

Abstract
To reduce culture artifacts by conventional repeated passaging and long-term culture in vitro, the isolation of synovial fibroblasts (SFB) was attempted from rheumatoid arthritis (RA) synovial membranes by trypsin/collagenase digest, short-term in vitro adherence (7 days), and negative isolation using magnetobead-coupled anti-CD14 monoclonal antibodies. This method yielded highly enriched SFB (85% prolyl-4-hydroxylase+/74% Thy-1/CD90+ cells; <2% contaminating macrophages; <1% leukocytes/endothelial cells) that, in comparison with conventional fourth-passage RA-SFB, showed a markedly different phenotype and significantly lower proliferation rates upon stimulation with platelet-derived growth factor and IL-1β. This isolation method is simple and reliable, and may yield cells with features closer to the in vivo configuration of RA-SFB by avoiding extended in vitro culture.
Keywords: isolation, phenotype/function, primary culture, rheumatoid arthritis, synovial fibroblasts
Synopsis
Introduction:
Activated SFB in the invasive pannus tissue appear to play a major role in the crippling destruction of cartilage and bone in the joints of patients with RA [1,2,3]. Several studies indicate that RA-SFB are morphologically altered [1,4], grow in an anchorage-independent fashion [5], and strongly express markers of activation, including major histocompatibility complex (MHC)-II and other surface molecules, proto-oncogenes, or matrix degrading enzymes (reviewed in [2]). Finally, activated RA-SFB elicit erosive arthritis upon intra-articular injection or engraftment into severe combined immunodeficiency mice [6,7].
Aims:
To investigate ex vivo the pathophysiological properties of activated FB from the RA synovial membrane (SM), it is desirable to obtain freshly isolated SFB with phenotypic features as close as possible to the configuration observed in vivo. Because the generation of cell lines requires a number of passages in vitro to eliminate contaminating cells (especially macrophages), the risk of substantial in vitro alteration/growth selection exists.
We attempted to isolate FB from primary cultures of synovial cells, using a negative separation technique to eliminate macrophages, to minimize these artifacts.
Methods:
Synovial tissue was obtained from a total of 16 patients fulfilling the American Rheumatism Association criteria for RA [8] and 21 patients with osteoarthritis (OA) under approval of the local Ethics Committees. The tissue was placed in cell culture medium at ambient temperature and subjected to tissue digestion within 2 h. Synovectomy samples of RA and OA SM were finely minced, digested for 30 min at 37°C in phosphate-buffered saline (PBS) containing 0.1% trypsin (Sigma, Deisenhofen, Germany), and thereafter digested in 0.1% collagenase P (Boehringer Mannheim, Mannheim, Germany) in Dulbecco's modified Eagle medium (DMEM)/10% fetal calf serum (FCS) for 2 h at 37°C, 5% CO2. The cell suspension was then filtered and the cells collected by centrifugation. Cells were kept in primary culture for 7 days (DMEM/10% FCS, 25 mM HEPES, 100 U/ml penicillin, 100 μg/ml streptomycin, and 2.5 μg/ml amphotericin B [Gibco BRL, Eggenstein, Germany], including removal of non-adherent cells on days 1, 3, 5, and 7) and subsequently used for SFB isolation. The samples were randomly tested to exclude Mycoplasma contamination. For negative isolation of SFB from primary culture, adherent synovial cells were detached by short-term trypsinization for 2 min (0.25% trypsin/0.2% EDTA; Gibco) and 107/ml synovial cells were incubated with 4 × 107/ml Dynabeads® M-450 CD14 (clone RMO52; Dynal, Hamburg, Germany) in PBS/2% FCS for 1 h at 4°C. Nine milliliters of PBS/2% FCS were then added and the conjugated cells collected using the Dynal magnetic particle concentrator®. The compositions of magnetobead-conjugated cells and unconjugated cells were analyzed by flow cytometry. Phenotype analysis of the expression of FB markers, as well as that of SFB features previously reported at a tissue level, was conducted by flow cytometry in RA-SFB, either negatively isolated from primary culture or obtained from conventional fourth passage. The findings were compared with those of normal skin-FB (lineage control) and OA-SFB (disease control). The proliferation of RA-SFB, either negatively isolated from primary culture or obtained from conventional fourth passage, was assayed by [3H]-thymidine incorporation.
Results:
The primary culture of RA synovial cells resulting from trypsin/collagenase digestion of the RASM contained large, spindle-shaped Thy-1+ SFB (CD90+; Fig. 1C) (monoclonal antibody [mAb] AS02; Dianova, Hamburg, Germany) and small, round CD14+ cells, most probably macrophages (Fig. 1D) (mAb Tyk4; Dako, Hamburg, Germany), as detected by immunohistochemical staining [6,9]. Endothelial cells were absent, as confirmed by lack of staining for von Willebrand Factor (Fig. 1F) (mAb 4F9; Immunotech, Hamburg, Germany) and CD144 (Fig. 1G) (mAb Cadherin 5; Immunotech), which clearly identified human umbilical vein endothelial cells (HUVEC) (data not shown). The FB nature of the spindle-shaped cells was confirmed by intracellular staining for procollagen I and III (Fig. 1E,H) (rabbit antibodies MP I and MP III; Prof. Schuppan, Berlin, Germany). An average of 62% of the cells stained with the anti-Thy-1 mAb AS02 (n = 4 RA patients; Table 1a) in flow cytometry (FACS) [10]; the average percentage of CD14+ cells was approximately 15% (n = 4; Table 1a). There were <1% T cells (CD3+) (mAb UCHT-1; ATCC, Manassas, VA, USA), B cells (CD19+/20+) (mAbs HD 37 and B-Ly 1; Dako), plasma cells (CD38+) (mAb AT 13/5; Dako), natural killer (NK) cells (CD56+) (mAb NKH/1; Immunotech), dendritic cells (CD83+) (mAb HB 15a; Immunotech), endothelial cells (CD144+), or PMN (CD15+) (mAb80H5; Immunotech), indicating that non-adherent cells had been efficiently removed during primary culture. The total yield of cells following 7 days of primary culture averaged (5.2 ± 1.1) × 107 cells (mean ± SEM; n = 7).
Figure 1.
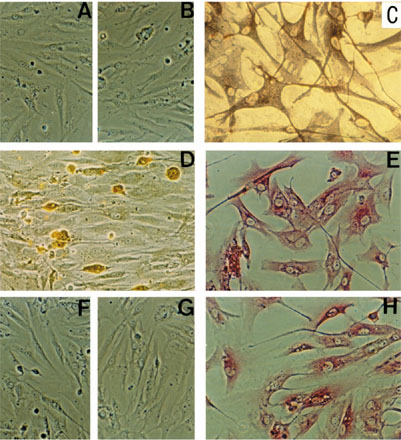
Immunohistochemical staining of primary-culture RA synovial cells in chamber slides: (A), (B), (D)-(H) phase contrast, and (C) lightfield. While (A) control isotype-matched mAbs (peroxidase) or (B) rabbit serum (APAAP) showed no positive reaction, (C) the anti-Thy-1 mAb AS02 stained large, flat, spindle-shaped or stellate-shaped cells (ie cells with FB morphology; alkaline phosphatase, purple staining) and (D) the anti-CD14 mAb RMO52 identified small, round cells, conceivably macrophages (peroxidase, brown staining). The RA-SFB clearly expressed (E) procollagen I and (H) procollagen III (APAAP, red staining). The primary culture contained no endothelial cells, as documented by the lack of staining for (F) von Willebrand factor (APAAP) and (G) CD144 (APAAP). Original magnification: (A), (B), (F), and (G), 184 ×; (C)-(E) and (H) 368 ×.
Table 1.
Composition of rheumatoid arthritis (RA)-synovial fibroblasts (SFB), osteoarthritis (OA)-SFB, and normal skin-FB, as determined by flow cytometry for various numbers of patients/donors
| Percent of positive cells | Mean fluorescence intensity | |||
| Composition | Mean ± SEM | Range | Mean ± SEM | Range |
| (a) Primary-culture RA synovial cells | ||||
| Thy-1 (CD90) (n = 4) | 62.0 ± 14.6 | 24–93 | 152 ± 49 | 46–283 |
| CD14 (n = 4) | 14.8 ± 6.0 | 2–25 | 230 ± 87 | 64–445 |
| (b)RA-SFB (negative isolation from primary culture; negative fraction) | ||||
| Thy-1 (n = 9) | 73.8 ± 9.6 | 21–99 | 109 ± 20 | 53–222 |
| Prolyl-4-hydroxylase (n = 9) | 84.8 ± 2.9 | 66–95 | 164 ± 45 | 20–391 |
| CD14 (n = 9) | 0.7 ± 0.2 | 0.1–1.7 | 62 ± 12 | 30–126 |
| CD68 (mAb PG-M1) (n = 5) | 1.7 ± 0.6 | 0.4–3.2 | 120 ± 78 | 10–430 |
| (c)RA-SFB (fourth-passage; isolated cells) | ||||
| Thy-1 (n = 3) | 86.6 ± 4.6 | 79–95 | 41 ± 7 | 27–53 |
| Prolyl-4-hydroxylase (n = 3) | 98.7 ± 0.8 | 97–100 | 164 ± 62 | 82–285 |
| CD14 (n = 3) | 0.6 ± 0.2 | 0.2–1.0 | 50 ± 30 | 20–110 |
| (d)RA-SFB (fourth-passage; conventionally passaged cells) | ||||
| Thy-1 (n = 6) | 85.9 ± 3.9 | 70–95 | 95 ± 15 | 43–167 |
| Prolyl-4-hydroxylase (n = 6) | 98.3 ± 0.6 | 95–99 | 189 ± 75 | 20–518 |
| CD14 (n = 6) | 1.6 ± 0.6 | 0.2–4.9 | 83 ± 32 | 22–255 |
| CD68 (mAb PG-M1) (n = 3) | 0.5 ± 0.0 | 0.5–0.6 | 29 ± 6 | 19–40 |
| (e)OA-SFB (negative isolation from primary culture; negative fraction) | ||||
| Thy-1 (n = 9) | 62.1 ± 9.0 | 28–92 | 120 ± 36 | 53–404 |
| Prolyl-4-hydroxylase (n = 7) | 92.2 ± 2.8 | 77–98 | 212 ± 44 | 30–382 |
| CD14 (n = 9) | 1.3 ± 0.3 | 0.7–3.0 | 268 ± 97 | 14–754 |
| CD68 (mAb PG-M1) (n = 7) | 1.3 ± 0.2 | 0.6–2.1 | 58 ± 18 | 14–140 |
| (f)OA-SFB (fourth-passage; conventionally passaged cells) | ||||
| Thy-1 (n = 3) | 80.9 ± 5.0 | 72–93 | 56 ± 6 | 46–70 |
| Prolyl-4-hydroxylase (n = 3) | 98.8 ± 0.3 | 98–99 | 250 ± 27 | 205–313 |
| CD14 (n = 3) | 1.0 ± 0.5 | 0.2–2.2 | 24 ± 1 | 22–26 |
| (g) Primary culture or first-passage normal skin-FB | ||||
| Thy-1 (n = 8) | 91.4 ± 3.0 | 78–99 | 156 ± 36 | 34–349 |
| Prolyl-4-hydroxylase (n = 8) | 70.7 ± 15.2 | 2–99 | 101 ± 31 | 10–269 |
| CD14 (n = 8) | 1.5 ± 0.4 | 0.3–4.0 | 70 ± 14 | 22–130 |
| CD68 (mAb PG-M1) (n = 5) | 0.5 ± 0.3 | 0.1–1.4 | 82 ± 25 | 12–164 |
mAb, Monoclonal antibody.
Negative isolation of SFB from primary culture resulted in RA-SFB that were Thy-1+ (on average, approximately 74%; n = 9; Fig. 2A and Table 1b) and, more importantly, prolyl-4-hydroxylase+ (on average, approximately 85%; n = 9; Table 1b) (mAb3-2B12; Dianova), as shown by FACS analysis and confirmed by immunohistochemistry in chamber slides. There were very few contaminating non-specific esterase+ (three RA patients; Fig. 2C), CD14+, CD68+, and/or CD11b+ macrophages (<2%; Fig. 2B,D and Table 1b), as well as <1% T cells (CD3+), B cells (CD19+/20+), plasma cells (CD38+), NK cells (CD56+), dendritic cells (CD83+), PMN (CD15), or endothelial cells (CD144+; von Willebrand factor-positive). The positive fraction, in turn, contained virtually no SFB, indicating minimal cell loss, and thereby also excluding major subset selection. The average yield of RA-SFB negatively isolated from primary culture was (2.8 ± 0.9) × 107 cells (mean ± SEM; n = 7). Phenotype analyses yielded two main results. The first, regarding expression of SFB features previously reported at a tissue level, was that, strikingly, approximately 66% of the cells expressed MHC-II molecules (normal skin-FB, 2%; OA-SFB, 17%). A low or moderate and variable percentage of RA-SFB, but also normal skin-FB and OA-SFB, expressed vascular cell adhesion molecule-1 (VCAM-1) (using two different anti-VCAM-1 mAbs) without statistically significant differences between the three different FB preparations. RA-SFB showed a higher (although non-significant) mean fluorescence intensity (MFI) for the cytoskeletal protein vimentin than normal skin-FB. Approximately 45% and 50% of the cells, respectively, expressed procollagen I and procollagen III (similar to normal skin-FB); however, the MFI for procollagen III was significantly higher in RA-SFB. Approximately 57% of RA-SFB expressed c-Fos. Neither the percentage nor the MFI were significantly different from those of normal skin-FB (approximately 87%) or OA-SFB (approximately 54%). In general, the differences between RA-SFB and normal skin-FB were not specific to RA, since they were also observed in the comparison between the disease control OA-SFB and normal skin-FB. The second outcome of phenotype analysis was that the percentages of RA-SFB positive for MHC-II as well as the MFI for VCAM-1 and c-Jun were significantly decreased in conventional fourth passage compared with isolated primary RA-SFB. In contrast, the percentages of cells positive for MHC-I, CD13, prolyl-4-hydroxylase, vimentin, procollagen I and III, c-Fos, and Jun-D were significantly increased in conventional fourth passage.
