

Short abstract
Mesenchymal precursor cells found in the blood (BMPCs) of normal persons adhere to plastic and glass and proliferate logarithmically in DMEM-20% fetal calf serum (FCS) without growth factors. They form cells with fibroblast-like and stromal morphology, which is not affected by eliminating CD34, CD3, or CD14 cells. Osteogenic supplements (dexamethasone, ascorbic acid, and β-glycerophosphate) added to the culture inhibited fibroblast formation, and BMPCs assumed the cuboidal shape of osteoblasts. After 5 days in supplemented medium, the elutriated cells displayed alkaline phosphatase (AP), and the addition of bone morphogenetic protein (BMP)2 (1 ng) doubled AP production (P < 0.04). Two weeks later, 30% of the cells were very large and reacted with anti-osteocalcin antibody. The same cultures also contained sudanophlic adipocytes and multinucleated giant cells that stained for tartrate-resistant acid phosphatase (TRAP) and vitronectin receptors. Cultured BMPCs immunostain with antibodies to vimentin, type I collagen, and BMP receptors, heterodimeric structures expressed on mesenchymal lineage cells. In addition, BMPCs stain with anti-CD105 (endoglin), a putative marker for bone-marrow mesenchymal stem cells (MSCs).
Keywords: adipocytes, bone marrow progenitor cells, bone morphogenetic protein receptors, CD34, mesenchymal precursor cells, osteoblasts, osteoclasts, rheumatoid arthritis, SDF-1
Abstract
Introduction:
Adult human bone marrow contains a minority population of MSCs that contribute to the regeneration of tissues such as bone, cartilage, muscle, ligaments, tendons, fat, and stroma. Evidence that these MSCs are pluripotent, rather than being a mixture of committed progenitor cells each with a restricted potential, includes their rapid proliferation in culture, a characteristic morphology, the presence of typical marker proteins, and their consistent differentiation into various mesenchymal lineages. These attributes are maintained through multiple passages and are identifiable in individual stem cells.
Aims:
Since stem cells are present in both the bone marrow and other tissues, we thought it possible that cells with a similar appearance and pluripotent mesenchymal potential would be present in the blood. We applied techniques used successfully with marrow MSCs to identify similar cells in elutriation fractions of normal human blood.
Methods:
BMPCs were elutriated by diluting the buffy coats from 500 ml of anticoagulant-treated, platelet-depleted blood 1:4 in RPMI-1640 medium (RPMI) and layering 25-ml portions over 20 ml of Lymphoprep™. These samples were centrifuged at 2000 rpm for 20 min. The leukocyte-rich interface cells were collected, made up to 20 ml in RPMI, and separated by density-gradient centrifugation. The interface cells, now depleted of red blood cells, were collected, resuspended in 50 ml of sterile RMPI and 5% heat-inactivated FCS, and introduced into the sample line of the flow system of a Beckman JE-50 cell elutriator charged with elutriation buffer. The chamber was centrifuged at 25 000 rpm at 10°C and the flow rate adjusted to 12 ml/min. After about 150 ml had been collected, the flow rate was increased by 1 ml/min. Fractions nos. 1-6 (flow rates of 12-16 ml/min) contained most of the lymphocytes. Monocytes usually appeared in fractions 6 or 7 (as determined by flow cytometric analysis in a fluorescence-activated cell sorter (FACS). BMPCs were concentrated in fractions 7 and 8, along with monocytes and lymphocytes. Elutriation fractions with more than 50% and less than 75% monocytes were collected and concentrated by centrifugation at 1200 rpm for 5 min, and the cell pellets were combined, reconstituted in DMEM plus 20% sterile heat-inactivated FCS, counted, washed in medium, repelleted, and then resuspended in DMEM to 5 × 106/ml and dispensed into either tissue-culture plastic slides or glass chamber slides. Cells thus obtained were observed in time-lapse cinematography, assayed for proliferation, and examined immunohistologically and histochemically, and their ability to become fibroblasts, osteoclasts, osteoblasts, and adipocytes was documented.
Results:
BMPCs were found in elutriation fractions containing less than 30% T cells and more than 60% monocytes from the blood of more than 100 normal persons. BMPCs adhered to plastic and glass and proliferated logarithmically in DMEM-20% FCS without added growth factors. The initial elutriate had only small, round, mononuclear cells; upon culture, these were replaced by fibroblast-like cells and large, round, stromal cells. The formation of cells with fibroblast-like and stromal morphology was not affected by eliminating CD34, CD3, or CD14 cells from the elutriation fraction. Osteogenic supplements (dexamethasone, ascorbic acid, and β-glycero-phosphate) added to the culture inhibited fibroblast formation, and BMPCs assumed the cuboidal shape of osteoblasts. After 5 days in supplemented medium, the elutriated cells displayed AP and its production was doubled by the addition of BMP2 (1 ng) (P < 0.04). Two weeks later, 30% of the cells were very large and reacted with anti-osteocalcin antibody. The same cultures contained two other types of cell: sudanophlic adipocytes and multinucleated giant cells, which stain for TRAP and vitronectin receptors (attributes of osteoclasts). Cultured BMPCs were immunostained by antibodies to vimentin, type I collagen, and BMP receptors (heterodimeric structures expressed on mesenchymal lineage cells). The cultured cells also stained strongly for the SH-2 (endoglin) antigen, a putative marker for marrow MSCs. BMPCs express the gene for SDF-1, a potent stroma-derived CXCα chemokine.
Discussion:
In the circulation of normal individuals is a small population of CD34- mononuclear cells that proliferate rapidly in culture as an adherent population with a variable morphology, display cytoskeletal, cytoplasmic, and surface markers of mesenchymal precursors, and differentiate into several lineages (fibroblasts, osteoblasts, and adipocytes). These are all features found in bone-marrow-derived MSCs. Therefore, autologous blood could provide cells useful for tissue engineering and gene therapy. In addition, the demonstration of similar cells in the inflammatory joint fluids and synovium of patients with rheumatoid arthritis (RA) suggests that these cells may play a role in the pathogenesis of RA.
Introduction
Bone marrow is a complex tissue containing hematopoietic cell progenitors and their progeny and a connective-tissue network of mesenchymally derived cells known as stroma. Marrow stroma includes a subpopulation of undifferentiated cells that are capable of becoming one of a number of phenotypes, including chondrocytes, osteoblasts, adipocytes, fibroblasts, possibly muscle cells, and the reticular cells that support hematopoietic cell differentiation [1,2]. Extensive experimentation has defined conditions for the isolation, propagation, and differentiation in vitro and in vivo of the stromal cells referred to as MSCs. They are a population of firmly adherent cells with a high proliferative capacity and potential for self-renewal. Their developmental potential is retained even after repeated subcultivation in vitro, supporting their designation as stem cells [3].
Identification of MSCs in situ has been difficult, partly because they have few unique products or molecular markers. A series of monoclonal antibodies (SH antibodies) purportedly specific reagents have been used to isolate MSCs from a population of bone-marrow cells [1,3]. The one used most often (SH-2) was recently shown to react with endoglin (CD105), a member of the transforming growth factor (TGF)-β receptor family usually found on the endothelium of postcapillary venules [4]. Two other reagents may be more specific. One consists of a group of antibodies to BMP receptors (BMPRs) present on embryonic mesenchyme and postnatally on osteoblasts and chondrocytes [5]. Another antibody, Stro-1 made against marrow fibroblastic cells, blocks hematopoiesis in vitro by interfering with the interaction of reconstituted human hematopoietic stem cells (HSCs) and stromal cells [6].
