

Abstract
Protein prenylation in plants was studied by in vivo metabolic 3H-mevalonate labeling in combination with a range of protein synthesis inhibitors. In spinach cotyledons, this posttranslational protein modification was found to be divided into two categories, one representing the conventional prenylation involving farnesyl and geranylgeranyl groups bound to cysteine residues via thioether linkages. This category revealed a similar pattern of prenylated proteins to that observed in mammalian cells and depends on nuclear gene expression. The other category was shown to represent a type of prenylation confined to chloroplasts. It depends on plastid gene expression and does not involve a thioether bond. The modifying isoprenoid could be released from the chloroplastic polypeptides by alkaline treatment and was identified as phytol upon GC-MS analysis. The phytol could readily be derived from all-trans-[3H]farnesol, which, like all-trans-[3H]geranylgeraniol, was taken up by the cotyledons, resulting in incorporation of radiolabel into proteins.
The prenyl groups that are covalently bound to proteins serve important functions. For instance, they make the protein more hydrophobic, thus facilitating membrane association. Additionally, it has been shown that prenyl groups can be required for protein–protein interactions (1) and perhaps protein targeting (2).
In vivo studies have revealed the presence of prenylated proteins in plants (3, 4). A broad range of proteins, with molecular masses between 6 and 200 kDa, was shown to be prenylated. The pattern of prenylation differed markedly from that observed in both mammalian and plant cells in tissue culture (5, 6), where the majority of the prenylated proteins are restricted to the 21- to 26-kDa molecular mass region.
Prenylated proteins identified to date in mammals are all nuclear-encoded such as rhodopsin kinase (7), small GTP-binding proteins (8, 9), and nuclear lamin B (10). In plants only a few proteins, including ANJ1 (11), a GTP-binding protein (12), and the calmodulin CaM53 (13), have been specifically identified as carrying a prenyl modification.
The isoprenoids so far firmly established as being covalently bound to proteins are farnesyl (C15) and geranylgeranyl (C20) groups. Both isoprenoids are bound to one or two cysteine residues at or near the C terminus of the protein via thioether bonds. The isoprenoid substrates for the protein prenylating enzymes are the pyrophosphate forms of the prenyl alcohols farnesol and geranylgeraniol (14–16). Farnesyl pyrophosphate and geranylgeranyl pyrophosphate both are synthesized at various locations in the cell, with the bulk of these compounds being synthesized in the cytosol (17–19). Other isoprenoids that have been implicated in protein prenylation include hydroxylated farnesol, dolichol, pentaprenol, and hexaprenol (20–22).
In general, in vivo studies of protein prenylation are performed by metabolic labeling using mevalonate as a precursor of the isoprenoid compounds. Recently however, it has been demonstrated that both farnesol and geranylgeraniol also can be used as precursors for protein prenylation in tissue culture cells (23, 24). Evidently, these prenyl alcohols can be phosphorylated in vivo by kinases and the existence of such enzymes has been established in both bacteria (25) and rat liver (26, 27).
In a recent study, it was demonstrated that chloroplasts contain several prenylated polypeptides, the majority of which are associated with the thylakoid membrane or the inner envelope membrane (28). These polypeptides largely accounted for the differences observed between the labeling pattern of prenylated polypeptides in mammalian cells in tissue cultures and plant tissue.
In the present study, a plant experimental method was used to investigate the genetic origin (nuclear or extra-nuclear) of the proteins/enzymes involved in plant protein prenylation through the use of specific protein synthesis inhibitors. The results reveal the existence of a type of prenylation that depends on chloroplastic gene products.
MATERIALS AND METHODS
Materials.
Spinach seedlings (Spinacea oleracea L.) were grown on vermiculite at 22–25°C, with a 10-hr photoperiod, for 2–3 weeks. (R,S)-[5-3H]mevalonate (specific activity 2.8 and 22.2 Ci/mmol) was prepared as described by Keller (29). Before use, the mevalonolactone and mevinolin were converted to their metabolically active forms (3). All-trans-[3H]farnesol (1.6 Ci/mmol) and all-trans-[3H]geranylgeraniol (1.6 Ci/mmol) were prepared as described (30). Farnesol and geranylgeraniol were the kind gift of T. Takigawa of the Kururay Co., Okayama, Japan, and phytol was from Aldrich. Protein molecular mass standards and Hyperfilm-MP were obtained from Amersham Pharmacia. All other chemicals were obtained from Sigma, and solvents were of reagent grade.