Figure 2.
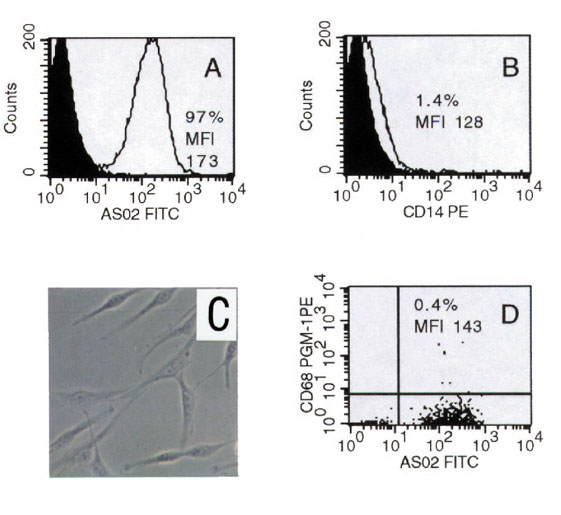
FACS/histochemical analysis of the negative fraction following isolation of RA-SFB from primary culture using Dynabeads® M-450 CD14. The AS02/Thy-1+ RA-SFB (A) were almost free of contaminating CD14+(B) (mAb Tyk4) or CD68+ macrophages (D) (mAb PG-M1). The RA-SFB were accordingly negative for the macrophage marker non-specific esterase (histochemistry (C)). Original magnification: (C) 184 ×. PE, phycoerythrine; FITC, fluoresceine isothiocyanate.
Proliferation assays showed that, at rest, the proliferation rates of isolated first-passage RA-SFB did not significantly differ from those of the corresponding cells in conventional fourth passage. Upon stimulation with platelet-derived growth factor (PDGF) (2.5 and 5 U/ml) and IL-1β (150 U/ml), however, the mean proliferation rates of conventional fourth-passage RA-SFB were significantly higher than those observed in first-passage cells (3.9-fold and 4.2-fold, respectively, in the case of 2.5 and 5 U/ml PDGF; 2.1-fold in the case of 150 U/ml IL-1β).
Discussion:
The advantages of negative isolation are as follows: first, there was minimal contamination with other cells, especially macrophages (<2%); second, negative isolation avoided direct contact of SFB with mAbs and/or magnetobeads, thereby limiting possible functional alterations of the cells; and third, the FB marker mAb AS02 (anti-Thy-1; used in the parallel attempt to positively isolate SFB) identified 91% of primary-culture/first-passage normal skin-FB but only 74% of isolated primary RA-SFB, probably due the variable density of Thy-1 molecules on the cell surface (Table 1). Thus, Thy-1-independent isolation appears to avoid selection of Thy-1+ SFB subpopulations, a critical point since Thy-1+ and Thy-1- FB diverge in several characteristics potentially relevant to RA.
Potential disadvantages of the negative isolation method include the limited number of cells obtained (approximately 2.8 × 107), due to the lack of in vitro expansion, and the necessity for reliable access to freshly obtained synovial specimens. However, the applicability of this procedure not only to joint replacement samples, but also to arthroscopic synovectomy samples, augments the potential sources of fresh synovial tissue. The use of enzymes for tissue digestion may also represent at least a transient stress for the cells. On the other hand, this method may avoid selection of premitotic FB, a problem usually linked to tissue-outgrowth techniques [11].
The advantages of limited-passage isolation include the following. The present technique avoids a high number of passages in vitro; indeed, even a low number of passages increased the proliferation rates in response to cytokine stimulation and altered the cellular phenotype of RA-SFB (as exemplified by a strong decrease of the percentage of MHC-II+ cells, on one hand, and a striking increase of the percentage of procollagen I and III+, as well as c-Fos and Jun-D+ cells, on the other). Caution should thus be applied to the interpretation of data from passaged cells, especially in the case of proto-oncogenes. A second advantage is that this isolation procedure reduces the time of exposure (7 days) to surviving macrophages (unlike the weeks of exposure upon conventional isolation), thus possibly minimizing long-term in vitro changes due to monokine secretion or cell–cell contact between SFB and synovial macrophages. Finally, a high percentage of RA-SFB from limited-passage isolation expressed MHC-II molecules (in analogy to findings in synovial tissue) without in vitro stimulation with cytokines, which in conventionally isolated cells requires stimulation with interferon-gamma. Therefore, the present in vitro system may be particularly useful in studies on the inter-relationship of SFB with T cells or macrophages, in which the influence of exogenous mediators may complicate the design and/or interpretation of the experiments.
Negative isolation of cells from synovial primary culture with magnetobead-conjugated anti-CD14 mAbs thus yielded highly enriched RA-SFB. This technique, by avoiding repeated passaging and numerous cumulative population doublings, may minimize the risk of in vitro alteration/growth selection; in RA-SFB, this includes increased proliferation rates and changes of several phenotypic features.
Introduction
Activated SFB play a major role in the crippling destruction of cartilage and bone in the joints of patients with RA [1,2,12,13,14,15], either autonomously [1] or in concert with T cells and macrophages [16,17,18,19]. RA-SFB are a major part of the invasive pannus, a vascular and fibrous granulation tissue arising from the joint recessus and extending onto the surface of cartilage [20]. Several studies indicate that RA-SFB are morphologically and functionally altered [4,5,12], and strongly express markers of activation, including surface molecules (eg MHC-II, CD13, or VCAM-1 and other integrins) [1,2,13,21,22] and proto-oncogenes [23,24,25].
Activation of SFB in vitro generates several functional responses that may considerably contribute to joint pathology in RA; that is, the production of matrix components [11,26], soluble mediators [17,27], or matrix degrading enzymes [28,29,30]. Because of the capability to act as antigen-presenting cells, activated SFB can also potentially fuel local antigen-driven processes relevant to the immunopathogenesis of RA [19,31]. Finally, stimulated RA-SFB elicit erosive arthritis upon intra-articular injection or engraftment into severe combined immunodeficiency mice [6,7].
To investigate ex vivo the pathophysiological properties of activated FB from the RASM, it is desirable to obtain freshly isolated SFB with phenotypic features as close as possible to the configuration observed in vivo. Because the generation of cell lines requires a number of passages in vitro to eliminate contaminating cells (especially macrophages), the risk of substantial in vitro growth selection exists [32]. Indeed, increasing the number of passages modifies the surface phenotype of synoviocytes [19] and increases the frequency of genotype changes [33]. We attempted to isolate FB from primary cultures of synovial cells to minimize these artifacts, with careful characterization of each preparation step and comparison of the phenotypic and functional features of primary-culture SFB with those of repeated-passage cells. Both positive and negative isolation approaches were pursued.
Positive identification/isolation of SFB was attempted using the mAb AS02, an antibody initially described as FB-specific [9] and subsequently shown to recognize the Thy-1 molecule (CD90, highly glycosylated membrane-bound protein; molecular weight, 30–35 kDa; core protein, 17 kDa; anchored via glycosylphosphatidyl-inositol [34]).
Negative isolation was performed using magnetobead-conjugated anti-CD14 mAbs. The choice of CD14 was based on the consideration that this is regarded as a reliable surface marker for RA synovial macrophages [23,35]; CD14 is also the macrophage marker most clearly correlated with disease activity and radiologic progression of joint destruction in RA [36]. The use of the most valid alternative (ie anti-CD68 mAbs) is limited by predominant expression of the CD68 antigen in the lysosomal compartment (requiring permeabilization of all cells for negative selection), and by the fact that CD68 epitopes recognized by certain mAbs are not specific for macrophages, but are also expressed by SFB [37,38]. The possibility of using anti-CD11b mAbs was also discarded, because CD11b is not specific for monocytes/macrophages and is also found on granulocytes and NK cells. CD11b expression on subpopulations of RA synovial macrophages may also be variable [39], possibly due to the local influence of cytokine stimulation [40,41]. Therefore, although the expression of CD14 on RA synovial macrophages may also be influenced by cytokines [36,42,43], the anti-CD14 mAb appeared the most appropriate choice for the purpose of this study.
Negative isolation from primary culture with magnetobead-conjugated anti-CD14 mAbs yielded highly enriched RA-SFB, positive for the known FB marker prolyl-4-hydroxylase (85%) and the recently suggested FB marker Thy-1 (74% [9]), with <2% macrophage contamination and <1% T cells, B cells, plasma cells, NK cells, dendritic cells, endothelial cells, or PMN. These cells bore surface and intracellular characteristics typical of the FB lineage when compared with primary-culture normal skin-FB (lineage control) or OA-SFB (disease control).
Materials and methods
Patients
Synovial tissue was obtained during open joint replacement surgery or arthroscopic synovectomy from a total of 16 patients with the clinical diagnosis of RA (13 knee joints, one hip joint, one wrist, one metacarpo-phalangeal joint) as well as 21 patients with the clinical diagnosis of OA (all knee joints; Table 2) from the Department of Orthopedics, University of Leipzig, Germany, the Clinic of Orthopedics, Bad Düben, Germany, and the Clinic of Orthopedics, Eisenberg, Germany, as well as the Department of Orthopedic Surgery, University of Michigan, Ann Arbor, MI, USA. All RA patients fulfilled the American Rheumatism Association criteria for RA [8]. The study was approved by the Ethics Committees of the University of Leipzig and the University of Jena, Germany, and the University of Michigan, MI, USA. One portion of each sample was immediately frozen in isopentane (Merck, Darmstadt, Germany), cooled in liquid nitrogen and stored at –70°C for immunohistochemistry. The remaining tissue was placed in cell culture medium at ambient temperature and subjected to tissue digestion within 2 h.
Table 2.
Clinical characteristics of the patients at the time of synovectomy
| Disease | ||||||||
| Patients | Gender | Age | duration | RF | ESR | CRP | Number of ARA | Concomitant |
| (total) | (m/f) | (years) | (years) | (+/-) | (mm/h) | (mg/l) | criteria (RA) | medication |
| Rheumatoid arthritis | ||||||||
| 16 | 7/9 | 61.4 ± 2.2 | 12.0 ± 2.4 | 11/4 (n.d. = 1) | 33.0 ± 4.6 | 41.8 ± 7.7 | 5.2 ± 0.2 | MTX (n = 10) |
| Prednisolone (n = 13) | ||||||||
| Sulfasalazine (n = 4) | ||||||||
| Gold salts (n = 1) | ||||||||
| NSAIDs (n = 12) | ||||||||
| Osteoarthritis | ||||||||
| 21 | 5/16 | 70.4 ± 2.6 | 9.8 ± 3.0 | 0/8 (n.d. = 13) | 15.6 ± 1.9 | 4.6 ± 1.4 | 0.3 ± 0.1 | MTX (n = 1) |
| Prednisolone (n = 1) | ||||||||
| NSAIDs (n = 11) | ||||||||
| None (n = 10) | ||||||||
For the parameters age, disease duration, erythrocyte sedimentation rate (ESR), C-reactive protein (CRP) (normal range, < 5 mg/l), and number of American Rheumatism Association (ARA) criteria (rheumatoid arthritis [RA]), means ± SEM are given; for the remaining parameters, numbers are provided. m, Male; f, female; RF, rheumatoid factor; +/-, positive/negative; n.d. = not determined; MTX, Methotrexate; NSAIDs, non-steroidal anti-inflammatory drugs.
Histochemical detection of non-specific esterase
Non-specific esterase activity was analyzed in methanol-fixed (10 min at room temperature) first-passage RA-SFB and OA-SFB on chamber slides, and in air-dried, unfixed rat kidney sections or rat peritoneal macrophages, by incubation for 1 h at 37°C with a reaction medium containing 0.02% 5-bromo-4-chloro-3-indoxylacetate (initially dissolved in dimethylformamide), 1 mM potassiumferricyanide, and 1 mM potassiumferrocyanide (final concentrations) (all from Sigma) in 2 mM Tris7#150;HCl buffer (pH 6.8). The positive controls (rat kidney or peritoneal macrophages) were stained in all experiments, while negative control cell preparations/kidney sections incubated with substrate-free medium did not show any positive reaction (respective data not shown). The cells/sections were then washed in PBS (see later) and mounted in glycerin gelatin.
Immunohistochemistry
Immunohistochemical analysis was performed on cryostat sections of RASM (fixed with acetone for 10 min at 4°C) or chamber slides (fixed with 100% methanol for 10 min at room temperature; Nunc, Wiesbaden, Germany) using the antibodies presented in Table 3. The mouse mAbs, diluted in Tris–buffered saline (TBS)/1% bovine serum albumin for per-oxidase/alkaline phosphatase detection, were added for 30 min in a humid chamber at room temperature following pre-incubation of the sections/chambers with 20% normal human serum in TBS for 20 min (pH 7.4). A peroxidase-coupled rabbit anti-mouse antibody (Dako) and a peroxi-dase-coupled swine anti-rabbit antibody (Dako; both 1:30 in TBS/20% human serum) were added for 30 min each. The peroxidase was revealed using diaminobenzidine (5 min, 0.5 mg diaminobenzidine in 1 ml PBS [pH 7.4] containing 30 μl H2O2). Blocking of endogenous peroxidase (0.03% H2O2) before immunohistochemical analysis, as performed in some experiments, did not alter the staining results.
Table 3.