Attempts to demonstrate MSCs in peripheral blood have been unrewarding, except for a report by Fernandez et al [7], who identified cells with the features of MSCs in growth-factor-mobilized peripheral-blood cells from breast-cancer patients. Low-density mononuclear cells grown for 1 week in tissue culture with fetal calf serum (FCS) become adherent fibroblast-like cells and a few were large, flat, round cells. Immunohistology and flow cytometric analysis in a fluorescence-activated cell sorter (FACS) revealed fibronectin and three types of collagen (I, III, and VI) in the cytoplasm of the cultured cells. They expressed adhesion ligands and antigens recognized by SH-2 and SH-3 monoclonal antibodies. No stromal cells were demonstrated in normal peripheral-blood cells not mobilized by granulocyte-macrophage CSF [7]. Bucala et al [8] separated human blood cells by density centrifugation, cultured them on a fibronectin matrix, and identified a population of circulating cells that had fibroblast properties and a distinctive phenotype (collagen+/vimentin+/ CD34+). This novel circulating cell, termed a fibrocyte, has both mesenchymal and hematopoietic features.
Now we report for the first time that cells with the morphology and phenotype of mesenchymal precursors are normally present in the circulation. Hereafter these are referred to as `blood-derived mesenchymal precursor cells' (BMPCs). The observations that support these conclusions and the significance of the findings are discussed.
Methods
Reagents and laboratory ware
Dexamethasone, ascorbic acid-2 phosphate, β-glycerophosphate, bovine serum albumin (BSA), and FCS were purchased from Sigma Diagnostics (St Louis, MO, USA); penicillin, streptomycin, DMEM, and RPMI-1640 from Biowhittaker (Watersville, MD, USA); Lymphoprep™ from Nycomed, Oslo, Norway.
Monoclonal antibodies were purchased from commercial vendors unless otherwise stated: CD3, IgG1; CD68, IgG1; CD34, IgG1; CD45, IgG1, D105, IgG1; and IgG2a controls (Dako Corporation, Carpinteria, CA, USA); anti-HLA-DR, IgG2a; CD14, IgG2b; CD34, IgG1 (Becton Dickinson, San Jose, CA, USA); anti-vimentin, IgG1; IgG2b control (Serotec, Kidlington, Oxfordshire, UK); anti-VCAM-1, IgG1; anti-αvβ3 (vitronectin receptor), IgG1 (Pharmingen, San Diego, CA, USA); anti-collagen-type-1, IgG1 (Sigma Diagnostics); anti-osteocalcin, IgG1 (ABOC-5021, Haematologic Technologies, Essex Junction, VT, USA) anti-IgG1. Stro-1 is a culture supernatant, monoclonal IgM, from Developmental Studies Hybridoma Bank, University of Iowa (Iowa City, IA, USA). Biotinylated mouse Ig, streptavidin/horseradish peroxidase conjugate, diamino benzidine, and Vectastain ABC were from Vector (Burlingame, CA, USA).
Tissue-culture treated glass slides, Petri dishes, and six-well tissue-culture plates and eight-chamber tissue culture slides (Falcon) were from Becton Dickinson Labware (Franklin Lakes, NJ, USA); 12-well sterile glass slides were from ICN (Costa Mesa, CA, USA).
Elutriation procedure for BMPCs
Anticoagulant-treated, platelet-depleted buffy coat was obtained in sterile packages from the North London Blood Transfusion Service. About 50 ml of the buffy coat was diluted 1:4 in RPMI and 25 ml was layered over 20 ml of Lymphoprep™ in a 50-ml conical centrifuge tube. The tubes (approximately eight) were centrifuged at 2000 rpm for 20 min. The supernatant was discarded and leukocyte-rich interface cells were collected and combined. These were made up to 20 ml in RPMI and layered again over Lymphoprep™ and centrifuged at 2000 rpm for 20 min more. The buffy coat, now depleted of red blood cells, was collected from the interface, resuspended in 50 ml of sterile RMPI and 5% heat-inactivated FCS, and introduced into the sample line of the flow system of a Beckman JE-50 cell elutriator which had been charged with elutriation buffer. The chamber was centrifuged at 25 000 rpm at 10°C and the flow rate adjusted to 12 ml/min. The eluate fractions were collected in sterile, conical, 50-ml tubes. After about 150 ml had been collected, the flow rate was increased by 1 ml/min. Fractions nos. 1-6 (flow rates of 12-16 ml/min) contained most of the lymphocytes. Monocytes usually made their appearance (as determined by FACS analysis) in fraction 6 or 7. In fractions 7 and 8, monocytes constituted up to two thirds of the cells, and BMPCs were concentrated in these fractions. Elutriation fractions containing more than 50% and less than 75% monocytes were concentrated by centrifugation at 1200 rpm for 5 min, and the cell pellets were combined, reconstituted in DMEM plus 20% sterile heat-inactivated FCS (hereafter referred to as complete medium, unless otherwise stated), counted in a hemocytometer, washed in DMEM medium, repelleted, resuspended to 5 × 106/ml, and dispensed into either plastic tissue-culture slides or glass chamber slides.
More than 100 consecutive buffy coats from normal individuals were processed by this method and cultured. In every case, the appropriate elutriation fractions had cells with the BMPC morphology.
Cell-proliferation assay
The BMPC-rich elutriation fractions were plated at 5 × 105 in 500 μl of complete medium in polystyrene chambers on treated glass tissue-culture slides (Falcon). At various times, cultures were rinsed twice with Tyrode's balanced salt solution, fixed with 1% glutaraldehyde (v/v) in Tyrode's for 15 min, rinsed twice with deionized water, and air-dried. Cultures were then stained with 0.1% crystal violet (w/v) in deionized water for 30 min and washed 3 times with deionized water; the crystal-violet dye was extracted by rocking the cultures gently in 1% Triton X-100 for 4 h at room temperature and read at 595 nm on a microplate reader (BioRad, Hercules, CA, USA). Absorbance values (optical densities; ODs) were converted into absolute cell numbers on the basis of established standard curves [9].
Immunohistochemistry
BMPC-rich elutriation fractions (500 μl, containing 5 × 106 cells per ml) or other sources of BMPCs were placed into the wells of sterile, 12-well multitest slides (ICN) in complete medium and left to adhere at 37°C for 4 h. The slides were then placed into 100 × 20 mm Petri dishes containing 5-7 ml DMEM-20% FCS. The nonadherent cells floated off, while mesenchymal cells adhered, spread, and grew. Their daily progress was assessed by phase-contrast microscopy. The medium was changed every 3 to 5 days and the cells were studied after 5-7 days. The growing, adherent cells were rinsed in phosphate-buffered saline (PBS), fixed in ice-cold 4% paraformaldehyde for 20 min, and then washed in PBS. All further incubations and washes were carried out using PBS. Endogenous peroxidase activity was blocked with 0.1 mol sodium azide containing 1% hydrogen peroxide, and the specimens were incubated with 10% normal goat serum, 2% normal rat serum, and 1% bovine serum albumin for 30 min at room temperature to eliminate non-specific binding. Specimens were then incubated with primary antibodies at 4°C overnight and were then incubated with a biotinylated secondary antibody (Vector). The antibody-biotin conjugates were detected with an avidin-biotin-peroxidase complex (Vector), applied for 30 min at room temperature. A color reaction was developed with 3-amino-9-ethylcarbazole and specimens were lightly counterstained with Mayer's hematoxylin.
Controls included normal rabbit or mouse IgG, 1% BSA in PBS, or, in the case of BMPR antibodies, preabsorbed with the respective peptide used for immunization.
Quantification of BMPCs by immunohistochemistry
BMPC-rich elutriation fractions (5 × 105 cells in 500 μl of complete medium) were placed into the chambers of sterile, eight-chamber, treated glass tissue-culture slides. Two to 4 h later, nonadherent cells were removed. Cultures were fed every 3 days. At regular intervals, the slides were rinsed in PBS, fixed in ice-cold 4% paraformaldehyde for 20 min, washed in PBS, stained with anti-BMPR antibodies, and visualized by the ABC immunoperoxidase method described above. The specimens were examined using an Olympus BH-2 microscope and analyzed by computer image analysis (AnalySIS, Soft Imaging System GmbH, Münster, Germany). Six digital images (400×) per specimen were recorded and quantitative analysis was performed according to the color cell separation. Images chosen at random were analyzed and the data are presented as the mean of the total number of cells per six images examined at 400×. Slender cells with a small, centrally localized nucleus were scored as fibroblast-like. Large, round cells and intermediate-sized cells with more cytoplasm and a large, round nucleus were scored as large cells.