Labeling of Cotyledon Proteins with [3H]Mevalonate/Farnesol/Geranylgeraniol.
Cotyledons were labeled as described (28) in medium supplemented with 0.05% n-octyl β-d-glucopyranoside and 750 μCi of [3H]mevalonate (22.2 Ci/mmol), farnesol, or geranylgeraniol for 24 hr at room temperature under laboratory light. When studying the effects of chloramphenicol (CAP) (150 μM) or cycloheximide (CH) (150 μM), the samples were preincubated with the specified inhibitor for 1 hr before the addition of the [3H]mevalonate. Stock solutions of CAP and CH were stored in ethanol, and the final concentration of ethanol in the labeling medium, also present in control samples, was 0.1% (vol/vol).
[3H]Mevalonate Labeling of Mammalian Cells in Tissue Culture.
Mouse neuroblastoma N2a cells were maintained in MEM supplemented with 10% FCS in the presence of 100 units/ml of penicillin and 100 μg/ml of streptomycin. Labeling was performed with 70 μCi/ml of [3H]mevalonate (2.8 Ci/mmol) for 24 hr, after which the cells were washed with PBS and pelleted, and proteins were subjected to lipid extraction as described below.
Labeling of Rat Proteins in Vivo.
Rats were injected i.p. with 150 μCi of [3H]mevalonate (22.2 Ci/mmol) in 50 μl of 25 mM Tris⋅HCl, pH 7.5, every fifth hour. Each rat received a total of five injections. Twenty-five hours after the first injection, the rats were decapitated and the organs were homogenized in 1 ml of double-distilled water by using an Ultra-Turrax blender. Proteins were precipitated immediately in 10 ml of ice-cold acetone and extracted as described below.
Lipid Extraction and Protein Analysis.
After labeling, each sample was rapidly homogenized in 1.5 ml of ice-cold double-distilled water by using an Ultra-Turrax blender at 18,000 rpm for approximately 15 s. The homogenate was filtered immediately through one layer of nylon net (mesh size 20 μm) into 10 ml of ice-cold acetone to precipitate the proteins that were lipid-extracted and subjected to SDS/PAGE as described (3, 28, 31). The gels were fixed, impregnated with AMPLIFY (Amersham Pharmacia), dried, and exposed to film (Hyperfilm-MP, Amersham Pharmacia) at −80°C for as long as 3–39 weeks. Of the total radioactivity supplied 0.01–0.05% was incorporated into polypeptides.
Cellular Subfractionation of Spinach Seedlings.
Cellular subfractionation was performed according to the method of Bartlett et al. (32) from 3.0 g of 3H-mevalonate-labeled cotyledons. Seedlings were homogenized by using a mortar and pestle. The homogenate was filtered through one layer of nylon net and centrifuged at 1,000 × g for 5 min. The pellet, containing intact chloroplasts and nuclei, was resuspended in buffer and layered onto a Percoll-step gradient to purify intact chloroplasts (32). The supernatant from the initial centrifugation was recentrifuged for 3.5 min at 20,000 × g to pellet the crude mitochondrial fraction. The supernatant was centrifuged at 150,000 × g for 1.5 h to separate the microsomal and cytosolic fractions. The microsomes were washed in 0.15 M Tris (pH 8.5) to remove adsorbed cytosolic proteins. These techniques give intact organellar preparations essentially free of contamination. All pelleted fractions were resuspended in double-distilled water and immediately precipitated by the addition of 10% trichloroacetic acid. Proteins were delipidated with organic solvents, as described above, before electrophoretic analysis or scintillation counting.
Cleavage and Analysis of Protein-Bound Lipids. After the final lipid extraction, the proteins were subjected to methyl iodide, acidic (HCl) or alkaline (KOH) hydrolyses (33, 34). After hydrolysis, the isoprenoids were extracted with chloroform/methanol (9:1). Sixty percent of the total radioactivity from methyl iodide-treated proteins labeled in the presence of CAP and 40% of the total radioactivity from alkaline-treated proteins labeled in the presence of CH were recovered in the respective organic phases. Analysis of released isoprenoids was performed by RP-HPLC by using a C-18 column (Hewlett Packard Hypersil ODS 3 μm) and a two-step linear gradient, basically as described (4, 17). The absorbance at 210 nm and radioactivity of the eluate were monitored by using a UV detector and a radioactivity flow detector (Radiomatic Instruments, Tampa, FL).
