemzadeh MB. The temporal sequence of changes in gene expression by drugs of abuse. In: Wang JQ, editor. Methods in Molecular Medicine: Drugs of Abuse: Neurological Reviews and Protocols. Vol. 79. Humana Press; Totowa, NJ: 2003. pp. 3–11. [DOI] [PubMed] [Google Scholar]
- 37.Kalivas PW, Volkow N, Seamans J. Unmanageable motivation in addiction: A pathology in prefrontal-accumbens glutamate transmission. Neuron. 2005;45:647–650. doi: 10.1016/j.neuron.2005.02.005. [DOI] [PubMed] [Google Scholar]
- 38.Kohlert JG, Meisel RL. Sexual experience sensitizes mating-related nucleus accumbens dopamine responses of female Syrian hamsters. Behav Brain Res. 1999;99:45–52. doi: 10.1016/s0166-4328(98)00068-0. [DOI] [PubMed] [Google Scholar]
- 39.Kohlert JG, Olexa N. The role of vaginal stimulation for the acquisition of conditioned place preference in female Syrian hamsters. Physiol Behav. 2005;84:135–139. doi: 10.1016/j.physbeh.2004.10.020. [DOI] [PubMed] [Google Scholar]
- 40.Kohlert JG, Rowe RK, Meisel RL. Intromissive stimulation from the male increases extracellular dopamine release from fluoro-gold-identified neurons within the midbrain of female hamsters. Horm Behav. 1997;32:143–154. doi: 10.1006/hbeh.1997.1415. [DOI] [PubMed] [Google Scholar]
- 41.Kumar V, Zhang MX, Swank MW, Kunz J, Wu GY. Regulation of dendritic morphogenesis by Ras-PI3K-Akt-mTOR and Ras-MAPK signaling pathways. J Neurosci. 2005;25:11288–11299. doi: 10.1523/JNEUROSCI.2284-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Lanier DL, Estep DQ, Dewsbury DA. Copulatory behavior of golden hamsters: Effects on pregnancy. Physiol Behav. 1975;15:209–212. doi: 10.1016/0031-9384(75)90237-1. [DOI] [PubMed] [Google Scholar]
- 43.Lee KW, Kim Y, Kim AM, Helmin K, Nairn AC, Greengard P. cocaine-induced dendritic spine formation in D1 and D2 dopamine receptor-containing medium spiny neurons in nucleus accumbens. Proc Natl Acad Sci USA. 2006;103:3399–3404. doi: 10.1073/pnas.0511244103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Li Y, Acerbo MJ, Robinson TE. The induction of behavioural sensitization is associated with cocaine-induced structural plasticity in the core (but not shell) of the nucleus accumbens. Eur J Neurosci. 2004;20:1647–1654. doi: 10.1111/j.1460-9568.2004.03612.x. [DOI] [PubMed] [Google Scholar]
- 45.Li Y, Kolb B, Robinson TE. The location of persistent amphetamine-induced changes in the density of dendritic spines on medium spiny neurons in the nucleus accumbens and caudate-putamen. Neuropsychopharmacol. 2003;28:1082–1085. doi: 10.1038/sj.npp.1300115. [DOI] [PubMed] [Google Scholar]
- 46.Lisk RD, Ciaccio LA, Catanzaro C. Mating behaviour of the golden hamster under seminatural conditions. Anim Behav. 1983;31:659–666. [Google Scholar]
- 47.Lonze BE, Ginty DD. Function and regulation of CREB family transcription factors in the nervous system. Neuron. 2002;35:605–623. doi: 10.1016/s0896-6273(02)00828-0. [DOI] [PubMed] [Google Scholar]
- 48.Marinissen MJ, Gutkind JS. G-protein-coupled receptors and signaling networks: emerging paradigms. Trends Pharmacol Sci. 2001;22:368–376. doi: 10.1016/s0165-6147(00)01678-3. [DOI] [PubMed] [Google Scholar]
- 49.McClintock MK, Adler NT. The role of the female during copulation in wild and domestic Norway rats (Rattus norvegicus) Behaviour. 1978;67:67–96. [Google Scholar]
- 50.McClintock MK, Anisko JJ. Group mating among Norway rats I. Sex differences in the pattern and neuroendocrine consequences of copulation. Anim Behav. 1982;30:398–409. [Google Scholar]
- 51.McClintock MK, Anisko JJ, Adler NT. Group mating among Norway rats II. The social dynamics of copulation: competition, cooperation, and mate choice. Anim Behav. 1982;30:410–425. [Google Scholar]
