

Short abstract
Mast cell (MC) activation in the rheumatoid lesion provides numerous mediators that contribute to inflammatory and degradative processes, especially at sites of cartilage erosion. MC activation in rheumatoid synovial tissue has often been associated with tumour necrosis factor (TNF)-α and interleukin (IL)-1β production by adjacent cell types. By contrast, our in situ and in vitro studies have shown that the production of IL-15 was independent of MC activation, and was not related to TNF-α and IL-1β expression. Primary cultures of dissociated rheumatoid synovial cells produced all three proinflammatory cytokines, with production of IL-1β exceeding that of TNF-α, which in turn exceeded that of IL-15. In vitro cultures of synovial macrophages, synovial fibroblasts and articular chondrocytes all produced detectable amounts of free IL-15, macrophages being the most effective.
Keywords: interleukin-15, interleukin-1β, mast cells, rheumatoid arthritis, tumour necrosis factor-α
Abstract
Introduction:
Increased numbers of mast cells (MCs) are found in the synovial tissues and fluids of patients with rheumatoid arthritis (RA), and at sites of cartilage erosion. MC activation has been reported for a significant proportion of rheumatoid specimens. Because the MC contains potent mediators, including histamine, heparin, proteinases, leukotrienes and multifunctional cytokines, its potential contributions to the processes of inflammation and matrix degradation have recently become evident.
Proinflammatory cytokines are important mediators of inflammation, immunity, proteolysis, cell recruitment and proliferation. Tumour necrosis factor (TNF) reportedly plays a pivotal role in the pathogenesis of RA, especially its ability to regulate interleukin (IL)-1β expression, this being important for the induction of prostanoid and matrix metalloproteinase production by synovial fibroblasts and chondrocytes. IL-15 has been assigned numerous biological effects and has been implicated as an important factor in TNF-α expression by monocyte/macrophages. Some in vitro studies have placed IL-15 upstream from TNF-α in the cytokine cascade, suggesting an interdependence between TNF, IL-1 and IL-15 for the promotion of proinflammatory cytokine expression in the rheumatoid joint.
Aims:
To examine the in situ relationships of TNF-α, IL-1β and IL-15 in relation to MC activation in rheumatoid tissues by use of immunolocalization techniques; and to compare quantitatively the proinflammatory cytokine production by specific cell cultures and rheumatoid synovial explants with and without exposure to a MC secretagogue.
Materials and methods:
Samples of rheumatoid synovial tissue and cartilage–pannus junction were obtained from patients (n = 15) with classic late-stage RA. Tissue sections were immunostained for MC (tryptase) and the proinflammatory cytokines IL-1, TNF-α and IL-15. Rheumatoid synovial tissue explants were cultured in Dulbecco's modified Eagles medium (DMEM) containing either the MC secretagogue rabbit antihuman immunoglobulin (Ig)E, or control rabbit IgG. Primary rheumatoid synovial cell cultures, human articular chondrocytes, synovial fibroblasts and synovial macrophages were prepared as described in the full article. Conditioned culture media from these cultures were collected and assayed for IL-1β, TNF-α and IL-15 using enzyme-linked immunosorbent assay methodology.
Results:
Immunohistological studies of rheumatoid synovial tissues have demonstrated local concentrations of MCs in most specimens of the rheumatoid lesion. Sites of MC activation were associated with localized oedema, and TNF-α, IL-1α and IL-1β production by a proportion of mononuclear inflammatory cells. By contrast, no evidence was found for IL-15 production in tissue sites containing either intact or activated MCs, and IL-15 expression, when observed, bore no relation to tissue sites where TNF-α and IL-1β were evident. The immunodetection of IL-15 was restricted to microfocal sites and was not typical of most junctional specimens, but was associated with a proportion of articular chondrocytes in a minority of junctional specimens.
MC activation within synovial explant cultures was induced by the addition of polyclonal antibody to human IgE. MC activation significantly reduced the levels of TNF-α and IL1β released into the medium, this representing approximately 33% of control values. By contrast, MC activation had little effect on the levels of IL-15 released into the culture medium, the average value being very low in relation to the release of TNF-α and IL-1β . Thus, induced MC activation brings about changes in the amounts of released tryptase, TNF-α and IL-1β , but not of IL-15.
Four preparations of primary rheumatoid synovial cell cultures produced more IL-1β than TNF-α, with only modest values for IL-15 production, indicating that all three cytokines are produced and released as free ligands by these cultures. Of specific cell types that produced IL-15 in vitro, macrophages produced more than fibroblasts, which in turn produced more than chondrocytes. This demonstrates that all three cell types have the potential to produce IL-15 in situ.
Discussion:
The biological consequences of MC activation in vivo are extremely complex, and in all probability relate to the release of various combinations of soluble and granular factors, as well as to the expression of appropriate receptors by neighbouring cells. The subsequent synthesis and release of cytokines such as TNF-α and IL-1 may well follow at specific stages after activation, or may be an induced cytokine response by adjacent macrophagic or fibroblastic cells. However, because no IL-15 was detectable either in or around activated or intact MCs, and the induced MC activation explant study showed no change in IL-15 production, it seems unlikely that the expression of this cytokine is regulated by MCs. The immunohistochemistry (IHC) demonstration of IL-15 at sites of cartilage erosion, and especially by some chondrocytes of articular cartilage, showed no spatial relationship with either T cells or neutrophils, and suggests other functional properties in these locations. The lack of evidence for an in situ association of IL-15 with TNF and IL-1 does not support a role for IL-15 in a proinflammatory cytokine 'cascade', as proposed by other in vitro experiments. We believe that sufficient evidence is available, however, to suggest that MC activation makes a significant contribution to the pathophysiological processes of the rheumatoid lesion.