For MS analysis, 6-week-old spinach was used after establishing that it possessed a similar set of prenylated polypeptides and an alkaline hydrolysis releasable isoprenoid, as did the spinach seedlings. Thylakoid proteins from 1 kg of spinach were subjected to alkaline hydrolysis, and the eluate corresponding to the elution time of phytol standard was collected. The solvents were evaporated, and the residue, in parallel with phytol standard, was converted to the acetate form by incubation with 15 μl of acetic anhydride and 10 μl of pyridine in 0.5 ml of benzene at room temperature for 2 hr. The yield of this reaction was close to 100% as estimated by TLC, developed in ethyl acetate/benzene (95:5, vol/vol).
GC was performed on a DB-5 column (30 meters, i.d. 0.2 μm), which was connected to a Nermag R10–10H mass analyzator. The temperature gradient started at 130°C for 1 min and was followed by an increase of 3°C/min up to 250°C. Thereafter, the increase was 10°C/min up to 290°C at which the temperature was kept for 20 min. Ionization was by electron impact at 70 eV and the scan range was 50 to 400. The samples were dissolved in hexane.
RESULTS
The Effect of Protein Synthesis Inhibitors on Protein Prenylation. It has been shown previously that cells derived from the cotyledons of spinach seedlings possess a complicated pattern of prenylated polypeptides (4). In a recent study, it was demonstrated that a number of these are associated with chloroplast membranes (28).
The effects of two specific protein synthesis inhibitors, CAP, which inhibits organellar protein translation, and CH, which inhibits translation of nuclear-encodes genes, on the pattern of prenylated proteins in spinach cotyledon tissue, were studied under in vivo conditions. The results revealed clear and differential effects of CAP and CH on the prenylation pattern as illustrated in Fig. 1A. Only very few of the total prenylated polypeptides could be detected after 3H-mevalonate labeling in the presence of 150 μM CAP. These CAP-insensitive-labeled polypeptides are found predominantly in the 21- to 26-kDa region. Additionally, two bands could be observed at molecular masses of about 15 and 60 kDa. Similar results were obtained with 350 μM streptomycin, another inhibitor of organellar protein synthesis (not shown). The CAP-insensitive bands, however, were found to be sensitive to labeling in the presence of 150 μM CH. The most strongly labeled polypeptides insensitive to CH have molecular masses of approximately 35 and 100 kDa. In addition, several labeled bands were seen in the entire 14-to 200-kDa molecular mass region. When combined, the sum of the CAP-sensitive and CH-sensitive bands comprise the total labeling pattern observed in cotyledons labeled in the absence of protein synthesis inhibitors.
Figure 1.

Effect of protein synthesis inhibitors on protein prenylation. (A) Protein prenylation in spinach cotyledons. Cotyledons were labeled with [3H]mevalonate. After the labeling period, the samples were homogenized, lipid-extracted, and analyzed by SDS/PAGE followed by fluorography. C, control; CH, prenylation in the presence of 150 μM CH, CAP, prenylation in the presence of 150 μM CAP. (B) Prenylated proteins in isolated, intact chloroplasts. Labeling as in A. (C) Prenylated proteins in: lane 1, neuroblastoma N2a cells; lane 2, rat kidney.
In a previous study where spinach seedlings with their roots excised were fed the [3H]mevalonate solution, it was concluded that the nuclei and the mitochondria were the most intensely labeled cellular fractions (3). For cotyledon leaves, however, this is not the case and the cellular subfraction that showed the highest labeling, on a protein basis, was the chloroplasts (Table 1). Furthermore, the yield of proteins in the chloroplast subfraction was a lot higher than that of the others, i.e. the vast majority of tritium-labeled, protein-bound isoprenoids were recovered from chloroplasts. The effect of CAP and CH on the in vivo protein prenylation pattern in various organelle fractions was investigated after subfractionation of the cotyledon cells. Notably, the presence of CAP during the labeling period completely abolished the prenylation of chloroplastic proteins (Fig. 1B, lane 2). On the other hand, CH did not appear to alter the pattern of prenylated proteins in this organelle on a qualitative basis (Fig. 1B, lane 3), although the overall labeling intensity was extensively increased on a protein basis. The prenylation of microsomal and cytosolic proteins was abolished by CH, but was not affected by CAP. The low yield of mitochondrial proteins (10–20 μg) excluded fluorography of the labeled proteins in this fraction. Nevertheless, estimations of the effect of CAP and CH on overall labeling in the mitochondrial fraction indicated that the isoprenoid modification of these proteins was sensitive to CH and not to CAP.
Table 1.