- 52.McClung CA, Nestler EJ. Regulation of gene expression and cocaine reward by CREB and ΔFosB. Nat Neurosci. 2003;6:1208–1215. doi: 10.1038/nn1143. [DOI] [PubMed] [Google Scholar]
- 53.McClung CA, Ulery PG, Perrotti LI, Zachariou V, Berton O, Nestler EJ. ΔFosB: a molecular switch for long-term adaptation in the brain. Mol Brain Res. 2004;132:146–154. doi: 10.1016/j.molbrainres.2004.05.014. [DOI] [PubMed] [Google Scholar]
- 54.McEwen BS. Estrogen effects on the brain: multiple sites and molecular mechanisms. J Appl Physiol. 2001;91:2785–2801. doi: 10.1152/jappl.2001.91.6.2785. [DOI] [PubMed] [Google Scholar]
- 55.Meisel RL, Camp DM, Robinson TE. A microdialysis study of ventral striatal dopamine during sexual behavior in female Syrian hamsters. Behav Brain Res. 1993;55:151–157. doi: 10.1016/0166-4328(93)90111-3. [DOI] [PubMed] [Google Scholar]
- 56.Meisel RL, Joppa MA. Conditioned place preference in female hamsters following aggressive or sexual encounters. Physiol Behav. 1994;56:1115–1118. doi: 10.1016/0031-9384(94)90352-2. [DOI] [PubMed] [Google Scholar]
- 57.Meisel RL, Joppa MA, Rowe RK. Dopamine receptor antagonists attenuate conditioned place preference following sexual behavior in female Syrian hamsters. Eur J Pharmacol. 1996;309:21–24. doi: 10.1016/0014-2999(96)00389-5. [DOI] [PubMed] [Google Scholar]
- 58.Mermelstein PG, Becker JB. Increased extracellular dopamine in the nucleus accumbens and striatum of the female rat during paced copulatory behavior. Behav Neurosci. 1995;109:354–365. doi: 10.1037//0735-7044.109.2.354. [DOI] [PubMed] [Google Scholar]
- 59.Mullins AJ, Sengelaub DR, Meisel RL. Society for Neuroscience. San Diego: 2004 Abstract Viewer and ItineraryPlanner; Washington, DC: 2004. Effects of sexual experience in female hamsters on MAP kinase signaling and dendritic morphology. [Google Scholar]
- 60.Nestler EJ. Molecular basis of long-term plasticity underlying addiction. Nat Rev Neurosci. 2001;2:119–128. doi: 10.1038/35053570. [DOI] [PubMed] [Google Scholar]
- 61.Nestler EJ. Molecular mechanisms of drug addiction. Neuropharmacol. 2004;47:24–32. doi: 10.1016/j.neuropharm.2004.06.031. [DOI] [PubMed] [Google Scholar]
- 62.Noble RG. The sexual responses of the female hamster: A descriptive analysis. Physiol Behav. 1979;23:1001–1005. doi: 10.1016/0031-9384(79)90288-9. [DOI] [PubMed] [Google Scholar]
- 63.Noble RG. Sexual responses of the female hamster: Effects on male performance. Physiol Behav. 1980;24:237–242. doi: 10.1016/0031-9384(80)90080-3. [DOI] [PubMed] [Google Scholar]
- 64.Norrholm SD, Bibb JA, Nestler EJ, Ouimet CC, Taylor JR, Greengard P. Cocaine-induced proliferation of dendritic spines in nucleus accumbens is dependent on the activity of cyclin-dependent kinase-5. Neuroscience. 2003;116:19–22. doi: 10.1016/s0306-4522(02)00560-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Oldenberger WP, Everitt BJ, De Jonge FH. Conditioned place preference induced by sexual interaction in female rats. Horm Behav. 1992;26:214–228. doi: 10.1016/0018-506x(92)90043-u. [DOI] [PubMed] [Google Scholar]
- 66.Opazo P, Watabe AM, Grant SGN, Odell TJ. Phosphatidylinositol 3-kinase regulates the induction of long-term potentiation through extracellular signal-related kinase-independent mechanisms. J Neurosci. 2003;23:3679–3688. doi: 10.1523/JNEUROSCI.23-09-03679.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Paredes RG, Alonso A. Sexual behavior regulated (paced) by the female induces conditioned place preference. Behav Neurosci. 1997;111:123–128. doi: 10.1037//0735-7044.111.1.123. [DOI] [PubMed] [Google Scholar]
- 68.Paredes RG, Martínez I. Naloxone blocks place preference conditioning after paced mating in female rats. Behav Neurosci. 2001;115:1363–1367. [PubMed] [Google Scholar]
- 69.Paredes RG, Vazquez B. What do female rats like about sex? Paced mating Behav Brain Res. 1999;105:117–127. doi: 10.1016/s0166-4328(99)00087-x. [DOI] [PubMed] [Google Scholar]