Introduction
Rheumatoid arthritis (RA) is characterized by chronic inflammation, cartilage destruction and joint deformity. Histopathological observations of the 'rheumatoid lesion' — a term used to describe cartilage-pannus junctions and cartilage erosion sites [1] — have identified a number of cell types, each of which may contribute different mediators to the inflammatory and degradative processes; these usually being microenvironmental in nature [1,2]. Although macrophages, fibroblastic synoviocytes, lymphocytes and neutrophils are recognized as important contributors to RA joint pathology, the mast cell (MC) has generally been neglected. Increased numbers of MCs are found in the synovial tissue and fluid of patients with RA [3,4], and especially at sites of cartilage erosion [5]. MC activation has been reported in these locations for a significant proportion of rheumatoid specimens [3]. Because the MC contains a variety of potent mediators, including histamine, heparin, proteinases, leukotrienes and multifunctional cytokines [6,7], its potential contributions to the processes of inflammation and matrix degradation have recently become evident [2,8].
Proinflammatory cytokines are important mediators of inflammation, immunity, proteolysis, cell recruitment and proliferation. Tumour necrosis factor (TNF)-α and TNF-β, and interleukin (IL)-1α and IL-1β have received much attention over the past decade [9,10,11], and both TNF and IL-1 have been demonstrated in the rheumatoid lesion by immunohistochemistry [2,12]. TNF reportedly plays a pivotal role in the pathogenesis of RA [10,11], especially its ability to regulate IL-1β expression, this being important for the induction of prostanoid and matrix metalloproteinase production by synovial fibroblasts and chondrocytes [9,13]. Cellular interactions mediated by TNF and IL-1, cytokines that are mainly produced by activated macrophages, have become prominent factors in the numerous reviews that have proposed a sequence of events leading to cartilage damage in RA [10,11,13]. The precise factors that induce monocyte/macrophages to produce TNF and IL-1 remain obscure, however, although recent studies [14,15,16] have suggested a role for IL-15.
IL-15 is reportedly expressed by activated monocytes, epithelial cells and fibroblasts [17]. IL-15 has been assigned numerous biological effects: it is a potent T-cell chemokine; it induces proliferation of mitogen-activated T cells; it contributes to B-cell proliferation and immunoglobulin (Ig) synthesis; and it enhances neutrophil and natural killer cell responses [17]. It was recently implicated as an important factor in TNF-α expression by monocyte/ macrophages. IL-15 together with IL-6 and TNF-α was shown to induce monocyte TNF-α production, a response similarly induced by IL-15-stimulated T cells via contact with monocytes [14,15,16]. Other studies have placed IL-15 upstream from TNF-α in the cytokine cascade, showing that IL-15 activation of T cells directly produces TNF-α as well as amplifying inflammatory responses [15,17]. Such reports, derived from in vitro cell studies, have suggested an interdependence between TNF, IL-1 and IL-15, and between monocyte/macrophages and T cells, for the promotion of proinflammatory cytokine expression in the rheumatoid joint. The present study examines these possible relationships in situ, especially in relation to MC activation, by use of immunolocalization techniques on freshly fixed rheumatoid tissues, and by a comparative quantitative assessment of proinflammatory cytokine production by rheumatoid synovial explants and specific cell cultures. We report here that IL-15 production in situ does not appear to be related to sites of TNF or IL-1 production, or with MC activation, and that synovial explants, synoviocytes, macrophages and chondrocytes in vitro produce quite modest amounts of IL-15 compared with those of TNF-α and IL-1β .
Materials and methods
Tissue samples
Samples of rheumatoid synovial tissue and cartilage-pannus junction were obtained from arthroplasty specimens from patients (n = 15) with classic late-stage rheumatoid arthritis (RA). Each sample was fixed in Carnoy's fixative at 20°C for 1–2 h and embedded in paraffin wax, and 5 μ m sections were cut. Tissue sections were dewaxed, rehydrated and examined for the presence of MCs and the proinflammatory cytokines IL-1, TNF-α and IL-15 using immunohistochemical techniques.
Immunolocalization of mast cells
Tissue sections were dewaxed and pretreated for 30 min with 10% rabbit serum (vol/vol) in Tris buffered saline (TBS) at pH 7.6. Mouse monoclonal antibody to human MC tryptase (Biogenesis, Poole, Dorset, UK) was diluted 1:200 and applied to the sections for 2 h at 20°C. After three 10-min washes in TBS, alkaline phosphatase-conjugated rabbit antimouse IgG (Dako Ltd, Cambridge, UK) diluted 1:50 was applied for 1 h at 20°C. After further washing the alkaline phosphatase was developed using new fuchsin substrate. Tissue sections were lightly counterstained with Harris's haematoxylin or Toluidine blue as previously described [2,3].
Dual immunolocalization of mast cell tryptase and cytokines IL-1α, IL-1β or TNF-α
Tissue sections were pretreated for 30 min with rabbit serum at 10% (vol/vol) final concentration in TBS. After draining, the sections were incubated at 20°C for 2 h with a combination of primary antibodies, namely mouse monoclonal antitryptase (diluted 1 : 200) with either a goat polyclonal antibody to IL-1α or IL-1β (diluted 1 : 50) or goat polyclonal antibody to TNF-α (diluted 1 : 50). Polyclonal antibodies were purchased from R&D Systems (Abingdon, UK). After three 10 min washes in TBS, secondary antibodies alkaline phosphatase-conjugated rabbit antimouse and horseradish peroxidase-conjugated rabbit antigoat (both from Dako) diluted 1 : 50 in TBS were applied together for 1 h at 20°C followed by three 10-min washes in TBS. Peroxidase was developed first using diaminobenzidine (brown colour) and the sections were washed, followed by development of alkaline phosphatase using new fuchsin (red colour) as described previously [2]. The sections were lightly counterstained in Harris's haematoxylin to demonstrate nuclei, dehydrated, mounted in Histomount (Mensura, Wigan, UK) and photographed using a Zeiss (Oberkochen, Germany) photomicroscope III and Ektachrome (Rochester, New York, USA) 160 tungsten film.
Immunolocalization of interleukin-15
IL-15 was immunolocalized using a goat polyclonal antibody from R&D Systems.