Antibodies used in the present study
| Antibody (technique) | Antigen recognized | Cellular localization | Main cellular expression |
| MOPC 21 (F/I) | - | [IgG1 isotype control] | |
| UPC-10 (F/I) | - | [IgG2a isotype control] | |
| Rabbit Ig/rabbit serum (F/I) | - | [rabbit Ig control] | |
| Monoclonal | |||
| AS02 (F/I) | Thy-1 (CD 90) | Membrane | Fibroblasts, endothelial cells, thymocytes |
| W6/32 (F) | MHC-I (A, B, C) | Membrane | Ubiquitous |
| CII 1.47 (F) | MHC-II (Pan) | Membrane | Antigen presenting cells, fibroblasts |
| SJD1 (F) | CD13 | Membrane | Macrophages, fibroblasts, PMN, T cells |
| 3-2B12 (F/I) | Prolyl-4-hydroxylase | Cytoplasm | Fibroblasts, endothelial cells |
| V9 (F) | Vimentin | Cytoplasm | Fibroblasts, endothelial cells |
| 4F9 (F/I) | von Willebrand factor | Cytoplasm | Endothelial cells |
| UCHT-1 (F) | CD3 | Membrane | T cells |
| TM316 (F) | CD11b | Membrane | Monocytes/macrophages, PMN, NK cells |
| MoS39 (F/I) | CD14 | Membrane | Monocytes/macrophages |
| Tyk 4 (F/I) | CD14 | Membrane | Monocytes/macrophages |
| 80H5 (F) | CD15 | Membrane | PMN, monocytes |
| HD 37 (F) | CD19 | Membrane | B cells |
| B-Ly 1 (F) | CD20 | Membrane | B cells |
| Cadherin 5(F/I) | CD144 | Membrane | Endothelial cells |
| AT 13/5 (F) | CD38 | Membrane | Plasma cells, activated T cells |
| NKH/1 (F) | CD56 | Membrane | NK cells |
| PG-M1 (F/I) | CD68 | Membrane/cytoplasm | Macrophages* |
| HB 15a (F) | CD83 | Membrane | Dendritic cells, activated lymphocytes |
| BBIG-V1(F/I) | VCAM-1 (CD 106) | Membrane | Fibroblasts, endothelial cells |
| 1.4C3 (F) | VCAM-1 (CD 106) | Membrane | Fibroblasts, endothelial cells |
| Polyclonal | |||
| MP I (F/I) | Procollagen I | Cytoplasm | Fibroblasts |
| MP III (F/I) | Procollagen III | Cytoplasm | Fibroblasts |
| Anti-c-Fos(F) | c-Fos | Nucleus/cytoplasm | Ubiquitous |
| Anti-c-Jun(F) | c-Jun | Nucleus/cytoplasm | Ubiquitous |
*The only anti-CD68 monoclonal antibody reported to be specific for macrophages (data not shown) [38]. F, Antibody used for flow cytometry (FACS) analysis only; F/I, antibody used for FACS and immunohistochemistry; MHC, major histocompatibility complex; VCAM-1, vascular cell adhesion molecule-1; NK, natural killer.
Alternatively, either the indirect alkaline phosphatase or the alkaline phosphatase anti-alkaline phosphatase (APAAP) techniques were used for detection of the monoclonal or polyclonal primary antibodies [35]. An alkaline phosphatase-coupled goat-anti-mouse antibody (Dako) was applied for 30 min (1:30 in TBS/20% human serum; pH 7.4) in the case of the indirect alkaline phosphatase, and the alkaline phosphatase was revealed in substrate buffer (TBS; pH 9.5) containing 0.16 mg/ml 5-chromo-4-chloro-3-indoxylphosphate and 0.15 mg/ml nitroblue tetra-zolium chloride (both from Boehringer Mannheim). In the case of the APAAP, a rabbit anti-mouse antibody (Dako) was applied for 45 min (1:100 in TBS/20% human serum; pH 7.4). The sections/chamber slides were washed, incubated for 45 min with APAAP complex (1:80 in TBS/1% bovine serum albumin; Dianova), and washed again with TBS. To amplify the staining, incubation with the rabbit anti-mouse mAb and the APAAP was repeated once. The alkaline phosphatase was revealed in Tris–HCl buffer (pH 8.6) containing 0.3 mg/ml Naphthol AS-MX phosphate (first dissolved at 15 mg/ml in dimethylformamide; both from Sigma), 1 mg/ml Fast Red TR salt or Fast Blue BB salt, and 0.24 mg/ml levamisole (all from Sigma). The solution was filtered and added to the sections/chamber slides for 30 min in a humid chamber. For the APAAP technique, polyclonal primary antibodies (against procollagen I and III) were incubated with a secondary mouse anti-rabbit antibody (Dako) before using the secondary detection systems.
Double-staining experiments on cryostat sections of the RA SM were performed by combining the peroxidase technique with the APAAP technique. The sections were blocked with 20% normal human serum in TBS for 30 min following completion of peroxidase staining. Isotype-matched mouse mAbs, rabbit sera, or purified rabbit immunoglobulins at identical concentrations to the primary antibodies were used as controls for all immunohistochemical analyses (Table 3), which gave no positive results. Cross-reactivity of the peroxidase and the alkaline phosphatase detection (theoretically possible on the basis of employing mouse primary mAbs and anti-mouse secondary antibodies in both systems) was never observed when using control Ig or when omitting one or both primary mAbs, presumably because the peroxidase reaction with diaminobenzidine eliminates all immunoreactive epitopes from this step.
For comparative immunohistochemical and FACS analyses, the percentage of cells stained for the individual markers following immunohistochemistry on chamber slides was determined (total of 100 cells analyzed) and compared with the percentage of positive cells in FACS analysis (see later).
Tissue digestion and cell culture
Samples of RA or OA SM were finely minced with scissors (tissue pieces of approximately 30 mm3) and digested for 30 min at 37°C in 20 ml PBS containing 0.1% trypsin (Sigma). After removal of trypsin/PBS, the samples were digested in 20 ml 0.1% collagenase P (Boehringer Mannheim) in DMEM/10% FCS for 2 h at 37°C, 5% CO2. The cell suspension was then filtered through a sterile sieve (Sigma), the cells collected by centrifugation, washed twice with serum-free DMEM medium, and subsequently cultured for 7 days in DMEM/10% FCS, 25 mM HEPES, 100 U/ml penicillin, 100 μg/ml streptomycin, and 2.5 μg/ml amphotericin B (all from Gibco BRL, Karlsruhe, Germany). Primary-culture skin-FB from normal donors (kindly provided by Dr Sauer, Leipzig, Germany) were prepared by first incubating skin samples with 0.5 U/ml Dispase II (Boehringer Mannheim) overnight at 4°C and, following removal of the epidermis, by digesting with 0.25% collagenase P (Boehringer Mannheim) in DMEM/1% FCS at 37°C and 5% CO2 for 4 h. The resulting cell suspension was then cultured for 14 days as already described. Repeated-passage HUVEC (kindly provided by Dr C. Syring, Institute of Clinical Immunology and Transfusion Medicine, University of Leipzig) were cultured in DMEM/10% FCS/25 mM HEPES containing 10 U/ml heparin, 10 ng/ml basic fibroblast growth factor (Sigma), and 100 μg/ml recombinant endothelial growth factor (Sigma). The medium was changed every 2-3 days in all cases. Adherent RA-SFB or OA-SFB were subjected to very short trypsinization (2 min, 0.25% trypsin/0.2% EDTA; Gibco), removed from the culture dish by mechanical dislocation, washed in PBS/2% FCS, and used for negative isolation. The samples were randomly tested to exclude Mycoplasma contamination.
Negative isolation from primary culture
Trypsinized and washed RA and OA synovial cells from primary culture (107/ml) were incubated with 4 × 107/ml Dynabeads® M-450 CD14 (clone RMO52; Dynal) in PBS/2% FCS for 1 h at 4°C under bidirectional rotation. Nine milliliters of PBS/2% FCS were then added and the conjugated cells collected using the Dynal magnetic particle concentrator®. Magneto-bead-conjugated cells and unconjugated cells were collected and washed twice in PBS/2% FCS; cell composition and phenotype were analyzed by flow cytometry using the antibodies presented in Table 3. For the purpose of comparison with fourth-passage cells obtained by conventional isolation (ie repeated passaging; see later), negatively isolated RA-SFB and OA-SFB were then passaged four times by culture in complete DMEM/10% FCS (see earlier) with a 1:3 split of confluent cells in each passage.
Conventional isolation of RA-SFB and OA-SFB by repeated passage
Synovial cells were obtained by trypsin/collagenase digestion of the RASM as already described. The cells were subsequently cultured for four passages in DMEM/10% FCS containing the aforementioned additives by splitting confluent cells in each passage at a ratio of 1:3.
Flow cytometry
The antibodies presented in Table 3 were used for FACS analyses of primary-culture RA and OA synovial cells (following 7 days of culture), isolated RA-SFB and OA-SFB (immediately following isolation or at fourth-passage), conventional fourth-passage RA-SFB and OA-SFB, normal skin-FB (primary-culture or fourth-passage), or repeated-passage endothelial cells (HUVEC). Primary antibodies were used at concentrations of 10–20 μg/ml. Standard single and double-staining procedures for surface molecules were performed as previously described [10]. The specificity of staining was confirmed using isotype-matched control mAbs, rabbit serum, or rabbit Ig at identical concentrations (Table 3). The FACS analysis in RA-SFB and OA-SFB was not always performed with the complete set of antibodies due to the initial establishment of the isolation procedure.
Trypsinized cells were washed twice with serum-free medium and fixed for 10 min at 4°C in 4% paraformalde-hyde (Fluka, Deisenhofen, Germany) for detection of antigens in cytoplasm and/or nucleus (eg prolyl-4-hydroxylase; Table 3). After washing twice in PBS/2% FCS, the pellet was resuspended in permeabilization buffer (PBS/1% FCS, 0.01% NaN3, and 0.5% saponine; Serva, Heidelberg, Germany) and incubated for 10 min at room temperature. Unlabeled primary mAbs were added at saturating concentrations and detected with a secondary phycoerythrine-labeled goat anti-mouse antibody (Dako), both for 45 min at 4°C in permeabilization buffer. Polyclonal antibodies were incubated with a secondary mouse anti-rabbit antibody (Dako) before using the phycoerythrine-labeled goat anti-mouse antibody. After washing twice in permeabilization buffer, the cells were incubated for 10 min in PBS/5% FCS and 0.1% NaN3 without saponine. In the special case of the CD68 epitope recognized by the mAb PG-M1, present on the cell surface and in the cytoplasm, FACS analysis was performed in both non-permeabilized and permeabilized cells. For double-staining analyses, fluoresceine isothiocyanate (FITC)-labeled Anti-Thy-1 mAb AS02 (20 μg/ml in PBS/2% FCS) or a matched concentration of the FITC-labeled IgG1 isotype control mAb were added for 30 min at 4°C.
Analyses were performed on a FACSCAN® using the software Cell Quest (Becton Dickinson, San Jose, CA, USA). Forward and side scatter gates were set to include all viable cells. In single-staining experiments, a gate was set to exclude 99% of the cells stained with control Ig. The gates for control Ig in double-staining experiments were placed to limit, to less than 1%, not only the individual percentages in the upper left, upper right, and lower right quadrants, but also the sum of the percentages in the upper left and upper right quadrant, or in the upper right and lower right quadrant.
Proliferation assays
The proliferation rates of normal skin-FB as well as OA-SFB and RA-SFB, derived/negatively isolated from primary culture, from the fourth passage of the isolated cells (in the case of OA-SFB and RA-SFB), or from conventional fourth passage, were assessed by seeding the cells at a density of 1.7 × 103/well in 96-well plates and subsequent culture in DMEM/10% FCS with the aforementioned additives for 24 h at 37°C and 5% CO2. The cells were then starved in DMEM/1% FCS for 72 h and subsequently stimulated for 18 h by addition of 50, 100, or 150 U/ml IL-1β (Genzyme, Rüsselsheim, Germany) or 2.5, 5, or 10 U/ml PDGF-BB (R&D Systems, Wiesbaden, Germany). A total of 1 μCi [3H]-thymidine (Amersham, Braunschweig, Germany) was added to each well, the cells cultured for an additional 18 h, harvested, and the incorporated radioactivity determined in a β-counter (Canberra-Packard, Frankfurt/Main, Germany).
Statistical analysis
The data were first subjected to the multi-group Kruskal-Wallis test because of multiple comparisons. Only those parameters showing significant differences (P ≤ 0.05) underwent further analysis. The non-parametric Mann–Whitney U test was then applied for analysis of the phenotypic and functional features. The Spearman Rank correlation test was used to analyze correlations among phenotypic features of SFB and between these features and the clinical status/treatment of individual patients. Differences were considered statistically significant in all cases for P ≤ 0.05. Analyses were performed using the SPSS 9.0™ program (SPSS Inc., Chicago, IL, USA).
Results
Expression of Thy-1 in RA synovial tissue
The mAb AS02 was tested by immunohistochemistry on cryostat sections of RASM to verify the feasibility of this anti-Thy-1 mAb for positive identification/isolation of SFB. The mAb AS02 stained connective tissue cells, while largely sparing the lining layer (see Fig. 3A for details), therefore reproducing the known distribution of FB within the SM [21,23,24]. The mAb AS02 showed no overlap with an anti-CD14 mAb in double-staining experiments, thereby excluding crossreactivity with monocytes/macrophages (Fig. 3C,E). However, the mAb AS02 stained endothelial cells, as demonstrated by double-staining with rabbit Ig against von Willebrand factor (Fig. 3D,F,G). Cultured, non-stimulated HUVEC, in contrast, did not express Thy-1 (data not shown). These results, in line with cytokine induction of Thy-1 expression on endothelial cells in culture [44,45,46], suggest that Thy-1 expression on RASM endothelial cells may reflect ongoing inflammation [46].
Figure 3.