Prenylation of proteins in cotyledon subcellular fractions
| Fraction | Labeling, dpm per μg of protein |
|---|---|
| Chloroplasts | 2,020 ± 370 |
| Mitochondria | 1,380 ± 400 |
| Microsomes | 1,380 ± 490 |
| Cytosol | 400 ± 160 |
Cotyledons were labelled with [3H]mevalonate and subfractionated. After lipid extraction and solubilization, the radiolabel associated with the proteins of each fraction was determined by scintillation counting and protein estimation. Data shown represent the mean values of four fractionation experiments ±SE.
Comparison with 3H-Mevalonate Labeling of Mammalian Cells.
To readily compare the pattern of prenylated polypeptides in mammalian cells and spinach cotyledons, N2a neuroblastoma cells in tissue culture and rats were metabolically labeled with [3H]mevalonate. This resulted in the prenylation of several proteins, and the pattern of labeled polypeptides is shown in Fig. 1C. The pattern of prenylated polypeptides in neuroblastoma cells and rat kidney closely resembled that observed in the CAP-treated spinach cotyledons (Fig. 1A, lane 1).
Hydrolysis of Prenylated Proteins.
Methyl iodide hydrolysis is a mild cleavage procedure specific for thioether bonds (35). All prenylated proteins identified to date carry thioether-bound isoprenoids on C-terminal cysteine residues. The isoprenoids farnesol and geranylgeraniol were released by hydrolysis of cotyledon polypeptides labeled in the presence of CAP (Fig. 2A), demonstrating that the CH-sensitive polypeptides are prenylated with these isoprenoids bound to cysteine residues via thioether linkages. Remarkably, no isoprenoids were released from CH-treated, 3H-mevalonate-labeled material (Fig. 2A, Inset). This finding indicates that a different type of prenylation, not involving isoprenoids thioether-linked to cysteine residues, takes place in chloroplasts. Upon alkaline hydrolysis of the polypeptides that were not prenylated in the presence of CAP, a single isoprenoid was released. This isoprenoid corresponded to the elution time of phytol (Fig. 2B), and it also comigrated with phytol upon TLC analysis (not shown). On the other hand, no isoprenoids were released after acidic hydrolysis of polypeptides labeled in the presence of CAP (not shown).
Figure 2.

HPLC analysis of covalently bound isoprenoids. Cotyledons were labeled with [3H]mevalonate in the presence of CAP (150 μM). After labeling, proteins were lipid-extracted and subjected to hydrolysis. The released isoprenoids were analyzed by RP- HPLC, and the resulting chromatograms are shown. (A) Methyl iodide hydrolysis. (Inset) Isoprenoids released from cotyledon proteins labeled in the presence of CH (150 μM). (B) Alkaline hydrolysis of proteins labeled in the presence of CH. The elution of farnesol (F), geranylgeraniol (GG), and phytol (PH) are indicated.
MS Analysis of the Released Isoprenoid.
To further establish the identity of the isoprenoid modifying the chloroplastic polypeptides, the alkaline hydrolyzable isoprenoid from thylakoids from 1 kg of spinach was derivatized to its acetate form. When phytyl acetate standard was analyzed on GC-MS, the compound eluting at 36.2 min demonstrated a fragmentation pattern typical of phytol (Fig. 3A). The peak at m/z 278 corresponds to phytyl acetate that has lost acetate and two hydrogens. The isoprenoid from the thylakoid sample also contained a compound that eluted at 36.2 min, and its fragmentation pattern was almost identical to that of the phytyl acetate standard (Fig. 3B). This finding confirms that phytol is modifying the polypeptides whose prenylation is sensitive to CAP.
Figure 3.

Identification of phytol by MS. Lipid-extracted thylakoid proteins from 1 kg of spinach were subjected to alkaline hydrolysis followed by RP-HPLC. The eluate corresponding to the elusion time of phytol standard was collected, the solvents were evaporated, and the residue was derivatized to the acetate form. The resulting product was analyzed on GC-MS. (A, Upper) GC chromatogram of phytyl acetate standard. (A, Lower) The fragmentation pattern of phytyl acetate (elution time 36.2 min). (B, Upper) GC chromatogram of the spinach isoprenoid. (B, Lower) The fragmentation pattern of the compound eluting at 36.24 min.
In Vivo Metabolic Labeling of Cotyledons with Farnesol and Geranylgeraniol.