Anti-BMPR antibodies
Rabbit polyclonal antibodies to BMPRs were provided by K Funa (Göteborg University, Gothenburg, Sweden). Polyclonal rabbit antisera were prepared using synthetic peptides corresponding to the intracellular transmembrane portions of the types IA, IB, and II BMPRs [10]. The antisera were affinity-purified and tested for specificity by immunoprecipitation of cross-linked complexes of cultured cells transfected with receptor complementary deoxyribonucleic acids (cDNAs) [11].
Magnetic antibody-coated-bead separation (MACS) was performed in accordance with the manufacturer's recommendations (Miltenyi Biotec, Inc., Auburn, CA, USA). Elutriation fractions with 50-75% monocytes were centrifuged at 900 × g , washed with MACS buffer (PBS pH7.2, + 0.5% SSA + 2 mmol EDTA) and counted in a hemocytometer. The cell pellet was resuspended in 80 μl MACS buffer per 107 total cells, and 20 μl of MACS antibody-coated beads was added to the cells, mixed, and incubated for 15 min at 6-12°C. The cells were washed with a 20-fold volume of MACS buffer, spun, and resus-pended in 500 μl buffer. The cell suspension was applied to a positive selection column washed previously with 1 ml MACS buffer and placed in a magnetic separator and the cells were eluted. The column was rinsed four times with 500 μl buffer and the cells that passed through were combined as the antigen-free fraction. The column was removed from the magnetic separator, 1 ml of buffer was added to the column, and the positive cells were flushed out with a syringe plunger. This was repeated with another 1 ml of buffer. The elutriated cells were combined as the antigen-containing fraction.
Alkaline phosphatase activity of circulating BMPCs
BMPC-rich elutriation fractions were prepared from four individual blood packs as described and plated into four-well chamber slides (Lab-Tek) at 5 × 106 cells per ml in DMEM-10% FCS. After 24 h at 37°C, the nonadherent cells were removed and new medium was added containing BMP2 (a gift from the Genetics Institute, Cambridge, MA, USA) at concentrations of 0, 1, 10, or 100 ng/ml. The cells were incubated at 37°C in 5% CO2 and the medium was changed every 5 days. Supernatants were taken at 5, 10, and 15 days and stored at -20°C for later analysis. AP activity in the supernatants was estimated using a p-nitro-phenol colorimetric assay. Cell supernatants were assayed for AP activity in 50 mmol glycine, 0.05% Triton X-100, 4 mmol MgCl2, and 5 mmol p-nitrophenol phosphate, pH10.3, for 15 min at 37°C (Sigma Diagnostics, St Louis, MO). OD was measured at 405 nm and compared with that of standards.
Stromal-cell-derived factor (SDF)-1 RT-PCR
RNA was isolated from BMPCs cultured for 7-12 days using the Qiagen RN Easy kit (Qiagen, Santa Clarita, CA, USA). RNA was then used for the first-strand cDNA synthesis in the SuperScript Preamplification System (GIBCO, BRL, Rockville, MD, USA) in accordance with the manufacturer's instructions. SDF-1 specific primers: 5'-GAGGATCCGACGGGAAGCCC-GTCAGC; 3'-GAA-TTCACATCTTGAACCTCTTG. The annealing temperature was 58°C and the reaction proceeded for 35 cycles. The glyceraldehyde-3-phosphate dehydrogenase (GA3PD) gene was included as a reverse transcriptase polymerase chain reaction (RT-PCR) control and performed under similar conditions to normalize for the amount of RNA. Reaction products were analyzed in 2% agarose gel containing 0.25 mg/ml ethidium bromide.
Data analysis and statistics
Results are shown as the standard error about the mean (SEM) of at least three experiments each. For statistical comparison between groups, the Student paired t test or Bonferroni t test was used. Analyses were performed using the Biostatistics software developed by Stanton A Glantz (UC San Francisco, CA, USA). Flow cytometry data were analyzed using the FlowJo software.
Results
BMPCs selected by elutriation of normal human blood
When the elutriated cells from the fractions between the smaller T cells and the larger, more granular monocytes were cultured in complete medium without any other supplements, they appeared small and round on examination by phase-contrast microscopy. Some of them were nonadherent (presumably T lymphocytes) and were removed with the initial feeding of the culture. After 72 h, elongated cells with a fibroblast-like morphology and large cells with a clear, thin, adherent cytoplasm around a central nucleus made their appearance. Over the ensuing 7-14 days, they became the predominant cells in the culture (Fig. 1a). At higher magnification they could often be seen to have a splayed, spreading cap at the end and a small, central nucleus. Another cell population, consisting of larger and wider cells, with more cytoplasm and a larger nucleus, was intermediate in morphology between the large, round cells and the thinner, fibroblast-like cells (Fig. 1b). Culture conditions modified the morphology of the elutriated cells. Adding dexamethasone (100 nmol) at the initiation of the culture significantly reduced (by 60% ± 10%) the total number of cells at the 7th to 10th days and decreased the formation of fibroblast-like cells (data not shown). Cultures supplemented with a mixture of 100 nmol dexamethasone, 0.05 mmol ascorbic acid-2-phosphate, and 10 mmol β-glycerophosphate (conditions that favor the development of osteoblasts) developed only round or cuboidal cells, and not fibroblast-like ones (Fig. 1c). Dexamethasone alone added at days 6 to 8 reduced fibroblast numbers, but not the total cell numbers (data not shown).
Figure 1.
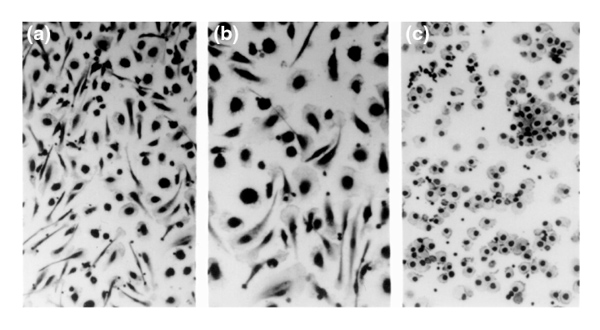
Appearance of cells isolated from a BMPC-rich elutriation fraction of healthy human blood and cultured in DMEM-20% FCS for 8 days (a, b). At this time, the predominant cells consist of both fibroblast-like cells with a central nucleus, filmy cytoplasm, and adherent pseudopods, and large, round cells with a thin, adherent cytoplasm and a round, central nucleus. Hematoxylin staining. (a) ×200; (b) ×400. (c) The appearance of cells isolated from the same BMPC-rich elutriation fraction of healthy human blood and cultured in DMEM-20% FCS supplemented at the initiation of culture with dexamethasone (10-7mol), ascorbic-acid-2-phosphate (0.05 mmol), and β-glycerophosphate (10 mmol) for 8 days. At this time, all the cells have a round or cuboidal morphology with a centrally placed nucleus. ×200. Compare with the nonsupplemented cultures in (a).
BMPC-rich elutriation fractions observed in time-lapse cinematography
Clusters of small round cells formed within 24 h. Cell processes occasionally extended from them, but these retracted minutes later (Fig. 2a). Individual cells were motile and often left the field, but the clusters remained intact. After 72 h, a few cells with a fibroblast-like morphology could be seen beneath and at the edges of the clusters. The fibroblast-like cells were much larger than the initial cells and quite mobile, extending and retracting usually about a broad, fixed cup, or pseudopod. By 6 days, a significant portion of the cells retained their elongated form and looked like the cells in Fig. 2b. Large, round cells were also present. Thus, it appears that BMPCs in the circulation were present as small, round mononuclear cells and their subsequent morphology and function were dictated by culture conditions.
Figure 2.
