

Abstract
Our results show that cytokines derived from macrophages play an important role in pathogenesis of arthritis triggered by CpG oligodinucleotide (CpG ODN). IL-12 is in this respect an important immunomodulator during the development of joint inflammation.
Keywords: arthritis, CpG dinucleotides, cytokine, IL-12 knockout
Synopsis
Introduction:
Bacterial infections can be localized to the joints, causing septic arthritis, the most rapidly progressing joint disease. We have recently shown that unmethylated CpG motifs of bacterial DNA give rise to arthritis characterized by an influx of monocytic, Mac-1 antibody positive (Mac-1+ ) cells and by a scarcity of T lymphocytes. Cytokines have been shown to exert an important influence in the pathogenesis of arthritis in several mouse models. Tumor necrosis factor (TNF)-α, IL-1β, IFN-γ, and IL-12 are all produced in various quantities in the joints of patients with rheumatoid arthritis and in experimental (eg collagen-induced and septic) arthritides.
Aims:
To investigate patterns of local cytokine mRNA expression for IL-1β, IL-12, TNF-α, and IFN-γ in mice with arthritis induced by CpG ODN and the role of IL-12 in the development of CpG ODN-induced arthritis.
Methods:
CpG ODN was injected into the knee joints of mice, which were then killed after various intervals (0, 1, 3, 7, 14, or 21 d). At the end of each time interval, the synovial tissues were excised under an inverted microscope. We analyzed the mRNA expression of the cytokines in synovial tissue using a technique of hybridization in situ. IL-12 P40 knockout mice were used in an analysis of the role of IL-12 in the development of CpG ODN-induced arthritis.
Results:
None of the cytokine mRNAs studied was detected in the synovia of mice injected intra-articularly with calf thymus DNA or phosphate-buffered saline solution. In contrast, after intra-articular injection of CpG ODN, the monocyte/macrophage-derived cytokines TNF-α, IL-1β, and IL-12 were induced rapidly, being detected within the first day. The expression of TNF-α mRNA peaked on day 3 and then decreased, whereas IL-1β mRNA expression was high from day 1 onwards. IL-12 mRNA rose to peak values between days 3 and 21. The T helper (Th)1 cytokine IFN-γ mRNA was undetected throughout the experiment. The arthritis had a lower incidence and was less severe in IL-12 knockout mice than in their congenic littermates. The frequency of TNF-α and IL-1β mRNA expression in synovia was lower at day 3 in IL-12 knockout mice than in wild-type mice, while IFN-γ mRNA expression was not detectable. In vitro, TNF-α levels were lower in supernatants from mononuclear cells originating from IL-12 knockout (IL–/–) mice and incubated with CpG ODN than in corresponding supernatants originating from IL-12+/+ mice.
Discussion:
Previous studies have shown that bacterial DNA and CpG ODN directly activate macrophages to secrete proinflammatory cytokines, such as TNF-α, IL-1β, and IL-12. These cytokines exert proinflammatory activities in both septic and aseptic arthritides. TNF-α is produced primarily by monocytes and macrophages and stimulates macrophage production of IL-1. These two cytokines interact synergistically, stimulating each other's release and thereby amplifying the cascade of other inflammatory mediators.
IL-12 too is produced primarily by monocytes/macrophages, but mostly in response to microbial agents. It induces differentiation of Th1 cells and the production of IFN-γ by natural killer and T cells and is involved in the inflammatory cascade as synovitis develops. Mice that are genetically unable to produce IL-12 have a decreased incidence of septic arthritis. The present study shows sustained expression of IL-12 mRNA in synovia of CpG ODN-triggered arthritis. Histopathological examination showed that arthritis had a lower incidence and was less severe in IL-12 knockout mice than in control mice. Although the differences were not statistically significant, these findings suggest that IL-12 may play a role in the pathogenesis of arthritis triggered by CpG ODN.
How does IL-12 participate in the induction of this arthritis? Our data suggest that TNF-α and IL-1β mRNA are expressed less strongly in synovial tissue of IL-12 knockout mice than in that of of wild-type controls. Furthermore, the levels of TNF-α released in vitro by spleen mononuclear cells in response to stimulation with CpG ODN were markedly lower in IL-12 knockout mice. IL-12, one of the cytokines produced in joints, has proinflammatory properties.