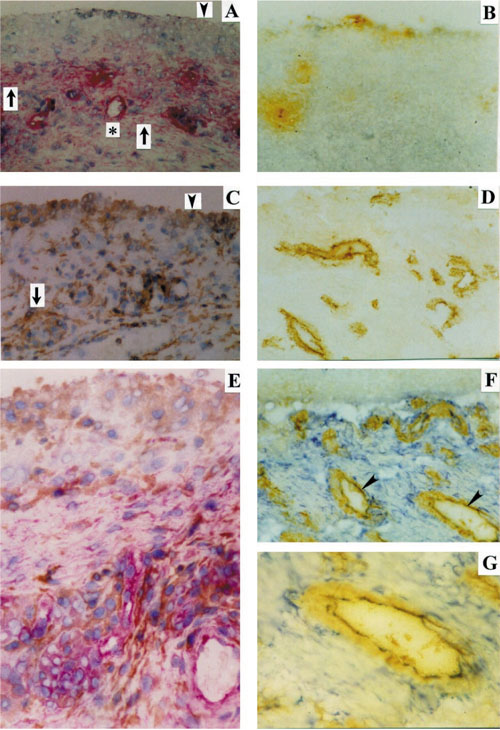
Immunohistochemical staining of the RA synovial membrane with the anti-Thy-1 mAb AS02. (A) This mAb strongly stained cells in the connective tissue layer beneath the lining layer (APAAP, red staining; arrows), while largely sparing cells of the lining layer (arrowhead). Endothelial cells (asterisk) and connective tissue cells in diffuse infiltrates were also stained, more weakly in the latter case. In serial sections, the anti-CD14 mAb MoS39 (kindly provided by Prof GR Burmester, Berlin, Germany) stained macrophages in the lining layer (arrowhead in (C)) and diffuse inflammatory infiltrates (peroxidase, brown staining; arrow in (C), (E)); the mAb against von Willebrand factor stained endothelial cells (peroxidase, brown staining; (D), (F), (G)). In double-staining experiments, the anti-Thy-1 mAb AS02 showed no overlap with anti-CD14 staining (E). In contrast, endothelial cells were double-stained with both the antibody against von Willebrand factor and the anti-Thy-1 mAb AS02 [arrowheads in (F)(G)]. There was no positive reaction with control isotype-matched mAbs (APAAP and peroxidase) (B), employed as controls for double-staining [(E), (F), (G)]. Original magnification: (A)-(D) and (F), 92 ×; (E) and (G), 184 ×.
Phenotype analysis of RA synovial cells in primary culture
RA synovial cells were subjected to extensive analysis to carefully characterize the starting population before negative isolation (ie the 7-day primary culture of synovial cells resulting from trypsin/collagenase digestion of the RASM). The primary culture contained large, spindle-shaped Thy-1+ SFB (Fig. 1C) and small, round CD14+ cells, most probably macrophages (Fig. 1D; mAb Tyk4), as detected by immunohistochemical staining [6,9]. Endothelial cells were absent, as confirmed by lack of staining for von Willebrand factor (Fig. 1F; mAb 4F9) and CD144 (Fig. 1G; mAb Cadherin 5), which clearly identified HUVEC (data not shown). The FB nature of the spindle-shaped cells was confirmed by intracellular staining for procollagen I and III (Fig. 1E,H; rabbit antibodies MP I and MP III). An average of approximately 62% of the cells stained with the anti-Thy-1 mAb AS02 (n = 4 RA patients; Table 1a and Fig. 4A) in FACS analysis [10]. The average percentage of CD14+ cells was approximately 15% (n = 4; Table 1a and Fig. 4B). There were <1% T cells (CD3+; mAb UCHT-1), B cells (CD19+/20+; mAbs HD 37 and B-Ly 1), plasma cells (CD38+; mAb AT 13/5), NK cells (CD56+; mAb NKH/1), dendritic cells (CD83+; mAb HB 15a), endothelial cells (CD144+), or PMN (CD15+; mAb 80H5), indicating that non-adherent cells had been efficiently removed during primary culture. Endothelial cells presumably did not adhere to the culture dish due to the absence of gelatin coating and unfavorable medium composition (see Materials and Methods).
Figure 4.
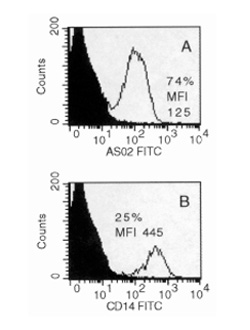
FACS analysis of primary-culture RA synovial cells (following 7 days of culture). The primary culture contained virtually only cells staining either with the FB-directed anti-Thy-1 mAb AS02 (A) or with the monocyte/macrophage-specific anti-CD14 mAb Tyk4 (B). FITC, fluoresceine isothiocyanate.
Adherent synovial cells were then detached by short-term trypsinization for 2 min (0.25% trypsin/0.2% EDTA; Gibco) and used for negative isolation. The total yield of cells following 7 days of RA primary culture averaged (5.2 ± 1.1) × 107 cells (mean ± SEM; n = 7; open synovectomy samples). The yield of cells from arthroscopic synovectomy samples ([3.1 ± 0.6] × 107 cells; n = 3) was comparable. There was no significant difference between the yield of cells from the RA or OA SM.
Flow cytometry/histochemical analysis of cells negatively isolated from primary culture
Negative isolation from primary culture using Dynabeads® M-450 CD14 (clone RMO52) resulted in cells that were Thy-1+ (on average, approximately 74%; n = 9; Fig. 2A and Table 1b) and, more importantly, prolyl-4-hydroxylase+ (on average, approximately 85%; n = 9; Table 1b) (mAb 3-2B12, Dianova), as shown by FACS analysis (Table 1b) and confirmed by immunohistochemistry in chamber slides. There were very few contaminating non-specific esterase+ (n = 3 RA and n = 3 OA patients; Fig. 2C), CD14+, CD68+, and/or CD11b+ macrophages (<2%; Fig. 2B,D and Table 1b), as well as <1% T cells (CD3+), B cells (CD19+/20+), plasma cells (CD38+), NK cells (CD56+), dendritic cells (CD83+), PMN (CD15), or endothelial cells (CD144+; von Wille-brand factor-positive).
The average yield of RA-SFB negatively isolated from primary culture was (2.8 ± 0.9) × 107 cells (mean ± SEM; n = 7; open synovectomy samples). A similar yield was observed upon isolation of cells from arthroscopic synovectomy samples ([1.7 ± 0.6] × 107 cells; n = 3). There was no significant difference between the yield from the primary culture of the RASM and that of the OA SM.
Inclusion of the trypsin component in the initial tissue digestion led to a considerably higher yield of cells following negative isolation from primary culture (1.2-fold to 5-fold; n = 3) than without trypsin or with DNAse instead of trypsin.
The composition of the cells was very similar to that obtained with primary-culture or first-passage normal skin-FB (Table 1g). Comparable results, although with a considerably lower percentage of Thy-1+ cells, were also obtained when OA-SFB were negatively isolated from primary culture (Table 1e). Conventional fourth-passage RA-SFB (Table 1d) or OA-SFB (Table 1f) showed FB markers on a high percentage of cells (>98% prolyl-4-hydroxylase+ cells; >80% Thy-1+ cells) and contained virtually no contaminating macrophages (<2% CD14+-positive or CD68+-positive cells). The same was true for isolated primary RA-SFB kept in culture until fourth passage (Table 1c).
In vitro morphology of negatively isolated RA-SFB upon reculture
Negatively isolated RA-SFB showed almost exclusively spindle-shaped or stellate, flat morphology when recultured (Fig. 5A). Recultured CD14+ cells (Fig. 5B) exhibited small, round morphology with attached Dynabeads® and contained only very few cells with FB morphology.
Figure 5.
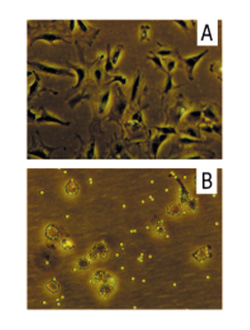
Morphology of negatively isolated RA-SFB and macrophages upon reculture. When recultured, the RA-SFB obtained by negative isolation with Dynabeads® M-450 CD14 showed almost exclusively spindle-shaped, flat or stellate morphology (A). Recultured CD14+ cells (B) exhibited small, round, macrophage-like morphology with attached magnetobeads, and contained only very few cells with FB morphology. Original maginification: 368 ×.
Phenotype characterization of negatively isolated RA-SFB
To verify whether isolated RA-SFB displayed the features observed in situ in the RASM [1,2,12,13,14,15, 21,22,23,24,25,28], the expression of several surface or intracellular/nuclear molecules was investigated by FACS analysis (Fig. 6 and Table 4). The phenotype was compared with that of primary-culture normal skin-FB and isolated primary-culture OA-SFB (Fig. 6 and Table 4).
Figure 6.
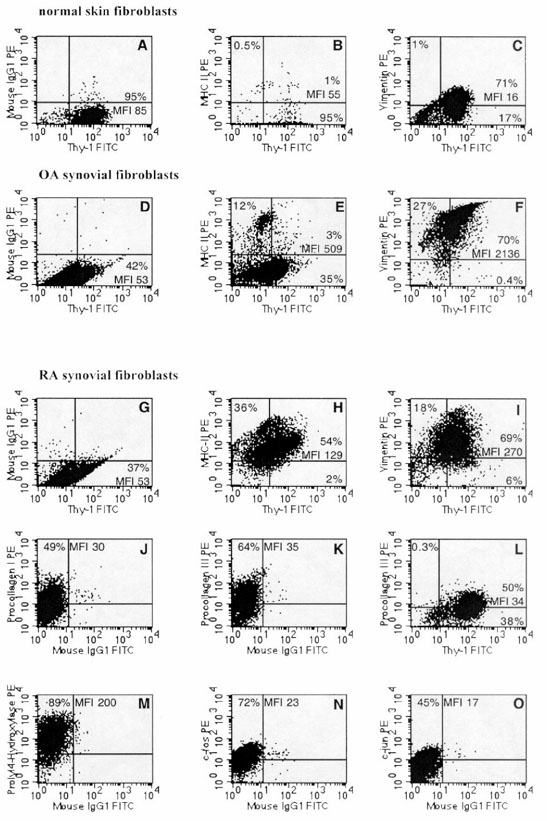
Phenotype of isolated primary-culture RA-SFB (lower panels) in comparison with primary-culture normal skin-FB (upper panel) and isolated primary-culture OA-SFB. The double-staining experiments were performed with the anti-Thy-1 mAb AS02. (A)-(C) The expression of Thy-1 (A), MHC-II/Thy-1 (B) (double-staining), and vimentin/Thy-1 (C) (double-staining) in normal skin-FB; (D)-(F) the expression of Thy-1 (D), MHC-II/Thy-1 (E) (double-staining), and vimentin/Thy-1 (F) (double-staining) in OA-SFB; (G)-(I) the expression of the antigens Thy-1 (G), MHC-II/Thy-1 (H) (double-staining), vimentin/Thy-1 (I) (double-staining) on RA-SFB; (J)-(L) the expression of the cytoplasmic antigens procollagen I (J) and procollagen III/Thy-1 (K) (single-staining for procollagen III) and (L) (double-staining) in RA-SFB. Expression of prolyl-4-hydroxylase (M) and the proto-oncogenes c-Fos (N), and c-Jun (O) in RA-SFB is also shown. See Results and Table 4 for mean values and statistical comparison among the different FB types. PE, phycoerythrine; FITC, fluoresceine isothiocyanate.
Table 4.
Phenotype of negatively isolated rheumatoid arthritis (RA)-synovial fibroblasts (SFB) in comparison with osteoarthritis (OA)-SFB and normal skin-FB
| skin-FB (n = 8) | OA-SFB (n = 9) | RA-SFB (n = 9) | ||||
| Antigen | Percent | MFI | Percent | MFI | Percent | MFI |
| Thy-1 (CD90; mAb AS02) | 91.4 ± 3.0 | 156 ± 36 | 62.1 ± 9.0# | 120 ± 36 | 73.8 ± 9.6 | 109 ± 20 |
| MHC-I | 90.2 ± 8.5 | 414 ± 119 | c87.7 ± 6.8 | 759 ± 170 | a77.2 ± 8.1 | 130 ± 54x |
| MHC-II | 1.5 ± 0.4 | 225 ± 67 | c17.0 ± 7.3# | 678 ± 137# | a65.8 ± 12.3#x | 223 ± 84x |
| Aminopeptidase N (CD13) | b84.0 ± 10.2 | 365 ± 163 | a83.2 ± 8.7 | 794 ± 207 | 84.8 ± 5.9 | 444 ± 226 |
| VCAM-1 (CD 106) | c8.3 ± 3.4 | 66 ± 33 | a28.9 ± 13.2 | 156 ± 82 | c17.5 ± 9.8 | 122 ± 42 |
| Prolyl-4-hydroxylase | 70.7 ± 15.2 | 101 ± 31 | a92.2 ± 2.8 | 212 ± 44 | 84.8 ± 2.9 | 164 ± 45 |
| Vimentin | a90.3 ± 4.2 | 133 ± 34 | b94.2 ± 1.7 | 1083 ± 530 | a93.2 ± 1.5 | 592 ± 290 |
| Procollagen I | d68.2 ± 8.3 | 18 ± 1 | d27.5 ± 10.4 | 39 ± 5# | a44.8 ± 13.0 | 25 ± 2x |
| Procollagen III | d55.1 ± 9.8 | 15 ± 2 | d22.1 ± 7.8 | 45 ± 9# | a50.0 ± 14.3 | 26 ± 3# |
| c-Fos | d87.2 ± 2.1 | 17 ± 3 | d53.9 ± 10.4 | 35 ± 2# | a57.4 ± 14.0 | 20 ± 2x |
| c-Jun | d42.7 ± 15.6 | 13 ± 3 | d22.2 ± 10.3 | 29 ± 3 | b42.5 ± 14.0 | 21 ± 5 |
| Jun-D | d87.4 ± 1.5 | 19 ± 5 | d37.6 ± 17.7 | 33 ± 1# | a53.0 ± 14.8 | 21 ± 4 |
Data presented as means ± SEM. MFI, Mean fluorescence intensity; mAb, monoclonal antibody; MHC, major histocompatibility complex; VCAM-1, vascular adhesion molecule-1. Significant differences among groups are indicated in bold. All cell populations were isolated/obtained from primary culture (in the case of skin-FB from primary culture or first passage; see Methods for details). #P ≤ 0.05 Mann-Whitney U test versus skin-FB. xP ≤ 0.05 Mann-Whitney U test versus OA-SFB. an = 7 Normals, OA or RA patients. bn = 6 Normals, OA or RA patients. cn = 5 Normals, OA or RA patients. dn = 3 Normals or OA patients.