Tissue sections were pretreated with 10% (vol/vol) rabbit serum in TBS for 30 min. The primary antibody, diluted 1 : 100 in TBS, was applied to the sections and incubated for 2 h at 20°C. After three 10-min washes, biotinylated rabbit antigoat IgG was applied to the sections for 45 min, followed by a further washing in TBS. Alkaline phosphatase conjugated avidinbiotin complex was diluted as recommended by the supplier (Dako) and applied to the sections for 45 min at 20°C. After further washing alkaline phosphatase was developed using new fuchsin substrate (red). Consecutive sections of cartilage-pannus junction and rheumatoid synovium were stained for MC tryptase, IL-15, TNF-α and IL-1β, respectively.
Control tissue sections
Normal IgG from the same species and in concentrations similar to those used for the primary antibodies was substituted for each primary antibody and consistently gave negative results. In addition, each cytokine antibody was preabsorbed with its relevant antigen and used in place of the primary antibody; this also produced negative data and confirmed specificity.
Explant cultures
Rheumatoid synovial tissue obtained shortly after surgery was cut into small pieces of approximately 3 mm3 in Hanks balanced salt solution (HBSS; Gibco, Paisley, Scotland). Randomized explants (8–10) were placed in each of four or six preweighed sterile culture dishes. After reweighing to determine the wet weight of the tissue per dish, 2 ml Dulbecco's modified Eagles medium (DMEM, Gibco) containing either the MC secretagogue rabbit antihuman IgE or control rabbit IgG, each at a final concentration of 150 μ g/ml, was added to the cultures in duplicate or triplicate [18]. The dishes were incubated at 37°C in a 5% air incubator with humidified atmosphere.CO2/95% air incubator with humidified atmosphere.Culture medium was removed at 20 h, aliquoted and stored at -20°C until it was assayed for the cytokines TNF-α, IL-1β and IL-15.
Preparation of rheumatoid synovial cell cultures
Rheumatoid synovial tissue was washed with HBSS, chopped into fine pieces and enzymically digested by incubation with 10 ml DMEM containing 1 mg/ml bacterial collagenase (Clostridium hystoliticum; Boehringer, Mannheim, Germany) in a shaking water bath for 3 h at 37°C. The resultant cell suspension was filtered, washed with DMEM and the pellet of cells was resuspended in DMEM + 10% (vol/vol) foetal calf serum (FCS), as described previously [19]. An aliquot of 200 μ l dissociated synovial cells were plated at a density of 2 ×105 cells/well in 12-well culture dishes and incubated at 37°C with a 5% CO2 incubator. After 48 h the conditionedmedium was harvested, centrifuged to remove nonadherent cells and stored at -20°C until assaying for the cytokines TNF-α, IL-1β and IL-15.
Synovial macrophage-enriched cultures
Dissociated rheumatoid synovial cells prepared as described above were plated into 80 cm2 culture flasks and, after 24 h, the medium containing the nonadherent cells was removed and discarded. The remaining adherent cells, mainly fibroblasts and macrophages, were first washed with HBSS and then harvested by trypsinization. The cells were resuspended in HBSS containing 2% FCS using 80 μ l buffer per 107 cells. Cells of anti-fibroblast microbeads 20 μ l per 107 (Miltenyi Biotec, Bisley, Surrey, UK) was added to the cell suspension and the mixture was incubated at 20°C for 30 min. The cells were washed in 20 times the volume of the HBSS and centrifuged at 2000 revolutions/min for 10 min. After resuspension in 1 ml HBSS + 2% FCS, the cells were separated using a MS + magnetic separation column (Miltenyi Biotec). The resultant eluant gave cell preparations that were depleted of fibroblasts and rich in macrophages. The cells were plated out in DMEM + 10% FCS into 12-well culture dishes overnight and then transferred to DMEM + 2% FCS for 48 h. Cultures were shown to contain more than 90% macrophages by immunostaining with CD68 macrophage marker (Dako; data not shown). The medium was harvested and stored at -20°C until assaying for the cytokines IL-1β, TNF-α and IL-15 by enzyme-linked immunosorbent assay (ELISA). The cells were fixed in the culture wells, stained and counted.
Rheumatoid synovial fibroblasts
Subcultures of adherent synovial fibroblasts were used at passages two to four. Cells were plated out in DMEM + 10% FCS into 12-well culture dishes and, when confluent, the medium was replaced with DMEM + 2% FCS for 48 h. Medium was collected, stored and assayed for cytokines. Cells were fixed, stained and counted.
Chondrocyte cultures
Macroscopically normal articular cartilage was enzymically digested as described previously [20]. Chondrocytes were grown to confluence in 12-well culture dishes in DMEM + 10% FCS and used as described above for the synovial macrophage and fibroblast cultures, after which cells were fixed, stained and counted.
Measurement of cytokines
IL-1β, TNF-α and IL-15 were measured using Quantikine ELISA kits from R&D Systems, following the manufacturer's instructions. Detection limits of the ELISAs were <1pg/ml, 4pg/ml and 1pg/ml for IL-1β, TNF-α and IL-15, respectively.
Results
Immunohistological studies of rheumatoid synovial tissues have demonstrated local concentrations of MCs in most specimens of the rheumatoid lesion. Previous studies have shown evidence of MC activation in situ, as judged by the release of the MC-specific enzyme tryptase. Figure 1a shows MC activation with evidence of local oedema, associated with sites of TNF-α production by a proportion of mononuclear inflammatory cells. Figure 1b also shows dual immunolocalization of MC tryptase together with IL-1β production by some neighbouring cells. Associations of IL-1β and sites of MC activation were similarly observed [2]. Although MCs are reported to have the potential to express TNF-α and IL-1, only occasionally has TNF-α production by MCs been demonstrated in our rheumatoid specimens.
Figure 1.