Introduction
Several reports on the immunostimulatory properties of bacterial DNA have recently been published. Bacterial DNA directly activates B cells, monocytes, macrophages, and dendritic cells in vitro to upregulate their expression of costimulatory molecules that drive immune responses and secrete a variety of cytokines, including high levels of interleukin (IL)-12, IL-1, and tumor necrosis factor (TNF)-α [1,2,3]. Bacterial DNA indirectly activates natural killer (NK) cells and T cells [1,4,5], whereas vertebrate DNA lacks immunostimulatory effects. Unmethylated CpG motifs are common in bacterial DNA and considerably less common in vertebrate DNA [6,7]. In addition, whereas CpG motifs in bacterial DNA are unmethylated, the great majority of C and G nucleotides are methylated in all eukaryotic organisms, including mammals [6,7]. Unmethylated CpG oligodinucleotides (CpG ODNs) are responsible for the immunostimulatory properties of bacterial DNA [8,9].
Our group has recently reported that intra-articular bacterial DNA induces arthritis [10]. Histopathological signs of the arthritis were evident within two hours and lasted for at least three weeks, and it was characterized by an influx of monocytic, Mac-1+ cells and a scarcity of T lymphocytes. Unmethylated CpG motifs were responsible for the induction of this arthritis. This proinflammatory effect of bacterial DNA did not appear to be caused by contamination with endotoxins, since mice that did not respond to lipopolysaccharides developed arthritis in response to CpG ODNs but not in response to non-DNA bacterial contamination. Neither T cells, B cells, NK cells, nor neutrophils were found to be mandatory for induction of CpG ODN-mediated arthritis, whereas macrophages played a major role in induction of arthritis triggered by CpG motifs in bacterial DNA [10].
Cytokines have been shown to exert an important role in the pathogenesis of arthritis in several mouse models. TNF-α, IL-1β, IFN-γ, and IL-12 are all produced in various quantities in the joints of patients with rheumatoid arthritis and in experimental arthritides such as collagen-induced [11,12,13,14] and septic [15] arthritis. These cytokines play an important role in the induction and development of aseptic and septic arthritis [16,17,18,19,20]. To better understand the pathogenesis of CpG ODN-mediated arthritis, we wanted to know more about the expression and role of these cytokines during its early phase. We therefore investigated the patterns of local cytokine mRNA using hybridization in situ and assessed the role of IL-12 in the induction of CpG ODN-mediated arthritis.
Materials and methods
Mice
C57BL/6 mice were purchased from ALAB (Stockholm, Sweden). IL-12 P40 knockout mice were kindly provided by Dr J Magram (Nutley, NJ, USA) [21]. All mice were housed in the animal facility of the Department of Rheumatology, University of Göteborg. Male mice 6–8 weeks of age were used in all the experiments.
Oligonucleotides and injection
Phosphorothioate-modified oligonucleotide (CpG ODN) 1668 were synthesized by Scandinavian Gene Synthesis AB (Köping, Sweden). The sequence of oligodinucleotide (ODN) 1668 (containing the CpG motif) has been reported elsewhere [10]: 5'-TCC ATG ACG TTC CTG ATG CT-3'. CpG ODN (6 μg in a volume of 20 μl) was injected into the knee joints of the mice.
Tissue preparation
The mice were killed 0, 1, 3, 7, 14, or 21 d after inoculation. At the end of each time interval, the synovial tissues from the joints of four animals were excised under an inverted microscope. These tissues were then snap-frozen in OCTTM compound (Tissue-TeK®; Sakura Finetek Europe B.V., The Netherlands) by immersion in liquid nitrogen. Frozen tissue was stored at -70°C until use. Serial 6-μm sections were cut and thaw-mounted onto probe-on-slides (Fisher Scientific, Pittsburgh, PA, USA).