Surface antigens
Approximately 74% of RA-SFB expressed the Thy-1 antigen (Fig. 6G and Table 4), compared with 91% of normal skin-FB (Fig. 6A and Table 4). Approximately 77% of the cells expressed MHC-I molecules, compared with 90% of normal skin-FB (Table 4). Strikingly, approximately 66% of the cells expressed MHC-II molecules (Fig. 6H and Table 4), significantly more than OA-SFB (17%; Fig. 6E and Table 4) and normal skin-FB (2%; Fig. 6B and Table 4). At the same time, however, the MFI for MHC-II did not significantly differ between RA-SFB and normal skin-FB (Table 4).
The expression of CD13 (aminopeptidase N; EC 3.4.11.2) on 85% of RA-SFB was comparable with that of normal skin-FB (84%; Table 4).
A low or moderate and variable percentage of normal skin-FB, as well as OA-SFB and RA-SFB, expressed VCAM-1 (using two different anti-VCAM-1 mAbs; see Table 3) without statistically significant differences among these three different FB preparations (Table 4).
Cytoplasmic antigens
Approximately 85% of the RA-SFB expressed the FB marker prolyl-4-hydroxylase (Fig. 6M, and Tables 1 and 4), comparable with the expression in normal skin-FB (71%; Tables 1 and 4).
RA-SFB showed a similar percentage but a higher MFI for the cytoskeletal protein vimentin (Fig. 6I) than normal skin-FB (Fig. 6C and Table 4), although the MFI difference did not attain statistical significance.
A mean of approximately 45% and 50% of the cells, respectively (Table 4), expressed procollagen I (Fig. 6J) and procollagen III (Fig. 6K), as confirmed by double-staining (Fig. 6L). The percentage of cells expressing procollagen I and III was similar to that of normal skin-FB (68% and 55%, respectively; Table 4); however, the MFI for procollagen III was significantly higher in RA-SFB (26 versus 15; Table 4).
Cytoplasmic/intranuclear antigens
Approximately 57% of RA-SFB expressed c-Fos (Fig. 6N and Table 4). Neither the percentage nor the MFI were significantly different from those of normal skin-FB.
The proto-oncogenes c-Jun and Jun-D (both percentage of positive cells and MFI) were also comparably expressed in RA-SFB and normal skin-FB (Fig. 6O and Table 4).
The differences between RA-SFB and normal skin-FB were, in general, not specific to RA since they were also observed in the comparison between the disease control OA-SFB and normal skin-FB (Table 4). In fact, some of the differences in OA were even more pronounced than in RA, attaining statistical significance in the direct comparison between OA and RA (Table 4). Briefly, significantly higher MFI levels in OA-SFB were noted for MHC-I, MHC-II and procollagen I, as well as c-Fos. Opposite differences among the different FB preparations, when considering the percentage of positive cells or the MFI (eg for the comparison of MHC-II and procollagen I and III expression in RA-SFB versus OA-SFB), appeared due to large variations in one or both parameters.
Comparison of FACS analysis and immunohistochemistry
To verify whether the short-term trypsin digestion (2 min at 37°C) employed for removal of the cells from the culture dishes and subsequent FACS analysis altered the expression of the antigens, selected experiments were carried out to compare the expression of surface and intracellular antigens in FACS analysis (Table 1b,e, and Table 4) and immunohistochemistry on chamber slides. Negatively isolated SFB from n = 3 OA and n = 3 RA patients showed comparable percentages of positive cells for surface antigens (RA-Thy-1, 95.3% FACS versus 85% immunohistochemistry; CD14, 1.1% versus 0%; VCAM-1, 0.7% versus 0%) and intracellular antigens (RA-prolyl-4-hydroxylase, 94.7% versus 85%; CD68, 1.7% versus 0%) with both methods, confirming previously published results [47] and excluding the possibility that short-term trypsinization significantly influences antigen expression.
Phenotype comparison of primary-culture and fourth-passage RA-SFB
In order to assess differences between the primary RA-SFB derived from the isolation technique developed in the present study (immediately following isolation or at fourth passage) and those obtained by conventional repeated passaging, the phenotypic features of the three preparations were compared.
Isolated primary RA-SFB versus conventional fourth-passage RA-SFB
The percentages of RA-SFB positive for MHC-II, as well as the MFI for VCAM-1 and c-Jun, were significantly decreased in conventional fourth passage in comparison with isolated primary RA-SFB (Fig. 7C,D,K,L and Table 5). The percentages of cells positive for MHC-I, CD13, prolyl-4-hydroxylase, vimentin, procollagen I and III, c-Fos and Jun-D were, in contrast, significantly increased in conventional fourth passage (Fig. 7E,F,G,H,I,J and Table 5).
Figure 7.
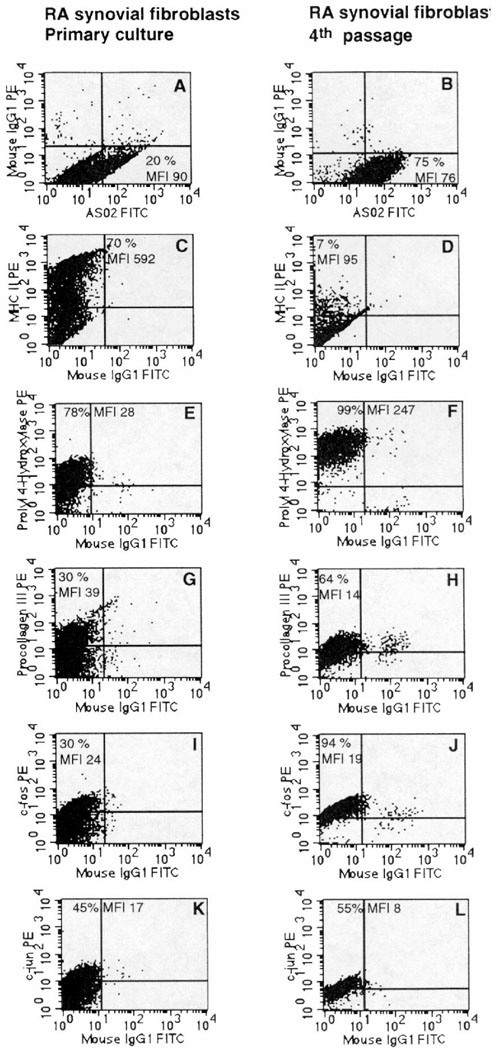
Phenotype comparison of isolated primary-culture (left column) and conventional fourth-passage RA-SFB (right column) from one representative patient. Expression of Thy-1 (A, B), MHC-II (C, D), prolyl-4-hydroxylase (E, F), procollagen III (G, H), c-Fos (I, J), and c-Jun (K, L). While the percentages of cells positive for MHC-II (C) and the MFI for c-Jun (K) were significantly higher in isolated primary-culture RA-SFB, the percentages of cells positive for prolyl-4-hydroxylase (F), procollagen III (H), and c-Fos (J) were significantly higher in conventional fourth passage (see Table 5 for details). PE, phycoerythrine; FITC, fluoresceine isothiocyanate.
Table 5.
Phenotype of negatively isolated primary-culture rheumatoid arthritis (RA)-synovial fibroblasts (SFB) in comparison with fourth-passage RA-SFB
| Primary-culture | Fourth-passage | Fourth-passage | ||||
| RA-SFB | RA-SFB | RA-SFB | ||||
| Isolated cells | Isolated cells | Conventionally passaged cells | ||||
| (n = 9 patients) | (n = 3 patients) | (n = 6 patients) | ||||
| Antigen | Percent | MFI | Percent | MFI | Percent | MFI |
| Thy-1 (CD90; mAb AS02) | 73.8 ± 9.6 | 109 ± 20 | 86.6 ± 4.6 | 41 ± 7# | 85.9 ± 4.3 | 95 ± 17x |
| MHC-I | a77.2 ± 8.1 | 130 ± 54 | 99.2 ± 0.2# | 117 ± 48 | 97.4 ± 1.4# | 175 ± 78 |
| MHC-II | a65.8 ± 12.3 | 223 ± 84 | 4.7 ± 2.9# | 59 ± 25 | 4.8 ± 2.4# | 80 ± 21 |
| Aminopeptidase N (CD13) | 84.8 ± 5.9 | 444 ± 226 | 97.2 ± 1.6 | 108 ± 49 | 97.9 ± 1.2# | 236 ± 95 |
| VCAM-1 (CD106) | c17.5 ± 9.8 | 122 ± 42 | n.d. | n.d. | c0.6 ± 0.1 | 21 ± 5# |
| Prolyl-4-hydroxylase | 84.8 ± 2.9 | 164 ± 45 | 98.7 ± 0.8# | 164 ± 62 | 98.3 ± 0.7# | 189 ± 82 |
| Vimentin | a93.2 ± 1.5 | 592 ± 290 | 99.5 ± 0.4# | 717 ± 126 | 99.7# | 325 |
| Procollagen I | a44.8 ± 13.0 | 25 ± 2 | 92.7 ± 2.8# | 25 ± 12 | 91.0 ± 4.0# | 28 ± 5 |
| Procollagen III | a50.0 ± 14.3 | 26 ± 3 | 94.1 ± 1.8# | 21 ± 12 | 90.2 ± 5.2# | 22 ± 4 |
| c-Fos | a57.4 ± 14.0 | 20 ± 2 | 63.5 ± 16.4 | 10 ± 5 | 95.0 ± 0.6#x | 17 ± 2 |
| c-Jun | b42.5 ± 14.0 | 21 ± 5 | n.d. | n.d. | d42.1 ± 12.0 | 9 ± 2# |
| Jun-D | a53.0 ± 14.8 | 21 ± 4 | 68.1 ± 13.2 | 11 ± 5 | 93.0 ± 1.3#x | 18 ± 3 |
Data presented as means ± SEM. MFI, Mean fluorescence intensity; mAb, monoclonal antibody; MHC, major histocompatibility complex; VCAM-1, vascular adhesion molecule-1; n.d., not determined. For vimentin expression in conventionally passaged fourth-passage RA-SFB, the median is shown because of a large deviation from the mean. Significant differences between groups are indicated in bold. #P ≤ 0.05 Mann–Whitney U test versus primary-culture RA-SFB. xP ≤ 0.05 Mann–Whitney U test versus fourth-passage RA-SFB (isolated cells). an = 7 RA patients. bn = 6 RA patients. cn = 5 RA patients. dn = 4 RA patients.
The upregulation of the proto-oncogenes c-Fos and Jun-D was limited to RA, since the percentages of cells positive for these molecules were significantly decreased upon passaging in OA-SFB (c-Fos: isolated primary-culture OA-SFB, 53.9 ± 10.4%; conventional fourth-passage OA-SFB, 15.0 ± 6.6%; Jun-D: isolated primary-culture OA-SFB, 37.6 ± 17.7%; conventional fourth-passage OA-SFB, 25.1 ± 4.2%; n =3 for both SFB preparations) and numerically decreased upon passaging in normal skin-FB (c-Fos: primary-culture/first-passage skin-FB, 87.2 ± 2.1%; conventional fourth-passage skin-FB, 59.4 ± 26.0%; Jun-D: primary-culture/first-passage skin-FB, 87.4 ± 1.5%; conventional fourth-passage skin-FB, 66.2 ± 18.6%; n = 3 for both). These decreases were also noted for the MFI (data not shown). The percentage of positive cells and/or MFI for c-Fos and Jun-D in conventional fourth-passage RA-SFB were significantly higher than in OA-SFB as a consequence of these reciprocal changes, under these circumstances confirming previously published data [25].
Isolated primary RA-SFB versus isolated fourth-passage RA-SFB
Some of the differences observed between isolated primary RA-SFB and conventional fourth passage also arose when the isolated primary RA-SFB were passaged until the fourth passage; this applied to the significantly increased percentage of cells positive for MHC-I, prolyl-4-hydroxylase, vimentin, and procollagen I and III, as well as to the decreased percentage of MHC-II-positive cells (Table 5). In addition, isolated fourth-passage RA-SFB showed a significantly decreased MFI (but a numerically increased percentage) for Thy-1 in comparison with isolated primary RA-SFB (Table 5).
Conventional fourth-passage RA-SFB versus isolated fourth-passage RA-SFB
The most important difference between these two preparations was the significantly increased percentage of cells positive for the proto-oncogenes c-Fos and Jun-D, observed only in conventional fourth-passage RA-SFB (Table 5). The MFI for Thy-1 was also significantly higher in conventional fourth-passage cells.
Proliferation rates of first- and fourth-passage FB
The proliferation rates of skin-FB and SFB were determined and the different preparations compared.
Isolated first-passage FB versus conventional fourth-passage FB
At rest, the proliferation rates of conventional fourth-passage normal skin-FB did not significantly differ from those of the corresponding cells in first passage (Fig. 8A). The same was true for conventional fourth passage OA-SFB and RA-SFB (Fig. 8B,C). Upon stimulation with PDGF (2.5 and 5 U/ml for all cells, 10 U/ml only for OA-SFB), however, the proliferation rates of skin, RA, and OA conventional fourth-passage FB were significantly higher (maximum mean increase, 5.4-fold) than those observed in first-passage cells (Fig. 8A,B,C). Following stimulation with IL-1β, in contrast, only conventional fourth-passage RA-SFB (150 U/ml IL-1β) showed significantly higher proliferation rates than first-passage cells (Fig. 8C).
Figure 8.