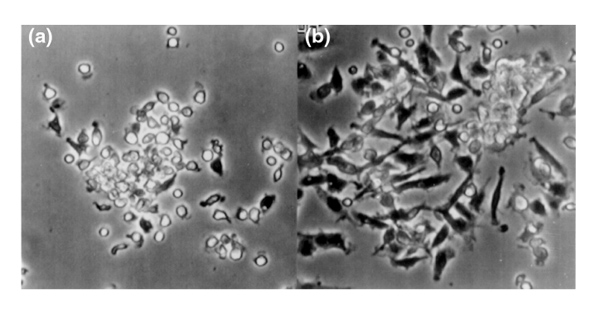
Time-lapse video cinematography of a BMPC-rich elutriation fraction cultured in DMEM-20% FCS observed by phase-contrast microscopy. At day 2 (a), there are small, round cells in clusters and a few cells with pseudopods. By day 6 (b), there are many large, fibroblast-like cells and large, round, stromal cells arising from a cluster of small, round cells.
Cell numbers in the BMPC-rich elutriation fraction from 500 ml of normal human blood
Elutriation fractions were selected for quantification of BMPCs based on cell size (intermediate between lymphocytes and monocytes) and granularity (FACS). This population comprised less than 35% lymphocytes and more than 50% monocytes. Nineteen consecutive samples had an average total cell number of 2.14 ± 0.22 (SEM) × 107, of which 64.4% ± 1.5% (SEM) were monocytes. A sub-population, estimated as 0.3-0.7% of the starting elutriation fractions, was judged to consist of BMPCs on the basis of their morphology, their strong adherence to plastic or glass, and their ability to proliferate in DMEM-20% FCS without added growth factors (ie <1% of the starting 2 × 107 elutriated cells represents 1000 to 10 000 BMPCs). Therefore, it is likely that 500 ml of normal blood will have several thousand BMPCs.
Cultures were established with 5 × 105 cells from the elutriation fractions and proliferation was measured on days 3, 10, and 17. Nonadherent cells were removed in the first 24 h and the cultures were fed twice weekly. The cells grew logarithmically, with an approximate doubling time of 2.5 days. By day 17, the initial 5 × 105 cells multiplied to 6.7 × 107 (Table 1). The culture conditions are not conducive to growth of lymphocytes or monocytes; therefore, by week 3, most of the proliferating population in cultures were mesenchymal cells (<20% CD14-staining cells; data not shown).
Table 1.
Proliferation of BMPCs
| Day | Observations | OD* | No. of cells |
| 0 | 10 | 0.098 ± 0.02 | 0.5 × 106 |
| 3 | 6 | 0.111 ± 0.28 | 1.2 × 106 |
| 10 | 3 | 0.282 ± 0.40 | 7.5 × 106 |
| 17 | 6 | 0.505 ± 0.51 | 6.7 × 107 |
OD = optical density; see Methods.
Cells from a BMPC-rich elutriation fraction of healthy human blood were cultured in DMEM-20% FCS. At days 3, 5, 8, and 11 the cultured cells were fixed, stained with anti-BMPR antibodies, and quantified with an autoanalyzer (described in Methods). The results are presented in Fig. 3 as the means of the total number of cells in six individual images. Big cells plus fibroblast-like cells constituted 37% to 59% of the total. Cells with a fibroblast-like morphology varied from 21% to 34% of the BMPCs. The 1:2 ratio of fibroblast-like to big cells remained relatively stable over 11 days of culture, even as the total cell numbers increased (see Table 1).
Figure 3.
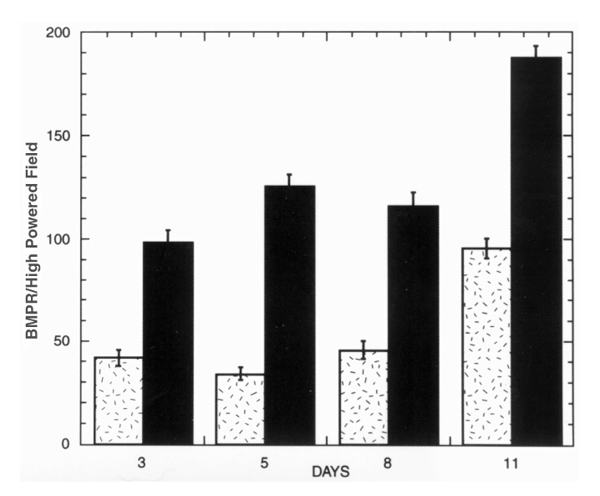
BMPC-rich elutriation fraction from healthy human blood cultured in DMEM-20% FCS. Cells were fixed, stained with anti-BMPR 1A antibody (see Methods), and analyzed on days 3, 5, 8, and 11. BMPCs were determined by morphology (fibroblast-like [hatched bars] or big cells [filled bars]) and immunoperoxidase staining. Results are presented as the means of total number of cells in six individual images. ×400. Computer image analysis (AnalySIS).
Immunohistology of BMPCs
Elutriated cells were grown on sterile 12-well glass slides (ICN) in DMEM-20% FCS and 6 to 12 days later the cells were fixed, stained, and examined by immunohistology (Table 2). The large cells and the fibroblast-like cells stained with antibodies to both vimentin and collagen type I. They were also identified by antibodies to one or the other chain (type IA and type II) of the BMPR (Fig. 4a), but did not react with an anti-type-IB-receptor-chain antibody. Monoclonal Stro-1 antibody stained most of the large BMPCs and a few of the fibroblast-like cells, while anti-CD105 reacted with both populations (Fig. 4b). BMPCs stained strongly with anti-CD44 antibody. Conventional T-cell (CD3), monocyte (CD14, CD68), and B-cell (CD20) antibodies stained neither of the two BMPC populations, nor did they react to anti-LCA (CD45), anti-VCAM (CD106), or MHC-Class II (anti-DR).
Table 2.
Antibody profile of BMPCs (at days 7-10)
| Antibody | Large cells | Fibroblastoid cells |
| Vimentin | + | + |
| Collagen type I | + | + |
| BMPR IA | + | + |
| IB | 0 | 0 |
| II | + | + |
| STRO-1 | + | 0 |
| CD3, CD14, CD20 | 0 | 0 |
| CD34, CD45, Class II | 0 | 0 |
Figure 4.
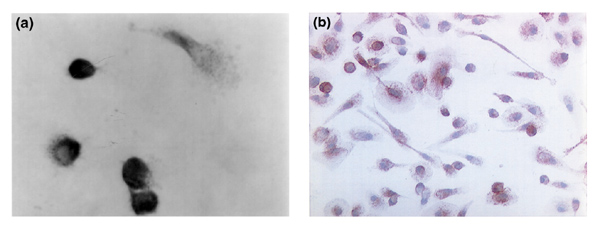
BMPC-rich elutriation fraction cultured on a sterile glass slide in DMEM-20% FCS and allowed to proliferate. (a) On day 7, the slide was washed and the cells were fixed and stained with anti-BMPR Type 1A antibody. In the background are many unstained, small, mononuclear cells. Two types of cell are stained: one looks like a fibroblast, and the other is a large, round cell with an adherent cytoplasm. (b) A similar culture on day 12, stained with anti-endoglin (CD105) antibody. The cytoplasm of almost all the large mesenchymal cells shows stippled staining with perinuclear accentuation.
Cell separation by magnetic beads to analyze the contribution of CD34+ progenitors, monocytes, and T lymphocytes to BMPC formation
BMPC-rich elutriation fractions were incubated with magnetic beads coated with specific antibodies and subsequently separated into adherent (antigen-enriched) and nonadherent (antigen-depleted) populations. These were cultured in complete medium in six-well plates and observed daily by phase-contrast microscopy. The CD34-depleted fraction always developed many examples of both types of mesenchymal cells (seven experiments). The fibroblast-like cells appeared in the CD34-depleted cultures at the same time as in untreated controls (usually day 3 or 4). There were not enough CD34+ cells to establish cultures. A representative experiment is shown in Table 3. In two additional experiments, the cells from the elutriation fraction were exposed to CD34 beads and the negative fraction was separated again on fresh CD34 beads. There was no reduction in the time of appearance or number of fibroblast-like cells. Thus, although BMPCs were present in an elutriation fraction that contained CD34+ cells, the two types of cell could be distinguished from one another.
