

Abstract
Cancer gene therapy approaches will derive considerable benefit from adenovirus (Ad) vectors capable of self-directed localization to neoplastic disease or immunomodulatory targets in vivo. The ablation of native Ad tropism coupled with active targeting modalities has demonstrated that innate gene delivery efficiency may be retained while circumventing Ad dependence on its primary cellular receptor, the coxsackie and Ad receptor. Herein, we describe advances in Ad targeting that are predicated on a fundamental understanding of vector/cell interplay. Further, we propose strategies by which existing paradigms, such as nanotechnology, may be combined with Ad vectors to form advanced delivery vehicles with multiple functions.
Keywords: review, vector targeting, adenovirus
Introduction
The understanding of the molecular basis of many pathologies, especially cancer, now demands rationally designed molecular interventions for therapeutic regimes. Over the last 15 years, the field of gene therapy has emerged as a potentially powerful therapeutic platform to serve this end.
The central dogma of gene therapy is employment of gene delivery as a therapeutic molecular intervention to selectively correct or eradicate defective tissues. Early gene therapy efforts, however, revealed that the clinical benefit of gene delivery modalities was irrevocably linked to specific localization of the therapeutic agent. For example, insufficient transduction of cancer cells and solid tumor masses may limit clinical efficacy of cancer gene therapy approaches.1-5 Hence, a fundamental requirement for the achievement of cancer gene therapy is the employment of advanced molecular biology and disease-specific cellular physiology to design targetable gene delivery vectors.
Vectors based on human adenovirus (Ad) serotypes 2 and 5 continue to show increasing promise as gene therapy delivery vehicles, particularly in the context of cancer gene therapy, due to several key attributes: These replication-deficient Ad vectors display in vivo stability and superior gene transfer efficiency to numerous dividing and nondividing cell targets in vivo, and are rarely linked to any severe disease in humans. Further, production parameters for clinical grade Ad vectors are well established. In 2004, Ad vectors comprised one-fourth of all clinical trial gene therapy vectors (256 of 987), with almost two-thirds of gene therapy trials being for cancer (565 of 987).6
Nonetheless, clinical trial results of nontargeted Ad vectors have clearly exposed the need to advance Ad vector technology. The relatively poor clinical performance of Ad vectors can be attributed, in large part, to their broad native tropism, emphasizing the need for the derivation of Ad agents that have the capacity for intrinsic, self-directed, specific localization to the disease-affected target tissue. The following is a discussion of Ad biology, barriers to Ad targeting and a review of the strategies applied toward increasing the targeting capacity of this delivery vector. Lastly, we propose unique future applications for Ad-based vectors that derive from recently established targeting and molecular imaging feasibilities reported herein.
Adenovirus structure
The family Adenoviridae contains, among others, 51 human Ad serotypes that are divided into six species (A–F) based on genome homology and organization, oncogenicity and hemagglutination properties.7-9 The human Ad is a nonenveloped icosahedral particle that encapsulates up to a 36-kilobase double-stranded DNA genome. The Ad capsid is comprised of several minor and three major capsid proteins: hexon is the most abundant structural component and constitutes the bulk of the protein shell; five subunits of penton form the penton base platform at each of the 12 capsid vertices to which the 12 fiber homotrimers attach (Figure 1). At the distal tip of each linear fiber is a globular knob domain, which serves as the major viral attachment site for cellular receptors. Hexon appears to play only a structural role as a coating protein, while the penton base and the fiber are responsible for virion–cell interactions that constitute Ad tropism. Detailed structures of hexon,10-12 penton base13 and fiber14,15 have been determined by crystallography; the high-resolution structure of the entire virion has been determined by various methods.16,17
Figure 1.
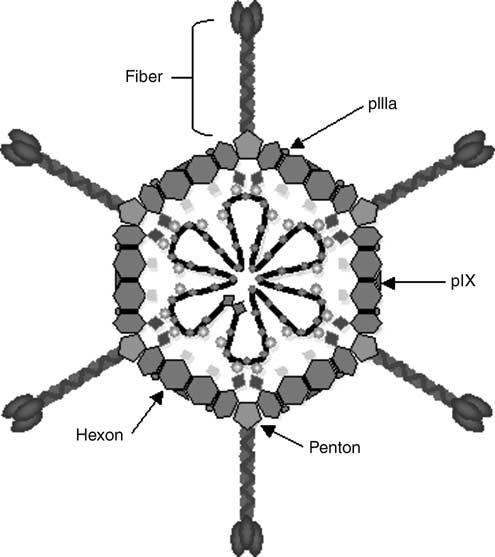
Generalized Ad structure depicting major structural components of a wild-type Ad capsid. Hexon, penton base, pIX, pIIIa and fiber structures are shown. Adenovirus capsids contain up to a 36-kilobase double-stranded DNA genome, shown as the dark line inside the capsid.
Adenovirus entry biology
Entry of Ad into cells involves two distinct steps: attachment to a primary receptor molecule at the cell surface, followed by interaction with molecules responsible for virion internalization. Initial high-affinity binding of the virion occurs via direct binding of the fiber knob domain to its cognate primary cellular receptor, which is the 46 kDa coxsackie and adenovirus receptor (CAR) for most serotypes, including Ad2 and Ad5, which are widely used in gene therapy approaches.18,19 Other receptors have been described for Ad5, although the nature of their interaction(s) with the Ad5 virion is unclear and their roles appear limited. These receptors include heparin sulfate glycosaminoglycans,20,21 class I major histocompatability complex22 and vascular cell adhesion molecule-1.23 Following receptor binding, receptor-mediated endocytosis of the virion is effected by interaction of penton base Arg–Gly–Asp (RGD) motifs with cellular integrins, including αvβ3 and αvβ5,24 αvβ1,25 α3b1 and α5β1.26 Virus enters the cell in clathrin-coated vesicles27 and is transported to endosomes. Subsequent acidification of the endosome results in virion disassembly and release of the virus remains into the cytosol, then to the nucleus where viral replication takes place.
Transductional targeting of Ad
This mechanistic understanding of Ad cellular entry explains clinical findings by numerous groups that have demonstrated that cells expressing low levels of CAR are refractory to Ad infection and gene delivery. This CAR dependency results in a scenario wherein nontarget but high-CAR cells can be infected, whereas target tissues, if low in CAR, remain poorly infected. Of key relevance to cancer gene therapy, increased CAR expression appears to have a growth-inhibitory effect on some cancer cell lines, while loss of CAR expression correlates with tumor progression and advanced disease. In addition, CAR has been shown to play a role in cell adhesion, and its expression may be cell cycle dependent.28,29 In short, while Ad delivery is uniquely efficient in vivo, CAR biodistribution is incompatible with many gene therapy interventions. We therefore hypothesized that if CAR expression could be induced in target tissues, resultant increases in Ad-mediated infection and gene expression would result in therapeutic gain. Indeed, a number of chemical agents predicted to affect cell cycle or cell adhesion were identified that increased CAR levels and Ad gene expression in ovarian cancer cells in vitro and in vivo.30
Adenovirus vector biodistribution in vivo, however, is not determined solely by receptor biodistribution.31 Intravenous administration of Ad results in accumulation in the liver, spleen, heart, lung and kidneys of mice, although these tissues may not necessarily be the highest in CAR expression.32,33 This is true with regard to the liver in particular, which sequesters the majority of systemically administered Ad particles via hepatic macrophage (Kupffer cell) uptake34 and hepatocyte transduction,35 leading to Ad-mediated inflammation and liver toxicity.36-39 Thus, the nature of Ad–host interactions dictating the fate of systemically applied Ad has come under considerable scrutiny.
Initial attempts to ‘de-target’ the liver were based on the supposition that CAR- and integrin-based interactions were required for liver transduction in vivo. Strategies to inhibit hepatocyte and/or liver Kupffer cell uptake by ablating CAR- or integrin-binding motifs in the Ad capsid have been largely unsuccessful, however, indicating that native Ad tropism determinants contribute little to vector hepatotropism in vivo.40-44 These data notwithstanding, work by several groups has implicated the fiber protein as a major structural determinant of liver tropism in vivo (reviewed by Nicklin et al.45). For example, shortening of the native fiber shaft domain of the Ad5 fiber46 or replacement of the Ad5 shaft with the short Ad3 shaft domain47 was shown to attenuate liver uptake in vivo following intravenous delivery. In related work, Smith et al.48 examined the role of a putative heparan sulfate proteoglycan (HSPG)-binding motif, KKTK, in the third repeat of the native fiber shaft. Replacement of this motif with an irrelevant peptide sequence reduced reporter gene expression in the liver by 90%. This was also the first indication of the importance of HSPG as an Ad receptor in vivo.
More recently, Shayakhmetov et al.49 uncovered a major role for coagulation factor IX (FIX) and complement component C4-binding protein (C4BP) in hepatocyte and Kupffer cell uptake of intravenous Ad. Indeed, Ad5 vectors containing fibers genetically modified to ablate FIX and C4BP binding provided 50-fold lower liver transduction with reduced inflammation and hepatotoxicity. Further analysis demonstrated that these blood factors mediated in vivo tropism by crosslinking Ad to hepatocellular HSPG and the low-density lipoprotein (LDL)-receptor-related protein. Kupffer cell sequestration of Ad particles was likewise heavily dependent on Ad association with FIX and C4BP.
These efforts serve to highlight the complexity of vector/host interplay, and have identified important genetic modifications that have important practical implications for designing safer and more effective Ad-based vectors for clinical applications. In the absence of a clinically defined upper limit for ectopic liver transduction in humans, it is clear that the concepts of ‘de-targeting’ and ‘re-targeting’ must be simultaneously employed to allow for maximum vector efficacy at the lowest possible dose. Therefore, engineering of targeted delivery with Ad requires the elimination of native tropism to be replaced by an alternative tropism based on targeting other receptor molecules, all while retaining innate Ad gene transfer efficiency. Two distinct approaches have been employed to transductionally target Ad-based therapeutic vectors: (1) adapter molecule-based targeting and (2) targeting achieved via structural manipulation of the Ad capsid via genetic means.
Adapter-based Ad targeting
The formation of a ‘molecular bridge’ between the Ad vector and a cell surface receptor constitutes the adapter-based concept of Ad targeting (Figure 2). Adapter function is performed by so-called ‘bi-specific’ molecules that crosslink the Ad vector to alternative cell surface receptors, bypassing the native CAR-based tropism. This approach is predicated by the aforementioned two-step entry mechanism of the Ad virion, wherein attachment is distinct from its ability to internalize into the targeted cell. In this way, alternative means of cellular attachment do not impede Ad cell entry. The majority of current adapter-based Ad targeting approaches incorporate the two mandates of delivery targeting, that of ablation of native CAR-dependent Ad tropism and formation of a novel tropism to previously identified cellular receptors. Bispecific adapter molecules include, but are not limited to: bi-specific antibodies, chemical conjugates between antibody fragments (Fab) and cell-selective ligands such as folate, Fab-antibody conjugates using antibodies against target cell receptors, Fab-peptide ligand conjugates and recombinant fusion proteins that incorporate Fabs and peptide ligands (Figure 3a).
Figure 2.
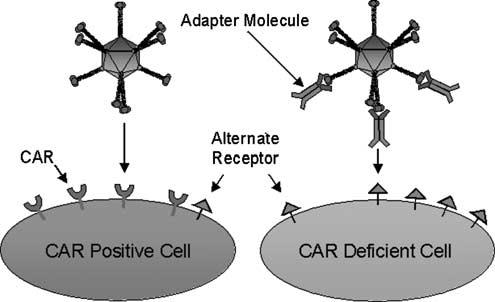
The Ad infection pathway. Shown are the basic steps of initial high-affinity binding of the virion fiber to its primary cellular receptor, CAR (left). Alternatively, a generalized adapter molecule ablates native CAR-based tropism and targets Ad to an alternate cellular receptor molecule (right). The dual specificity of the adapter molecule for both the Ad and the alternative receptor provides novel, CAR-independent cell binding.
Figure 3.