Hybridization in situ
Hybridization was conducted in situ to analyze cytokine mRNA expression, as previously detailed [10]. Briefly, synthetic oligonucleotide probes (Table 1) — TNF-α, IFN-γ, IL-1β, and IL-12 (the gift of Dr Tomas Olsson, Karolinska Institute, Stockholm, Sweden) — were labeled at the 3' end using terminal deoxynucleotidyl transferase (Advanced Biotechnologies, Leatherhead, UK) and [35S]ATP (Dupont Scandinavia, Stockholm, Sweden). Sections (6 μm thick) of freshly frozen synovial tissues were thaw-mounted onto slides and hybridized with 1 × 106 cpm of labeled probe per 100 μl hybridization mixture. After emulsion autoradiography, development, and fixation, the coded slides were examined by dark-field microscopy for positive cells, which were defined as those containing >15 silver grains in a star-like distribution. The intracellular distribution of the grains was checked by light microscopy at higher magnification. The data were expressed as the number of cells (mean ± SEM) expressing mRNA per mm2 of the tissue section.
Table 1.
Oligonucleotide probes used for hybridization in situ
Histopathological examination
Joints were fixed, decalcified, and embedded in paraffin for histopathological examination. Tissue sections from knee joints were cut and stained with hematoxylin–eosin. All the slides were coded and evaluated blind. The specimens were evaluated with regard to synovial hypertrophy, pannus formation, and destruction of cartilage and sub-chondral bone [22].
TNF-α levels in supernatant and serum
Spleen mononuclear cells were prepared as described previously [20]. The cells (1×106/ml) were cultured in Iscove's complete medium (10% fetal calf serum, 5 × 10-5 m 2-mercaptoethanol, 2 mm L-glutamine, and 50 μg/ml gentamicin) and stimulated with 1 μg/ml lipopolysaccharides or 1 μm CpG ODN. The cultures were maintained in 24-well plates (Nunc; Roskilde, Denmark) at 37°C in 5% CO2 and 95% humidity. The supernatants were collected after 18 h for determination of TNF-α. TNF-α levels in supernatant and serum taken from IL-12 knockout and wild-type mice at day 3 after intra-articular inoculation with 6 μg CpG ODN were analyzed using a TNF-α ELISA kit (Genzyme, Cambridge, MA, USA).
Statistical analysis
The differences between mean values were tested for significance with the Fisher's Exact test and the Mann–Whitney U test.
Results
Kinetics of synovial cytokine mRNA expression
Various workers have suggested that locally released cytokines are a key mediator of the inflammation and joint destruction observed in inflammatory arthritis [11,12,13,14,15]. To assess the local induction of cytokines, we analyzed the expression of their mRNA in synovia, using hybridization in situ. Figure 1 shows a typical result. None of the cytokine mRNAs studied were detected in the synovia of mice given intra-articular injections of calf-thymus DNA or phosphate-buffered saline solution. In contrast, after intra-articular inoculation with CpG ODN, the monocyte/macrophage-derived cytokines TNF-α, IL-1β, and IL-12 were induced rapidly, being detected within the first day. The expression of TNF-α mRNA peaked on day 3 and then decreased, whereas that of IL-1β mRNA rose on day 1 and remained high from then onwards. IL-12 mRNA rose to peak values between days 3 and 21. The Th1 cytokine IFN-γ mRNA was undetectable throughout the experiment (Fig. 2a,b).
Figure 1.
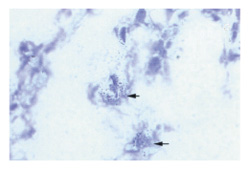
Hybridization in situ of synovium from the knee of an arthritic mouse, showing expression of TNF-α mRNA 3 d after intra-articular injection of ODN 1668.
Figure 2.
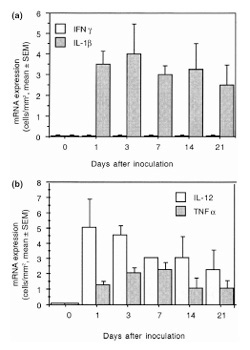
Kinetics of synovial cytokine mRNA expression in knees of mice given an intra-articular injection (6 μg) of CpG ODN. (a) IFN-γ and IL-1β; (b) TNF-α and IL-12. Each bar represents the mean number and SEM of stained cells per mm2 in synovial tissue from four mice for each interval after inoculation.
Histopathological examination
Since IL-12 mRNA expression was high during arthritis induced by CpG ODN, we studied its role further using IL-12 knockout mice. The incidence (62.5%) and severity (0.75 ± 0.75) of arthritis were decreased in IL-12 knockout mice as compared with their congenic littermates (100% and 1.33 ± 0.5, respectively) (Fig. 3a,b). These differences do not reach statistical significance.