Table 3.
Outcome of separation of BMPC-rich elutriation fraction using magnetic beads coated with specific antibodies
| Starting fraction | Post separation | ||||
| Cells (×106) | Monocytes (%) | T cells (%) | CD antibody coating beads | Cells (×106) | Fibroblasts/HPFs* |
| 8.7 | 59 | 30 | 34(-) | 6.5 | ++++ |
| 10.5 | 70 | 24 | 14(-) | 2.8 | + |
| 14(+) | 4.8 | + | |||
| 14(+/-) | 2.1 | +++ | |||
*Mean of 6-10 fields. + = up to 5; ++ = 5-10; +++ = 10-15; ++++ = 15-20.
Findings with anti-CD14 beads were somewhat different. Both CD14+ and CD14- eluates developed fibroblast-like cells in culture, but the numbers were less than in unfractionated controls. In four of five experiments, the fibroblast-like cells in the CD14+ fraction appeared sooner and in greater numbers than the CD14-fraction. In all instances, however, each fraction contained both large, round cells and fibroblast-like cells and their numbers become more equal with time (usually by day 13). A representative experiment is shown in Table 3. When the CD14+ and CD14- populations were combined, the number of fibroblast-like cells and their time of appearance was the same as in unfractionated controls.
T-cell depletion had no effect on the number, morphology, or time of appearance of mesenchymal cells (data not shown).
Pluripotent BMPC-fibroblast, osteoblast, and adipocyte formation
BMPC-rich elutriation fractions cultured in complete medium supported growth of fibroblasts (spindle-shaped cells stained by antibodies to vitronectin and type I collagen). When the same elutriated cells were supplemented with dexamethasone, ascorbic acid, and β-glycerophosphate their morphology altered, and they became uniform, polygonal cells reminiscent of osteoblasts (Fig. 1c). By 10 days, many of the cells stained for AP (not shown). Over the next 1 to 2 weeks, a subpopulation (approximately 30%) of large cells (which were 3- to 6-fold bigger than monocytes) developed. They accumulated an ill-defined pericellular matrix and the osteoblast-specific protein osteocalcin (Fig. 5a and b). In the same supplemented cultures were sudanophilic adipocytes (Fig. 5a). Another large cell present after 1 week in the supplemented cultures had multiple nuclei (Fig. 6a) and stained for tartrate-resistant acid phosphatase (TRAP) and the vitronectin receptor (Fig. 6b), which are features of osteoclasts (OCs).
Figure 5.
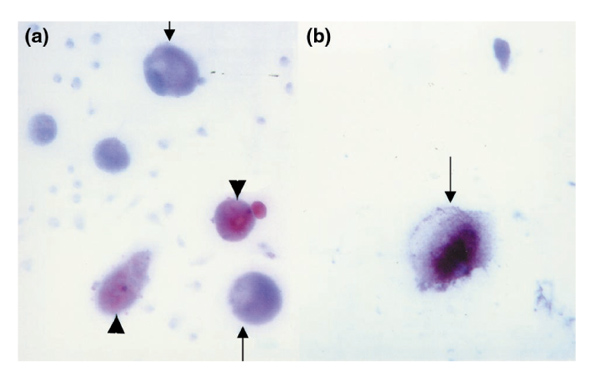
BMPC-rich elutriation fraction cultured in DMEM-20% FCS supplemented with dexamethasone, ascorbic acid, and β-glycerophosphate (as described in Fig. 1). After 20 days, about one-third of the cells are very large. Their cytoplasm stains with an anti-calcitonin antibody. Immunoperoxidase staining is also observed in the matrix formed around some of the largest cells. In the same supplemented cultures are large cells containing neutral lipid (Sudan IV; red stain). (a) Shows both anti-calcitonin-stained cells (arrows) and sudanophilic cells (arrowheads) in the same supplemented cultures (×250) and (b) is at higher magnification (×400) showing a cell and its surrounding matrix stained with an anticalcitonin antibody (arrow).
Figure 6.
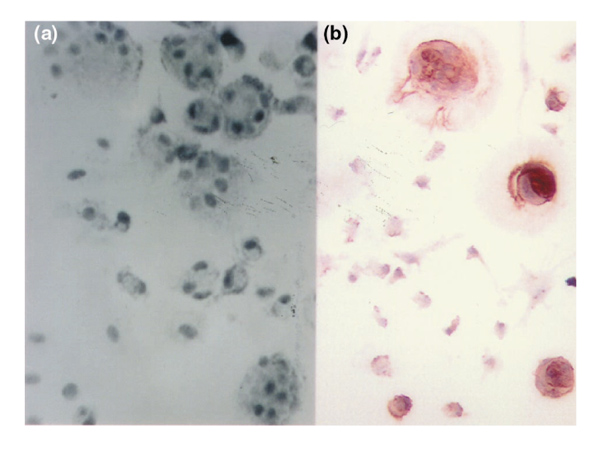
BMPC-rich elutriation fraction cultured in DMEM-20% FCS supplemented with dexamethasone, ascorbic acid, and β-glycerophosphate (as described in Fig. 1) and examined daily thereafter. (a) By day 7 there are many large, multinucleated cells (phase-contrast microscopy). (b) These cells stain with an anti-vitronectin receptor antibody (×250).
OCs developed in the supplemented BMPC cultures because both monocytes and stem cells (and/or pre-osteoblasts) were present together in the elutriated cells.
Alkaline phosphatase production
BMPC-rich elutriation fractions from four separate blood packs were cultured in complete medium with varying concentrations of BMP2 for 5 days and AP in the supernatants was measured (Fig. 7). The lowest concentration of BMP2 (1 ng/ml) caused a significant increase in AP activity (P = 0.004). This represents an increased AP production per cell, because BMP2 had no effect on BMPC proliferation over 5 days (data not shown).
Figure 7.
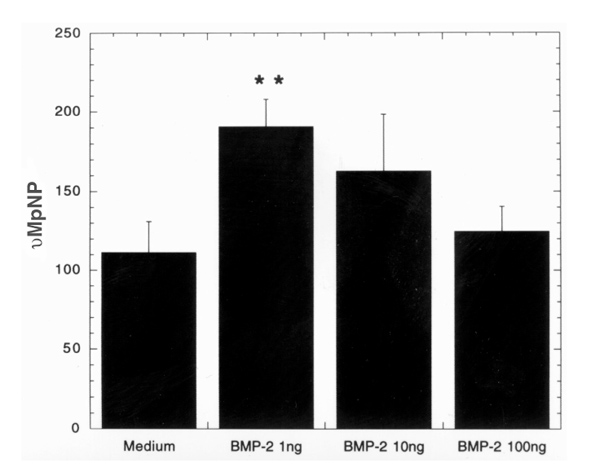
BMPC-rich elutriation fractions (n = 4) cultured in DMEM-20% FCS with varying concentrations of BMP2 for 5 days. Supernatants were collected and analyzed for alkaline phosphatase activity (AP; the release of pNP is a measure of AP activity). The lowest concentration of BMP2 (1 ng/ml) caused a significant increase in AP activity. **P > 0.004.
Stromal-cell-derived factor 1
SDF-1 is a potent CXCα chemokine produced by bone-marrow spindle-shaped stromal cells and other cells of mesenchymal origin, but not blood leukocytes [12,13]. RT-PCR analysis for SDF-1 mRNA expression in two cDNA samples of cultured BMPCs (lanes 2 and 3) and in a cDNA from RA synoviocytes (lane 4) are shown in Fig. 8.
Figure 8.