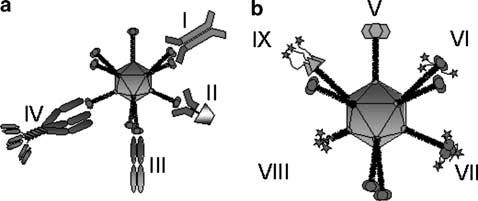
(a). Adenoviral particles can be re-targeted to TAAs, using bifunctional adapter molecules. Examples of such adapter molecules are chemical conjugates of a Fab fragment derived from an antiknob monoclonal antibody with a whole antibody directed against the target antigen (I), or naturally occurring ligands such as folate or basic fibroblast growth factor (II). The other class of adapter molecules consists of recombinant bispecific fusion proteins, examples of which include so-called diabodies, comprised of two single-chain antibodies (scFv), with one scFv recognizing the fiber knob and the other the TAA (III). Another example is the trimeric sCAR-fibritin-scFv, which uses the soluble ectodomain of the native Ad receptor CAR to bind to the Ad, thus ablating CAR recognition (IV). (b) Adenoviral particles can be re-targeted to TAAs using genetic targeting strategies in which the targeting ligands are incorporated into capsid proteins. The Ad5 knob protein can, for instance, be replaced with that of other serotypes (V), or it can be altered by incorporation of targeting ligands, such as RGD-containing peptide sequences or six histidine residues, at the C-terminus of the protein (VI). As an alternative to the C-terminus, peptide sequences can also be incorporated in the HI-loop of the Ad5 knob (VII). The entire fiber and knob domain can be replaced by an artificial fiber, for instance, consisting of the bacteriophage T4 fibritin trimerization domain and a targeting motif (VIII). Finally, multiple genetic modifications can be combined in a single particle, forming a so-called ‘complex mosaic Ad’. An example is a particle containing Ad3 knob instead of Ad5 knob, in which a six-histidine targeting motif is incorporated at the C-terminus (IX).
The first in vitro demonstration of Ad targeting via the adapter method resulted in CAR-independent, folate receptor-mediated cellular uptake of the virion by cancer cells overexpressing this receptor.50 This was accomplished using a bispecific conjugate consisting of an anti-knob neutralizing Fab chemically linked to folate. A similar targeting adapter comprised of the same anti-knob Fab as above fused to a basic fibroblast growth factor (FGF2) was utilized to target Ad vectors to FGF receptor-positive Kaposi's sarcoma (KS) cells in vitro.51 Importantly, this targeting system also reduced hepatic toxicity and resulted in increased survival in a melanoma xenograft mouse model.52 Other Fab–ligand conjugates targeted against Ep-CAM, Tag-72, epidermal growth factor (EGF) receptor, CD-40 and other cell markers have been employed in a similar manner with promising results.1,53-58
Dmitriev et al.60 developed an elegant alternative to the chemical conjugate approach by creating a single recombinant fusion molecule formed by a truncated, soluble form of CAR (sCAR) fused to either an anti-CD40 antibody59 or epidermal growth factor (EGF). Using the latter, a nine-fold increase in reporter gene expression was achieved in several EGFR-overexpressing cancer cell lines compared to untargeted Ad or EGFR-negative cells in vitro. EGF-directed targeting to EGFR-positive cells was shown to be dependent on cell surface EGFR density, an additional confirmation of Ad targeting specificity. Addressed also was the issue of virion/adapter complex stability, a critical issue if targeting adapters are to be employed in vivo. In this regard, preformed Ad/sCAR–EGF complexes subjected to gel filtration purification showed the same targeting profile as those not purified, indicating adequate Ad/adapter complex stability. To further increase Ad/sCAR–ligand complex stability, Kashentseva et al.61 developed a trimeric sCAR-antic-erbB2 single-chain antibody adapter molecule. While untested as yet in vivo, the trimeric sCAR-c-erbB2 adapter displayed increased affinity for the Ad fiber knob, while augmenting gene transfer up to 17-fold in six c-erbB2-positive breast and ovarian cancer cell lines in vitro. In similar work, Itoh et al.62 demonstrated improved efficiency of sCAR-based fusion molecule adapters. Kim et al.63 showed that adapter trimerization yielded a 100-fold increase in infection of CAR-deficient human diploid fibroblasts compared to the monomeric sCAR adapter. Importantly, in vivo employment of a nontargeted trimeric sCAR adapter attenuated liver transduction in mice following intravenous administration, indicating the excellent in vivo stability of this Ad/trimeric adapter complex. These studies have supplied preliminary evidence of the systemic stability of adaptor–virus complexes, although it is likely that clinical trials will ultimately be needed to investigate this crucial aspect for individual agents.
The above proof-of-principle studies and others have rationalized the further testing of targeting adapters in vivo. In this regard, Reynolds et al.56 employed a novel bispecific adapter composed of an anti-knob Fab chemically conjugated to a monoclonal antibody (9B9) raised against angiotensin-converting enzyme (ACE), a surface molecule expressed preferentially on pulmonary capillary endothelium and upregulated in various disease states of the lung. Following peripheral intravenous injection of the Ad/Fab–9B9 complex, reporter transgene expression and viral DNA in the lung were increased 20-fold over untargeted Ad. Importantly, reporter gene expression in the liver, a nontarget, high-CAR organ, was reduced by 83%. Further, in a unique adapter-based in vivo lung-targeting schema, Everts et al. used a bifunctional adapter molecule comprised of sCAR fused to a single-chain antibody (MFE-23) directed against carcinoembryonic antigen (CEA). Systemic administration of the Ad/sCAR–MFE-23 adapter complex increased gene expression in CEA-positive murine lung by 10-fold and reduced liver transduction, resulting in an improved lung-to-liver ratio of gene expression compared with untargeted Ad.
Overall, adapter-based Ad targeting studies provide compelling evidence that Ad tropism modification can be achieved by targeting alternate cellular receptors and that this modality augments gene delivery to CAR-deficient target cells in vitro. Adapter-targeted vectors have also performed well in vivo, although data so far are limited. While single-component systems have been favored for employment in human gene therapy trials, rigorous analysis of the pharmaco-dynamics and -kinetics, and systemic stability of vector/adapter complexes could potentially provide the rationale for clinical translation.
Adenovirus targeting via genetic modification: fiber
Genetic manipulation of capsid proteins has yielded increasingly promising data in terms of Ad targeting. Redirection of Ad tropism via genetic capsid modification is conceptually elegant, but genetic targeting efforts must work within narrow structural constraints. The success of this approach depends upon modulation of the complex protein structure/function relationships that result in Ad tropism modification, without disrupting the innate molecular interactions required for proper biological function. Based on a clear understanding of Ad infection biology, development of genetically targeted vectors has rationally focused on the fiber, the primary capsid determinant of Ad tropism. In general, there have been three basic strategies for genetic tropism modification via structural modification of the Ad fiber: (1) so-called ‘fiber pseudotyping’; (2) ligand incorporation into the fiber knob and (3) ‘de-knobbing’ of the fiber coupled with ligand addition (Figure 3b).
As previously mentioned, many clinically relevant tissues are refractory to Ad5 infection, including several cancer cell types, due to negligible CAR levels. Adenovirus fiber pseudotyping, the genetic replacement of either the entire fiber or knob domain with its structural counterpart from another human Ad serotype that recognizes a cellular receptor other than CAR, was first accomplished by Krasnykh et al.64 These vectors display CAR-independent tropism by virtue of the natural diversity in receptor recognition found in human subgroup B and D fibers.65 In this regard, primary receptors for subgroup B Ads have been recently identified, including the complement regulatory protein CD46,66-69 CD80 and CD86,70 although an additional unknown receptor is postulated.67 Subgroup D Ad receptors include CD46 and α(2–3)-linked sialic acid, a common element of glycolipids.71-73 This fiber pseudotyping approach has identified chimeric vectors with superior infectivity to Ad5 in several clinically relevant cell types, including primary ovarian carcinoma cells,74-76 vascular endothelial cells,77 dendritic cells,78 B-cells,79 CD34 + hematopoietic cells,80 synovial tissue,81 human cardiovascular tissue82 and others.83,84 Interestingly, this strategy has been extended to exploit fiber elements from non-human Ads85,86 and the fiber-like σ1 reovirus attachment protein, which targets cells expressing junctional adhesion molecule.87,88
Direct ligand incorporation into the Ad knob domain without ablating native CAR-binding has resulted in Ad vectors with expanded, rather than restricted, cell recognition. These efforts are based on rigorous structural analysis of the knob domain and have exploited two separate locations within the knob that tolerate genetic manipulation without loss of fiber function, the C-terminus and the HI-loop. Since the C-terminus of the Ad knob is solvent exposed, extension of the knob peptide to include a targeting peptide moiety is conceptually simple. Ads with C-terminal integrin-binding RGD motifs and poly-lysine ligands have yielded some promising results in vitro and in vivo, but other peptide ligands were rendered ineffective in the C-terminus structural context,89 presumably due to steric or other inhibition. Krasnykh et al.90 inserted a FLAG peptide sequence into an exposed loop structure that connects β-sheets H and I (HI-loop) within the Ad5 knob, showing that this locale is structurally permissive to modification. Indeed, the Ad5 HI-loop tolerates peptide insertions up to 100 amino acids with minimal negative effects on virion integrity, thus suggesting considerable potential for ligand incorporation at this site.91 Dmitriev et al.4 introduced an integrin-binding RGD peptide sequence into the HI-loop. The resulting vector, Ad5lucRGD, used the RGD/cellular integrin interaction to enhance gene delivery to ovarian cancer cell lines and primary tumors versus unmodified Ad.92,93 The expanded tropism of this vector has been useful in several other cancer contexts including carcinomas of the ovary, pancreas, colon cancer, and head and neck carcinomas, all of which frequently display highly variable CAR levels.94 Wu et al.95 demonstrated that Ad vectors with a double fiber modification consisting of a C-terminal poly-lysine stretch, which interacts with heparan sulfates, and the HI-loop RGD provided increased infectivity in several CAR-deficient cell lines, as well as human pancreatic islet cells,96 ovarian carcinoma97 and cervical cancer cells in vivo.98 Other targeting peptides have functioned in the HI-loop locale, including a vascular endothelial cell-binding motif SI-GYLPLP.99 This fiber modification also provided cancer cell selective infection.100
Korokhov et al.,101 Volpers et al.102 and others103 have developed similar targeting approaches that embody elements of both genetic fiber modification and adapter-based targeting by incorporating the immunoglobulin (Ig)-binding domain of Staphylococcus aureus protein A into the fiber C-terminus or HI-loop. As a result, these fiber-modified vectors form stable complexes with a wide variety of targeting molecules containing the Fc region of Ig. This provides the opportunity to screen numerous targeting molecules directed against a host of cell-surface elements. This approach was used to target and activate dendritic cells via an Fc-single-chain antibody directed against CD40.104 This system was also used to target ovarian cancer cells via an antibody directed against mesothelin,105 as well as the pulmonary endothelium in a rat model in vitro.106
The structural conflicts emerging from knob modifications and the observation that fiber-deleted Ad vectors could be produced107,108 provided the conceptual basis for replacing the native fiber with knobless fibers. Virions containing a knobless fiber would be ablated for CAR binding, a hallmark of targeted Ad vectors. Simultaneous addition of a targeting ligand to the knobless fiber would result in a more specifically targeted Ad. The technical barrier to this approach is the innate trimerization function of the knob, required for proper fiber function and capsid incorporation. To overcome this structural conflict, addition of foreign trimerization motifs have been used to replace the native fiber and/or knob.109 Krasnykh et al.110 replaced the fiber and knob domains with bacteriophage T4 fibritin containing a C-terminal 6-His motif. This novel Ad variant lacks the ability to interact with CAR and demonstrated up to a 100-fold increase in reporter gene expression to cells presenting an artificial 6-His-binding receptor. A similar ‘de-knobbing’ strategy was employed by Magnussen et al.,111 wherein an integrin-binding RGD motif was utilized, resulting in selective infection of integrin-expressing cell lines in vitro, as well as human glandular cells.112 Based on the feasibility of fiber replacement with T4 fibritin, an elegant system was devised wherein the trimeric CD40 ligand was fused to the C-terminus of this artificial fiber.113 Notably, this vector provided CD40-specific gene delivery in vivo following systemic delivery.114 Further, this vector accomplished CD40-mediated infection of human monocyte-derived dendritic cells, suggesting a possible utility for cancer immunotherapy ‘antigen-loading’ approaches. In addition, Ad vectors simultaneously incorporating multiple fiber types with distinct receptor specificities have been proposed.115,116
Given the target specificity demonstrated by antibody-mediated targeting of Ad vectors using adapter molecules, the development of single-component Ad vectors with genetically incorporated antibodies, antibody-derived moieties or other multi-domain ligands has been a longstanding field milestone. Genetic capsid incorporation of any moiety requires that the heterologous peptide be compatible with the nonreducing environment within the cytosol and nucleus, wherein Ad capsid proteins are translated and assembled. Indeed, capsid incorporation of several classes of complex-targeting ligands, including scFv and growth factors, has been severely hampered by the innate biosynthetic incompatibilities between the ligand and Ad capsid proteins, resulting in unstable or insoluble ligands and/or reduced Ad replication.117 On this basis, rational development of complex ligands with cytoplasmic solubility and stability will be required for their application to Ad vectors. Exemplifying this concept, Hedley et al.118 have developed a single-component antibody-targeted Ad vector by incorporating a novel, cytoplasmically stable scFv into a de-knobbed fiber. This vector demonstrated selective targeting to its cognate epitope expressed on the membrane surface of cells, and suggests that cytoplasmic stability of the targeting molecule, per se, allows retention of antigen recognition in the Ad capsid-incorporated context.