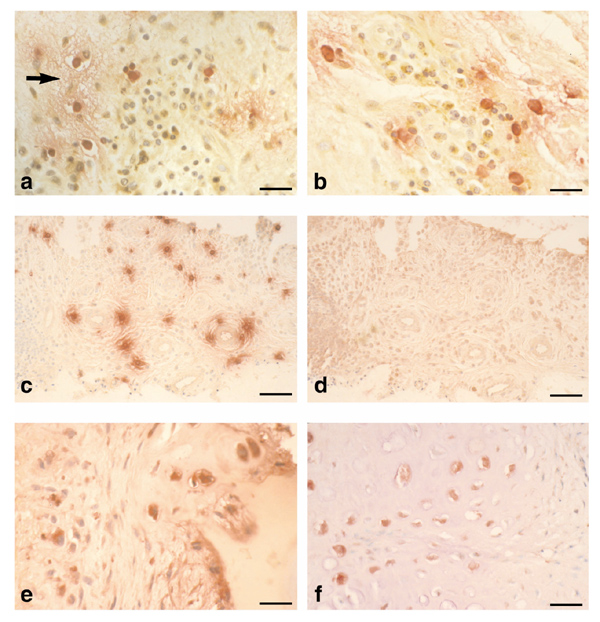
Immunolocalisation of mast cell tryptase, tumour necrosis factor (TNF)-α, interleukin (IL)-1α and IL-15 in the rheumatoid lesion. (a) Micrograph showing mast cell tryptase (red) with extracellular staining indicative of mast cell (MC) activation associated with localized oedema (arrow) and TNF-α expression (brown) by a proportion of mononuclear cells. (b) Micrograph showing mast cell tryptase (red), local MC activation and associated mononuclear cells stained for IL-1α (brown). (c) Low power micrograph showing distribution of mast cells (tryptase, red) and their degranulation in rheumatoid synovial tissue. (d) Consecutive section to (c) stained for IL-15. Note the absence of IL-15 from MC activation sites. (e) Micrograph of cartilage-pannus junction showing both extracellular and intracellular staining for IL-15 (red). Note the microfocal nature of cytokine production by a minority of cells. (f) Micrograph of rheumatoid lesion showing chondrocytic cells stained for IL-15 (red) with little evidence of cytokine production by the overlying pannus tissue (to right of micrograph). Bars: (a) and (b) 35 μm; (c) and (d) 120 μm; (e) 35 μm; and (f) 45 μm.
The association of TNF-α and IL-1α or IL-1β production by cells in similar locations to activated MCs was a common observation. By contrast, no evidence was found for IL-15 production in tissue sites containing either intact or activated MCs. Figures 1c and 1d show consecutive tissue sections stained for tryptase and IL-15, respectively; the latter showing negligible staining despite the presence of numerous MCs, some of which show evidence of degranulation. Indeed, IL-15 expression, when observed, bore no relation to tissue sites where TNF-α and IL-1β were evident. The immunodetection of IL-15 was variable between the different rheumatoid tissue specimens; whereas some showed prominent staining of synovial lining cells, especially CD68+ macrophages, other specimens were devoid of IL-15. Figure 1e shows a cartilage-pannus junction with evidence of extracellular IL-15 staining at cartilage erosion sites, and with intracellular staining localized to a few chondrocytic and macrophagic cells. This distribution of IL-15 was restricted to microfocal sites; it was not typical of most junctional specimens, but was not associated with the local expression of TNF-α or IL-1. Similarly, Figure 1f shows a proportion of articular chondrocytes at a cartilage-pannus junction stained for IL-15. Again, such observations were restricted to discrete sites where only a proportion of the cells were positive for IL-15. Thus, it appears that the production of IL-15 in rheumatoid tissues is independent of MC activation, and the local production of TNF-α and IL-1α and IL-1β.
We recognize that all of these specimens are fixed at one window in time, but despite the reported interdependence of these proinflammatory cytokines we have not as yet observed any colocalization of IL-15 with TNF or IL-1 in any of the 15 rheumatoid specimens fixed within minutes of surgical excision. Table 1 is a general summary of the immunolocalization data, which provides information on the relative frequencies for cytokine production in specimens of rheumatoid tissues and at the cartilage-pannus interface. Of special note is that most positive observations were microfocal in nature, seldom involving large groups of cells, with a significant proportion of the CD68 + macrophagic cells showing no evidence of cytokine production. Such observations suggest that at least in situ cytokine expression by macrophages and chondrocytes is subject to regulation, but the induction factors remain uncertain. These findings have been further examined using in vitro studies with rheumatoid synovial explants and monolayer cultures of specific cell types.
Table 1.
Summary of tryptase and cytokine immunolocalization data
| Antigen | Cartilage-pannus junctions (n = 35) | Rheumatoid synovial tissue (n = 26) |
| MC tryptase | Approximately 50% of specimens showed local | All specimens contained MCs, but with variable distributions and |
| accumulations of MCs, most with extracellular enzyme | states of activation both between and within individual specimens | |
| indicative of activation/degranulation | ||
| TNF-α | Approximately 80% of specimens stained; occasional | Most specimens showed only intracellular staining; usually |
| extracellular staining of cartilage matrix and intracellular | macrophages of the synovial lining layer | |
| staining of chondrocytes | ||
| IL-1β | Approximately 80% of specimens positive; extracellular | Commonly observed for a proportion of macrophages, especially |
| distribution often reflecting that for TNF-α | those in the lining layer | |
| IL-1α | Staining limited to about 30% of specimens and | Mainly restricted to a minor proportion of macrophages of the |
| usually associated with macrophages | synovial lining layer | |
| IL-15* | Microfocal staining observed for about 30% of | Restricted to approximately 40% of specimens; usually |
| specimens, usually associated with macrophages, | macrophages of the synovial lining and a few fibroblastic cells; | |
| and especially chondrocytes; no association with TNF | seldom associated with TNF+, IL-1+ or tryptase+ cells | |
| or IL-1 |
*Interleukin (IL)-15 was examined in 9 and 12 different specimens for cartilage-pannus junction and rheumatoid synovial tisue respectively. The tryptase, tumour necrosis factor (TNF) and IL-1 data are partly adapted from Tetlow and Woolley [2]. MC, mast cell.