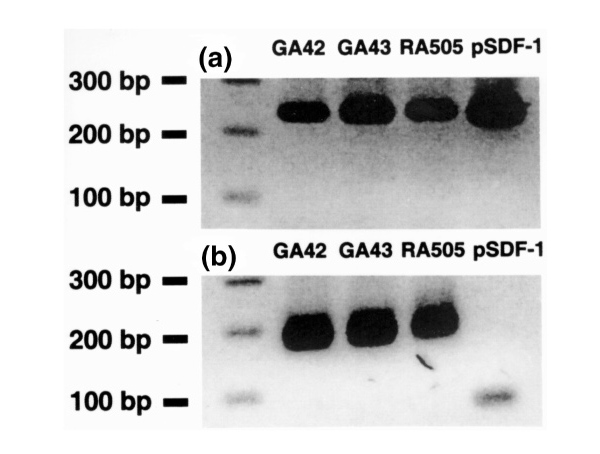
RT-PCR analysis for expression of mRNA for (a) the chemokine SDF-1 and (b) the housekeeping gene GA3PD in cultured BMPCs (GA42, GA43), and a rheumatoid arthritis synovial fibroblast line (RA505 passage 4). Note the similar RT-PCR fragment size in the samples to that in an SDF-1 plasmid (lane 5) included as a positive control. Conditions for RT-PCR and the primers used are described in the methods section.
Each sample displayed an amplification of a PCR fragment of the expected size for SDF-1 (296 bp) and was similar to the positive control (lane 5), a sequenced plasmid containing human SDF-1β cDNA.
Discussion
The abilities to self-replicate and to give rise to daughter cells that undergo an irreversible terminal differentiation are features of stem cells [14,15]. The best-characterized are HSCs and their progeny. Friedenstein et al proposed a similar scheme for mesenchymal cells and showed that bone marrow contained primitive cells that could generate progenitors committed to one or another mesenchymal line [2]. Such cells are called MSCs [1]. Conditions that direct marrow MSCs along myogenic [16], adipogenic [17,18], osteogenic [18,19,20], chondrogenic [19,21], and stromal pathways [22] have been defined. For instance, exposure of fibroblast-like marrow MSCs in vitro to optimal concentrations of dexamethasone, ascorbic acid and β-glycerophosphate induces a cuboidal morphology, upregulates AP and osteocalcin expression, and a mineralized (hydroxyapatite) matrix [20].
Lineage differentiation signals can be subtle. Dexamethasone at 10-9mol/l supports adipocyte differentiation, whereas osteogenesis is favored at 10-7 mol/l [19]. Human marrow MSCs obtained by Ficoll density gradient fractionation (1.078 g/ml) and cultured in 25% serum (half horse and half fetal calf) supplemented with hydrocortisone (1 μmol) gives rise to a heterogeneous population, in which fibroblast-like cells do not predominate [22]. Separation of the same population of marrow MSCs on a Percoll density gradient (1.090 g/ml) and culture in carefully selected 10% FCS resulted in a homogeneous population of spindle-shaped fibroblast cells. The higher-density Ficoll may isolate cells that sediment through the Percoll solution used for the marrow MSC isolation. Elutriation, as used in this investigation, probably selects a somewhat different population.
The CD34 status of mesenchymal cells is disputed [7,8]. A minority of adult marrow cells express CD34. The antigen is present on pluripotent HSCs, and all unipotent myeloid and erythroid-colony-forming cells [23], but CD34 is also recognized on vascular endothelial cells, basement-membrane structures, and dendritic and perifollicular cells in human skin [24,25]. Simmons and Torok-Storb separated human marrow cells on the basis of their CD34 expression [26]. More than 95% of the detectable colony forming unit fibroblasts were recovered in the adherent CD34+ population, but their CD34 density was much less than on CD34 high HSCs. Furthermore, only 5% of the CD34+ marrow cells reacted with the monoclonal antibody Stro-1, which identifies marrow MSCs [6]. All these studies were done on marrow MSCs before culture, because after culture in vitro, the same stromal cells no longer react with anti-CD34 antibodies [26]. Likewise, although CD34+ cells are identified in tissue sections of human umbilical vein endothelial cells, they are not found in vitro [27]. These and other reports suggest the CD34 glycoprotein is either down-regulated or modified in vitro to a form that is not reactive with the usual anti-CD34 antibodies [23]. Therefore, the failure of Fernandez et al to demonstrate CD34 on circulating stromal cells mobilized by growth factors was probably because the cells had been in culture for 10 days [7]. Similarly, the inability of Majumdar et al to demonstrate CD34 staining was on first-passage marrow MSCs [22]. In vitro culture conditions, however, cannot explain our failure to eliminate BMPCs in fresh elutriation fractions with anti-CD34-coated magnetic beads, a technique widely used to harvest CD34+ HSCs from growth-factor-mobilized human blood. The absorptions were performed on fresh BMPC-rich elutriation fractions. Therefore, BMPCs either lack CD34 or have only a very low density of this glycoprotein.
The CD34+ cells called fibrocytes, which are present in monocyte fractions of human blood and develop a fibroblast morphology when grown on fibronectin, have features identical to those of circulating vascular endothelial cell progenitors [28] and are probably not BMPCs.
Bone morphogenetic proteins (BMPs) were originally identified as proteins that induced bone formation at extraskeletal sites [29]. Currently, there are 20 or more known BMPs, all members of a larger TGF-β superfamily. BMPs are involved in morphogenesis and embryogenesis, influencing bone, cartilage, and skeletal formation [29,30,31,32]. Much of this information comes from animal cells and embryos, but the addition of BMP2 to cultured postnatal human bone marrow `preosteoblastic' cells in the presence of β-glycerophosphate and ascorbic acid increases the gene message and protein production of AP, osteopontin, bone sialoprotein, osteocalcin, and α-1 collagen [33]. BMPCs develop into osteocalcin-producing cells (Fig. 5a) and make increased amounts of AP in response to BMP2 (Fig. 7). This cannot be explained by proliferation, because BMP2 reduces the number of marrow MSCs in either serum or serum-free conditions [33].
BMPRs belong to the TGF-β receptor family of serine/threonine kinases [34]. Both type I and type II BMPRs bind their respective ligands, but heterodimerization is required for a signal to be transduced [34,35]. For instance, coexpression of type II BMPR with either IA or IB BMPR increases ligand-binding affinity and dramatically enhances biologic activity [11]. Human marrow MSCs express BMP2/4 type I and II receptors as shown in studies employing BMP2 as ligand in the presence or absence of a 100-fold excess of a competitor [33]. BMP structure is conserved across species, and antibodies to type I and typeII receptors react equally well with murine and human mesenchymal cells, but not with hematopoietic cells [34]. This is consistent with our findings that polyclonal antibody to BMPRs can be used to identify BMPCs and constitutes strong evidence that the circulating cells described in this report are mesenchymal precursors.
BMPCs stain with the Stro-1 monoclonal antibody made against human bone-marrow stromal cells [6]. Stro-1+ cells cultured in an osteogenic medium exhibit three markers of differentiated bone: AP; 1,25-dihydroxyvitamin-D3-dependent induction of osteocalcin; and a mineralized matrix (hydroxyapatite) [36,37]. Stro-1 is expressed by BMPCs, but the antibody also stains pericytes, cells that surround small-vessel endothelium. Pericytes, which are of mesodermal origin, can also differentiate into a variety of cell types, including osteoblasts and adipocytes (reviewed in [38]). The reactivity of pericytes with BMPR antibodies is not known, but we have used BMPR antibodies to analyze mesenchymal cells in synovial tissues. The antibodies identify large cells in the inflamed joint lining, but they do not stain blood vessels of either normal or inflamed synovium (Marinova-Mutafchieva, personal communication).
SH-2, a proprietary antibody developed against isolated bone marrow MSCs [1,3], was not available when these studies were performed. When SH-2 was identified as endoglin (CD105) [4], the cells in a BMPC-enriched elutriation fraction of blood were examined with an anti-CD105 monoclonal antibody. The strong cytoplasmic staining of the large blood mesenchymal cells (Fig. 4b) is more evidence of the great similarity of marrow MSCs and circulating BMPCs.