Adenovirus targeting via genetic modification: other capsid locales
The field-wide appreciation of the difficulty of incorporating complex ligands into the Ad fiber locale has prompted the identification of other capsid proteins amenable to ligand incorporation via genetic manipulation. These approaches have the potential, through increased capsid valency and unique capsid microenvironments, to incorporate an increased number of complex ligands per virion. To date, capsid protein hexon as well as minor capsid proteins polypeptide IX (pIX) and pIIIa have been used as platforms for incorporation of heterologous peptides.
Hexon is the largest and most abundant capsid protein, and as such is an attractive locale for peptide ligand incorporation due to both its surface exposure and high valency (240 hexon homotrimers per virion). The primary sequence of the hexon monomer is highly conserved among human serotypes, with the exception of nine nonconserved hypervariable regions (HVRs) of unknown function found mainly within solvent-exposed loops at the surface.11,119,120 In this regard, Vigne et al.121 exploited hexon hypervariable region 5 (HVR5), a loop structure in hexon, as a site for incorporation of an integrin-binding RGD motif. Notably, virion stability was unaffected by the addition of the foreign peptide, while providing enhanced, fiber-independent transduction to low-CAR vascular smooth muscle cells. Extending this work, Wu et al. identified HVRs 2, 3 and 5–7 as hexon sites tolerating 6-histidine (6-His) motifs without adverse affects to virion formation or stability. 6-His motifs in HVRs 2 and 5 mediated virion binding to anti-6-His antibodies; however, 6-His-mediated viral infection of cells was not observed, in contrast to Vigne et al. above, highlighting the importance of the nature and the length of the incorporated peptides.
Recently, pIX has emerged as a versatile capsid locale well suited for display of ligands with utility for both targeting and imaging modalities.122 The 14.3 kDa pIX is the smallest of the minor capsid proteins, a subset of capsid proteins that generally function to stabilize the capsid shell. In the mature Ad virion, 80 pIX homo-trimers123 stabilize hexon–hexon interactions during capsid assembly, and it is therefore termed a ‘cement’ protein. Indeed, virions deleted for pIX have decreased thermostability and a DNA capacity that is approximately 2 kb less than the normal length.124-126 Interest in employing pIX as a capsid site for incorporation of peptide ligands stemmed from the observation that the C-terminus of pIX is located at the capsid surface,127,128 which prompted several groups to explore the fusion of several polypeptides to this terminus.
Dmitriev et al.129 first reported the incorporation of functional targeting peptides at the pIX C-terminus by inserting poly-lysine or FLAG motifs, resulting in augmented, CAR-independent gene transfer via binding to cellular heparan sulfate moieties. In a similar approach, Vellinga et al.130 fused an integrin-binding RGD peptide to pIX, and used α-helical spacers (up to 7.5 nm in length and 113 amino acids) to extend the RGD motif away from the virion surface. Increased gene transfer to CAR-deficient endothelioma cells was observed with increased spacer length, giving support to the notion that the pIX C-terminus may reside in a cavity formed by surrounding hexons. In order to evaluate the utility of multiple ligand types at the pIX capsid locale, Campos et al.131 fused to the pIX-C-terminus a 71-amino-acid fragment of the Propionibacterium shermanii 1.3S transcarboxylase protein, which functions a biotin acceptor peptide (BAP). During virus propagation the BAP is metabolically biotinylated, rendering this virus compatible with a host of avidin-tagged ligands, including peptides, antibodies and carbohydrates. Importantly, it was noted in this study that coupling transferrin to virions via pIX-BAP resulted in specific transferrin receptor-mediated infection of C2C12 cells, but the use of an antibody directed against the transferrin receptor (CD71) did not. This dichotomy was not observed when these two ligands successfully redirected Ad tropism when incorporated into fiber. The authors speculate that the difference was not due to a lack of receptor recognition by the pIX–anti-CD71 complex, but rather a difference between the dissociation of targeted fiber and pIX from the Ad particle in endosomes, resulting in trapping of the pIX–anti-CD71 variant, but not the fiber–anti-CD71 in the endosome. If this notion is fully validated, it will represent a key finding showing that the nature of pIX-incorporated ligands may influence successful redirection of Ad infection.
Polypeptide IX has also been in use for the display of relatively large imaging molecules as C-terminal fusions. While Ad vector imaging is beyond the scope of this work, the successful incorporation of the 240-amino-acid enhanced green fluorescent protein (eGFP) into pIX bears mentioning due to the size and complexity of this fusion. Of note, the presence of the pIX–eGPF fusion in purified Ad virions did not appreciably decrease virus viability or capsid stability, and has allowed monitoring of Ad localization in vitro and in vivo.132,133 Further, other Ad vectors harboring complex ligands at the pIX locale have been reported.134-136 As a whole, these studies have established pIX as a highly relevant capsid locale marked by the highest structural compatibility, with diverse targeting and imaging ligands observed to date.
Based on its surface capsid position, pIIIa has been proposed as a platform for ligand display for modification of Ad tropism.137 High-resolution imaging techniques originally indicated that pIIIa is an elongated protein penetrating the capsid and is located along the icosahedral edges of the virion.16 More recent structural studies performed at higher resolution place pIIIa only at the surface of the virion.17 The minor capsid protein pIIIa is a 67-kDa monomer that is cleaved at the C-terminus during maturation of the virion, giving rise to the final 63.5 kDa form. To evaluate the utility of pIIIa for ligand display, Dmitriev et al.138,139 have incorporated a 6-His motif singly into the N-terminus of pIIIa. The modified pIIIa proteins were well tolerated in mature virions, although whether pIIIa ligands can redirect Ad tropism remains to be conclusively demonstrated.
In the aggregate, these results highlight that genetic manipulation of a variety of Ad capsid proteins is currently feasible, and has brought to fruition several novel targeting and imaging paradigms. These successes confirm a level of capsid flexibility that was largely unexpected. There remains, however, an ongoing struggle to identify true targeting ligands that are structurally and/or biosynthetically compatible with Ad capsid formation and stability.
Identification of new tumor-associated antigens
Targeted gene delivery is ultimately predicated on the ability of the vector to discriminate between target cell or tissues types via interaction with unique cell- or disease-specific surface markers. On this basis, the discovery and characterization of novel cell-surface targets is of paramount importance. In the context of targeted gene delivery for cancer, considerable effort is being expended to identify new tumor-associated antigens (TAAs) for a variety of cancers, utilizing advanced technologies such as mRNA microarrays, proteomics and ‘serological identification of antigens by recombinant expression cloning’ (SEREX). The identification of more than 20 000 genes by the human genome project has provided a pool of possible new targets in cancer, and the screening for those has become significantly more facile in recent years due to commercialization and increased availability of mRNA microarray technology. Several new targets have been identified using this technology, such as the carcinoma-associated antigen GA733-2, and hepsin, a transmembrane serine protease found in prostate cancer.140 Another example is the high expression of ‘preferentially expressed antigen of melanoma’ gene (PRAME), detected by microarray in ovarian cancer samples.141
Compared to the number of genes present in the human genome, the number of possible protein targets is increased several-fold due to the intermediate steps between mRNA and protein expression, and the protein variability potentially introduced with each of these steps. These variabilities include mRNA splicing, post-translational processing, glycosylation, as well as others. The Human Proteome Organization (HUPO), formed in 2001, aims to systematically characterize protein expression in health and disease, and its Plasma Proteome Project is a prime example in which several laboratories throughout the world participate to identify tumor markers that can be used for both screening as well as therapeutic targets.142 The mRNA and protein array technologies can also be combined in order to discover new biomarkers, such as demonstrated by Nishizuka et al.,143 who identified villin as a new marker for colon cancer using this combination.
Another method to identify new tumor targets is ‘serological identification of antigens by recombinant expression cloning’ (SEREX), which was first described by Sahin et al.144 in 1995 and is based on the presence of TAA recognizing antibodies in the serum of cancer patients. In this approach, a cDNA library is constructed from tumor specimens and cloned into expression vectors. Clones are then screened for reactivity with the serum of the autologous patient, and the nucleotide sequence of the cDNA insert is determined.144 In previous years, several antigens that can be classified into different groups have been identified, including cancer-testis antigens, differentiation antigens, overexpressed gene products, mutated gene products, splice variants and cancer-related auto-antigens. Examples of antigens discovered using this technology are melanoma antigen gene-1 (MAGE-1) in melanoma and metastasis-associated protein-1 (MTA1) in prostate cancer.145
Identification of new ligands
In addition to the identification of new TAAs, new ligands that are able to target these antigens and are suitable for incorporation into adenoviral vectors need to be identified as well. One of such technologies is phage display, which was first pioneered by Smith146 in 1985, that can identify high-affinity peptides for a target receptor of choice. As an example, Peletskaya et al.147,148 screened a 15-mer library for peptides able to bind the Thomson–Friedenreich (TF) antigen, a glycoantigen present on several carcinomas, and identified peptides with an affinity around 1 μm. One of the most well-known examples of a peptide identified by phage display is the RGD amino-acid sequence, which has a high affinity for integrins that are expressed in tumor vasculature. This peptide, identified by the group of Ruoslathi149 in 1993, has extensively been used in drug- or gene-targeting approaches, either a linear or cyclic format. For example, Schiffelers et al.150 coupled this peptide to a liposome carrier, demonstrating selective localization of this drug-targeting construct in angiogenic vasculature in a dorsal skin chamber model, whereas genetic incorporation of RGD in both adenoviral and adeno-associated viral vectors has led to a dramatic infectivity enhancement of these vectors in a variety of cancer cells.4,151
Since some promising targeting ligands lose their receptor specificity upon incorporation into adenoviral capsid proteins, it would be useful to screen these ligands in their appropriate context in a high-throughput manner. Pereboev et al.152 have elegantly approached this problem by modifying the pJuFo phage system in such a way that the (modified) Ad5 knob is expressed on the surface of filamentous bacteriophage, allowing screening of incorporated ligands with traditional phage display techniques. This system may prove very useful in screening for candidate ligand fidelity and Ad structural compatibility, prior to genetic Ad modification.
Future applications
Despite the advances in tumor-targeted gene therapy as described in this article, several obstacles remain. The vascular endothelial wall is a significant physical barrier prohibiting access of systemically administered vectors to the tumor cell. To overcome this obstacle, strategies are currently being endeavored to route adenoviral vectors via transcytosis pathways through the endothelium. As an example, Zhu et al.153 redirected Ad vectors to the transcytosing transferrin receptor pathway, using the bifunctional adapter molecule. The transcytosed Ad virions retained the ability to infect cells, establishing the feasibility of this approach. However, efficiency of Ad trafficking via this pathway is poor, and current efforts are directed towards exploring other transcytosing pathways such as the melanotransferrin pathway,154 the poly-Ig A receptor pathway,155 or caveolae-mediated transcytosis pathways.156 One can envision the development of mosaic Ad vectors incorporating both targeting ligands directed to such transcytosis pathways as well as ligands mediating subsequent targeting and infection of tumor cells present beyond the vascular wall.