To investigate the nature of this nonthioether-bound phytol of chloroplastic proteins, metabolic labeling of cotyledons was performed with all-trans-[3H]farnesol or all-trans-[3H]geranylgeraniol. These isoprenoids both are dephosphorylated forms of mevalonate pathway intermediates/end-products. In vivo labeling of cotyledons with [3H]geranylgeraniol resulted in intense incorporation of radiolabel into polypeptides of the 21- to 26-kDa molecular mass region (Fig. 4). The pattern of the prenylated polypeptides was similar to that of cotyledons labeled in the presence of CAP (Fig. 1A, lane 1), representing the extra-chloroplast labeling. However, metabolic labeling with [3H]farnesol (Fig. 4) yielded a protein pattern similar to that observed after mevalonate labeling. This finding demonstrates that the phytol bound to the chloroplastic polypeptides via the novel type of linkage can readily be derived from exogenously supplied farnesol.
Figure 4.
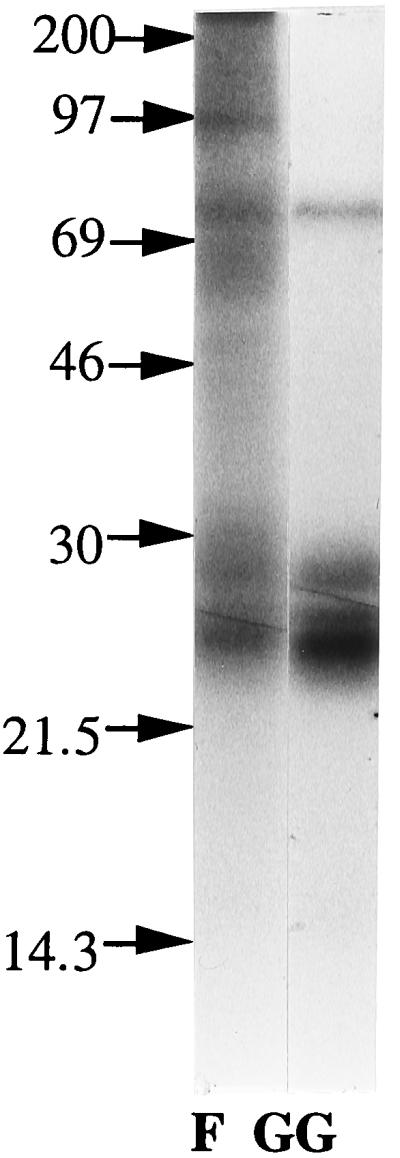
In vivo labeling of cotyledon proteins with farnesol or geranylgeraniol. Labeling was performed with either [3H]geranylgeraniol (GG) or [3H]farnesol (F). After labeling, proteins from homogenates were precipitated, lipid-extracted, and subjected to SDS/PAGE and fluorography.
DISCUSSION
The use of an in vivo labeling method has enabled extensive analysis of protein prenylation in plant tissue (3, 4, 28). The present study used this method to investigate the genetic origin of the proteins/enzymes involved in plant protein prenylation, through the use of the specific protein synthesis inhibitors CH and CAP. It was demonstrated that prenylation in plant tissues can be divided into two categories (Table 2), one of which is the conventional modification involving thioether-linked farnesyl and geranylgeranyl groups with a pattern of prenylated proteins equivalent to that observed in mammalian cells. This type of prenylation depended on nuclear gene products. The second category of protein prenylation displayed several unique features (Table 2). It was found to be confined to chloroplasts and dependent on gene products of the plastid genome. Another remarkable observation was that this type of prenylation did not involve a thioether bond, but another, as-yet-uncharacterized binding mechanism.
Table 2.
Characteristics of protein prenylation in spinach cotyledons
| Feature | Type 1 | Type 2 |
|---|---|---|
| Main prenylated proteins | 21–26 kDa | 30–55 kDa and 100 kDa |
| Inhibitor sensitivity | CAP-insensitive, CH-sensitive | CAP-sensitive, CH-insensitive |
| Cellular location | Extra chloroplast | Chloroplast |
| Relation to other systems | Analogous to yeast and mammalian prenylation | Unique to plants |
| Type of bond | Thioether linkage | Unknown, nonthioether linkage |
| Prenylating group | Farnesyl and geranylgeranyl | Phytyl |
Protein prenylation requires the presence of both isoprenoid and protein substrates as well as prenylating enzymes. It is not very likely that the effects described above can be explained at the enzyme level. It would require an enzyme with an extremely short turnover time to be depleted in 1 hr, which was the time of the preincubation with the protein synthesis inhibitors before the addition of the labeled substrates. Furthermore, the chloroplast genome has not been reported to encode any prenyl protein transferases or other isoprenoid-metabolizing enzymes. However, several ORFs still have to be functionally assigned. It seems more likely that the effect of CAP on chloroplast prenylation should be sought at the protein substrate level. Yet, the relatively high number of prenylated chloroplastic polypeptides, and their uniform response to CAP, is somewhat puzzling. If all of the chloroplastic polypeptides that are not prenylated in the presence of CAP are plastid-encoded, this would mean that a substantial fraction of the approximately 120 gene products encoded by the chloroplast genome harbors this modification. Another possible explanation is that the presence of CAP results in the depletion of short-lived plastid-encoded isoprenoid carriers, which would result in a shortage of isoprenoid substrates inside the chloroplasts. The revelation of the mechanism behind the CAP effect on chloroplastic protein prenylation will require some further experimentation.