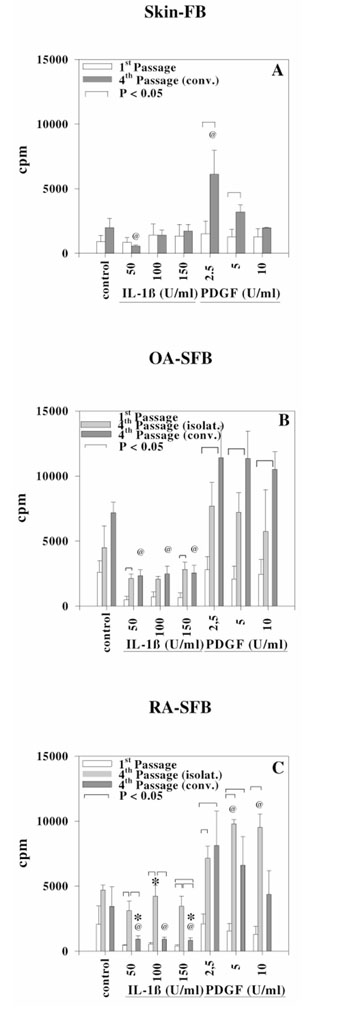
Proliferation rates, as assessed by incorporation of [3H]-thymidine (counts per minute [cpm]) in: first-passage (white bars) and conventional (conv.) fourth-passage (dark gray bars) normal skin-FB (n = 3 each) (A) isolated first-passage (white bars), isolated (isolat.) fourth-passage (light gray bars), and conventional fourth-passage (dark gray bars) OA-SFB (n = 3, 3, and 4, respectively) (B) and isolated first-passage (white bars), isolated (isolat.) fourth-passage (light gray bars), and conventional fourth-passage (dark gray bars) RA-SFB (n = 4, 3, and 5, respectively) (C), at rest (control) or following stimulation with IL-1β (50, 100, or 150 U/ml) or PDGF-BB (2.5, 5, or 10 U/ml). See Results for details. @ P ≤ 0.05 for the comparison between cytokine-stimulated FB and non-stimulated control FB within the same FB preparation. *P ≤ 0.05 for the comparison between OA-SFB and RA-SFB.
Isolated first-passage versus isolated fourth-passage OA-SFB and RA-SFB
The proliferation rates of isolated fourth-passage cells in OA-SFB differed significantly from those of isolated first-passage cells only upon stimulation with IL-1β (50 and 150 U/ml; Fig. 8B). Such differences were not observed at rest or following stimulation with PDGF.
Isolated fourth-passage RA-SFB, in contrast, showed significantly higher proliferation rates than first-passage cells upon stimulation with all concentrations of IL-1β and PDGF, but not at rest (Fig. 8C).
Conventional fourth-passage versus isolated fourth-passage OA-SFB and RA-SFB
The proliferation rates of isolated fourth-passage SFB in both OA and RA were comparable with those of conventional fourth-passage SFB, whether at rest or PDGF-stimulated (Fig. 8B,C). Stimulation with IL-1β (at all concentrations), in contrast, induced significantly higher proliferation in isolated fourth-passage RA-SFB than in conventional fourth-passage RA-SFB (Fig. 8C). This difference was specific for RA-SFB, since it was not observed in OA-SFB (Fig. 8B).
As a consequence of differential IL-1β sensitivity in RA-SFB and OA-SFB, there were significant differences between isolated fourth-passage RA-SFB and OA-SFB (P ≤ 0.05; RA>OA, for 100 U/ml IL-1β), as well as between conventional fourth-passage RA-SFB and OA-SFB (P ≤ 0.05; OA>RA, for 50 and 150 U/ml IL-1β; Fig. 8B,C).
Alternative attempts to isolate RA-SFB
The following isolation techniques were also tested to compare the quality and reliability of negative isolation from primary culture with possible alternative methods. The first technique was positive isolation directly from the trypsin/collagenase digest using the FB-directed anti-Thy1/CD90 mAb AS02 and Dynabeads® M-450 goat anti-mouse IgG. This approach, however, considered legitimate on the basis of the immunostaining in RA synovial tissue (Fig. 3A,E), remained unsatisfactory (yield, only approximately 6% of the initial cell numbers). Another technique involved negative isolation directly from the trypsin/collagenase digest. This approach proved technically prohibitive for the amount of different anti-leukocyte and anti-endothelial mAbs and Dynabeads® M-450 goat anti-mouse IgG necessary for complete removal of all contaminating cell types. Positive isolation from primary culture with the anti-Thy-1 mAb AS02 and Dynabeads® M-450 goat anti-mouse IgG was also tested. This method was unsuitable because: first, the percentages of contaminating macrophages were too high/variable (2-15%); second, the isolated SFB showed a significantly reduced surface density for Thy-1 (MFI 19 versus 152 before isolation; n = 4, P ≤ 0.05), presumably due to contact with the mAb used for isolation; third, the magnetobeads remained attached to the positively isolated SFB for as long as 1 week, thus requiring further manipulation for their removal (eg with anti-idiotypic mAbs); fourth, a substantial proportion of SFB (approximately 30%) was lost into the negative fraction; and, finally, a Thy-1-dependent isolation technique bore the risk of selecting for SFB subpopulations, since the prolyl-4-hydroxylase+ cells obtained by negative isolation with Dynabeads® M-450 CD14 contained both a Thy-1+ fraction (RA, 49.3± 14.9%, n = 6; OA, 43.6 ± 12.0%, n = 4) and a Thy-1– fraction, as demonstrated by double-staining FACS analysis.
Discussion
Negative isolation from primary culture
Negative isolation of SFB from primary cultures of RASM using magnetobead-coupled anti-CD14 mAbs proved successful, providing satisfactory numbers of cells and avoiding repeated passaging and numerous cumulative population doublings. With culture times as short as 7 days, fresh SFB express surface and intracellular antigens typical of the FB lineage and/or SFB features as reported at a tissue level (Fig. 6 and Table 4) [1,2,12,13,14,15, 21,22,23,24,25,28]. This approach may also reduce the risk of in vitro growth selection. In the case of conventionally isolated RA-SFB, for example, the higher the number of passages, the higher the percentage of cells with trisomy 7 [33]. Some SFB functions (eg cell density at saturation and expression of transcription factors), in contrast, decrease at later passages [5,32,48].
While the use of enzymes for the initial tissue digestion represents at least a transient stress for the cells, this method may also represent an alternative to tissue-outgrowth techniques, known to select for premitotic FB [26]. This is important since the relative contribution of pre-mitotic or postmitotic FB to the pathogenetic features of RA is presently unclear.
Advantages of negative isolation
Negative isolation with magnetobead-coupled anti-CD14 mAbs shows the following advantages. There is minimal contamination with macrophages (<2%) and other inflammatory cells (all <1%). Thus, although CD14 may be expressed to a lower degree than CD68 on RA synovial lining macrophages [36,39], the degree of CD14-positivity appears more than sufficient to conduct the isolation procedure. The positive fraction, in turn, contains virtually no SFB, indicating minimal cell loss, and thereby also excluding major subset selection. Another advantage is that negative isolation with Dynabeads® M-450 CD14 avoids direct contact of SFB with mAbs and/or magnetobeads, thereby reducing possible functional alterations of the cells. Finally, the anti-Thy-1 mAb AS02 (used in the parallel attempt to positively isolate SFB) identifies 91% of normal skin-FB, but not more than 74% of RA-SFB (Tables 1 and 4). In addition, the prolyl-4-hydroxylase+ cells obtained by negative isolation with Dynabeads® M-450 CD14 contain both a Thy-1+ and a Thy-1– fraction. Thy-1-independent isolation thus circumvents the danger of selecting for Thy-1+ SFB subpopulations, especially relevant since Thy-1+ and Thy-1- FB diverge in several characteristics potentially relevant to RA; for example, cytokine and matrix production, as well as antigen presentation [49] (reviewed in [50,51,52]).
The negative isolation technique, however, also bears possible disadvantages: the limited number of cells obtained (approximately 2.8 × 107), due to the lack of in vitro expansion, and the necessity for reliable access to fresh synovial specimens. On the other hand, the applicability of this procedure not only to joint replacement samples, but also to arthroscopic synovectomy samples, augments the potential sources of fresh synovial tissue.
Advantages of limited-passage isolation
Limited-passage isolation shows the following advantages. It avoids phenotypic and functional in vitro alterations due to repeated passaging; indeed, even a low number of passages increases the proliferation rates in response to cytokine stimulation (Fig. 8). Passaging also alters the cellular phenotype of RA-SFB, as exemplified by a strong decrease of the percentage of MHC-II+ cells, on one hand, and a striking increase of the cells positive for procollagen I and III, and the proto-oncogenes c-Fos and Jun-D, on the other (Figs. 7 and 8, and Table 5). These results are relevant especially in view of the question whether proliferation and some phenotypic features of RA-SFB (in particular, the expression of proto-oncogenes) can be assimilated to those of dysregulated growth [1,2]. As these features seem to depend on the length of in vitro culture, caution should be applied to the interpretation of data from passaged cells [1,2,12,13,14,15, 23,24,25]. Another advantage is that this isolation procedure reduces the time of exposure to surviving macrophages to 7 days (in contrast to the weeks typical of conventional passaging), thus limiting long-term in vitro changes resulting from monokine secretion or cell-cell contact between SFB and synovial macrophages. Limited-passage isolation may therefore allow one to differentiate between FB-inherent changes (eg growth selection and cell senescence) and the changes mediated by the degree or length of contact with surviving macrophages during the initial passages. A high percentage of RA-SFB (and, to a lesser degree, also OA-SFB) from limited-passage isolation express MHC-II molecules without in vitro stimulation with cytokines (Table 4), which in conventionally isolated cells requires stimulation with interferon-gamma (used not only to explore the antigen-presentation potential of RA-SFB, but also to inhibit collagen biosynthesis [19,31,53]). In this aspect, the present in vitro system may be very useful in studies on the interrelationship of SFB with T cells or macrophages, in which the influence of exogenous mediators complicates the design and/or interpretation of the experiments.
Similarities/discrepancies between the in situ and in vitro phenotype of RA-SFB
Similarities
The cells obtained by negative isolation generally showed features similar to those reported in in situ analyses. The expression of Thy-1, CD13 (aminopeptidaseN), and vimentin exemplify this. The expression of these molecules, particularly their local upregulation by cytokines [54,55] and/or heterogeneous expression at different anatomical sites [9,49], may be pathogenetically relevant, either in terms of defining functionally heterogeneous SFB subpopulations [49] (reviewed in [50,51,52]), by providing pro-inflammatory enzymes [17,47,56,57,58,59], or as targets of autoimmune reactions [60].
MHC-II. The constitutive MHC-II expression in isolated RA-SFB and OA-SFB, confirming previous in situ observations in the RA and OA SM [13,21,22,28] and in systemic scleroderma [61], is probably related to the inflammatory microenvironment, since normal skin-FB (Table 4) and normal SFB [31] hardly express MHC-II. This is further supported by the a strong decrease of the percentage of MHC-II+ cells upon repeated passage (both conventional and following negative isolation), possibly due to lack of or a progressive decrease of external stimulation with pro-inflammatory mediators (Fig. 7 and Table 5). Notably, there was a significant, negative correlation between the percentage of MHC-II-positive SFB and treatment of the RA patients with Methotrexate (ρ=-0.866, P = 0.01, n = 7), indicating that effective antirheumatic therapy may be reflected by decreased MHC-II expression on RA-SFB [28].
Procollagen III. The significant increase of the MFI for intracellular procollagen III in RA-SFB and OA-SFB (Table 4) suggests that the pattern of SFB activation may be functional (among other things) to the increase of collagen metabolism; this is supported by the increased expression of collagen α2I and α1III mRNA in the RASM in situ [2,23,24], in comparison with normal or non-RA synovial tissue. Also, in as much as procollagen III is the fetal form of collagen used in wound healing and tissue repair (reviewed in [62,63]), ongoing fibrosis may be a considerable component of the disease process in RA (and OA), in analogy to systemic scleroderma [61] or interstitial kidney fibrosis [11,26]. A striking increase of procollagen I and III expression in RA-SFB from primary culture to fourth passage (increase of positive cells by ≥ 38%; Fig. 7 and Table 5), on the other hand, indicates that primary-culture cells do not exploit their full potential of matrix production.
Discrepancies
The expression of VCAM-1 [1,15,64,65] and Jun and Fos proto-oncogenes [23,25,66,67] showed some in situ / in vitro discrepancies.
VCAM-1. Surprisingly, only a moderate and variable percentage of RA-SFB (whether conventionally passaged or negatively isolated from primary culture) expressed the adhesion molecule VCAM-1 (Tables 4 and 5). There were also no significant differences between RA-SFB and OA-SFB or normal skin-FB (Table 4). This is in apparent contrast to the enhanced expression of VCAM-1 reported in the lining layer of RA and OA synovial tissue [64,65]; however, it is well compatible with the large variability in VCAM-1 expression observed in vitro (Table 4) [1,15,68] and in situ [1,15]. The significant, positive correlation between VCAM-1 expression in isolated primary-culture RA-SFB and the erythrocyte sedimentation rate in RA patients (ρ=1.00, P = 0.000, n = 4) indicates that the variability may depend on disease activity, as also suggested in recent reports [68,69,70].
Proto-oncogene expression. Negatively isolated RA-SFB expressed c-Fos, c-Jun, and Jun-D (Table 4), in analogy to the features of SFB in RASM [23,25,66]. The expression of c-Jun and Jun-D in the present study could be unequivocally demonstrated by FACS analysis (Table 4), while previous immunohistochemical studies had failed to detect these molecules in situ [23]. Notably, however, the degree of proto-oncogene expression (both percentage of positive cells and MFI) did not significantly differ from that of normal skin-FB (Fig. 6 and Table 4) or, as previously reported, of SFB from traumatic joint injury [67]. This finding supports in situ observations that, at a single-cell level, the degree of proto-oncogene expression in RA, OA, and joint trauma is similar [23], thereby questioning whether proto-oncogene expression in RA reflects per se cell transformation and/or severe metabolic abnormalities.