Figure 3.
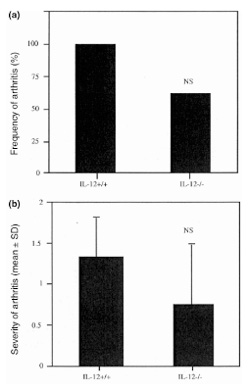
Incidence and severity of arthritis in knees of IL-12 knockout mice (IL-12–/–) (n = 8) in comparison with their wild-type littermates (IL-12+/+) (n = 9), 3 d after intra-articular inoculation with 6 μg CpG ODN. (a) Incidence. (b) Severity. NS = not significant.
Proinflammatory cytokine mRNA levels
To explore how IL-12 influenced the CpG ODN-triggered arthritis, synovial tissues taken from the knees of wild-type and IL-12 knockout mice at day 3 after intra-articular inoculation with CpG ODN were evaluated for TNF-α, IL-1β, and IFN-γ mRNA expression using hybridization in situ. As shown in Fig. 4, the frequency of TNF-α and IL-1β mRNA expression in synovia was lower at day 3 in IL-12 knockout mice than in wild-type mice, and IFN-γ mRNA expression was undetectable.
Figure 4.
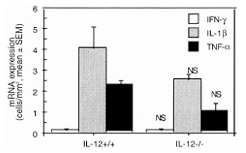
Expression of cytokine mRNA in synovial tissue from knees of IL-12 knockout mice (IL-12–/–) in comparison with their wild-type littermates (IL-12+/+), 3 d after intra-articular inoculation with 6 μg CpG ODN. Each bar represents the mean number and SEM of positive cells per mm2 in synovial tissue from four mice. NS = not significant.
TNF-α levels in supernatant and serum
To assess why the incidence and severity of CpG ODN-triggered arthritis were decreased in IL-12 knockout mice, we compared TNF-α levels in IL-12 knockout mice with those in wild-type mice, using CpG ODN as stimulus in vitro or in vivo. The reason for this approach is that TNF-α is an essential mediator of CpG ODN-mediated arthritis [10]. TNF-α levels in supernatants from mononuclear cells stimulated with CpG ODN were lower in IL-12 knockout mice than in the wild-type control mice (Fig. 5). In contrast, the TNF-α levels were similar in supernatants from mononuclear cells stimulated with lipopolysaccharides, irrespective of IL-12 phenotype (Fig. 5). This suggests that the TNF-α response by macrophages to CpG ODN stimulation is at least partly influenced by expression of IL-12. Serum TNF-α levels collected at day 3 after intra-articular inoculation with 6 μg CpG ODN from wild-type and IL-12 knockout mice did not show detectable amounts of this cytokine.
Figure 5.
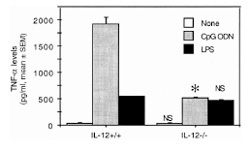
TNF-α production in vitro by spleen mononuclear cells from IL-12 knockout mice (IL-12–/–) in comparison with cells from their wild-type littermates (IL-12+/+), after stimulation with CpG ODN (1 μm) or lipopolysaccharides (LPS) (1 μg/ml). Values are means and SEM from three mice per group. *P < 0.05. NS = not significant.
Discussion
Bacterial DNA in general, and unmethylated CpG ODN in particular, trigger joint inflammation and thus may play a pathogenic role in septic arthritis. Macrophages participate in innate cellular immunity and initiate many host defense responses. How does CpG ODN activate macrophages and induce arthritis? Previous studies have demonstrated that bacterial DNA and CpG ODN activate macrophages directly [23]. The first step of activation comprises the uptake of bacterial DNA or synthetic oligonucleotides by macrophages in a saturable, sequence-independent, temperature- and energy-dependent fashion [24,25] into an acidified intracellular compartment, where DNA becomes degraded to oligodeoxynucleotides [26]. Once there, unmethylated CpG dinucleotides activate the stress-kinase/jun pathway within minutes, yielding transcriptionally active activating protein-1 and NF-κB [27]. These transcription factors control mRNA expression of a variety of cytokines and secretion of proinflammatory cytokines, such as TNF-α, IL-1β, and IL-12 [28]. These cytokines are considered to exert proinflammatory activities both in septic and aseptic arthritides [16,17,18,19,20].