The often-limited therapeutic effects achieved by gene or virotherapy approaches mandate the development of alternative ‘amplifying strategies’, in which delivery of a vector to a single cell will result in the destruction of its neighboring cells as well. In this regard, the development of so-called multifunctional particles (MFPs) that incorporate targeting, imaging and amplifying capacities is an exciting possible new area of investigation (Figure 4). Adenovirus-based vectors are ideally positioned to represent such an MFP, by virtue of genetic capsid modifications, to incorporate functionalities as cited above. For example, incorporation of genetically modified fibers, combined with imaging motifs on the pIX protein, could be used to simultaneously target tumor cells while monitoring viral replication and spread. In this regard, in addition to optical imaging-based ligands that have been incorporated at pIX as mentioned above, recently Li et al.136 have been successful in incorporating the enzyme Herpes Simplex Virus thymidine kinase (HSV-tk) at this capsid locale. This enzyme is compatible with available PET imaging ligands such as 18F-penciclovir, providing an imaging system for viral replication that can directly be translated for clinical applications. Interestingly, HSV-tk is an enzyme that has utility in so-called suicide gene therapy, in which the expressed enzyme converts a substrate such as ganciclovir to its toxic phosphorylated metabolite, resulting in cell death.157 Also, tumor cells expressing this gene product induce the death of adjacent cells via the so-called ‘bystander effect’, thus representing an ‘amplifying strategy’ as mentioned above. Incorporation of this enzyme on the capsid surface of a targeted Ad could be exploited in an amplifying strategy for the induction of cell death, in addition to its use for imaging.
Figure 4.
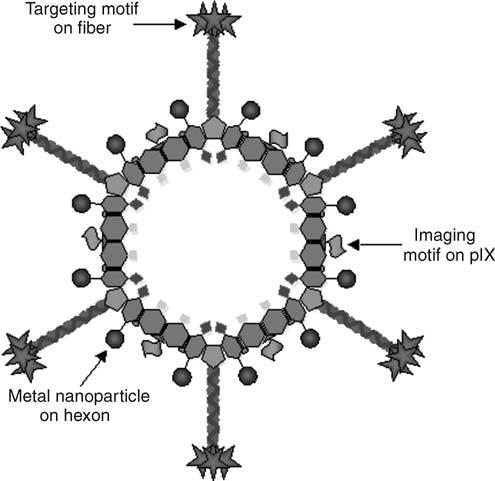
Schematic diagram showing an idealized MFP based on an empty Ad capsid. Adenoviral vectors can theoretically be used as a platform for the development of MFPs incorporating targeting, imaging and therapeutic motifs. For example, targeting motifs can be incorporated into the fiber protein, whereas simultaneous incorporation of imaging motifs such as thymidine kinase on the pIX protein will provide the ability to monitor particle and thus tumor localization, utilizing noninvasive technologies such as PET. Incorporation of metal nanoparticles with magnetic properties into such a platform will provide additional imaging opportunities via MRI, as well as therapeutic opportunities via hyperthermia induction.
The concept of MFPs has also been introduced recently in the context of nanotechnology, often defined as the development of devices of 100 nm or smaller, having unique properties due to their scale. Applications in medicine are starting to emerge, with ‘nanomedicine’ recently being defined as the utilization of nanotechnology for treatment, diagnosis, monitoring and control of biological systems. The devices that are being developed generally incorporate inorganic or biological material. In this regard, the coupling of inorganic nanoscale materials to targeted Ad vectors would expand MFP functions, capitalizing on the new treatment strategies being developed using nanotechnology. For example, magnetic nanoparticles have recently received much attention due to their potential application in clinical cancer treatment,158-162 targeted drug delivery163-165 and MRI contrast agents.166,167 However, despite the useful functionalities that might derive from metal nanoparticle systems, the lack of targeting strategies has limited their application to locoregional disease. Thus, tumor-selective delivery is key to improve therapeutic applications of metal nanoparticle systems, rationalizing incorporation of such particles into targeted, multifunctional Ad vectors via conjugation to capsid proteins or other means.
For the development of targeted MFPs, it would be beneficial to minimize the adenoviral vector to empty capsid shells, based on the consideration that the so-called first-generation Ad vectors (i.e. Ad5- or Ad2-based nonreplicating viruses lacking E1, and sometimes E3 regions) still express low levels of the remaining viral genes. This low level of viral gene expression leads to direct toxicity and immunogenicity. Consequently, duration of transgene expression is shortened as the immune response clears the vector-transduced cells.168,169 To overcome these problems, high-capacity Ad vectors have been developed. The only viral genomic elements retained in these vectors are the inverted terminal repeats (ITR) and packaging signal. This allows up to 36 kb of nonviral DNA to be inserted. Reduction in in vivo toxicity and immunogenicity are seen,170-173 and in mice long-term transgene expression for more than a year has been observed.173 The deletion of all the viral genes requires helper viruses to trans-complement all viral genes, including those for capsid proteins. Well-defined and efficient systems have been developed to allow production of high-capacity adenoviral vectors without contamination of helper virus, based on recombinase-mediated excision of the helper-virus packaging signal.174-176 It is logical that the capsid proteins expressed by the helper virus would be genetically modified, allowing re-targeting of the resultant high-capacity adenoviral vector. This concept was recently demonstrated by using the RGD motif in the HI-loop of the fiber knob,177 and this demonstration of combining re-targeting with high-capacity Ad illustrates the potential for utilization of gutless Ad as a nanoscale platform.
To extend the paradigm of limiting foreign gene expression, high-capacity adenoviral vectors have been further developed into empty Ad capsid shells.178,179 The production of empty capsids has been based on a modification of the systems utilized in the construction of high-capacity Ad vectors, utilizing a Cre/lox recombination event. It has been demonstrated that, compared to recombinant Ad vectors, empty Ad capsid shells induce a significantly reduced number of genes after cellular interaction.179 This suggests that these empty capsid shells could serve as an improved platform for the development of MFP systems, which would incorporate targeting, imaging and therapeutic elements, with minimal adverse effects after in vivo administration.
Summary
Adenovirus-based vectors are the most widely used platform for gene delivery. They are of particular utility for cancer gene therapy applications, where temporary gene expression is acceptable or even beneficial. The history of Ad-based gene therapy studies clearly illustrates and confirms the critical linkage between an improved delivery vector and increases in therapeutic potential. Very often, clinical breakthroughs have been dependent on advances in vector development. With regard to Ad-mediated cancer treatment, high-level tumor transduction remains a key developmental hurdle. To this end, Ad vectors possessing infectivity enhancement and targeting capabilities should be evaluated in the most stringent model systems possible. Advanced Ad-based vectors with imaging, targeting and therapeutic capabilities have yet to be fully realized; however, the feasibilities leading to this accomplishment are within close reach.
Acknowledgements
This work was supported by grants from the National Institutes of Health: R01 CA083821, R01 CA094084 and R01 CA111569-01A1, and grant W81XWH-05-1-0035 from the US Department of Defense. We wish to thank Dr Long P Le for assistance with graphic images.
References
- 1.Miller CR, Buchsbaum DJ, Reynolds PN, Douglas JT, Gillespie GY, Mayo MS, et al. Differential susceptibility of primary and established human glioma cells to adenovirus infection: targeting via the epidermal growth factor receptor achieves fiber receptor-independent gene transfer. Cancer Res. 1998;58:5738–5748. [PubMed] [Google Scholar]
- 2.Li Y, Pong RC, Bergelson JM, Hall MC, Sagalowsky AI, Tseng CP, et al. Loss of adenoviral receptor expression in human bladder cancer cells: a potential impact on the efficacy of gene therapy. Cancer Res. 1999;59:325–330. [PubMed] [Google Scholar]
- 3.Hemminki A, Alvarez RD. Adenoviruses in oncology: a viable option? Bio Drugs. 2002;16:77–87. doi: 10.2165/00063030-200216020-00001. [DOI] [PubMed] [Google Scholar]
- 4.Dmitriev I, Krasnykh V, Miller CR, Wang M, Kashentseva E, Mikheeva G, et al. An adenovirus vector with genetically modified fibers demonstrates expanded tropism via utilization of a coxsackievirus and adenovirus receptor-independent cell entry mechanism. J Virol. 1998;72:9706–9713. doi: 10.1128/jvi.72.12.9706-9713.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Cripe TP, Dunphy EJ, Holub AD, Saini A, Vasi NH, Mahller YY, et al. Fiber knob modifications overcome low, heterogeneous expression of the coxsackievirus-adenovirus receptor that limits adenovirus gene transfer and oncolysis for human rhabdomyosarcoma cells. Cancer Res. 2001;61:2953–2960. [PubMed] [Google Scholar]
- 6.Clinicals Trials Database The Journal of Gene Medicine Clinical Trials Database. 2004 http://www.wiley.co.uk/genmed/clinical. [Google Scholar]
- 7.Davison AJ, Benko M, Harrach B. Genetic content and evolution of adenoviruses. J Gen Virol. 2003;84(Part 11):2895–2908. doi: 10.1099/vir.0.19497-0. [DOI] [PubMed] [Google Scholar]
- 8.Mei YF, Wadell G. Epitopes and hemagglutination binding domain on subgenus B:2 adenovirus fibers. J Virol. 1996;70:3688–3697. doi: 10.1128/jvi.70.6.3688-3697.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Shenk T. Adenoviridae: and their replication. In: Fields B, Howley P, Knipe D, editors. Virology. Raven Press; New York: 1996. pp. 2111–2148. (translator and editor) [Google Scholar]
- 10.Stewart PL, Burnett RM. Adenovirus structure by X-ray crystallography and electron microscopy. Curr Top Microbiol Immunol. 1995;199(Part 1):25–38. doi: 10.1007/978-3-642-79496-4_2. [DOI] [PubMed] [Google Scholar]
- 11.Rux JJ, Kuser PR, Burnett RM. Structural and phylogenetic analysis of adenovirus hexons by use of high-resolution x-ray crystallographic, molecular modeling, and sequence-based methods. J Virol. 2003;77:9553–9566. doi: 10.1128/JVI.77.17.9553-9566.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Roberts MM, White JL, Grutter MG, Burnett RM. Three-dimensional structure of the adenovirus major coat protein hexon. Science. 1986;232:1148–1151. doi: 10.1126/science.3704642. [DOI] [PubMed] [Google Scholar]
- 13.Zubieta C, Schoehn G, Chroboczek J, Cusack S. The structure of the human adenovirus 2 penton. Mol Cell. 2005;17:121–135. doi: 10.1016/j.molcel.2004.11.041. [DOI] [PubMed] [Google Scholar]
- 14.van Raaij MJ, Louis N, Chroboczek J, Cusack S. Structure of the human adenovirus serotype 2 fiber head domain at 1.5 A resolution. Virology. 1999;262:333–343. doi: 10.1006/viro.1999.9849. [DOI] [PubMed] [Google Scholar]