Notably, the expression of the proto-oncogenes c-Fos and Jun-D was strikingly increased in conventional fourth-passage RA-SFB as compared with isolated primary-culture RA-SFB (increase of positive cells ≥ 38%; Fig. 7 and Table 5). This increase was limited to RA, since the percentage of cells positive for these molecules was significantly decreased in OA-SFB and numerically decreased in normal skin-FB upon passaging. As a consequence of these reciprocal changes, the percentage of positive cells and/or MFI for c-Fos and Jun-D in conventional fourth-passage RA-SFB was significantly higher than in OA-SFB, under these circumstances confirming previously published data [25].
Finally, a significant, positive correlation between the percentage of c-fos+ and procollagen I and III+ isolated primary-culture SFB in both RA and OA patients (RA: c-fos/procollagen I, ρ=1.000, P = 0.00, n = 7; c-fos/procollagen III, ρ=0.964, P = 0.00, n = 7) suggests a link between c-fos expression and augmented matrix production by SFB in rheumatic disorders, in line with the known regulation of collagen expression by the activator protein-1 transcription factor (reviewed in [13]).
Suitability of fibroblast markers for unequivocal identification of SFB
None of the markers used to recognize SFB [9,23,24,62] proved to be universal (Table 4) or stable upon passaging (Fig. 7 and Table 5). Cellular uridine diphosphoglucose dehydrogenase activity, a useful histochemical marker for intimal SFB [37], while clearly detectable by immunohistochemistry in the lining layer of RASM tissue sections, was also only positive in a small subpopulation of cultured SFB (90% negative, 9% weakly positive, 1% strongly positive). Thus, despite its parallel recognition of endothelial cells [62], prolyl-4-hydroxylase at present remains the most suitable FB marker (see Tables 1,4 and 5).
The variable degree of positivity for different markers observed in the present study supports the existence of different SFB subpopulations or maturational stages (Tables 4 and 5, and Fig. 6E,H,I) [1,15]. By providing isolated SFB that do not undergo major subset selection, the present technique may allow, on one hand, the further characterization of these subpopulations and, on the other, the identification of more universal and/or less regulated pan-FB markers.
In summary, the present technique allows the isolation of highly enriched SFB with very limited in vitro culture time. These cells clearly show different phenotypic and functional properties in comparison with conventional fourth-passage RA-SFB. In conjunction with simultaneously derived synovial macrophages from the same patient (eg the positive fraction of the CD14 negative isolation), normally lost upon repeated passage due to their inability to divide in vitro (reviewed in [1]), and non-adherent, viable cells from the supernatant of the primary culture, recreation of a simplified 'rheumatoid' system can be attempted to investigate the relative contribution of the different cells to joint pathology [18,71].
Note added in proof
Increased levels of c-Fos protein in late-passage versus early-passage RA-SFB, but decreased levels of c-Fos in late-passage OA-SFB, ie an RA-specific increase of c-Fos transcription/translation in late-passage SFB, has recently been confirmed by Shimizu et al [72].
Acknowledgments
Acknowledgement
D Claus, B Niescher, and B Ukena are gratefully acknowledged for assistance, and Dr MA Kukuruga (University of Michigan, Ann Arbor, MI, USA) for advice on FACS analysis. Dr R Bravo (Bristol Meyers Squibb, Pharmaceutical Research Institute, Princeton, NJ, USA), Dr D Schuppan (Department of Gastroenterology, Klinikum Benjamin Franklin, Berlin, Germany), Dr A Saalbach (Department of Dermatology, University of Leipzig, Germany) and Dr K. Herrmann (Institute of Biomedical Research and Counseling, Leipzig, Germany) are gratefully acknowledged for providing anti-Jun/Fos, anti-procollagen I/III, and anti-Thy-1 antibodies, respectively. Prof J Dippold and Prof G von Salis-Soglio (Department of Orthopedics, University of Leipzig), Dr D Jungmichel (Clinic of Orthopedics, Bad Düben), Prof R Venbrocks (Clinic of Orthopedics, Eisenberg), Dr Sauer (plastic surgeon, Leipzig; Germany), and Dr A Freyberg (Department of Orthopedic Surgery, University of Michigan, Ann Arbor, MI, USA) are gratefully acknowledged for providing SM and skin samples.
The authors are particularly grateful to Prof DA Fox, Prof BJ Roessler, and Prof CW Castor (University of Michigan, Ann Arbor, MI, USA) for support and critical reading of the manuscript.
Sponsorship
This work was supported by grants IZKF Leipzig, 01KS9504 (T Zimmermann), 01KS9504A1 (H-D Stahl) and 01KS9504A3 (U Sack), grants 01VM9311/3 and 01ZZ9602 (RW Kinne) from the Bundesministerium für Bildung und Forschung, and the Novartis Foundation (U Sack).
References
- Firestein GS. Invasive fibroblast-like synoviocytes in rheumatoid arthritis. Passive responders or transformed aggressors? Arthritis Rheum. 1996;39:1781–1790. doi: 10.1002/art.1780391103. [DOI] [PubMed] [Google Scholar]
- Kinne RW, Palombo-Kinne E, Emmrich F. Activation of synovial fibroblasts in rheumatoid arthritis. Ann Rheum Dis. 1995;54:501–504. doi: 10.1136/ard.54.6.501-b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Firestein GS, Alvaro-Gracia JM, Maki R. Quantitative analysis of cytokine gene expression in rheumatoid arthritis. J Immunol. 1990;144:3347–3353. [PubMed] [Google Scholar]
- Faßbender HG. Histomorphological basis of articular cartilage destruction in rheumatoid arthritis. Coll Relat Res. 1983;3:141–155. doi: 10.1016/s0174-173x(83)80040-5. [DOI] [PubMed] [Google Scholar]
- Lafyatis R, Remmers EF, Roberts AB, Yocum DE, Sporn MB, Wilder RL. Anchorage-independent growth of synoviocytes from arthritic and normal joints. Stimulation by exogenous platelet-derived growth factor and inhibition by transforming growth factor-beta and retinoids. J Clin Invest. 1989;83:1267–1276. doi: 10.1172/JCI114011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sack U, Kuhn H, Ermann J, Kinne RW, Vogt S, Jungmichel D, Emmrich F. Synovial tissue implants from patients with rheumatoid arthritis cause cartilage destruction in knee joints of SCID.bg mice. J Rheumatol. 1994;21:10–16. [PubMed] [Google Scholar]
- Müller-Ladner U, Kriegsmann J, Franklin BN, Matsumoto S, Geiler T, Gay RE, Gay S. Synovial fibroblasts of patients with rheumatoid arthritis attach to and invade normal human cartilage when engrafted into SCID mice. Am J Pathol. 1996;149:1607–1615. [PMC free article] [PubMed] [Google Scholar]
- Arnett FC, Edworthy SM, Bloch DA, Mc Shane DJ, Fries JF, Cooper NS, Healey LA, Kaplan SR, Liang MH, Luthra HS. The American Rheumatism Association 1987 revised criteria for the classification of rheumatoid arthritis. Arthritis Rheum. 1988;31:315–324. doi: 10.1002/art.1780310302. [DOI] [PubMed] [Google Scholar]
- Saalbach A, Anderegg U, Bruns M, Schnabel E, Herrmann K, Haustein UF. Novel fibroblast-specific monoclonal antibodies: properties and specificities. J Invest Dermatol. 1996;106:1314–1319. doi: 10.1111/1523-1747.ep12349035. [DOI] [PubMed] [Google Scholar]
- Pelegrí C, Kühnlein P, Buchner E, Schmidt CB, Franch A, Castell M, Hünig T, Emmrich F, Kinne RW. Depletion of γ/δ T cells does not ameliorate, but rather aggravates, adjuvant arthritis. Arthritis Rheum. 1996;39:204–215. doi: 10.1002/art.1780390206. [DOI] [PubMed] [Google Scholar]
- Müller GA, Rodemann HP. Characterization of human renal fibroblasts in health and disease: I. Immunophenotyping of cultured tubular epithelial cells and fibroblasts derived from kidneys with histologically proven interstitial fibrosis. Am J Kidney Dis. 1991;17:680–683. doi: 10.1016/s0272-6386(12)80351-9. [DOI] [PubMed] [Google Scholar]
- Case JP, Lafyatis R, Remmers EF, Kumkumian GK, Wilder RL. Transin/stromelysin expression in rheumatoid synovium. A transformation-associated metalloproteinase secreted by phenotypically invasive synoviocytes. Am J Pathol. 1989;135:1055–1064. [PMC free article] [PubMed] [Google Scholar]
- Ritchlin CT, Winchester RJ. Potential mechanisms for coordinate gene activation in the rheumatoid synoviocyte: implications and hypotheses. Springer Semin Immunopathol. 1989;11:219–234. doi: 10.1007/BF00197304. [DOI] [PubMed] [Google Scholar]
- Trabandt A, Aicher WK, Gay RE, Sukhatme VP, Nilson-Hamilton M, Hamilton RT, McGhee JR, Fassbender HG, Gay S. Expression of the collagenolytic and Ras-induced cysteine proteinase cathepsin L and proliferation-associated oncogenes in synovial cells of MRL/I mice and patients with rheumatoid arthritis. Matrix. 1990;10:349–361. doi: 10.1016/s0934-8832(11)80142-3. [DOI] [PubMed] [Google Scholar]
- Zvaifler NJ, Firestein GS. Pannus and pannocytes. Alternative models of joint destruction in rheumatoid arthritis. Arthritis Rheum. 1994;37:783–789. doi: 10.1002/art.1780370601. [DOI] [PubMed] [Google Scholar]
- Singer NG, Morita Y, Joo YS, Sarkar S, Kinne RW, Anderson CA, Endres J, Fox DA. Evidence for the expression of a second CD6 ligand by synovial fibroblasts. Arthritis Rheum. 2000;43:329–335. doi: 10.1002/1529-0131(200002)43:2<329::AID-ANR12>3.0.CO;2-Y. [DOI] [PubMed] [Google Scholar]
- Chomarat P, Rissoan MC, Pin JJ, Banchereau J, Miossec P. Contribution of IL-1, CD14, and CD13 in the increased IL-6 production induced by in vitro monocyte-synoviocyte interactions. J Immunol. 1995;155:3645–3652. [PubMed] [Google Scholar]
- Scott BB, Weisbrot LM, Greenwood JD, Bogoch ER, Paige CJ, Keystone EC. Rheumatoid arthritis synovial fibroblast and U937 macrophage/monocyte cell line interaction in cartilage degradation. Arthritis Rheum. 1997;40:490–498. doi: 10.1002/art.1780400315. [DOI] [PubMed] [Google Scholar]
- Tsai C, Diaz LA, Jr, Singer NG, Li LL, Kirsch AH, Mitra R, Nickoloff BJ, Crofford LJ, Fox DA. Responsiveness of human T lymphocytes to bacterial superantigens presented by cultured rheumatoid arthritis synoviocytes. Arthritis Rheum. 1996;39:125–136. doi: 10.1002/art.1780390117. [DOI] [PubMed] [Google Scholar]
- Mohr W. Gelenkkrankheiten Stuttgart: Georg Thieme Verlag. 1994.