TNF-α is produced primarily by monocytes and macrophages and acts as a mediator of inflammation and host responses to invasion by microbes [29]. It activates endothelial cells, upregulates expression of adhesion molecules, and stimulates macrophage production of IL-1 [29]. IL-1 and TNF-α act synergistically, stimulating each other's release and thereby amplifying the cascade of other inflammatory mediators [16]. High levels of TNF-α and IL-1 are found in the joints of patients with rheumatoid arthritis and such experimental arthritides as collagen-induced arthritis and septic arthritis [11,12,13,14]. A single intra-articular injection of IL-1β or TNF-α can induce acute synovitis [30,31]. In collagen-induced arthritis, neutralization of TNF-α and IL-1 lessens inflammation and joint destruction [32,33]. The incidence and severity of this arthritis are reduced significantly in TNF-α knockout mice [10].
IL-12 is a heterodimer consisting of two disulfide-linked subunits (P35 and P40). It is produced mainly by monocytes and macrophages, mostly in response to microbes. It induces differentiation of Th1 cells and the production of IFN-γ by NK and T cells. It takes part in the inflammatory cascade during the development of synovitis. Previous studies have shown that the role of IL-12 in the development of collagen-induced arthritis appears to depend upon a number of factors, including the timing of its administration, its dosage, and the immunization protocol used [33,34]. In addition, mice that are genetically unable to produce IL-12 display a decreased incidence of septic arthritis [35]. The present study shows sustained expression of IL-12 mRNA in synovia of mice with CpG ODN-triggered arthritis. Histopathological examination revealed that both the incidence and the severity of arthritis in IL-12 knockout mice were approximately half those in control animals, although the large within-group variation kept this difference from reaching statistical significance (see Fig. 3b). Taken altogether, these data suggest that IL-12 plays a role in the pathogenesis of arthritis triggered by CpG ODN. Further experiments should be performed to confirm this relation.
How might IL-12 contribute to the induction of CpG ODN-triggered arthritis? Previous studies showed that it might exert its influence in three ways. First, IFN-γ is an important intermediate for the action of IL-12, which is known to be able to induce IFN-γ production by NK and T cells [36]. However, our data show that IFN-γ mRNA was expressed neither in wild-type nor in IL-12 knockout mice. This finding suggests that the amelioration of CpG ODN-triggered arthritis in IL-12 knockout mice was not due to downregulation of IFN-γ production. A second possible means by which IL-12 exerts its influence is through its upregulation of B-cell production of autoantibodies [14,37]. Again, our previous studies revealed that B cells are not important for induction of this arthritis [10]. Finally, IL-12 could be thought to promote arthritis by favoring the production of proinflammatory cytokines other than IFN-γ. Indeed, previous studies in a model of pulmonary mycobacterial infection suggested that IL-12 is necessary for the local release of TNF-α [38,39], and that neutralization of IL-12 using monoclonal antibodies lowers production of TNF-α. We have recently shown that TNF-α is of importance in mediation of both septic [19] and CpG ODN-triggered arthritis [10]. In contrast to macrophage-derived cytokines, IFN-γ mRNA expression was not detected in arthritic synovia. This finding is not surprising, since there is a scarcity of T cells, a major source of IFN-γ, in CpG ODN-triggered arthritis [10], and NK cells are not important mediators for this arthritis [40].
Our data present in situ cytokine expression in CpG ODN-triggered arthritis. In the light of these data, we believe that IL-12, one of the cytokines produced in joints, has pro-inflammatory properties.
Acknowledgments
Acknowledgements
We thank Ing-Marie Nilsson and Zai-qing Liu for excellent technical assistance. The work was supported by grants from the Göteborg Medical Society, the Swedish Association against Rheumatism, King Gustaf V's Foundation, the Nanna Svartz Foundation, The Swedish Medical Research Council, the University of Göteborg, the A-G Crafoord Foundation, Börje Dahlin Foundation, The Inflammation Network, the Infection and Vaccination Network, and the A M E Wolff Foundation.