- 15.van Raaij MJ, Mitraki A, Lavigne G, Cusack S. A triple beta-spiral in the adenovirus fibre shaft reveals a new structural motif for a fibrous protein. Nature. 1999;401:935–938. doi: 10.1038/44880. [DOI] [PubMed] [Google Scholar]
- 16.Stewart PL, Fuller SD, Burnett RM. Difference imaging of adenovirus: bridging the resolution gap between X-ray crystallography and electron microscopy. EMBO J. 1993;12:2589–2599. doi: 10.1002/j.1460-2075.1993.tb05919.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Fabry CM, Rosa-Calatrava M, Conway JF, Zubieta C, Cusack S, Ruigrok RW, et al. A quasi-atomic model of human adenovirus type 5 capsid. EMBO J. 2005;24:1645–1654. doi: 10.1038/sj.emboj.7600653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Tomko RP, Xu R, Philipson L. HCAR and MCAR: the human and mouse cellular receptors for subgroup C adenoviruses and group B coxsackieviruses. Proc Natl Acad Sci USA. 1997;94:3352–3356. doi: 10.1073/pnas.94.7.3352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bergelson JM, Cunningham JA, Droguett G, Kurt-Jones EA, Krithivas A, Hong JS, et al. Isolation of a common receptor for Coxsackie B viruses and adenoviruses 2 and 5. Science. 1997;275:1320–1323. doi: 10.1126/science.275.5304.1320. [DOI] [PubMed] [Google Scholar]
- 20.Dechecchi MC, Melotti P, Bonizzato A, Santacatterina M, Chilosi M, Cabrini G. Heparan sulfate glycosaminoglycans are receptors sufficient to mediate the initial binding of adenovirus types 2 and 5. J Virol. 2001;75:8772–8780. doi: 10.1128/JVI.75.18.8772-8780.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Dechecchi MC, Tamanini A, Bonizzato A, Cabrini G. Heparan sulfate glycosaminoglycans are involved in adenovirus type 5 and 2-host cell interactions. Virology. 2000;268:382–390. doi: 10.1006/viro.1999.0171. [DOI] [PubMed] [Google Scholar]
- 22.Hong SS, Karayan L, Tournier J, Curiel DT, Boulanger PA. Adenovirus type 5 fiber knob binds to MHC class I alpha2 domain at the surface of human epithelial and B lymphoblastoid cells. EMBO J. 1997;16:2294–2306. doi: 10.1093/emboj/16.9.2294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Chu Y, Heistad D, Cybulsky MI, Davidson BL. Vascular cell adhesion molecule-1 augments adenovirus-mediated gene transfer. Arterioscler Thromb Vasc Biol. 2001;21:238–242. doi: 10.1161/01.atv.21.2.238. [DOI] [PubMed] [Google Scholar]
- 24.Wickham TJ, Mathias P, Cheresh DA, Nemerow GR. Integrins alpha v beta 3 and alpha v beta 5 promote adenovirus internalization but not virus attachment. Cell. 1993;73:309–319. doi: 10.1016/0092-8674(93)90231-e. [DOI] [PubMed] [Google Scholar]
- 25.Li E, Brown SL, Stupack DG, Puente XS, Cheresh DA, Nemerow GR. Integrin alpha(v)beta1 is an adenovirus coreceptor. J Virol. 2001;75:5405–5409. doi: 10.1128/JVI.75.11.5405-5409.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Davison E, Diaz RM, Hart IR, Santis G, Marshall JF. Integrin alpha5beta1-mediated adenovirus infection is enhanced by the integrin-activating antibody TS2/16. J Virol. 1997;71:6204–6207. doi: 10.1128/jvi.71.8.6204-6207.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Meier O, Boucke K, Hammer SV, Keller S, Stidwill RP, Hemmi S, et al. Adenovirus triggers macropinocytosis and endosomal leakage together with its clathrin-mediated uptake. J Cell Biol. 2002;158:1119–1131. doi: 10.1083/jcb.200112067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Okegawa T, Pong RC, Li Y, Bergelson JM, Sagalowsky AI, Hsieh JT. The mechanism of the growth-inhibitory effect of coxsackie and adenovirus receptor (CAR) on human bladder cancer: a functional analysis of car protein structure. Cancer Res. 2001;61:6592–6600. [PubMed] [Google Scholar]
- 29.Seidman MA, Hogan SM, Wendland RL, Worgall S, Crystal RG, Leopold PL. Variation in adenovirus receptor expression and adenovirus vector-mediated transgene expression at defined stages of the cell cycle. Mol Ther. 2001;4:13–21. doi: 10.1006/mthe.2001.0414. [DOI] [PubMed] [Google Scholar]
- 30.Hemminki A, Kanerva A, Liu B, Wang M, Alvarez RD, Siegal GP, et al. Modulation of coxsackie-adenovirus receptor expression for increased adenoviral transgene expression. Cancer Res. 2003;63:847–853. [PubMed] [Google Scholar]
- 31.Fechner H, Haack A, Wang H, Wang X, Eizema K, Pauschinger M, et al. Expression of coxsackie adenovirus receptor and alphav-integrin does not correlate with adenovector targeting in vivo indicating anatomical vector barriers. Gene Therapy. 1999;6:1520–1535. doi: 10.1038/sj.gt.3301030. [DOI] [PubMed] [Google Scholar]
- 32.Wood M, Perrotte P, Onishi E, Harper ME, Dinney C, Pagliaro L, et al. Biodistribution of an adenoviral vector carrying the luciferase reporter gene following intravesical or intravenous administration to a mouse. Cancer Gene Ther. 1999;6:367–372. doi: 10.1038/sj.cgt.7700090. [DOI] [PubMed] [Google Scholar]
- 33.Reynolds P, Dmitriev I, Curiel D. Insertion of an RGD motif into the HI loop of adenovirus fiber protein alters the distribution of transgene expression of the systemically administered vector. Gene Therapy. 1999;6:1336–1339. doi: 10.1038/sj.gt.3300941. [DOI] [PubMed] [Google Scholar]
- 34.Tao N, Gao GP, Parr M, Johnston J, Baradet T, Wilson JM, et al. Sequestration of adenoviral vector by Kupffer cells leads to a nonlinear dose response of transduction in liver. Mol Ther. 2001;3:28–35. doi: 10.1006/mthe.2000.0227. [DOI] [PubMed] [Google Scholar]
- 35.Connelly S. Adenoviral vectors for liver-directed gene therapy. Curr Opin Mol Ther. 1999;1:565–572. [PubMed] [Google Scholar]
- 36.Lieber A, He CY, Meuse L, Schowalter D, Kirillova I, Winther B, et al. The role of Kupffer cell activation and viral gene expression in early liver toxicity after infusion of recombinant adenovirus vectors. J Virol. 1997;71:8798–8807. doi: 10.1128/jvi.71.11.8798-8807.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Peeters MJ, Patijn GA, Lieber A, Meuse L, Kay MA. Adenovirus-mediated hepatic gene transfer in mice: comparison of intravascular and biliary administration. Hum Gene Ther. 1996;7:1693–1699. doi: 10.1089/hum.1996.7.14-1693. [DOI] [PubMed] [Google Scholar]
- 38.Alemany R, Suzuki K, Curiel DT. Blood clearance rates of adenovirus type 5 in mice. J Gen Virol. 2000;81(Part 11):2605–2609. doi: 10.1099/0022-1317-81-11-2605. [DOI] [PubMed] [Google Scholar]
- 39.Worgall S, Wolff G, Falck-Pedersen E, Crystal RG. Innate immune mechanisms dominate elimination of adenoviral vectors following in vivo administration. Hum Gene Ther. 1997;8:37–44. doi: 10.1089/hum.1997.8.1-37. [DOI] [PubMed] [Google Scholar]
- 40.Alemany R, Curiel DT. CAR-binding ablation does not change biodistribution and toxicity of adenoviral vectors. Gene Therapy. 2001;8:1347–1353. doi: 10.1038/sj.gt.3301515. [DOI] [PubMed] [Google Scholar]
- 41.Smith TA, Idamakanti N, Marshall-Neff J, Rollence ML, Wright P, Kaloss M, et al. Receptor interactions involved in adenoviral-mediated gene delivery after systemic administration in non-human primates. Hum Gene Ther. 2003;14:1595–1604. doi: 10.1089/104303403322542248. [DOI] [PubMed] [Google Scholar]
- 42.Smith T, Idamakanti N, Kylefjord H, Rollence M, King L, Kaloss M, et al. In vivo hepatic adenoviral gene delivery occurs independently of the coxsackievirus-adenovirus receptor. Mol Ther. 2002;5:770–779. doi: 10.1006/mthe.2002.0613. [DOI] [PubMed] [Google Scholar]
- 43.Martin K, Brie A, Saulnier P, Perricaudet M, Yeh P, Vigne E. Simultaneous CAR- and alpha V integrin-binding ablation fails to reduce Ad5 liver tropism. Mol Ther. 2003;8:485–494. doi: 10.1016/s1525-0016(03)00182-5. [DOI] [PubMed] [Google Scholar]
- 44.Mizuguchi H, Koizumi N, Hosono T, Ishii-Watabe A, Uchida E, Utoguchi N, et al. CAR- or alphav integrin-binding ablated adenovirus vectors, but not fiber-modified vectors containing RGD peptide, do not change the systemic gene transfer properties in mice. Gene Therapy. 2002;9:769–776. doi: 10.1038/sj.gt.3301701. [DOI] [PubMed] [Google Scholar]
- 45.Nicklin SA, Wu E, Nemerow GR, Baker AH. The influence of adenovirus fiber structure and function on vector development for gene therapy. Mol Ther. 2005;12:384–393. doi: 10.1016/j.ymthe.2005.05.008. [DOI] [PubMed] [Google Scholar]
- 46.Vigne E, Dedieu JF, Brie A, Gillardeaux A, Briot D, Benihoud K, et al. Genetic manipulations of adenovirus type 5 fiber resulting in liver tropism attenuation. Gene Therapy. 2003;10:153–162. doi: 10.1038/sj.gt.3301845. [DOI] [PubMed] [Google Scholar]
- 47.Breidenbach M, Rein DT, Wang M, Nettelbeck DM, Hemminki A, Ulasov I, et al. Genetic replacement of the adenovirus shaft fiber reduces liver tropism in ovarian cancer gene therapy. Hum Gene Ther. 2004;15:509–518. doi: 10.1089/10430340460745829. [DOI] [PubMed] [Google Scholar]
- 48.Smith TA, Idamakanti N, Rollence ML, Marshall-Neff J, Kim J, Mulgrew K, et al. Adenovirus serotype 5 fiber shaft influences in vivo gene transfer in mice. Hum Gene Ther. 2003;14:777–787. doi: 10.1089/104303403765255165. [DOI] [PubMed] [Google Scholar]
- 49.Shayakhmetov DM, Li ZY, Ni S, Lieber A. Analysis of adenovirus sequestration in the liver, transduction of hepatic cells, and innate toxicity after injection of fiber-modified vectors. J Virol. 2004;78:5368–5381. doi: 10.1128/JVI.78.10.5368-5381.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Douglas JT, Rogers BE, Rosenfeld ME, Michael SI, Feng M, Curiel DT. Targeted gene delivery by tropism-modified adenoviral vectors. Nat Biotechnol. 1996;14:1574–1578. doi: 10.1038/nbt1196-1574. [DOI] [PubMed] [Google Scholar]
- 51.Goldman CK, Rogers BE, Douglas JT, Sosnowski BA, Ying W, Siegal GP, et al. Targeted gene delivery to Kaposi's sarcoma cells via the fibroblast growth factor receptor. Cancer Res. 1997;57:1447–1451. [PubMed] [Google Scholar]
- 52.Gu DL, Gonzalez AM, Printz MA, Doukas J, Ying W, D'Andrea M, et al. Fibroblast growth factor 2 retargeted adenovirus has redirected cellular tropism: evidence for reduced toxicity and enhanced antitumor activity in mice. Cancer Res. 1999;59:2608–2614. [PubMed] [Google Scholar]
- 53.Haisma HJ, Pinedo HM, Rijswijk A, der Meulen-Muileman I, Sosnowski BA, Ying W, et al. Tumor-specific gene transfer via an adenoviral vector targeted to the pan-carcinoma antigen EpCAM. Gene Therapy. 1999;6:1469–1474. doi: 10.1038/sj.gt.3300969. [DOI] [PubMed] [Google Scholar]
- 54.Heideman DA, Snijders PJ, Craanen ME, Bloemena E, Meijer CJ, Meuwissen SG, et al. Selective gene delivery toward gastric and esophageal adenocarcinoma cells via EpCAM-targeted adenoviral vectors. Cancer Gene Ther. 2001;8:342–351. doi: 10.1038/sj.cgt.7700313. [DOI] [PubMed] [Google Scholar]
- 55.Kelly FJ, Miller CR, Buchsbaum DJ, Gomez-Navarro J, Barnes MN, Alvarez RD, et al. Selectivity of TAG-72-targeted adenovirus gene transfer to primary ovarian carcinoma cells versus autologous mesothelial cells in vitro. Clin Cancer Res. 2000;6:4323–4333. [PubMed] [Google Scholar]