Progenitor and precursor B cells require close contact with marrow MSCs for growth and maturation. Mouse marrow MSCs contain the gene for a protein (termed either stromal-cell-derived factor 1 (SDF-1) or pre-B-cell-growth-stimulating factor (PBSF) [12,13]. SDF-1 is a powerful CXC chemokine that recruits circulating lymphocytes, monocytes, and CD34+hematopoietic progenitors, but not neutrophils [39,40]. PBSF is responsible for converting `early' B cells into immunoglobulin-producing cells [41]. SDF-1 mRNA is constitutively expressed in many tissues, unlike other chemokines, which are only induced [41,42]. SDF-1 is expressed in marrow MSCs, dermal fibroblasts, and synovial fibroblasts, but not HSCs. The demonstration of constitutive expression of SDF-1 mRNA in cultured BMPCs (Fig. 8) and as protein in supernatants from cultured BMPCs (data not presented) is additional evidence that BMPCs are of mesenchymal lineage.
Human OCs arise from HSCs in close proximity to stromal cells or from blood monocytes. The OC is a TRAP-positive, large, multinucleated cell with receptors for calcitonin and vitronectin (αvβ3) (Fig. 5a) and the capacity to form resorption lacunae in bone slices [43]. Osteoblast production and OC production are tightly linked and regulated. Osteoblasts facilitate OC formation by providing physical support and critical soluble factors [43]. Our observation of spontaneous formation of cells with the morphology and phenotype of OCs in monocyte-rich (65%) elutriation fractions is best explained by the simultaneous presence of BMPCs in the same fractions.
More than 100 normal individuals had CD34- mononuclear cells in a fraction of elutriated blood cells that fulfilled criteria for mesenchymal precursors or stem cells. They proliferated rapidly in culture, had an adherent, spread morphology, displayed cytoskeletal, cytoplasmic, and surface markers of mesenchymal precursors, and had a capacity for differentiation into fibroblast, osteoblast, and adipocyte lineages. Thus, autologous blood could be an important source of cells for tissue engineering and gene therapy [44,45,46]. In addition, the finding of similar cells in the inflammatory joint fluids and synovial tissues of patients with RA suggests they may play a role in the pathogenesis of this disease [47].
Acknowledgments
Acknowledgements
This work was done at the Kennedy Institute of Rheumatology, London, while the first author (Nathan J Zvaifler) was on sabbatical leave from the University of California, San Diego. He is grateful to the members of the faculty and staff who facilitated these studies, especially the students and research staff in Professor Marc Feldmann's laboratory for their expertise in elutriation, M Kahan for cytofluorography, and S Quarantino, F Brennan, and Y Chernajovsky for helpful suggestions and discussion. Jennifer Allen provided excellent secretarial services. This work was supported by the Arthritis Research Campaign Core Grant to the Kennedy Institute and by the National Institutes of Health grants nos. AR40770 and AR45347 (NJ Zvaifler).
References
- Caplan AI. Mesenchymal stem cells. J Orthop Res. 1991;9:641–650. doi: 10.1002/jor.1100090504. [DOI] [PubMed] [Google Scholar]
- Friedenstein AJ, Gorskaja JF, Kulagina NN. Precursor cells of mechanocytes. ExpHematol. 1976;4:267–274. [PubMed] [Google Scholar]
- Bruder SP, Jaiswal N, Haynesworth SE. Growth kinetics, self-renewal, and the osteogenic potential of purified human mesenchymal stem cells during extensive subcultivation and following cryopreservation. . J Cell Biochem. 1997;64:278–294. doi: 10.1002/(sici)1097-4644(199702)64:2<278::aid-jcb11>3.0.co;2-f. [DOI] [PubMed] [Google Scholar]
- Barry FP, Boynton RE, Haynesworth S, Murphy JM, Zaia J. The monoclonal antibody SH-2, raised against human mesenchymal stem cells, recognizes an epitope on endoglin (CD105). Biochem Biophys Res Commun. 1999;265:134–139. doi: 10.1006/bbrc.1999.1620. [DOI] [PubMed] [Google Scholar]
- Rosen V. Signaling pathways in skeletal formation. A role for BMP receptors. In Ann NY Acad Sci No 785: Molecular and Developmental Biology of Cartilage Edited by de Crombrugghe B. 1996. pp. 56–59. [DOI] [PubMed]
- Simmons PJ, Torok-Storb B. Identification of stromal cell precursors in human bone marrow by a novel monoclonal antibody Stro-1. Blood. 1991;78:55–62. [PubMed] [Google Scholar]
- Fernandez M, Simon V, Herrera G, Cao C, Del Favero H, Minguell JT. Detection of stromal cells in peripheral blood progenitor cell collections from breast cancer patients. Bone Marrow Trans. 1997;20:265–271. doi: 10.1038/sj.bmt.1700890. [DOI] [PubMed] [Google Scholar]
- Bucala R, Spiegel LA, Chesney J, Hogan M, Cerami A. Circulating fibrocytes define a new leukocyte subpopulation that mediates tissue repair. . Mol Med. 1994;1:71–81. [PMC free article] [PubMed] [Google Scholar]
- Westergren-Thorsson G, Onnervik PO, Fransson LA, Malmstrom A. Proliferation of cultured fibroblasts is inhibited by L-iduronate containing glycosaminoglycan. J Cell Physiol. 1991;147:523–530. doi: 10.1002/jcp.1041470319. [DOI] [PubMed] [Google Scholar]
- Ten Dijke P, Yamashita H, Ichigo H, Franzer P, Laiho M, Miyazono K, Helden CH. Characterization of type I receptors for transforming growth factor-beta and activin. Science. 1994;264:101–104. doi: 10.1126/science.8140412. [DOI] [PubMed] [Google Scholar]
- Ten Dijke P, Yamashita H, Sampath TK, Reddi AH, Estevez M, Riddle DL, Ichijo H, Heldin CH, Miyanzono K. Identification of type I receptors for osteogenic protein-1 and bone morphogenetic protein-4. J Biol Chem. 1994;269:16985–16988. [PubMed] [Google Scholar]
- Tashiro T, Tada H, Heilker R, Shirozu R, Nakano T, Honjo T. Signal sequence trap: a cloning strategy for secreted proteins and type I membrane proteins. Science. 1993;61:600–603. doi: 10.1126/science.8342023. [DOI] [PubMed] [Google Scholar]
- Shirozu M, Nakano T, Inazawa J, Tashiro K, Tada H, Shinohara T, Honjo T. Structure and chromosomal location of the human stromal derived factor-1. Genomics. 1995;28:495–500. doi: 10.1006/geno.1995.1180. [DOI] [PubMed] [Google Scholar]
- Owen J. Marrow stromal cells. Cell Sci (Suppl) 1988;10:63–76. doi: 10.1242/jcs.1988.supplement_10.5. [DOI] [PubMed] [Google Scholar]
- Dennis JE, Merream A, Awadallah A, Yoo JU, Johnstone B, Caplan A. A quadripotential mesenchymal progenitor isolated from the marrow of an adult mouse. J Bone Mineral Res. 1999;5:700–709. doi: 10.1359/jbmr.1999.14.5.700. [DOI] [PubMed] [Google Scholar]
- Ferrari G, Cusella-de Angelis G, Coletta M, Paolucci E, Stornaueolo A, Cassu G, Mavilio F. Muscle regeneration by bone-marrow-derived myogenic progenitors. Science. 1998;279:1528–1530. doi: 10.1126/science.279.5356.1528. [DOI] [PubMed] [Google Scholar]
- Kelly KA, Gimble JM. 1,215-Dihydroxy vitamin D inhibits adipocyte differentiation and gene expression in murine bone marrow stromal cell clones and primary cultures. Endocrinology. 1998;139:2622–2628. doi: 10.1210/endo.139.5.5970. [DOI] [PubMed] [Google Scholar]