- 70.Pfaff DW. Estrogens and brain function. Springer-Verlag; New York: 1980. [Google Scholar]
- 71.Pfaff D, Ogawa S, Kia K, Vasudevan N, Krebs C, Frolich J, Kow L-M. Genetic mechanisms in neural and hormonal controls over female reproductive behaviors. In: Pfaff DW, Arnold AP, Etgen AM, Fahrbach SE, Rubin RT, editors. Hormones Brain and Behavior. Vol. 3. Academic Press; Amsterdam: 2002. pp. 441–509. [Google Scholar]
- 72.Poo MM. Neurotrophins as synaptic modulators. Nat Rev Neurosci. 2001;2:24–32. doi: 10.1038/35049004. [DOI] [PubMed] [Google Scholar]
- 73.Radwanska K, Valjent E, Trzaskos J, Caboche J, Kaczmarek L. Regulation of cocaine-induced activator protein 1 tanscription factors by the extracellular-regulated kinase pathway. Neruoscience. 2006;137:253–264. doi: 10.1016/j.neuroscience.2005.09.001. [DOI] [PubMed] [Google Scholar]
- 74.Robinson TE, Berridge KC. The psychology and neurobiology of addiction: an incentive-sensitization view. Addiction. 2000;95 (suppl 2):S91–117. doi: 10.1080/09652140050111681. [DOI] [PubMed] [Google Scholar]
- 75.Robinson TE, Berridge KC. Addiction. Annu Rev Psychol. 2003;54:25–53. doi: 10.1146/annurev.psych.54.101601.145237. [DOI] [PubMed] [Google Scholar]
- 76.Robinson TE, Gorny G, Mitton E, Kolb B. Cocaine self-administration alters the morphology of dendrites and dendritic spines in the nucleus accumbens and neocortex. Synapse. 2001;39:257–266. doi: 10.1002/1098-2396(20010301)39:3<257::AID-SYN1007>3.0.CO;2-1. [DOI] [PubMed] [Google Scholar]
- 77.Robinson TE, Gorny G, Savage VR, Kolb B. Widespread but regionally specific effects of experimenter- versus self-administered morphine on dendritic spines in the nucleus accumbens, hippocampus, and neocortex of adult rats. Synapse. 2002;46:271–279. doi: 10.1002/syn.10146. [DOI] [PubMed] [Google Scholar]
- 78.Robinson TE, Kolb B. Alterations in the morphology of dendrites and dendritic spines in the nucleus accumbens and prefrontal cortex following repeated treatment with amphetamine or cocaine. Eur J Neurosci. 1999;11:1598–1604. doi: 10.1046/j.1460-9568.1999.00576.x. [DOI] [PubMed] [Google Scholar]
- 79.Roitman MF, Na E, Anderson G, Jones TA, Bernstein IL. Induction of a salt appetite alters dendritic morphology in nucleus accumbens and sensitizes rats to amphetamine. J Neurosci. 2002;22:RC225. doi: 10.1523/JNEUROSCI.22-11-j0001.2002. (1–5) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Salamone JD, Correa M, Mingote SM, Weber SM. Beyond the reward hypothesis: alternative functions of nucleus accumbens dopamine. Curr Opin Pharmacol. 2005;5:34–41. doi: 10.1016/j.coph.2004.09.004. [DOI] [PubMed] [Google Scholar]
- 81.Steward O, Worley PF. A cellular mechanism for targeting newly synthesized mRNAs to synaptic sites on dendrites. Proc Natl Acad Sci USA. 2001;98:7062–7068. doi: 10.1073/pnas.131146398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Sweatt JD. The neuronal MAP kinase cascade: a biochemical signal integration system subserving synaptic plasticity and memory. J Neurochem. 2001;76:1–10. doi: 10.1046/j.1471-4159.2001.00054.x. [DOI] [PubMed] [Google Scholar]
- 83.Valjent E, Pascoli V, Svenningsson P, Paul S, Enslen H, Corvol JC, Stipanovich A, Caboche J, Lombroso PJ, Nairne AC, Greengard P, Herve D, Girault JA. Regulation of a protein phosphatase cascade allows convergent dopamine and glutamate signals to activate ERK in the striatum. Proc Nat Acad Sci (USA) 2005;102:491–496. doi: 10.1073/pnas.0408305102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Xiao L, Becker JB. Hormonal activation of the striatum and the nucleus accumbens modulates paced mating behavior in the female rat. Horm Behav. 1997;32:114–124. doi: 10.1006/hbeh.1997.1412. [DOI] [PubMed] [Google Scholar]