- 56.Reynolds PN, Zinn KR, Gavrilyuk VD, Balyasnikova IV, Rogers BE, Buchsbaum DJ, et al. A targetable, injectable adenoviral vector for selective gene delivery to pulmonary endothelium in vivo. Mol Ther. 2000;2:562–578. doi: 10.1006/mthe.2000.0205. [DOI] [PubMed] [Google Scholar]
- 57.Tillman BW, de Gruijl TD, Luykx-de Bakker SA, Scheper RJ, Pinedo HM, Curiel TJ, et al. Maturation of dendritic cells accompanies high-efficiency gene transfer by a CD40-targeted adenoviral vector. J Immunol. 1999;162:6378–6383. [PubMed] [Google Scholar]
- 58.Hakkarainen T, Hemminki A, Pereboev AV, Barker SD, Asiedu CK, Strong TV, et al. CD40 is expressed on ovarian cancer cells and can be utilized for targeting adenoviruses. Clin Cancer Res. 2003;9:619–624. [PubMed] [Google Scholar]
- 59.Pereboev AV, Asiedu CK, Kawakami Y, Dong SS, Blackwell JL, Kashentseva EA, et al. Coxsackievirus-adenovirus receptor genetically fused to anti-human CD40 scFv enhances adenoviral transduction of dendritic cells. Gene Therapy. 2002;9:1189–1193. doi: 10.1038/sj.gt.3301767. [DOI] [PubMed] [Google Scholar]
- 60.Dmitriev I, Kashentseva E, Rogers BE, Krasnykh V, Curiel DT. Ectodomain of coxsackievirus and adenovirus receptor genetically fused to epidermal growth factor mediates adenovirus targeting to epidermal growth factor receptor-positive cells. J Virol. 2000;74:6875–6884. doi: 10.1128/jvi.74.15.6875-6884.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Kashentseva EA, Seki T, Curiel DT, Dmitriev IP. Adenovirus targeting to c-erbB-2 oncoprotein by single-chain antibody fused to trimeric form of adenovirus receptor ectodomain. Cancer Res. 2002;62:609–616. [PubMed] [Google Scholar]
- 62.Itoh A, Okada T, Mizuguchi H, Hayakawa T, Mizukami H, Kume A, et al. A soluble CAR–SCF fusion protein improves adenoviral vector-mediated gene transfer to c-Kit-positive hematopoietic cells. J Gene Med. 2003;5:929–940. doi: 10.1002/jgm.430. [DOI] [PubMed] [Google Scholar]
- 63.Kim J, Smith T, Idamakanti N, Mulgrew K, Kaloss M, Kylefjord H, et al. Targeting adenoviral vectors by using the extracellular domain of the coxsackie-adenovirus receptor: improved potency via trimerization. J Virol. 2002;76:1892–1903. doi: 10.1128/JVI.76.4.1892-1903.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Krasnykh VN, Mikheeva GV, Douglas JT, Curiel DT. Generation of recombinant adenovirus vectors with modified fibers for altering viral tropism. J Virol. 1996;70:6839–6846. doi: 10.1128/jvi.70.10.6839-6846.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Havenga MJ, Lemckert AA, Ophorst OJ, van Meijer M, Germeraad WT, Grimbergen J, et al. Exploiting the natural diversity in adenovirus tropism for therapy and prevention of disease. J Virol. 2002;76:4612–4620. doi: 10.1128/JVI.76.9.4612-4620.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Gaggar A, Shayakhmetov DM, Lieber A. CD46 is a cellular receptor for group B adenoviruses. Nat Med. 2003;9:1408–1412. doi: 10.1038/nm952. [DOI] [PubMed] [Google Scholar]
- 67.Segerman A, Arnberg N, Erikson A, Lindman K, Wadell G. There are two different species B adenovirus receptors: sBAR, common to species B1 and B2 adenoviruses, and sB2AR, exclusively used by species B2 adenoviruses. J Virol. 2003;77:1157–1162. doi: 10.1128/JVI.77.2.1157-1162.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Segerman A, Atkinson JP, Marttila M, Dennerquist V, Wadell G, Arnberg N. Adenovirus type 11 uses CD46 as a cellular receptor. J Virol. 2003;77:9183–9191. doi: 10.1128/JVI.77.17.9183-9191.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Sirena D, Lilienfeld B, Eisenhut M, Kalin S, Boucke K, Beerli RR, et al. The human membrane cofactor CD46 is a receptor for species B adenovirus serotype 3. J Virol. 2004;78:4454–4462. doi: 10.1128/JVI.78.9.4454-4462.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Short JJ, Pereboev AV, Kawakami Y, Vasu C, Holterman MJ, Curiel DT. Adenovirus serotype 3 utilizes CD80 (B7.1) and CD86 (B7.2) as cellular attachment receptors. Virology. 2004;322:349–359. doi: 10.1016/j.virol.2004.02.016. [DOI] [PubMed] [Google Scholar]
- 71.Arnberg N, Edlund K, Kidd AH, Wadell G. Adenovirus type 37 uses sialic acid as a cellular receptor. J Virol. 2000;74:42–48. [PMC free article] [PubMed] [Google Scholar]
- 72.Arnberg N, Kidd AH, Edlund K, Nilsson J, Pring-Akerblom P, Wadell G. Adenovirus type 37 binds to cell surface sialic acid through a charge-dependent interaction. Virology. 2002;302:33–43. doi: 10.1006/viro.2002.1503. [DOI] [PubMed] [Google Scholar]
- 73.Wu E, Trauger SA, Pache L, Mullen TM, von Seggern DJ, Siuzdak G, et al. Membrane cofactor protein is a receptor for adenoviruses associated with epidemic keratoconjunctivitis. J Virol. 2004;78:3897–3905. doi: 10.1128/JVI.78.8.3897-3905.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Kanerva A, Zinn KR, Chaudhuri TR, Suzuki K, Uil T, Hakkarainen T, et al. Enhanced therapeutic efficacy for ovarian cancer with a serotype 3 receptor targeted oncolytic virus. Mol Ther. 2003;8:449–458. doi: 10.1016/s1525-0016(03)00200-4. [DOI] [PubMed] [Google Scholar]
- 75.Kanerva A, Wang M, Bauerschmitz GJ, Lam JT, Desmond RA, Bhoola SM, et al. Gene transfer to ovarian cancer versus normal tissues with fiber-modified adenoviruses. Mol Ther. 2002b;5:695–704. doi: 10.1006/mthe.2002.0599. [DOI] [PubMed] [Google Scholar]
- 76.Kanerva A, Mikheeva GV, Krasnykh V, Coolidge CJ, Lam JT, Mahasreshti PJ, et al. Targeting adenovirus to the serotype 3 receptor increases gene transfer efficiency to ovarian cancer cells. Clin Cancer Res. 2002;8:275–280. [PubMed] [Google Scholar]
- 77.Zabner J, Chillon M, Grunst T, Moninger TO, Davidson BL, Gregory R, et al. A chimeric type 2 adenovirus vector with a type 17 fiber enhances gene transfer to human airway epithelia. J Virol. 1999;73:8689–8695. doi: 10.1128/jvi.73.10.8689-8695.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Rea D, Havenga MJ, van Den Assem M, Sutmuller RP, Lemckert A, Hoeben RC, et al. Highly efficient transduction of human monocyte-derived dendritic cells with subgroup B fiber-modified adenovirus vectors enhances transgene-encoded antigen presentation to cytotoxic T cells. J Immunol. 2001;166:5236–5244. doi: 10.4049/jimmunol.166.8.5236. [DOI] [PubMed] [Google Scholar]
- 79.Von Seggern DJ, Huang S, Fleck SK, Stevenson SC, Nemerow GR. Adenovirus vector pseudotyping in fiber-expressing cell lines: improved transduction of Epstein–Barr virus-transformed B cells. J Virol. 2000;74:354–362. doi: 10.1128/jvi.74.1.354-362.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Shayakhmetov DM, Papayannopoulou T, Stamatoyannopoulos G, Lieber A. Efficient gene transfer into human CD34(+) cells by a retargeted adenovirus vector. J Virol. 2000;74:2567–2583. doi: 10.1128/jvi.74.6.2567-2583.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Goossens PH, Havenga MJ, Pieterman E, Lemckert AA, Breedveld FC, Bout A, et al. Infection efficiency of type 5 adenoviral vectors in synovial tissue can be enhanced with a type 16 fiber. Arthritis Rheum. 2001;44:570–577. doi: 10.1002/1529-0131(200103)44:3<570::AID-ANR105>3.0.CO;2-Y. [DOI] [PubMed] [Google Scholar]
- 82.Havenga MJ, Lemckert AA, Grimbergen JM, Vogels R, Huisman LG, Valerio D, et al. Improved adenovirus vectors for infection of cardiovascular tissues. J Virol. 2001;75:3335–3342. doi: 10.1128/JVI.75.7.3335-3342.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Gall J, Kass-Eisler A, Leinwand L, Falck-Pedersen E. Adenovirus type 5 and 7 capsid chimera: fiber replacement alters receptor tropism without affecting primary immune neutralization epitopes. J Virol. 1996;70:2116–2123. doi: 10.1128/jvi.70.4.2116-2123.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Chillon M, Bosch A, Zabner J, Law L, Armentano D, Welsh MJ, et al. Group D adenoviruses infect primary central nervous system cells more efficiently than those from group C. J Virol. 1999;73:2537–2540. doi: 10.1128/jvi.73.3.2537-2540.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Stoff-Khalili MA, Rivera AA, Glasgow JN, Le LP, Stoff A, Everts M, et al. A human adenoviral vector with a chimeric fiber from canine adenovirus type 1 results in novel expanded tropism for cancer gene therapy. Gene Therapy. 2005;12:1696–1706. doi: 10.1038/sj.gt.3302588. [DOI] [PubMed] [Google Scholar]
- 86.Glasgow JN, Kremer EJ, Hemminki A, Siegal GP, Douglas JT, Curiel DT. An adenovirus vector with a chimeric fiber derived from canine adenovirus type 2 displays novel tropism. Virology. 2004;324:103–116. doi: 10.1016/j.virol.2004.03.028. [DOI] [PubMed] [Google Scholar]
- 87.Tsuruta Y, Pereboeva L, Glasgow JN, Luongo CL, Komarova S, Kawakami Y, et al. Reovirus sigma1 fiber incorporated into adenovirus serotype 5 enhances infectivity via a CAR-independent pathway. Biochem Biophys Res Commun. 2005;335:205–214. doi: 10.1016/j.bbrc.2005.07.054. [DOI] [PubMed] [Google Scholar]
- 88.Mercier GT, Campbell JA, Chappell JD, Stehle T, Dermody TS, Barry MA. A chimeric adenovirus vector encoding reovirus attachment protein sigma1 targets cells expressing junctional adhesion molecule 1. Proc Natl Acad Sci USA. 2004;101:6188–6193. doi: 10.1073/pnas.0400542101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Wickham TJ, Tzeng E, Shears LL, Roelvink PW, II, Li Y, Lee GM, et al. Increased in vitro and in vivo gene transfer by adenovirus vectors containing chimeric fiber proteins. J Virol. 1997;71:8221–8229. doi: 10.1128/jvi.71.11.8221-8229.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Krasnykh V, Dmitriev I, Mikheeva G, Miller CR, Belousova N, Curiel DT. Characterization of an adenovirus vector containing a heterologous peptide epitope in the HI loop of the fiber knob. J Virol. 1998;72:1844–1852. doi: 10.1128/jvi.72.3.1844-1852.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Belousova N, Krendelchtchikova V, Curiel DT, Krasnykh V. Modulation of adenovirus vector tropism via incorporation of polypeptide ligands into the fiber protein. J Virol. 2002;76:8621–8631. doi: 10.1128/JVI.76.17.8621-8631.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Hemminki A, Wang M, Desmond RA, Strong TV, Alvarez RD, Curiel DT. Serum and ascites neutralizing antibodies in ovarian cancer patients treated with intraperitoneal adenoviral gene therapy. Hum Gene Ther. 2002;13:1505–1514. doi: 10.1089/10430340260185139. [DOI] [PubMed] [Google Scholar]
- 93.Hemminki A, Belousova N, Zinn KR, Liu B, Wang M, Chaudhuri TR, et al. An adenovirus with enhanced infectivity mediates molecular chemotherapy of ovarian cancer cells and allows imaging of gene expression. Mol Ther. 2001;4:223–231. doi: 10.1006/mthe.2001.0446. [DOI] [PubMed] [Google Scholar]
- 94.Bauerschmitz GJ, Barker SD, Hemminki A. Adenoviral gene therapy for cancer: from vectors to targeted and replication competent agents (review) Int J Oncol. 2002;21:1161–1174. [PubMed] [Google Scholar]
- 95.Wu H, Seki T, Dmitriev I, Uil T, Kashentseva E, Han T, et al. Double modification of adenovirus fiber with RGD and polylysine motifs improves coxsackievirus-adenovirus receptor-independent gene transfer efficiency. Hum Gene Ther. 2002;13:1647–1653. doi: 10.1089/10430340260201734. [DOI] [PubMed] [Google Scholar]