References
- Klinman DM, Yi AK, Beaucage SL, Conover J, Krieg AM. CpG motifs present in bacteria DNA rapidly induce lymphocytes to secrete interleukin 6, interleukin 12, and interferon gamma. Proc Natl Acad Sci U S A. 1996;93:2879–2883. doi: 10.1073/pnas.93.7.2879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sparwasser T, Koch ES, Vabulas RM, Heeg K, Lipford GB, Ellwart JW, Wagner H. Bacterial DNA and immunostimulatory CpG oligonucleotides trigger maturation and activation of murine dendritic cells. Eur J Immunol. 1998;28:2045–2054. doi: 10.1002/(SICI)1521-4141(199806)28:06<2045::AID-IMMU2045>3.0.CO;2-8. [DOI] [PubMed] [Google Scholar]
- Sparwasser T, Miethke T, Lipford G, Erdmann A, Hacker H, Heeg K, Wagner H. Macrophages sense pathogens via DNA motifs: induction of tumor necrosis factor-alpha-mediated shock. Eur J Immunol. 1997;27:1671–1679. doi: 10.1002/eji.1830270712. [DOI] [PubMed] [Google Scholar]
- Yamamoto S, Yamamoto T, Kataoka T, Kuramoto E, Yano O, Tokunaga T. Unique palindromic sequences in synthetic oligonucleotides are required to induce IFN and augment IFN-mediated natural killer activity. J Immunol. 1992;148:4072–4076. [PubMed] [Google Scholar]
- Ballas ZK, Rasmussen WL, Krieg AM. Induction of NK activity in murine and human cells by CpG motifs in oligodeoxynucleotides and bacterial DNA. J Immunol. 1996;157:1840–1845. [PubMed] [Google Scholar]
- Bird AP. CpG-rich islands and the function of DNA methylation. Nature. 1986;321:209–213. doi: 10.1038/321209a0. [DOI] [PubMed] [Google Scholar]
- Sved J, Bird A. The expected equilibrium of the CpG dinucleotide in vertebrate genomes under a mutation model. Proc Natl Acad Sci U S A. 1990;87:4692–4696. doi: 10.1073/pnas.87.12.4692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krieg AM, Yi AK, Matson S, Waldschmidt TJ, Bishop GA, Teasdale R, Koretzky GA, Klinman DM. CpG motifs in bacterial DNA trigger direct B-cell activation. Nature. 1995;374:546–549. doi: 10.1038/374546a0. [DOI] [PubMed] [Google Scholar]
- Pisetsky DS. Immune activation by bacterial DNA: a new genetic code. Immunity. 1996;5:303–310. doi: 10.1016/s1074-7613(00)80256-3. [DOI] [PubMed] [Google Scholar]
- Deng GM, Nilsson IM, Verdrengh M, Collins LV, Tarkowski A. Intra-articularly localized bacterial DNA containing CpG motifs induces arthritis. Nat Med. 1999;5:702–705. doi: 10.1038/9554. [DOI] [PubMed] [Google Scholar]
- Feldmann M, Brennan FM, Maini RN. Role of cytokines in rheumatoid arthritis. Annu Rev Immunol. 1996;14:397–440. doi: 10.1146/annurev.immunol.14.1.397. [DOI] [PubMed] [Google Scholar]
- Miossec P. Cytokine abnormalities in inflammatory arthritis. Baillière's Clin Rheumatol. 1992;6:373–392. doi: 10.1016/s0950-3579(05)80180-3. [DOI] [PubMed] [Google Scholar]
- Dayer JM, Fenner H. The role of cytokines and their inhibitors in arthritis. Baillière's Clin Rheumatol. 1992;6:485–516. doi: 10.1016/s0950-3579(05)80186-4. [DOI] [PubMed] [Google Scholar]
- McIntyre KW, Shuster DJ, Gillooly KM, Warrier RR, Connaughton SE, Hall LB, Arp LH, Gately MK, Magram J. Reduced incidence and severity of collagen-induced arthritis in interleukin-12-deficient mice. Eur J Immunol. 1996;26:2933–2938. doi: 10.1002/eji.1830261219. [DOI] [PubMed] [Google Scholar]
- Zhao YX, Ljungdahl A, Olsson T, Tarkowski A. In situ hybridization analysis of synovial and systemic cytokine messenger RNA expression in superantigen-mediated Staphylococcus aureus arthritis. Arthritis Rheum. 1996;39:959–967. doi: 10.1002/art.1780390613. [DOI] [PubMed] [Google Scholar]