The data that demonstrate that the labeling intensity in chloroplasts was increased after CH treatment could be the result of an internal mevalonate pool of higher specific activity, after depletion of the short-lived enzyme hydroxymethyl glutaryl-CoA reductase (36). Because the prenyl transferase(s) involved in modifying the proteins whose prenylation is CAP sensitive is most certainly nuclear encoded, it should display a fairly slow turnover rate.
The well-characterized and gentle hydrolysis method using methyl iodide, specific for thioether bonds (35), did not result in the release any isoprenoids from the CAP-sensitive proteins. In the RP-HPLC system used, isoprenoids at least as long as 23 units can be observed. Evidently plants possess the capability for an additional prenylation pathway not observed to date in mammalian and yeast studies. The nature of this novel prenylation awaits characterization; however, the identity of the isoprenoid used for prenylation is phytol. This prenyl alcohol was released from the proteins after alkaline hydrolysis and its identity confirmed by GC-MS analysis.
Covalent bonds between a prenyl alcohol and an amino group have been described for isopentenyladenosine in yeast and mammalian tissue, where an isopentenyl moiety is attached to N6 of adenine via an amine bond (37). In addition to isopentenyladenosine, plant tRNAs can contain two hydroxylated derivatives of that species (38). It is distinctly possible that the proteins whose prenylation is sensitive to CAP contain phytol bound to amino group side chains.
The fact that plant prenylated proteins can be labeled from farnesol or geranylgeraniol strongly suggests that plants possess prenyl alcohol phosphorylating systems. Such systems recently have been reported in bacteria (25) and mammals (26, 27). Furthermore, the results presented here demonstrate that prenyl alcohols can be taken up and metabolized by intact plant cells.
It is interesting that the type of prenylation described here was not readily achieved when all-trans-geranylgeraniol was used as a substrate. However, all-trans-farnesol was shown to be an excellent precursor for the phytyl modification. This finding implied that the modifying moiety could be cis-phytol. Surprisingly, when the released phytol was subjected to high-performance TLC analysis, it contained both the trans and the cis isomer in a ratio of about 4 to 1. At present, it is not known if this unexpected distribution was a result of rearrangement occurring during the alkaline hydrolysis.
That farnesol was readily used as a precursor of the phytyl groups also can be discussed in relation to the recent line of investigations suggesting that chloroplasts of higher plants do not possess the mevalonate/acetate pathway (39). If this claim holds true, the data presented here strongly favor an import mechanism for cytoplasmically synthesized farnesyl pyrophosphate, but not geranylgeranyl pyrophosphate, into the chloroplast. Such an import mechanism has been described for the C5 isoprenoid isopentenyl pyrophosphate (40), and geranylgeranyl pyrophosphate synthase is localized in the chloroplast in several plant species (41, 42).
Current theories regarding the function of protein prenylation include increasing hydrophobicity thereby aiding membrane association and recognition signals for protein–protein interactions which also may well apply to the chloroplastic prenylated proteins as they have been shown to be components of membrane protein complexes (28). These prenylated proteins seem to be of low abundance as they could not be visualized in SDS/PAGE gels stained with Coomassie blue. In addition, these proteins presumably are long-lived because relatively high amount of radiolabel was incorporated during the 24-hr labeling period. They may represent proteins involved in the regulation of chloroplast functions, catabolism, or biosynthesis as well as stress responses.
Taken together, the data presented here provide clear evidence for a novel type of protein prenylation in plants, which depends on products of the chloroplast genome. The existence of such a pathway for protein prenylation opens up an area for exciting future studies.