- 96.Contreras JL, Wu H, Smyth CA, Eckstein CP, Young CJ, Seki T, et al. Double genetic modification of adenovirus fiber with RGD polylysine motifs significantly enhances gene transfer to isolated human pancreatic islets. Transplantation. 2003;76:252–261. doi: 10.1097/01.TP.0000066361.02042.CA. [DOI] [PubMed] [Google Scholar]
- 97.Wu H, Han T, Lam JT, Leath CA, Dmitriev I, Kashentseva E, et al. Preclinical evaluation of a class of infectivity-enhanced adenoviral vectors in ovarian cancer gene therapy. Gene Therapy. 2004;11:874–878. doi: 10.1038/sj.gt.3302249. [DOI] [PubMed] [Google Scholar]
- 98.Rein DT, Breidenbach M, Wu H, Han T, Haviv YS, Wang M, et al. Gene transfer to cervical cancer with fiber-modified adenoviruses. Int J Cancer. 2004;111:698–704. doi: 10.1002/ijc.20295. [DOI] [PubMed] [Google Scholar]
- 99.Nicklin SA, White SJ, Watkins SJ, Hawkins RE, Baker AH. Selective targeting of gene transfer to vascular endothelial cells by use of peptides isolated by phage display. Circulation. 2000;102:231–237. doi: 10.1161/01.cir.102.2.231. [DOI] [PubMed] [Google Scholar]
- 100.Nicklin SA, Dishart KL, Buening H, Reynolds PN, Hallek M, Nemerow GR, et al. Transductional and transcriptional targeting of cancer cells using genetically engineered viral vectors. Cancer Lett. 2003;201:165–173. doi: 10.1016/j.canlet.2003.07.003. [DOI] [PubMed] [Google Scholar]
- 101.Korokhov N, Mikheeva G, Krendelshchikov A, Belousova N, Simonenko V, Krendelshchikova V, et al. Targeting of adenovirus via genetic modification of the viral capsid combined with a protein bridge. J Virol. 2003;77:12931–12940. doi: 10.1128/JVI.77.24.12931-12940.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Volpers C, Thirion C, Biermann V, Hussmann S, Kewes H, Dunant P, et al. Antibody-mediated targeting of an adenovirus vector modified to contain a synthetic immunoglobulin g-binding domain in the capsid. J Virol. 2003;77:2093–2104. doi: 10.1128/JVI.77.3.2093-2104.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Henning P, Andersson KM, Frykholm K, Ali A, Magnusson MK, Nygren PA, et al. Tumor cell targeted gene delivery by adenovirus 5 vectors carrying knobless fibers with antibody-binding domains. Gene Therapy. 2005;12:211–224. doi: 10.1038/sj.gt.3302408. [DOI] [PubMed] [Google Scholar]
- 104.Korokhov N, de Gruijl TD, Aldrich WA, Triozzi PL, Banerjee PT, Gillies SD, et al. High efficiency transduction of dendritic cells by adenoviral vectors targeted to DC-SIGN. Cancer Biol Ther. 2005;4:289–294. doi: 10.4161/cbt.4.3.1499. [DOI] [PubMed] [Google Scholar]
- 105.Breidenbach M, Rein DT, Everts M, Glasgow JN, Wang M, Passineau MJ, et al. Mesothelin-mediated targeting of adenoviral vectors for ovarian cancer gene therapy. Gene Therapy. 2005;12:187–193. doi: 10.1038/sj.gt.3302404. [DOI] [PubMed] [Google Scholar]
- 106.Balyasnikova IV, Metzger R, Visintine DJ, Dimasius V, Sun ZL, Berestetskaya YV, et al. Selective rat lung endothelial targeting with a new set of monoclonal antibodies to angiotensin I-converting enzyme. Pulm Pharmacol Ther. 2005;18:251–267. doi: 10.1016/j.pupt.2004.12.008. [DOI] [PubMed] [Google Scholar]
- 107.Von Seggern DJ, Chiu CY, Fleck SK, Stewart PL, Nemerow GR. A helper-independent adenovirus vector with E1, E3, and fiber deleted: structure and infectivity of fiberless particles. J Virol. 1999;73:1601–1608. doi: 10.1128/jvi.73.2.1601-1608.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Falgout B, Ketner G. Characterization of adenovirus particles made by deletion mutants lacking the fiber gene. J Virol. 1988;62:622–625. doi: 10.1128/jvi.62.2.622-625.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Papanikolopoulou K, Forge V, Goeltz P, Mitraki A. Formation of highly stable chimeric trimers by fusion of an adenovirus fiber shaft fragment with the foldon domain of bacteriophage T4 fibritin. J Biol Chem. 2004;279:8991–8998. doi: 10.1074/jbc.M311791200. [DOI] [PubMed] [Google Scholar]
- 110.Krasnykh V, Belousova N, Korokhov N, Mikheeva G, Curiel DT. Genetic targeting of an adenovirus vector via replacement of the fiber protein with the phage T4 fibritin. J Virol. 2001;75:4176–4183. doi: 10.1128/JVI.75.9.4176-4183.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Magnusson MK, Hong SS, Boulanger P, Lindholm L. Genetic retargeting of adenovirus: novel strategy employing ‘deknobbing’ of the fiber. J Virol. 2001;75:7280–7289. doi: 10.1128/JVI.75.16.7280-7289.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Gaden F, Franqueville L, Magnusson MK, Hong SS, Merten MD, Lindholm L, et al. Gene transduction and cell entry pathway of fiber-modified adenovirus type 5 vectors carrying novel endocytic peptide ligands selected on human tracheal glandular cells. J Virol. 2004;78:7227–7247. doi: 10.1128/JVI.78.13.7227-7247.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Belousova N, Korokhov N, Krendelshchikova V, Simonenko V, Mikheeva G, Triozzi PL, et al. Genetically targeted adenovirus vector directed to CD40-expressing cells. J Virol. 2003;77:11367–11377. doi: 10.1128/JVI.77.21.11367-11377.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Izumi M, Kawakami Y, Glasgow JN, Belousova N, Everts M, Kim-Park S, et al. In vivo analysis of a genetically modified adenoviral vector targeted to human CD40 using a novel transient transgenic model. J Gene Med. 2005;7:1517–1525. doi: 10.1002/jgm.806. [DOI] [PubMed] [Google Scholar]
- 115.Pereboeva L, Komarova S, Mahasreshti PJ, Curiel DT. Fiber-mosaic adenovirus as a novel approach to design genetically modified adenoviral vectors. Virus Res. 2004;105:35–46. doi: 10.1016/j.virusres.2004.04.012. [DOI] [PubMed] [Google Scholar]
- 116.Takayama K, Reynolds PN, Short JJ, Kawakami Y, Adachi Y, Glasgow JN, et al. A mosaic adenovirus possessing serotype Ad5 and serotype Ad3 knobs exhibits expanded tropism. Virology. 2003;309:282–293. doi: 10.1016/s0042-6822(03)00067-9. [DOI] [PubMed] [Google Scholar]
- 117.Magnusson MK, Hong SS, Henning P, Boulanger P, Lindholm L. Genetic retargeting of adenovirus vectors: functionality of targeting ligands and their influence on virus viability. J Gene Med. 2002;4:356–370. doi: 10.1002/jgm.285. [DOI] [PubMed] [Google Scholar]
- 118.Hedley SJ, Auf der Maur A, Hohn S, Escher D, Barberis A, Glasgow JN, et al. An adenovirus vector with a chimeric fiber incorporating stabilized single chain antibody achieves targeted gene delivery. Gene Therapy. 2006;13:88–94. doi: 10.1038/sj.gt.3302603. [DOI] [PubMed] [Google Scholar]
- 119.Athappilly FK, Murali R, Rux JJ, Cai Z, Burnett RM. The refined crystal structure of hexon, the major coat protein of adenovirus type 2, at 2.9 A resolution. J Mol Biol. 1994;242:430–455. doi: 10.1006/jmbi.1994.1593. [DOI] [PubMed] [Google Scholar]
- 120.Crawford-Miksza L, Schnurr DP. Analysis of 15 adenovirus hexon proteins reveals the location and structure of seven hypervariable regions containing serotype-specific residues. J Virol. 1996;70:1836–1844. doi: 10.1128/jvi.70.3.1836-1844.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Vigne E, Mahfouz I, Dedieu JF, Brie A, Perricaudet M, Yeh P. RGD inclusion in the hexon monomer provides adenovirus type 5-based vectors with a fiber knob-independent pathway for infection. J Virol. 1999;73:5156–5161. doi: 10.1128/jvi.73.6.5156-5161.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Parks RJ. Adenovirus protein IX: a new look at an old protein. Mol Ther. 2005;11:19–25. doi: 10.1016/j.ymthe.2004.09.018. [DOI] [PubMed] [Google Scholar]
- 123.Vellinga J, van den Wollenberg DJ, van der Heijdt S, Rabelink MJ, Hoeben RC. The coiled-coil domain of the adenovirus type 5 protein IX is dispensable for capsid incorporation and thermostability. J Virol. 2005;79:3206–3210. doi: 10.1128/JVI.79.5.3206-3210.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Ghosh-Choudhury G, Haj-Ahmad Y, Graham FL. Protein IX, a minor component of the human adenovirus capsid, is essential for the packaging of full length genomes. EMBO J. 1987;6:1733–1739. doi: 10.1002/j.1460-2075.1987.tb02425.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Colby WW, Shenk T. Adenovirus type 5 virions can be assembled in vivo in the absence of detectable polypeptide IX. J Virol. 1981;39:977–980. doi: 10.1128/jvi.39.3.977-980.1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Boulanger P, Lemay P, Blair GE, Russell WC. Characterization of adenovirus protein IX. J Gen Virol. 1979;44:783–800. doi: 10.1099/0022-1317-44-3-783. [DOI] [PubMed] [Google Scholar]
- 127.Akalu A, Liebermann H, Bauer U, Granzow H, Seidel W. The subgenus-specific C-terminal region of protein IX is located on the surface of the adenovirus capsid. J Virol. 1999;73:6182–6187. doi: 10.1128/jvi.73.7.6182-6187.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Rosa-Calatrava M, Grave L, Puvion-Dutilleul F, Chatton B, Kedinger C. Functional analysis of adenovirus protein IX identifies domains involved in capsid stability, transcriptional activity, and nuclear reorganization. J Virol. 2001;75:7131–7141. doi: 10.1128/JVI.75.15.7131-7141.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Dmitriev IP, Kashentseva EA, Curiel DT. Engineering of adenovirus vectors containing heterologous peptide sequences in the C terminus of capsid protein IX. J Virol. 2002;76:6893–6899. doi: 10.1128/JVI.76.14.6893-6899.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Vellinga J, Rabelink MJ, Cramer SJ, van den Wollenberg DJ, Van der Meulen H, Leppard KN, et al. Spacers increase the accessibility of peptide ligands linked to the carboxyl terminus of adenovirus minor capsid protein IX. J Virol. 2004;78:3470–3479. doi: 10.1128/JVI.78.7.3470-3479.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Campos SK, Parrott MB, Barry MA. Avidin-based targeting and purification of a protein IX-modified, metabolically biotinylated adenoviral vector. Mol Ther. 2004;9:942–954. doi: 10.1016/j.ymthe.2004.03.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Meulenbroek RA, Sargent KL, Lunde J, Jasmin BJ, Parks RJ. Use of adenovirus protein IX (pIX) to display large polypeptides on the virion-generation of fluorescent virus through the incorporation of pIX-GFP. Mol Ther. 2004;9:617–624. doi: 10.1016/j.ymthe.2004.01.012. [DOI] [PubMed] [Google Scholar]
- 133.Le LP, Everts M, Dmitriev IP, Davydova JG, Yamamoto M, Curiel DT. Fluorescently labeled adenovirus with pIX-EGFP for vector detection. Mol Imaging. 2004;3:105–116. doi: 10.1162/15353500200404100. [DOI] [PubMed] [Google Scholar]
- 134.Le LP, Dmitriev I, Davydova J, Yamamoto M, Curiel D. Genetic labeling of adenovirus with red fluorescent proteins on PIX for vector detection. Mol Ther. 2004;9(S1):S296. doi: 10.1162/15353500200404100. [DOI] [PubMed] [Google Scholar]
- 135.Le LP, Dmitriev I, Davydova J, Yamamoto M, Curiel D. Genetic labeling of adenovirus with luciferase on pIX for vector detection. Mol Ther. 2004;9(S1):297–298. doi: 10.1162/15353500200404100. [DOI] [PubMed] [Google Scholar]