- Thomson BM, Mundy GR, Chambers TJ. Tumor necrosis factors alpha and beta induce osteoblastic cells to stimulate osteoclastic bone resorption. J Immunol. 1987;138:775–779. [PubMed] [Google Scholar]
- Mauritz NJ, Holmdahl R, Jonsson R, Van der Meide PH, Scheynius A, Klareskog L. Treatment with gamma-interferon triggers the onset of collagen arthritis in mice. Arthritis Rheum. 1988;31:1297–1304. doi: 10.1002/art.1780311012. [DOI] [PubMed] [Google Scholar]
- Joosten LA, Lubberts E, Helsen MM, van den Berg WB. Dual role of IL-12 in early and late stages of murine collagen type II arthritis. J Immunol. 1997;159:4094–4102. [PubMed] [Google Scholar]
- Hultgren O, Eugster HP, Sedgwick JD, Korner H, Tarkowski A. TNF/lymphotoxin-alpha double-mutant mice resist septic arthritis but display increased mortality in response to Staphylococcus aureus. J Immunol. 1998;161:5937–5942. [PubMed] [Google Scholar]
- Zhao YX, Nilsson IM, Tarkowski A. The dual role of interferon-gamma in experimental Staphylococcus aureus septicaemia versus arthritis. Immunology. 1998;93:80–85. doi: 10.1046/j.1365-2567.1998.00407.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Magram J, Connaughton SE, Warrier RR, Carvajal DM, Wu CY, Ferrante J, Stewart C, Sarmiento U, Faherty DA, Gately MK. IL-12-deficient mice are defective in IFN gamma production and type 1 cytokine responses. Immunity. 1996;4:471–481. doi: 10.1016/s1074-7613(00)80413-6. [DOI] [PubMed] [Google Scholar]
- Bremell T, Abdelnour A, Tarkowski A. Histopathological and serological progression of experimental Staphylococcus aureus arthritis. Infect Immun. 1992;60:2976–2985. doi: 10.1128/iai.60.7.2976-2985.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stacey KJ, Sweet MJ, Hume DA. Macrophages ingest and are activated by bacterial DNA. J Immunol. 1996;157:2116–2122. [PubMed] [Google Scholar]
- Zhao Q, Waldschmidt T, Fisher E, Herrera CJ, Krieg AM. Stage-specific oligonucleotide uptake in murine bone marrow B-cell precursors. Blood. 1994;84:3660–3666. [PubMed] [Google Scholar]
- Geselowitz DA, Neckers LM. Analysis of oligonucleotide binding, internalization, and intracellular trafficking utilizing a novel radiolabeled crosslinker. Antisense Res Dev. 1992;2:17–25. doi: 10.1089/ard.1992.2.17. [DOI] [PubMed] [Google Scholar]
- Bennett RM, Gabor GT, Merritt MM. DNA binding to human leukocytes. Evidence for a receptor-mediated association, internalization, and degradation of DNA. J Clin Invest. 1985;76:2182–2190. doi: 10.1172/JCI112226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hacker H, Mischak H, Miethke T, Liptay S, Schmid R, Sparwasser T, Heeg K, Lipford GB, Wagner H. CpG-DNA-specific activation of antigen-presenting cells requires stress kinase activity and is preceded by non-specific endocytosis and endosomal maturation. EMBO J. 1998;17:6230–6240. doi: 10.1093/emboj/17.21.6230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baeuerle PA, Henkel T. Function and activation of NF-kappa B in the immune system. Annu Rev Immunol. 1994;12:141–179. doi: 10.1146/annurev.iy.12.040194.001041. [DOI] [PubMed] [Google Scholar]
- Tracey KJ, Cerami A. Tumor necrosis factor: a pleiotropic cytokine and therapeutic target. Annu Rev Med. 1994;45:491–503. doi: 10.1146/annurev.med.45.1.491. [DOI] [PubMed] [Google Scholar]