MC activation within synovial explant cultures was induced by the addition of polyclonal antibody to human IgE, this being an effective MC secretagogue. Its effect was confirmed by the elevated values for MC tryptase in the culture medium, with most cultures showing more than a threefold increase over controls (Table 2). Because the cellular composition of each synovial specimen is known to be variable in terms of both total cell numbers and specific cell types, quantitative differences for tryptase and cytokine values between cultures were to be expected. MC activation significantly reduced the levels of TNF-α released into the medium, this representing approximately 30% of control values for most explant cultures (average values being 177.2 and 53.2 pg/ml per 100 mg tissue per 20 h for control and anti-IgE treatments, respectively). Similarly MC activation reduced the levels of IL-1β released into the medium, this representing approximately 33% of control values (averaged values being 50.1 and 16.1 pg/100 mg tissue per 20 h for control and anti-IgE treatments, respectively). By contrast, MC activation apparently had little effect on the levels of IL-15 released into the culture medium, the average value of less than 1 pg/ml per 100 mg tissue per 20 h being very low in relation to the release of TNF-α and IL-1β (Table 2). Thus, the explant data demonstrate that induced MC activation brings about changes in the amounts of released tryptase, TNF-α and IL-1-β, but not of IL-15.
Table 2.
Effects of mast cell activation on cytokine production by explanted rheumatoid synovial tissue
| Tryptase | TNF-α | IL-β | IL-15 | ||||||
| Sample | Control | Anti-IgE | Control | Anti-IgE | Control | Anti-IgE | Control | Anti-IgE | |
| 753 | 11.1 | 30.4 | 138 | 6.4 | 24.2 | 4.1 | 0.31 | 0.28 | |
| 764 | 28.1 | 181.6 | 271 | 204 | 11.4 | 20.9 | 0.55 | 0.4 | |
| 770 | 8.6 | 46.6 | 12.8 | 276 | 30.3 | 11.9 | 0.54 | 0.54 | |
| 773 | 8.9 | 47.9 | 103 | 8.5 | 19.1 | 5.8 | 0.27 | 0.3 | |
| 798 | 32.3 | 42.1 | 97.8 | 34.6 | 165.3 | 38 | 0.52 | 0.49 | |
| Averaged values | 17.8 | 69.7 | 177.2 | 53.2 | 50.1 | 16.1 | 0.44 | 0.40 | |
Values expressed as ng/ml per 100 mg tissue per 20 h for tryptase and as pg/ml per 100 mg tissue for cytokines. For tryptase P = 0.05, for tumour necrosis factor-α P = 0.02, and for interleukin (IL)-1β P = 0.28, by Student's T-test. Ig, immunoglobulin.
It is possible that the measurements of tryptase and cytokines in the conditioned culture medium do not necessarily reflect total release or production by the tissue explants, because the accessibility of each protein into the culture medium will depend to some extent on the relative interactions and binding properties to the extracellular matrix, cells and receptors. To overcome this problem of possible retention of cytokines by the three-dimensional properties of the explant tissue, production of the three cytokines by primary cultures of dissociated rheumatoid synovial cells was examined. Table 3 shows that, for four preparations of primary rheumatoid synovial cell cultures, variations were observed for the relative values of the three cytokines. This in all probability reflects the different compositions of specific cell types for each preparation. In three of the cultures, however, the IL-1β values were greater than those for TNF-α, with only modest values for IL-15 production. This trend is borne out by the averaged values for each cytokine; IL-1β production was greater than that of TNF-α, which in turn was greater than that of IL-15, these being 190, 85 and 14 pg/106 cells per 48 h, respectively (Table 3). It is recognized that membrane- or surface-bound forms of these cytokines exist for specific cell types, and these will not have contributed to the values presented here because all cells were removed from the conditioned medium before assay was carried out. Similarly, cytokine receptor expression by the cell cultures will contribute to some cytokine depletion of the conditioned medium. Nevertheless, these experiments with primary cultures of synovial cells give some indication that all three cytokines are produced and released as free ligands, with IL-15 representing approximately 5% of the three cytokines present in the medium after 48 h of culture.
Table 3.
Cytokine production by primary dissociated synovial cells
| Sample no. | TNF-α | IL-1β | IL-15 |
| 856 | 32 | 135 | 7 |
| 859 | 140 | 420 | 15 |
| 851 | 104 | 44 | 9 |
| 848 | 65 | 160 | 25 |
| Average values | 85.3 ± 23.4 | 189.8 ± 80.6 | 14.0 ± 4.04 |
Values are expressed as pg/ml per 106 cells per 48 h. IL, interleukin; TNF, tumour necrosis factor.
Several cell types have been shown to produce IL-15 in vitro, but few studies have compared production by different cell types. Table 4 shows the values for IL-15 production and release into the culture medium by synovial macrophages, synovial fibroblasts and human articular chondrocytes. The data were obtained from three different preparations of each cell type, maintained in medium containing 2% FCS supplement, the latter having no detectable IL-15. In relation to cell numbers the ability of these cells to produce IL-15 followed the order macrophages, followed by fibroblasts, followed by chondrocytes, thereby demonstrating that all three cell types have the potential to produce this cytokine in situ.
Table 4.
Comparison of interleukin (IL)-15 production by macrophages, fibroblasts and chondrocytes from rheumatoid tissues
| Cell type | IL-15 production in vitro |
| Synovial macrophages | 16.3 ± 4.1 |
| Synovial fibroblasts | 8.2 ± 4.4 |
| Chondrocytes | 3.7 ± 2.0 |
Values are expressed as means ± standard error of the mean, and are given as pg/ml per 106 cells per 48 h (n = 3).