- 136.Li J, Le LP, Sibley DA, Mathis JM, Curiel DT. Genetic incorporation of HSV-1 thymidine kinase into the adenovirus protein ix for functioal display on the virion. Virology. 2005;338:247–258. doi: 10.1016/j.virol.2005.04.005. [DOI] [PubMed] [Google Scholar]
- 137.Vellinga J, Van der Heijdt S, Hoeben RC. The adenovirus capsid: major progress in minor proteins. J Gen Virol. 2005;86(Part 6):1581–1588. doi: 10.1099/vir.0.80877-0. [DOI] [PubMed] [Google Scholar]
- 138.Dmitriev I, Kashentseva EA, Seki T, Curiel DT. Utilization of minor capsid polypeptides IX and IIIa for adenovirus targeting. Mol Ther. 2001;3:S167. [Google Scholar]
- 139.Curiel D. United States patent 6555368 Capsid-modified recombinant adenovirus vectors and methods of use. 2003
- 140.Welsh JB, Sapinoso LM, Su AI, Kern SG, Wang-Rodriguez J, Moskaluk CA, et al. Analysis of gene expression identifies candidate markers and pharmacological targets in prostate cancer. Cancer Res. 2001;61:5974–5978. [PubMed] [Google Scholar]
- 141.Welsh JB, Zarrinkar PP, Sapinoso LM, Kern SG, Behling CA, Monk BJ, et al. Analysis of gene expression profiles in normal and neoplastic ovarian tissue samples identifies candidate molecular markers of epithelial ovarian cancer. Proc Natl Acad Sci USA. 2001;98:1176–1181. doi: 10.1073/pnas.98.3.1176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Omenn GS. The human proteome organization plasma proteome project pilot phase: reference specimens, technology platform comparisons, and standardized data submissions and analyses. Proteomics. 2004;4:1235–1240. doi: 10.1002/pmic.200300686. [DOI] [PubMed] [Google Scholar]
- 143.Nishizuka S, Chen ST, Gwadry FG, Alexander J, Major SM, Scherf U, et al. Diagnostic markers that distinguish colon and ovarian adenocarcinomas: identification by genomic, proteomic, and tissue array profiling. Cancer Res. 2003;63:5243–5250. [PubMed] [Google Scholar]
- 144.Sahin U, Tureci O, Schmitt H, Cochlovius B, Johannes T, Schmits R, et al. Human neoplasms elicit multiple specific immune responses in the autologous host. Proc Natl Acad Sci USA. 1995;92:11810–11813. doi: 10.1073/pnas.92.25.11810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Li G, Miles A, Line A, Rees RC. Identification of tumour antigens by serological analysis of cDNA expression cloning. Cancer Immunol Immunother. 2004;53:139–143. doi: 10.1007/s00262-003-0471-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Smith GP. Filamentous fusion phage: novel expression vectors that display cloned antigens on the virion surface. Science. 1985;228:1315–1317. doi: 10.1126/science.4001944. [DOI] [PubMed] [Google Scholar]
- 147.Peletskaya EN, Glinsky VV, Glinsky GV, Deutscher SL, Quinn TP. Characterization of peptides that bind the tumor-associated Thomsen–Friedenreich antigen selected from bacteriophage display libraries. J Mol Biol. 1997;270:374–384. doi: 10.1006/jmbi.1997.1107. [DOI] [PubMed] [Google Scholar]
- 148.Peletskaya EN, Glinsky G, Deutscher SL, Quinn TP. Identification of peptide sequences that bind the Thomsen–Friedenreich cancer-associated glycoantigen from bacteriophage peptide display libraries. Mol Divers. 1996;2:13–18. doi: 10.1007/BF01718695. [DOI] [PubMed] [Google Scholar]
- 149.Koivunen E, Gay DA, Ruoslahti E. Selection of peptides binding to the alpha 5 beta 1 integrin from phage display library. J Biol Chem. 1993;268:20205–20210. [PubMed] [Google Scholar]
- 150.Schiffelers RM, Koning GA, ten Hagen TL, Fens MH, Schraa AJ, Janssen AP, et al. Anti-tumor efficacy of tumor vasculature-targeted liposomal doxorubicin. J Control Release. 2003;91:115–122. doi: 10.1016/s0168-3659(03)00240-2. [DOI] [PubMed] [Google Scholar]
- 151.Shi W, Bartlett JS. RGD inclusion in VP3 provides adeno-associated virus type 2 (AAV2)-based vectors with a heparan sulfate-independent cell entry mechanism. Mol Ther. 2003;7:515–525. doi: 10.1016/s1525-0016(03)00042-x. [DOI] [PubMed] [Google Scholar]
- 152.Pereboev A, Pereboeva L, Curiel DT. Phage display of adenovirus type 5 fiber knob as a tool for specific ligand selection and validation. J Virol. 2001;75:7107–7113. doi: 10.1128/JVI.75.15.7107-7113.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Zhu ZB, Makhija SK, Lu B, Wang M, Rivera AA, Preuss M, et al. Transport across a polarized monolayer of Caco-2 cells by transferrin receptor-mediated adenovirus transcytosis. Virology. 2004;325:116–128. doi: 10.1016/j.virol.2004.04.021. [DOI] [PubMed] [Google Scholar]
- 154.Moroo I, Ujiie M, Walker BL, Tiong JW, Vitalis TZ, Karkan D, et al. Identification of a novel route of iron transcytosis across the mammalian blood–brain barrier. Microcirculation. 2003;10:457–462. doi: 10.1038/sj.mn.7800213. [DOI] [PubMed] [Google Scholar]
- 155.Mostov KE. Transepithelial transport of immunoglobulins. Annu Rev Immunol. 1994;12:63–84. doi: 10.1146/annurev.iy.12.040194.000431. [DOI] [PubMed] [Google Scholar]
- 156.McIntosh DP, Tan XY, Oh P, Schnitzer JE. Targeting endothelium and its dynamic caveolae for tissue-specific transcytosis in vivo: a pathway to overcome cell barriers to drug and gene delivery. Proc Natl Acad Sci USA. 2002;99:1996–2001. doi: 10.1073/pnas.251662398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.van Dillen IJ, Mulder NH, Vaalburg W, de Vries EF, Hospers GA. Influence of the bystander effect on HSV-tk/GCV gene therapy. A review. Curr Gene Ther. 2002;2:307–322. doi: 10.2174/1566523023347733. [DOI] [PubMed] [Google Scholar]
- 158.Jordan A, Scholz R, Wust P, Fahling H, Krause J, Wlodarczyk W, et al. Effects of magnetic fluid hyperthermia (MFH) on C3H mammary carcinoma in vivo. Int J Hyperthermia. 1997;13:587–605. doi: 10.3109/02656739709023559. [DOI] [PubMed] [Google Scholar]
- 159.Brigger I, Dubernet C, Couvreur P. Nanoparticles in cancer therapy and diagnosis. Adv Drug Deliv Rev. 2002;54:631–651. doi: 10.1016/s0169-409x(02)00044-3. [DOI] [PubMed] [Google Scholar]
- 160.Ito A, Kuga Y, Honda H, Kikkawa H, Horiuchi A, Watanabe Y, et al. Magnetite nanoparticle-loaded anti-HER2 immunoliposomes for combination of antibody therapy with hyperthermia. Cancer Lett. 2004;212:167–175. doi: 10.1016/j.canlet.2004.03.038. [DOI] [PubMed] [Google Scholar]
- 161.Hergt R, Andra W, d'Ambly CG, Hilger I, Kaiser WA, Richter U, et al. Physical limits of hyperthermia using magnetite fine particles. IEEE Trans Mag. 1998;34:3745–3754. [Google Scholar]
- 162.Hilger I, Hergt R, Kaiser WA. Towards breast cancer treatment by magnetic heating. J Magn Magn Mater. 2005;293:314–319. [Google Scholar]
- 163.Neuberger T, Schoöpf B, Hofmann HMH, von Rechenberg B. Superparamagnetic nanoparticles for biomedical applications: possibilities and limitations of a new drug delivery system. J Magn Magn E. 2005;293:483–496. [Google Scholar]
- 164.Xu H, Song T, Bao X, Hu L. Site research of magnetic nanoparticles in magnetic drug targeting. J Magn Magn E. 2005;293:514–519. [Google Scholar]
- 165.Alexiou C, Jurgons R, Schmid R, Hilpert A, Bergemann C, Parak F, et al. In vitro and in vivo investigations of targeted chemotherapy with magnetic nanoparticles. J Magn Magn Mater. 2005;293:389–393. [Google Scholar]
- 166.Pankhurst QA, Connolly J, Jones SK, Dobson J. Applications of magnetic nanoparticles in biomedicine. J Phys D Appl Phys. 2003;36:R167–R181. [Google Scholar]
- 167.Choi H, Choi SR, Zhou R, Kung HF, Chen IW. Iron oxide nanoparticles as magnetic resonance contrast agent for tumor imaging via folate receptor-targeted delivery. Acad Radiol. 2004;11:996–1004. doi: 10.1016/j.acra.2004.04.018. [DOI] [PubMed] [Google Scholar]
- 168.Yang Y, Nunes FA, Berencsi K, Furth EE, Gonczol E, Wilson JM. Cellular immunity to viral antigens limits E1-deleted adenoviruses for gene therapy. Proc Natl Acad Sci USA. 1994;91:4407–4411. doi: 10.1073/pnas.91.10.4407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Yang Y, Li Q, Ertl HC, Wilson JM. Cellular and humoral immune responses to viral antigens create barriers to lung-directed gene therapy with recombinant adenoviruses. J Virol. 1995;69:2004–2015. doi: 10.1128/jvi.69.4.2004-2015.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Morral N, Parks RJ, Zhou H, Langston C, Schiedner G, Quinones J, et al. High doses of a helper-dependent adenoviral vector yield supraphysiological levels of alpha1-antitrypsin with negligible toxicity. Hum Gene Ther. 1998;9:2709–2716. doi: 10.1089/hum.1998.9.18-2709. [DOI] [PubMed] [Google Scholar]
- 171.Morsy MA, Gu M, Motzel S, Zhao J, Lin J, Su Q, et al. An adenoviral vector deleted for all viral coding sequences results in enhanced safety and extended expression of a leptin transgene. Proc Natl Acad Sci USA. 1998;95:7866–7871. doi: 10.1073/pnas.95.14.7866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Thomas CE, Schiedner G, Kochanek S, Castro MG, Lowenstein PR. Peripheral infection with adenovirus causes unexpected long-term brain inflammation in animals injected intracranially with first-generation, but not with high-capacity, adenovirus vectors: toward realistic long-term neurological gene therapy for chronic diseases. Proc Natl Acad Sci USA. 2000;97:7482–7487. doi: 10.1073/pnas.120474397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Schiedner G, Morral N, Parks RJ, Wu Y, Koopmans SC, Langston C, et al. Genomic DNA transfer with a high-capacity adenovirus vector results in improved in vivo gene expression and decreased toxicity. Nat Genet. 1998;18:180–183. doi: 10.1038/ng0298-180. [DOI] [PubMed] [Google Scholar]
- 174.Parks RJ, Graham FL. A helper-dependent system for adenovirus vector production helps define a lower limit for efficient DNA packaging. J Virol. 1997;71:3293–3298. doi: 10.1128/jvi.71.4.3293-3298.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Hardy S, Kitamura M, Harris-Stansil T, Dai Y, Phipps ML. Construction of adenovirus vectors through Cre-lox recombination. J Virol. 1997;71:1842–1849. doi: 10.1128/jvi.71.3.1842-1849.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Umana P, Gerdes CA, Stone D, Davis JR, Ward D, Castro MG, et al. Efficient FLPe recombinase enables scalable production of helper-dependent adenoviral vectors with negligible helper-virus contamination. Nat Biotechnol. 2001;19:582–585. doi: 10.1038/89349. [DOI] [PubMed] [Google Scholar]
- 177.Biermann V, Volpers C, Hussmann S, Stock A, Kewes H, Schiedner G, et al. Targeting of high-capacity adenoviral vectors. Hum Gene Ther. 2001;12:1757–1769. doi: 10.1089/104303401750476258. [DOI] [PubMed] [Google Scholar]
- 178.Stilwell JL, McCarty DM, Negishi A, Superfine R, Samulski RJ. Development and characterization of novel empty adenovirus capsids and their impact on cellular gene expression. J Virol. 2003;77:12881–12885. doi: 10.1128/JVI.77.23.12881-12885.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Stilwell JL, Samulski RJ. Role of viral vectors and virion shells in cellular gene expression. Mol Ther. 2004;9:337–346. doi: 10.1016/j.ymthe.2003.11.007. [DOI] [PubMed] [Google Scholar]