Acknowledgments
We thank Profs. Lars Ernster and Itzhak Ohad and Dr. Cathie Shipton for constructive discussion and helpful advice. We are also grateful to Dr. Ewa Swiezewska for syntheses of tritiated farnesol and geranylgeraniol, Elisabeth Peterson for assistance with the tissue cultures, and Gunvor Alvelius for assistance with MS analyses. Financial support was provided by grants from the Swedish Council for Forestry and Agricultural Research and the Swedish Cancer Society.
ABBREVIATIONS
- CAP
chloramphenicol
- CH
cycloheximide
Footnotes
This paper was submitted directly (Track II) to the Proceedings Office.
References
- 1.Casey P J. Science. 1995;268:221–225. doi: 10.1126/science.7716512. [DOI] [PubMed] [Google Scholar]
- 2.Siddiqui A A, Garland J R, Dalton M B, Sinensky M. J Biol Chem. 1998;273:3712–3717. doi: 10.1074/jbc.273.6.3712. [DOI] [PubMed] [Google Scholar]
- 3.Shipton C A, Parmryd I, Swiezewska E, Andersson B, Dallner G. J Biol Chem. 1995;270:566–572. doi: 10.1074/jbc.270.2.566. [DOI] [PubMed] [Google Scholar]
- 4.Parmryd I, Shipton C A, Andersson B, Dallner G. Arch Biochem Biophys. 1997;339:73–78. doi: 10.1006/abbi.1996.9816. [DOI] [PubMed] [Google Scholar]
- 5.Schmidt R A, Schneider C J, Glomset J A. J Biol Chem. 1984;259:10175–10180. [PubMed] [Google Scholar]
- 6.Randall S K, Marshall M S, Crowell D N. Plant Cell. 1993;5:433–442. doi: 10.1105/tpc.5.4.433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Inglese J, Glickman J F, Lorenz W, Caron M G, Lefkowitz R J. J Biol Chem. 1992;267:1422–1425. [PubMed] [Google Scholar]
- 8.Hancock J F, Magee A I, Childs J E, Marshall C J. Cell. 1989;57:1167–1177. doi: 10.1016/0092-8674(89)90054-8. [DOI] [PubMed] [Google Scholar]
- 9.Schafer W R, Kim R, Sterne R, Thorner J, Kim S-H, Rine J. Science. 1989;245:379–385. doi: 10.1126/science.2569235. [DOI] [PubMed] [Google Scholar]
- 10.Wolda S L, Glomset J A. J Biol Chem. 1988;263:5997–6000. [PubMed] [Google Scholar]
- 11.Zhu J-K, Bressan R A, Hasegawa P M. Proc Natl Acad Sci USA. 1993;90:8557–8561. doi: 10.1073/pnas.90.18.8557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Saalbach G, Christov V. Plant Mol Biol. 1994;24:969–972. doi: 10.1007/BF00014451. [DOI] [PubMed] [Google Scholar]
- 13.Rodriguez-Concepción M, Yalovsky S, Zik M, Fromm H, Gruissem W. EMBO J. 1999;18:1996–2007. doi: 10.1093/emboj/18.7.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Reiss Y, Goldstein J L, Seabra M C, Casey P J, Brown M S. Cell. 1990;62:81–88. doi: 10.1016/0092-8674(90)90242-7. [DOI] [PubMed] [Google Scholar]
- 15.Moores S L, Shaber M D, Mosser S D, Rands E, O’Hara M B, Garsky V M, Marshall M S, Pompliano D L, Gibbs J B. J Biol Chem. 1991;266:14603–14610. [PubMed] [Google Scholar]
- 16.Seabra M C, Brown M S, Slaughter C A, Südhof T C, Goldstein J L. Cell. 1992;70:1049–1057. doi: 10.1016/0092-8674(92)90253-9. [DOI] [PubMed] [Google Scholar]
- 17.Runquist M, Ericsson J, Thelin A, Chojnacki T, Dallner G. J Biol Chem. 1994;269:5804–5809. [PubMed] [Google Scholar]
- 18.Runquist M, Ericsson J, Thelin A, Chojnacki T, Dallner G. Biochem Biophys Res Commun. 1992;186:157–165. doi: 10.1016/s0006-291x(05)80788-0. [DOI] [PubMed] [Google Scholar]
- 19.Gray J C. Adv Bot Res. 1987;14:25–91. [Google Scholar]