Discussion
Recent histological studies of rheumatoid synovial tissues have demonstrated localized accumulations of MCs and evidence of their activation/degranulation, especially at cartilage erosion sites [2,3]. Because MCs contain or express numerous potent mediators, including histamine, heparin, proteinases, prostanoids, cytokines and growth factors, it seems most likely that MC secretion, activation or degranulation in situ will bring about changes in the local environment, not least the phenotype of neighbouring cells. For example, histamine induces tissue oedema via its effects on endothelial cells, activates chondrocytes via H1 and H2 receptors, and stimulates synoviocytes through H1 receptors [21,22]. Heparin interacts with growth factors, contributes to angiogenesis and stimulates IL-1 production by monocyte/macrophages [23,24]. Tryptase and chymase are MC-specific proteinases that degrade various matrix components, and are capable of activating the zymogen forms of the matrix metalloproteinases, prostromelysin and procollagenase [25,26]. These MC enzymes are used to identify two subsets of MCs in human tissues. One subsetcontains both tryptase and chymase, and is termed MCTC; the other contains only tryptase, and is known as MCT. Both subsets have been demonstrated in rheumatoid synovial tissues, where they are purported to exert distinct functions with regard to inflammatory and degradative processes [27,28]. In addition to proteinase heterogeneity the MC subsets are reported to have different cytokine profiles; whereas MCT preferentially produce IL-4, IL-5 and IL-6, the MCTC phenotype contains predominantly IL-4 [29]. Indeed, the realization that MCs can express several multi-functional cytokines (including the proinflammatory mediators TNF-α and IL-1β [7,8]) and various profibrotic cytokines (such as basic fibroblast growth factor and IL-4), and have the ability to release membrane-bound TGF-α [28], suggests numerous functional roles for MCs in the rheumatoid lesion [30].
Our earlier immunohistochemistry (IHC) studies of rheumatoid synovial tissue presented evidence for MC activation, as judged by the extracellular release of tryptase, and its association with the microenvironmental expression of TNF-α, IL-1α, IL-1β, stromelysin and collagenase-1 [2]. These observations were in accord with previous reports that soluble MC products stimulated collagenase production by cultures of synoviocytes and chondrocytes [31], and induced monocyte/macrophages to increase IL-1 production [24]. Such studies suggested that MC activation was likely to bring about increased proinflammatory cytokine and metalloproteinase production in situ, but paradoxically in the present study the induced MC activation of synovial explants resulted in a significant reduction in TNF-α and IL-1β release into the culture medium. Explanations for this are currently unresolved, but may relate to an increase in the expression of the relevant receptors or to some binding or sequestration of these ligands by the tissue matrix or released heparin.
The biological consequences of MC activation in vivo are extremely complex, and in all probability relate to the release of various combinations of soluble and granular factors, as well as the expression of appropriate receptors by neighbouring cells. As yet we have a poor understanding of the hierarchy of the MC mediators released upon degranulation. Although histamine is released rapidly and induces tissue oedema, the solubilization and release of factors from exocytosed granules provides a temporally regulated supply of specific signals within the localized domain of the degranulated MC [23]. The subsequent synthesis and release of specific cytokines may well follow at specific stages after activation, or, as demonstrated here by IHC, may be an induced cytokine response by adjacent macrophagic or fibroblastic cells. It is therefore possible that the IHC observations presented here reflect transient changes in cytokine expression, the extracellular tissue distribution observed for the tryptase protein possibly taking longer to process or remove from the tissue than the three cytokines. However, because no IL-15 was detectable either in or around activated or intact MCs, and the induced MC activation explant study showed no change in the IL-15 production, it seems unlikely that the expression of this cytokine is regulated by MCs.
The proinflammatory cytokines IL-1 and TNF-α are reported to play important roles in cartilage and bone degradation, with TNF-α occupying a primary position in the cytokine cascade through its ability to upregulate production of other cytokines, including IL-1, granulocyte-macrophage colony-stimulating factor, IL-6, IL-8 and IL-10 [10,11]. TNF-α expression by monocyte/macrophages has been demonstrated both in synovial tissue and at the cartilage-pannus junction [32]. Similarly, IL-1 and its receptor have been demonstrated at sites of cartilage erosion [12]. Although monocyte/macrophages are recognized sources of both TNF-α and IL-1, in many specimens it is only a proportion of the macrophage numbers that express these cytokines at the time of surgery and tissue fixation. Similarly with MCs, although reported to produce TNF and IL-1 only very few have been shown to do so in rheumatoid synovial tissue. Such observations indicate that both TNF and IL-1 expression is subject to regulation, but the nature of the stimulatory agents remains uncertain [10]. Endotoxin, immunoglobulins, heparin and rheumatoid factors have been invoked, but more recent studies have shown the importance of T cells in mediating TNF and IL-1 production by monocyte/macrophages, especially via cell surface signals from T cells after their priming with IL-15 [14,15,16].
Several cell types are capable of producing IL-15. These include activated monocytes, macrophages, epithelial cells, fibroblasts [17,33] and endothelial cells [34], and now chondrocytes as described in the present study. Other immunohistochemical studies of IL-15 in rheumatoid synovial tissues have identified CD68+ lining cells [14], macrophages, T cells and natural killer cells as positively stained for IL-15 [35]. One of its many functions is the recruitment, migration and activation of T cells, subsequently contributing to monocyte-derived TNF-α production via cell surface contact [17]. IL-15 is also reported to stimulate monocytes to produce the chemokines IL-8 and monocyte chemotactic protein-1, which play important roles in the regulation of leucocyte infiltration during inflammation [33], a concept also proposed for the cellular composition of rheumatoid synovial tissue [35].
The present observations are generally in accord with these reports, atleast with regard to IL-15 expression by some synovial lining cells. However, the demonstration of IL-15 at sites of cartilage erosion, and especially by some chondrocytes of articular cartilage, showed no spatial relationship with T cells or neutrophils, and therefore suggest other functional properties in these locations. This is supported to some extent by the lack of evidence for an in situ association of IL-15 with TNF and IL-1, observations that do not necessarily support a role for IL-15 in a proinflammatory cytokine 'cascade', as determined by in vitro experiments [14,15,16].