- Burmester GR, Locher P, Koch B, Winchester RJ, Dimitriu-Bona A, Kalden JR, Mohr W. The tissue architecture of synovial membranes in inflammatory and non-inflammatory joint diseases. I. The localization of the major synovial cell populations as detected by monoclonal reagents directed towards Ia and monocyte-macrophage antigens. Rheumatol Int. 1983;3:173–181. doi: 10.1007/BF00541597. [DOI] [PubMed] [Google Scholar]
- Burmester GR, Jahn B, Rohwer P, Zacher J, Winchester RJ, Kalden JR. Differential expression of Ia antigens by rheumatoid synovial lining cells. J Clin Invest. 1987;80:595–604. doi: 10.1172/JCI113111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kinne RW, Boehm S, Iftner T, Aigner T, Vornehm S, Weseloh G, Bravo R, Emmrich F, Kroczek RA. Synovial fibroblast-like cells strongly express jun-B and c -fos proto-oncogenes in rheumatoid- and osteoarthritis. Scand J Rheumatol. 1995;101(suppl):121–125. doi: 10.3109/03009749509100913. [DOI] [PubMed] [Google Scholar]
- Qu Z, Garcia CH, O'Rourke LM, Planck SR, Kohli M, Rosenbaum JT. Local proliferation of fibroblast-like synoviocytes contributes to synovial hyperplasia: results of proliferating cell nuclear antigen/cyclin, c -myc, and nucleolar organizer region staining. Arthritis Rheum. 1994;37:212–220. doi: 10.1002/art.1780370210. [DOI] [PubMed] [Google Scholar]
- Trabandt A, Aicher WK, Gay RE, Sukhatme VP, Fassbender H-G, Gay S. Spontaneous expression of immediately-early response genes c -fos and egr-1 in collagenase-producing rheumatoid synovial fibroblasts. Rheumatol Int. 1992;12:53–59. doi: 10.1007/BF00300977. [DOI] [PubMed] [Google Scholar]
- Rodemann HP, Müller GA. Characterization of human renal fibroblasts in health and disease: II. In vitro growth, differentiation, and collagen synthesis of fibroblasts from kidneys with interstitial fibrosis. Am J Kidney Dis. 1991;17:684–686. doi: 10.1016/s0272-6386(12)80352-0. [DOI] [PubMed] [Google Scholar]
- Goddard DH, Grossman SL, Williams WV, Weiner DB, Gross JL, Eidsvoog K, Dasch JR. Regulation of synovial cell growth. Coexpression of transforming growth factor [beta] and basic fibroblast growth factor by cultured synovial cells. Arthritis Rheum. 1992;35:1296–1303. doi: 10.1002/art.1780351109. [DOI] [PubMed] [Google Scholar]
- Firestein GS, Paine MM, Littman BH. Gene expression (collagenase, tissue inhibitor of metalloproteinases, complement, and HLA-DR) in rheumatoid arthritis and osteoarthritis synovium. Quantitative analysis and effect of intraarticular corticosteroids. Arthritis Rheum. 1991;34:1094–1105. doi: 10.1002/art.1780340905. [DOI] [PubMed] [Google Scholar]
- Gravallese EM, Darling JM, Ladd AL, Katz JN, Glimcher LH. In situ hybridization studies of stromelysin and collagenase messenger RNA expression in rheumatoid synovium. Arthritis Rheum. 1991;34:1076–1084. doi: 10.1002/art.1780340903. [DOI] [PubMed] [Google Scholar]
- McCachren SS. Expression of metalloproteinases and metal-loproteinase inhibitor in human arthritic synovium. Arthritis Rheum. 1991;34:1085–1093. doi: 10.1002/art.1780340904. [DOI] [PubMed] [Google Scholar]
- Boots AM, Wimmers-Bertens AJ, Rijnders AW. Antigen-presenting capacity of rheumatoid synovial fibroblasts. Immunology. 1994;82:268–274. [PMC free article] [PubMed] [Google Scholar]
- Castor CW, Prince RK, Dorstewitz EL. Characteristics of human 'fibroblasts' cultivated in vitro from different anatomical sites. Lab Invest. 1962;11:703–713. [PubMed] [Google Scholar]
- Ermis A, Henn W, Remberger K, Hopf C, Hopf T, Zang KD. Proliferation enhancement by spontaneous multiplication of chromosome 7 in rheumatic synovial cells in vitro. Hum Genet. 1995;96:651–654. doi: 10.1007/BF00210293. [DOI] [PubMed] [Google Scholar]
- Saalbach A, Kraft R, Herrmann K, Haustein U-F, Anderegg U. The monoclonal antibody AS02 recognizes a protein on human fibroblasts being homologous to Thy-1. Arch Dermatol Res. 1998;290:360–366. doi: 10.1007/s004030050318. [DOI] [PubMed] [Google Scholar]
- Fox DA, Millard JA, Kan L, Zeldes WS, Davis W, Higgs J, Emmrich F, Kinne RW. Activation pathways of synovial T lymphocytes: expression and function of the UM4D4/CDw60 antigen. J Clin Invest. 1990;86:1124–1136. doi: 10.1172/JCI114817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mulherin D, Fitzgerald O, Bresnihan B. Synovial tissue macrophage populations and articular damage in rheumatoid arthritis. Arthritis Rheum. 1996;39:115–124. doi: 10.1002/art.1780390116. [DOI] [PubMed] [Google Scholar]
- Wilkinson LS, Pittsilides AA, Worrall JG, Edwards JCW. Light microscopic characterization of the fibroblast-like synovial intimal cell (synoviocyte). Arthritis Rheum. 1992;35:1179–1184. doi: 10.1002/art.1780351010. [DOI] [PubMed] [Google Scholar]
- Falini B, Flenghi L, Pileri S, Gambacorta M, Bigerna B, Durkop H, Eitelbach F, Thiele J, Pacini R, Cavaliere A, Martelli M, Cardarelli N, Sabattini E, Poggi S, Stein H. PG-M1: a new monoclonal antibody directed against a fixative-resistant epitope on the macrophage-restricted form of the CD68 molecule. Am J Pathol. 1993;142:1359–1372. [PMC free article] [PubMed] [Google Scholar]
- El-Gabalawy H, Canvin J, Guoping MM, Van den Vieren M, Hoffman P, Gallatin M, Wilkins J. Synovial distribution of alphad/CD18, a novel leukointegrin: comparison with other integrins and their ligands. Arthritis Rheum. 1996;39:1913–1921. doi: 10.1002/art.1780391119. [DOI] [PubMed] [Google Scholar]
- Buhling F, Ittenson A, Kaiser D, Tholert G, Hoffmann B, Reinhold D, Ansorge S, Welte T. MRP8/MRP14, CD11b and HLA-DR expression of alveolar macrophages in pneumonia. Immunol Lett. 2000;71:185–190. doi: 10.1016/s0165-2478(00)00164-4. [DOI] [PubMed] [Google Scholar]
- Muller Kobold AC, Kallenberg CGM, Tervaert JWC. Monocyte activation in patients with Wegener's granulomatosis. Ann Rheum Dis. 1999;58:237–245. doi: 10.1136/ard.58.4.237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Firestein GS, Zvaifler NJ. Downregulation of human monocyte differentiation antigens by interferon gamma. Cell Immunol. 1987;104:343–354. doi: 10.1016/0008-8749(87)90036-0. [DOI] [PubMed] [Google Scholar]
- Alvaro-Gracia JM, Zvaifler NJ, Firestein GS. Cytokines in chronic inflammatory arthritis. IV. Granulocyte/macrophage colony-stimulating factor-mediated induction of class II MHC antigen on human monocytes: a possible role in rheumatoid arthritis. J Exp Med. 1989;170:865–875. doi: 10.1084/jem.170.3.865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ishizu A, Ishikura H, Nakamaru Y, Kikuchi K, Koike T, Yoshiki T. Interleukin-1 alpha regulates Thy-1 expression on rat vascular endothelial cells. Microvasc Res. 1997;53:73–78. doi: 10.1006/mvre.1996.1986. [DOI] [PubMed] [Google Scholar]
- Mason JC, Yarwood H, Tarnok A, Sugars K, Harrison AA, Robinson PJ, Haskard DO. Human Thy-1 is cytokine-inducible on vascular endothelial cells and is a signaling molecule regulated by protein kinase C. J Immunol. 1996;157:874–883. [PubMed] [Google Scholar]
- Saalbach A, Wetzig T, Haustein UF, Anderegg U. Detection of human soluble Thy-1 in serum by ELISA. Fibroblasts and activated endothelial cells are a possible source of soluble Thy-1 in serum. Cell Tissue Res. 1999;298:307–315. doi: 10.1007/s004419900079. [DOI] [PubMed] [Google Scholar]
- Schwachula A, Riemann D, Kehlen A, Langner J. Characterization of the immunophenotype and functional properties of fibroblast-like synoviocytes in comparison to skin fibroblasts and umbilical vein endothelial cells. Immunobiology. 1994;190:67–92. doi: 10.1016/S0171-2985(11)80284-6. [DOI] [PubMed] [Google Scholar]
- Aicher WK, Heer AH, Trabandt A, Bridges SL, Jr, Schroeder HW, Jr, Stransky G, Gay RE, Eibel H, Peter HH, Siebenlist U. Overexpression of zinc-finger transcription factor Z-225/Egr-1 in synoviocytes from rheumatoid arthritis patients. J Immunol. 1994;152:5940–5948. [PubMed] [Google Scholar]
- Smith TJ, Sempowski GD, Wang HS, Del Vecchio PJ, Lippe SD, Phipps RP. Evidence for cellular heterogeneity in primary cultures of human orbital fibroblasts. J Clin Endocrinol Metab. 1995;80:2620–2625. doi: 10.1210/jcem.80.9.7673404. [DOI] [PubMed] [Google Scholar]
- Borrello MA, Phipps RP. Differential Thy-1 expression by splenic fibroblasts defines functionally distinct subsets. Cell Immunol. 1996;173:198–206. doi: 10.1006/cimm.1996.0268. [DOI] [PubMed] [Google Scholar]
- Silvera MR, Phipps RP. Synthesis of interleukin-1 receptor antagonist by Thy-1+ and Thy-1- murine lung fibroblast subsets. J Interferon Cytokine Res. 1995;15:63–70. doi: 10.1089/jir.1995.15.63. [DOI] [PubMed] [Google Scholar]
- Sempowski GD, Derdak S, Phipps RP. Interleukin-4 and interferon-γ discordantly regulate collagen biosynthesis by functionally distinct lung fibroblast subsets. J Cell Physiol. 1996;167:290–296. doi: 10.1002/(SICI)1097-4652(199605)167:2<290::AID-JCP13>3.0.CO;2-C. [DOI] [PubMed] [Google Scholar]
- Amento EP, Bhan AK, McCullagh KG, Krane SM. Influences of gamma interferon on synovial fibroblast-like cells. Ia induction and inhibition of collagen synthesis. J Clin Invest. 1985;76:837–848. doi: 10.1172/JCI112041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burmester GR, Stuhlmüller B, Keyszer G, Kinne RW. Mononu-clear phagocytes and rheumatoid synovitis. Mastermind or workhorse in arthritis? Arthritis Rheum. 1997;40:5–18. doi: 10.1002/art.1780400104. [DOI] [PubMed] [Google Scholar]
- Westacott CI, Sharif M. Cytokines in osteoarthritis: mediators or markers of joint destruction? Semin Arthritis Rheum. 1996;25:254–272. doi: 10.1016/s0049-0172(96)80036-9. [DOI] [PubMed] [Google Scholar]
- Bathon JM, Proud D, Mizutani S, Ward PE. Cultured human synovial fibroblasts metabolize kinins and neuropeptides. J Clin Invest. 1992;90:981–991. doi: 10.1172/JCI115975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piela-Smith TH, Korn JH. Aminopeptidase N: a constitutive cell-surface protein on human dermal fibroblasts. Cell Immunol. 1995;162:42–48. doi: 10.1006/cimm.1995.1049. [DOI] [PubMed] [Google Scholar]
- Shipp MA, Look AT. Hematopoietic differentiation antigens that are membrane-associated enzymes: cutting is the key! Blood. 1993;82:1052–1070. [PubMed] [Google Scholar]
- Pierart ME, Najdovski T, Appelboom TE, Deschodt-Lanckman MM. Effect of human endopeptidase 24.11 ('enkephalinase') on IL-1-induced thymocyte proliferation activity. J Immunol. 1988;140:3808–3811. [PubMed] [Google Scholar]
- Kurki P, Helve T, Virtanen I. Antibodies to cytoplasmic intermediate filaments in rheumatic diseases. J Rheumatol. 1983;10:558–562. [PubMed] [Google Scholar]
- Gruschwitz MS, Vieth G. Upregulation of class II major histocompatibility complex and intercellular adhesion molecule 1 expression on scleroderma fibroblasts and endothelial cells by interferon-gamma and tumor necrosis factor alpha in the early disease stage. Arthritis Rheum. 1997;40:540–550. doi: 10.1002/art.1780400321. [DOI] [PubMed] [Google Scholar]
- Konttinen YT, Nykanen P, Nordstrom D, Saari H, Sandelin J, San-tavirta S, Kour T. DNA synthesis in prolyl-4-hydroxylase positive fibroblasts in situ in synovial tissue. An autoradiography-immunoperoxidase double labeling study. J Rheumatol. 1989;16:339–345. [PubMed] [Google Scholar]
- Peacock EE., Jr Collagenolysis and the biochemistry of wound healing. In Wound Repair, edn 3 Philadelphia: WB Saunders, 1984. p. 102.
- Kriegsmann J, Keyszer GM, Geiler T, Bräuer R, Gay RE, Gay S. Expression of vascular adhesion molecule-1 mRNA and protein in rheumatoid synovium demonstrated by in situ hybridization and immunohistochemistry. Lab Invest. 1995;72:209–214. [PubMed] [Google Scholar]
- Morales-Ducret J, Wayner E, Elices MJ, Alvaro-Gracia JM, Zvaifler NJ, Firestein GS. Alpha4/beta 1 integrin (VLA-4) ligands in arthritis. Vascular cell adhesion molecule-1 expression in synovium and on fibroblast-like synoviocytes. J Immunol. 1992;149:1424–1431. [PubMed] [Google Scholar]
- Dooley S, Herlitzka I, Hanselmann R, Ermis A, Henn W, Rem-berger K, Hopf T, Welter C. Constitutive expression of c-fos and c-jun, overexpression of ets-2, and reduced expression of metastasis suppressor gene nm23-H1 in rheumatoid arthritis. Ann Rheum Dis. 1996;55:298–304. doi: 10.1136/ard.55.5.298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anders M, Krohn K, Kroger H, Huser H, Sparmann M, Meissner A, Gombert W. Proto-oncogene expression in cultured synovial fibroblasts of patients with rheumatoid arthritis. Mol Cell Probes. 1991;5:207–213. doi: 10.1016/0890-8508(91)90042-i. [DOI] [PubMed] [Google Scholar]
- Croft D, McIntyre P, Wibulswas A, Kramer I. Sustained levels of VCAM-1 in cultured fibroblast-like synoviocytes can be achieved by TNF-α in combination with either IL-4 or IL-13 through increased mRNA stability. Am J Pathol. 1999;154:1149–1158. doi: 10.1016/s0002-9440(10)65367-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ping L, Sanz I, O'Keefe RJ, Schwarz EM. NF-κB regulates VCAM-1 expression on fibroblast-like synoviocytes. J Immunol. 2000;164:5990–5997. doi: 10.4049/jimmunol.164.11.5990. [DOI] [PubMed] [Google Scholar]
- Dolhain RJEM, Tak PP, Dijkmans BAC, De Kuiper P, Breedveld FC, Miltenburg AMM. Methotrexate reduces inflammatory cell numbers, expression of monokines and of adhesion molecules in synovial tissue of patients with rheumatoid arthritis. J Rheumatol. 1998;37:502–508. doi: 10.1093/rheumatology/37.5.502. [DOI] [PubMed] [Google Scholar]
- Schultz O, Keyszer G, Zacher J, Sittinger M, Burmester GR. Development of in vitro model systems for destructive joint diseases. Novel strategies to establish inflammatory pannus. Arthritis Rheum. 1997;40:1420–1428. doi: 10.1002/art.1780400809. [DOI] [PubMed] [Google Scholar]
- Shimizu K, Kawasaki H, Morisawa T, Nakamura M, Yamamoto E, Yoshikawa N, Doita M, Shiozawa K, Yonehara S, Chihara K, Shiozawa S. Spontaneous and cytokine regulated c-fos gene expression in rheumatoid synovial cells: resistance to cytokine stimulation when the c-fos gene is overexpressed. Ann Rheum Dis. 2000;59:636–640. doi: 10.1136/ard.59.8.636. [DOI] [PMC free article] [PubMed] [Google Scholar]