- 20.Sakagami Y, Yoshida M, Isogai A, Suzuki A. Science. 1981;212:1525–1527. doi: 10.1126/science.212.4502.1525. [DOI] [PubMed] [Google Scholar]
- 21.Hjertman M, Wejde J, Dricu A, Carlberg M, Griffiths W J, Sjövall J, Larsson O. FEBS Lett. 1997;416:235–238. doi: 10.1016/s0014-5793(97)01208-8. [DOI] [PubMed] [Google Scholar]
- 22.Parmryd I, Dallner G. Arch Biochem Biophys. 1999;364:153–160. doi: 10.1006/abbi.1999.1134. [DOI] [PubMed] [Google Scholar]
- 23.Crick D C, Andres D A, Waechter C J. Biochem Biophys Res Commun. 1995;211:590–599. doi: 10.1006/bbrc.1995.1854. [DOI] [PubMed] [Google Scholar]
- 24.Crick D C, Waechter C J, Andres D A. Biochem Biophys Res Commun. 1994;205:955–961. doi: 10.1006/bbrc.1994.2758. [DOI] [PubMed] [Google Scholar]
- 25.Inoue H, Korenaga T, Sagami H, Koyama T, Ogura K. Biochem Biophys Res Commun. 1994;200:1036–1041. doi: 10.1006/bbrc.1994.1554. [DOI] [PubMed] [Google Scholar]
- 26.Westfall D, Aboushadi N, Shackelford J E, Krisans S K. Biochem Biophys Res Commun. 1997;230:562–568. doi: 10.1006/bbrc.1996.6014. [DOI] [PubMed] [Google Scholar]
- 27.Bentinger M, Grünler J, Peterson E, Swiezewska E, Dallner G. Arch Biochem Biophys. 1998;353:191–198. doi: 10.1006/abbi.1998.0611. [DOI] [PubMed] [Google Scholar]
- 28.Parmryd I, Shipton C A, Swiezewska E, Dallner G, Andersson B. FEBS Lett. 1997;414:527–531. doi: 10.1016/s0014-5793(97)01063-6. [DOI] [PubMed] [Google Scholar]
- 29.Keller R K. J Biol Chem. 1986;261:12053–12059. [PubMed] [Google Scholar]
- 30.Keenan R W, Kruczek M E. Biochemistry. 1976;15:1586–1591. doi: 10.1021/bi00652a032. [DOI] [PubMed] [Google Scholar]
- 31.Laemmli U K. Nature (London) 1970;227:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- 32.Bartlett S G, Grossman A R, Chua N-H. In: Methods in Chloroplast Molecular Biology. Edelman M, Hallick R B, Chua N-H, editors. Amsterdam: Elsevier; 1982. pp. 1081–1092. [Google Scholar]
- 33.Casey P J, Solski P A, Der C J, Buss J E. Proc Natl Acad Sci USA. 1989;86:8323–8327. doi: 10.1073/pnas.86.21.8323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Thelin A, Löw P, Chojnacki T, Dallner G. Eur J Biochem. 1991;195:755–761. doi: 10.1111/j.1432-1033.1991.tb15763.x. [DOI] [PubMed] [Google Scholar]
- 35.Kamiya Y, Sakurai A, Tamura S, Takahashi N. Agric Biol Chem. 1979;43:1049–1053. [Google Scholar]
- 36.Edwards P A, Lan S, Fogelman A M. J Biol Chem. 1983;258:10219–10222. [PubMed] [Google Scholar]
- 37.Robins M J, Hall R H, Thedford R. Biochemistry. 1967;6:1837–1848. doi: 10.1021/bi00858a035. [DOI] [PubMed] [Google Scholar]
- 38.Hall R H, Csonka L, David H, McLennan B. Science. 1967;156:69–71. doi: 10.1126/science.156.3771.69. [DOI] [PubMed] [Google Scholar]
- 39.Lichtenthaler H K, Schwender J, Disch A, Rohmer M. FEBS Lett. 1997;400:271–274. doi: 10.1016/s0014-5793(96)01404-4. [DOI] [PubMed] [Google Scholar]
- 40.Soler E, Clastre M, Bantignies B, Marigo G, Ambid C. Planta. 1993;191:324–329. [Google Scholar]
- 41.Kuntz M, Romer S, Suire C, Hugueney P, Weil J H, Schantz R, Camara B. Plant J. 1992;2:25–34. doi: 10.1111/j.1365-313x.1992.00025.x. [DOI] [PubMed] [Google Scholar]
- 42.Scolnik P A, Bartley G E. Plant Physiol. 1994;104:1469–1470. doi: 10.1104/pp.104.4.1469. [DOI] [PMC free article] [PubMed] [Google Scholar]