The present study has shown that the production of IL-15 by synovial explants and by cells in vitro was rather modest when compared with that for TNF-α and IL-1β. Harada et al [36] reported that fibroblast-like synoviocytes in culture produced large amounts of IL-15 protein and messenger RNA, this being elevated when stimulated with IL-1 or TNF. Receptor expression for each of these cytokines is clearly an important aspect for the understanding of these cytokine interactions, and so too is the realization that some cell types manifest membrane-bound cytokines. Biologically active IL-15 was detected in a constitutively expressed, membrane-bound form on normal human monocytes and monocytic cell lines, prompting Musso et al [37] to speculate that most of the natural biological effects of IL-15 are exerted by the cell surface-bound form. Cell membrane-bound forms of IL-1, TNF-α and IL-10 have also been reported, but it is unclear at present how these relate to the corresponding free forms of TNF-α, IL-1β and IL-15 measured in the experiments described here. It is apparent that the cellular interactions and proinflammatory cytokine responses that promote the inflammatory processes of rheumatoid synovial tissue and joint destruction are far from being resolved. We believe, however, that sufficient evidence is available to suggest that MC activation makes a significant contribution to the pathophysiological processes of the rheumatoid lesion.
Acknowledgements
We thank consultant orthopaedic surgeons T Dunningham (Tameside Hospital, Manchester) and M Morris (Devonshire Royal Hospital, Buxton) for the supply of rheumatoid tissues. This work was supported by project grants for the Arthritis Research Campaign, UK.
References
- Woolley DE, Tetlow LC. Observations on the microenvironmental nature of cartilage destruction in rheumatoid arthritis. Ann Rheum Dis. 1997;56:151–161. doi: 10.1136/ard.56.3.151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tetlow LC, Woolley DE. Mast cells, cytokines and metalloproteinases at the rheumatoid lesion: dual immunolocalisation studies. Ann Rheum Dis. 1995;54:896–903. doi: 10.1136/ard.54.11.896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tetlow LC, Woolley DE. Distribution, activation and tryptase/chymase phenotype of mast cells at the rheumatoid lesion. Ann Rheum Dis. 1995;54:549–555. doi: 10.1136/ard.54.7.549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gruber BL, Kaplan AP. Mast cells and rheumatic diseases. Arthritis and Allied Conditions, 12th ed. Edited by McCarly DJ, Koopman WJ, Philadelphia: Lea & Febiger. 1993. pp. 417–429.
- Bromley M, Fisher WD, Woolley DE. Mast cells at sites of cartilage erosion in the rheumatoid joint. Ann Rheum Dis. 1984;43:76–79. doi: 10.1136/ard.43.1.76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Church MK, Caulfield JP. Mast cell and basophil functions. Allergy. Edited by Holgate ST, Church MK. London: Gower Medical Publishing. 1993. pp. 5.1–5.12.
- Galli SJ. New concepts about the mast cell. N Engl J Med . 1993;328:257–265. doi: 10.1056/NEJM199301283280408. [DOI] [PubMed] [Google Scholar]
- McNeil HP. The mast cell and inflammation. Aust NZ J Med . 1996;26:216–225. doi: 10.1111/j.1445-5994.1996.tb00887.x. [DOI] [PubMed] [Google Scholar]
- Arend WP, Dayer J-M. Inhibition of the production and effects of interleukin-1 and tumour necrosis factor α in rheumatoid arthritis. Arthritis Rheum. 1995;38:151–160. doi: 10.1002/art.1780380202. [DOI] [PubMed] [Google Scholar]
- Feldmann M, Brennan F, Maini RN. Role of cytokines in rheumatoid arthritis. Annu Rev Immunol. 1996;14:397–440. doi: 10.1146/annurev.immunol.14.1.397. [DOI] [PubMed] [Google Scholar]
- Brennan FM, Maini RN, Feldmann M. Role of pro-inflammatory cytokines in rheumatoid arthritis. Springer Semin Immunopathol. 1998;20:133–147. doi: 10.1007/BF00832003. [DOI] [PubMed] [Google Scholar]
- Chu CQ, Field M, Allard S, Abney E, Feldmann M, Maini RN. Detection of cytokines at the cartilage-pannus junction in patients with rheumatoid arthritis: implications for the role of cytokines in cartilage destruction and repair. Br J Rheumatol. 1992;31:653–661. doi: 10.1093/rheumatology/31.10.653. [DOI] [PubMed] [Google Scholar]
- Krane SM. Mechanisms of tissue destruction in rheumatoid arthritis. Arthritis and Allied Conditions. A Textbook of Rheumatology vol 1. Edited by McCarly DJ, Koopman WJ. Philadelphi: Lea & Febiger. 1993. pp. 763–780.