- Pettipher ER, Higgs GA, Henderson B. Interleukin 1 induces leukocyte infiltration and cartilage proteoglycan degradation in the synovial joint. Proc Natl Acad Sci U S A. 1986;83:8749–8753. doi: 10.1073/pnas.83.22.8749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Henderson B, Pettipher ER. Arthritogenic actions of recombinant IL-1 and tumour necrosis factor alpha in the rabbit: evidence for synergistic interactions between cytokines in vivo. Clin Exp Immunol. 1989;75:306–310. [PMC free article] [PubMed] [Google Scholar]
- van den Berg WB, Joosten LA, Helsen M, van de Loo FA. Amelioration of established murine collagen-induced arthritis with anti-IL-1 treatment. Clin Exp Immunol. 1994;95:237–243. doi: 10.1111/j.1365-2249.1994.tb06517.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Germann T, Hess H, Szeliga J, Rude E. Characterization of the adjuvant effect of IL-12 and efficacy of IL-12 inhibitors in type II collagen-induced arthritis. Ann N Y Acad Sci. 1996;795:227–240. doi: 10.1111/j.1749-6632.1996.tb52672.x. [DOI] [PubMed] [Google Scholar]
- Hess H, Gately MK, Rude E, Schmitt E, Szeliga J, Germann T. High doses of interleukin-12 inhibit the development of joint disease in DBA/1 mice immunized with type II collagen in complete Freund's adjuvant. Eur J Immunol. 1996;26:187–191. doi: 10.1002/eji.1830260129. [DOI] [PubMed] [Google Scholar]
- Hultgren OH, Stenson M, Tarkowski A. Role of IL-12 in Staphylococcus aureus-triggered arthritis and sepsis. Arthritis Res. 2001. [DOI] [PMC free article] [PubMed]
- Puddu P, Fantuzzi L, Borghi P, Varano B, Rainaldi G, Guillemard E, Malorni W, Nicaise P, Wolf SF, Belardelli F, Gessani S. IL-12 induces IFN-gamma expression and secretion in mouse peritoneal macrophages. J Immunol. 1997;159:3490–3497. [PubMed] [Google Scholar]
- Germann T, Szeliga J, Hess H, Storkel S, Podlaski FJ, Gately MK, Schmitt E, Rude E. Administration of interleukin 12 in combination with type II collagen induces severe arthritis in DBA/1 mice. Proc Natl Acad Sci U S A. 1995;92:4823–4827. doi: 10.1073/pnas.92.11.4823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wakeham J, Wang J, Magram J, Croitoru K, Harkness R, Dunn P, Zganiacz A, Xing Z. Lack of both types 1 and 2 cytokines, tissue inflammatory responses, and immune protection during pulmonary infection by Mycobacterium bovis bacille Calmette-Guerin in IL-12-deficient mice. J Immunol. 1998;160:6101–6111. [PubMed] [Google Scholar]
- Xing Z, Wang J, Croitoru K, Wakeham J. Protection by CD4 or CD8 T cells against pulmonary Mycobacterium bovis bacillus Calmette-Guerin infection. Infect Immun. 1998;66:5537–5542. doi: 10.1128/iai.66.11.5537-5542.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deng GM, Verdgrengh M, Liu ZQ, Tarkowski A. The major role of macrophages and their product: tumour necrosis factor α in the induction of arthritis triggered by bacterial DNA containing CpG motifs. Arthritis Rheum. 2000;43:2283–2289. doi: 10.1002/1529-0131(200010)43:10<2283::AID-ANR16>3.0.CO;2-9. [DOI] [PubMed] [Google Scholar]
- Shirai T, Shimizu N, Horiguchi S, Ito H. Rat TNF alpha. Agric Biol Chem. 1989;53:1733–1736. [Google Scholar]
- Schoenhaut DS, Chua AO, Wolitzky AG, Quinn PM, Dwye CM, McComas W, et al. Cloning and expression of murine IL-12. J Immunol. 1992;148:3433–3440. [PubMed] [Google Scholar]
- Dijkema R, van der Meide PH, Dubbeld M, Caspers M, Lubben J, Schellekens H. Cloning, expression and purification of rat IFN-γ. Methods Enzymol. 1986;119:453–464. doi: 10.1016/0076-6879(86)19065-3. [DOI] [PubMed] [Google Scholar]