- Grigoriadis AE, Heersche JN, Aubin JE. Differentiation of muscle, fat, cartilage and bone from progenitor cells present in a bone marrow-derived clonal cell population. Effect of dexamethasone. J Cell Biol. 1988;106:2139–2151. doi: 10.1083/jcb.106.6.2139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thies RS, Baudy M, Ashton B, Kurtzberg L, Wozney J, Rosen V. Recombinant human bone morphogenetic protein-2 induces osteoblast differentiation in W-20-17 stromal cells. Endocrinology. 1992;130:1318–1324. doi: 10.1210/endo.130.3.1311236. [DOI] [PubMed] [Google Scholar]
- Jaiswal N, Haynesworth SE, Caplan AI, Bruder SP. Osteogenic differentiation of purified, culture expanded human mesenchymal stem cells in vitro. J Cell Biochem. 1997;64:295–312. [PubMed] [Google Scholar]
- Yoo JU, Barethel TS, Nishimura L, Solchaga L, Caplan AI, Goldberg VM, Johnstone G. The chondrogenic potential of human bone-marrow-derived mesenchymal progenitor cells. J Bone Joint Surg (Am) 1998;80:1745–1757. doi: 10.2106/00004623-199812000-00004. [DOI] [PubMed] [Google Scholar]
- Majumdar MD, Thiede MA, Mosca JD, Moorman M, Gerson SL. Phenotypic and functional comparison of cultures of marrow derived mesenchymal stem cells (MSCs) and stromal cells. J Cell Physiol . 1998;176:57–66. doi: 10.1002/(SICI)1097-4652(199807)176:1<57::AID-JCP7>3.0.CO;2-7. [DOI] [PubMed] [Google Scholar]
- van de Rijn M, Rouse RV. CD34. A review. Applied Immunohistochem. 1994;2:71–80. [Google Scholar]
- Nickoloff BJ. The human progenitor cell antigen (CD34) is localized on endothelial cells, dermal dendritic cells and perifollicular cells in formalin fixed normal skin. Arch Dermatol. 1991;127:523–529. [PubMed] [Google Scholar]
- Brown J, Greaves MF, Molgaard HV. The gene encoding the stem cell antigen CD34 is conserved in the mouse and expressed in hematopoietic cell lines, brain and embryonic fibroblasts. Inter Immunol. 1991;3:75–184. doi: 10.1093/intimm/3.2.175. [DOI] [PubMed] [Google Scholar]
- Simmon PJ, Torok-Storb B. CD34 expression by stromal precursors in normal adult human bone marrow. Blood. 1991;78:2848–2853. [PubMed] [Google Scholar]
- Fina L, Molgaard H-V, Robertson D, Bradley NJ, Monaghan P, Delia D, Sutherland DR, Baker M, Greaves MF. Expression of the CD34 gene in vascular endothelial cells. . Blood. 1990;75:2417–2426. [PubMed] [Google Scholar]
- Asahara T, Murohara T, Sullivan A, Silver M, van der Zee R, Li T, Witzenbichler B, Schatteman G, Isner J. Isolation of putative progenitor endothelial cells for angiogenesis. Science. 1997;275:964–967. doi: 10.1126/science.275.5302.964. [DOI] [PubMed] [Google Scholar]
- Reddi AH. Regulation of cartilage and bone differentiation by bone morphogenetic proteins. Curr Opin Cell Biol. 1992;4:850–855. doi: 10.1016/0955-0674(92)90110-x. [DOI] [PubMed] [Google Scholar]
- Wang EA, Rosen V, D'Allesandro JS, Bauduy M, Cordes P, Harada T, Isreal DJ, Hiwick RM, Kerns KM, La Pan P, Luxenberg DP, McQuaid D, Nove J, Wozney JM. Recombinant human bone morphogenetic protein induces cartilage and bone differentiation. Proc Natl Acad Sci USA. 1990;87:2220–2224. doi: 10.1073/pnas.87.6.2220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Enomoto-Iwamoto M, Iwamoto M, Mukudai Y, Kawakami Y, Nohno T, Higuchi Y, Takemoto S, Ohuchi H, Noji S, Kurisu K. Bone morphogenetic protein signaling is required for maintenance of differentiated phenotype, control of proliferation, and hypertrophy in chondrocytes. J Cell Biol . 1998;140:409–418. doi: 10.1083/jcb.140.2.409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lyons KM, Pelton RW, Hogan BL. Organogenesis and pattern formation in the mouse: RNA distribution patterns suggest a role for bone morphogenetic protein 2 (BMP 2-A). Development. 1990;109:833–844. doi: 10.1242/dev.109.4.833. [DOI] [PubMed] [Google Scholar]
- Lecanda F, Avioli LV, Cheng SL. Regulation of bone matrix protein expression and induction of differentiation of human osteoblasts and human bone marrow stromal cells by bone morphogenetic protein. Cell Biochem. 1997;67:386–396. [PubMed] [Google Scholar]
- Yamashita H, Ten Dijke P, Heldin C-H, Miyazono K. Bone morphogenetic protein receptors. Bone. 1996;19:569–574. doi: 10.1016/s8756-3282(96)00259-1. [DOI] [PubMed] [Google Scholar]
- Massague J. TGF-beta signaling: receptors, transducers and Mad proteins. Cell. 1996;85:947–950. doi: 10.1016/s0092-8674(00)81296-9. [DOI] [PubMed] [Google Scholar]
- Gronthos S, Greaves SE, Ohta S, Simmons PJ. The STRO-1 fraction of adult human bone marrow contains the osteogenic precursors. . Blood. 1994;84:4164–4173. [PubMed] [Google Scholar]
- Stewert K, Screen J, Jeffries CM, Walsh S, Beresford JN. Coexpression of the STRO-1 antigen and alkaline phosphatase in cultures of human bone and marrow cells. J Bone Miner Res. 1996;11:208–211. [Google Scholar]
- Doherty MJ, Canfield AE. Gene expression during vascular pericyte differentiation. Crit Rev Eukaryotic Gene Expr. 1999;9:1–17. [PubMed] [Google Scholar]
- Bleul CC, Fuhlbrigge RC, Casanovas JM, Aiuti A, Springer TA. A highly efficacious lymphocyte chemoattractant, stromal cell derived factor 1 (SDF-1). J Exp Med. 1996;184:1101–1109. doi: 10.1084/jem.184.3.1101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aiuti A, Webb IJ, Bleul C, Springer T, Gutierrez-Ramos JC. The chemokine SDF-1 is a chemoattractant for human CD34+ hematopoietic progenitor cells and provides a new mechanism to explain the mobilization of CD34+ progenitors to peripheral blood. J Exp Med. 1997;185:111–120. doi: 10.1084/jem.185.1.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagasawa T, Kikutani H, Kishimoto T. Molecular cloning and structure of a pre-B cell growth-stimulating factor. Proc Natl Acad Sci USA. 1994;91:2305–2309. doi: 10.1073/pnas.91.6.2305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagasawa T, Hirota S, Tachibana N, Takakura S, Nishikawa Y, Kitamura N, Yoshida H, Kikutani T, Kishimoto T. Defects of B cell lymphopoiesis and bone marrow myelopoiesis in mice lacking the CXC chemokine PBSF/SDF-1. . Nature. 1996;382:635–638. doi: 10.1038/382635a0. [DOI] [PubMed] [Google Scholar]
- Suda T, Takahashi N, Martin TJ. Modulation of osteoclast differentiation. Endocr Rev. 1992;13:66–80. doi: 10.1210/edrv-13-1-66. [DOI] [PubMed] [Google Scholar]
- Prockop DJ. Marrow stromal cells as stem cells for nonhematopoietic tissues. Science. 1997;276:71–74. doi: 10.1126/science.276.5309.71. [DOI] [PubMed] [Google Scholar]
- Gerson SL. Mesenchymal stem cells: no longer second class citizens. . Nature Med. 1999;5:262–264. doi: 10.1038/6470. [DOI] [PubMed] [Google Scholar]
- Pittinger MP, Mackay AM, Beck SC, Jaiswal RM, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S, Marshak D. Multilineage potential of human mesenchymal stem cells. Science. 1999;284:143–147. doi: 10.1126/science.284.5411.143. [DOI] [PubMed] [Google Scholar]
- Marinova-Mutafchieva L, Taylor P, Funa K, Maini RN, Zvaifler NJ. Mesenchymal cells expressing bone morphogenetic protein receptors are present in the rheumatoid arthritis joint. Arthritis Rheumat . 2000. [DOI] [PubMed]