- McInnes IB, Al Mughales J, Field M, et al. The role of interleukin-15 in T-cell migration and activation in rheumatoid arthritis. . Nature Med. 1996;2:175–182. doi: 10.1038/nm0296-175. [DOI] [PubMed] [Google Scholar]
- McInnes IB, Leung BP, Sturrock RD, Field M, Liew FY. Interleukin-15 mediates T cell-dependent regulation of tumour necrosis factor-alpha in rheumatoid arthritis. Nature Med. 1997;3:189–195. doi: 10.1038/nm0297-189. [DOI] [PubMed] [Google Scholar]
- Sebbag M, Parry SL, Brennan FM, Feldmann M. Cytokine stimulation of T lymphocytes regulates their capacity to induce monocyte production of tumour necrosis factor-alpha, but not interleukin-10: Possible relevance to pathophysiology of rheumatoid arthritis. Eur J Immunol. 1997;27:624–632. doi: 10.1002/eji.1830270308. [DOI] [PubMed] [Google Scholar]
- McInnes IB, Liew FY. Interleukin-15: a proinflammatory role in rheumatoid arthritis synovitis. Immunol Today. 1998;19:75–79. doi: 10.1016/s0167-5699(97)01205-x. [DOI] [PubMed] [Google Scholar]
- Tetlow LC, Harper N, Dunningham T, et al. Effects of induced mast cell activation on prostaglandin E and metalloproteinase production by rheumatoid synovial tissue in vitro. . Ann Rheum Dis. 1998;57:25–32. doi: 10.1136/ard.57.1.25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tetlow LC, Lees M, Ogata Y, Nagase H, Woolley DE. Differential expression of gelatinase B (MMP-9) and stromelysin-1 (MMP-3) by rheumatoid synovial cells in vitro and in vivo. Rheumatol Int. 1993;13:53–59. doi: 10.1007/BF00307734. [DOI] [PubMed] [Google Scholar]
- Meats JE, McGuire MK, Russel RGG. Human synovium releases a factor which stimulates chondrocyte production of PGE and plasminogen activator. Nature. 1980;286:891–892. doi: 10.1038/286891a0. [DOI] [PubMed] [Google Scholar]
- Taylor DJ, Yoffe JR, Brown DM, Woolley DE. Histamine stimulates prostaglandin E production by rheumatoid synovial cells and human articular chondrocytes in culture. Arthritis Rheum. 1986;29:160–165. doi: 10.1002/art.1780290202. [DOI] [PubMed] [Google Scholar]
- Taylor DJ, Woolley DE. Evidence for both H1 and H2 histamine receptors on human articular chondrocytes. Ann Rheum Dis. 1987;47:431–435. doi: 10.1136/ard.46.6.431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Norrby K, Woolley DE. Role of mast cells in mitogenesis and angiogenesis in normal tissue and tumour tissue. Advances in the Biosciences, vol 89. Edited by Garcia-caballero M, Brandes LJ, Hosoda S. Oxford: Pergammon Press, 1993. pp. 71–116.
- Yoffe JR, Taylor DJ, Woolley DE. Mast cell products and heparin stimulate the production of mononuclear-cell factor by cultured human monocyte-macrophages. Biochem J. 1985;230:83–88. doi: 10.1042/bj2300083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lees M, Taylor DJ, Woolley DE. Mast cell proteinases activate precursor forms of collagenase and stromelysin but not of gelatinases A and B. Eur J Biochem. 1994;223:171–177. doi: 10.1111/j.1432-1033.1994.tb18980.x. [DOI] [PubMed] [Google Scholar]
- Saarinen J, Kalkinen N, Welgus HG, Kovaren PT. Activation of human interstitial procollagenase through direct cleavage of the leu 83- thr 84 bond by mast cell chymase. J Biol Chem. 1994;269:18134–18140. [PubMed] [Google Scholar]
- Gotis-Graham I, McNeil HP. Mast cell responses in rheumatoid synovium: association of the MCTC subset with matrix turnover and clinical progression. Arthritis Rheum. 1997;40:479–489. doi: 10.1002/art.1780400314. [DOI] [PubMed] [Google Scholar]
- Gotis-Graham I, Smith M, Parker A, McNeil HP. Synovial mast cell responses during clinical improvement in early rheumatoid arthritis. Ann Rheum Dis. 1998;57:664–671. doi: 10.1136/ard.57.11.664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradding P, Feather IH, Wilson S, et al. The mast cell as a source of IL-4, IL-5 and IL-6 in human allergic mucosal inflammation. J Immunol. 1993;151:3853–3865. [PubMed] [Google Scholar]
- Woolley DE. Mast cells in the rheumatoid lesion: ringleaders or innocent bystanders? Ann Rheum Dis. 1995;54:533–534. doi: 10.1136/ard.54.7.533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woolley DE, Yoffe JR, Evanson JM. Mast cells and matrix degradation in the rheumatoid joint. Proteinases in Inflammation and Tumour Invasion. Edited by Tschesche H. Berlin: De Gruyter & Co. 1986. pp. 61–76.
- Chu CQ, Field M, Feldmann M, Maini RN. Localisation of tumour necrosis factor α in synovial tissue and at the cartilage pannus junction in patients with rheumatoid arthritis. Arthritis Rheum. 1991;34:1125–1132. doi: 10.1002/art.1780340908. [DOI] [PubMed] [Google Scholar]
- Badolato R, Ponzi AN, Millesimo M, Notarangelo LD, Musso T. Interleukin 15 (IL-15) induces IL-8 and monocyte chemotactic protein 1 production in human monocytes. Blood. 1997;7:2804–2809. [PubMed] [Google Scholar]
- OppenheimerMarks N, Brezinschek RI, Mahamadzadeh M, Vita R, Lipsky PE. Interleukin 15 is produced by endothelial cells and increases the transendothelial migration of T cells in vitro and in the SCID mouse: human rheumatoid arthritis model in vivo. J Clin Invest. 1998;101:1261–1272. doi: 10.1172/JCI1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thurkow EW, VanDerHeijden IM, Breedveld FC, et al. Increased expression of IL-15 in the synovium of patients with rheumatoid arthritis compared with patients with Yersinia -induced arthritis and osteoarthritis. . J Pathol. 1997;181:444–450. doi: 10.1002/(SICI)1096-9896(199704)181:4<444::AID-PATH778>3.0.CO;2-O. [DOI] [PubMed] [Google Scholar]
- Harada S, Yamaura M, Okamoto H, et al. Production of interleukin-7 and interleukin-15 by fibroblast-like synoviocytes from patients with rheumatoid arthritis. Arthritis Rheum. 1999;42:1508–1516. doi: 10.1002/1529-0131(199907)42:7<1508::AID-ANR26>3.0.CO;2-L. [DOI] [PubMed] [Google Scholar]
- Musso T, Calosso L, Zucca M, et al. Human monocytes constitutively express membrane-bound, biologically active, and interferon-gamma-upregulated interleukin-15. Blood. 1999;93:3531–3539. [PubMed] [Google Scholar]