

Abstract
Organisms regularly modify local resource distributions, influencing both their ecosystems and the evolution of traits whose fitness depends on such alterable sources of natural selection in environments. We call these processes niche construction. We explore the evolutionary consequences of niche construction using a two-locus population genetic model, which extends earlier analyses by allowing resource distributions to be influenced both by niche construction and by independent processes of renewal and depletion. The analysis confirms that niche construction can be a potent evolutionary agent by generating selection that leads to the fixation of otherwise deleterious alleles, supporting stable polymorphisms where none are expected, eliminating what would otherwise be stable polymorphisms, and generating unusual evolutionary dynamics. Even small amounts of niche construction, or niche construction that only weakly affects resource dynamics, can significantly alter both ecological and evolutionary patterns.
Keywords: ecosystem engineering, adaptation
There is increasing recognition that all organisms modify their environments (1), a process that we call “niche construction” (2) but is elsewhere described as “ecosystem engineering” (3). Such modifications can have profound effects on the distribution and abundance of organisms, the influence of keystone species, the control of energy and material flows, residence and return times, ecosystem resilience, and specific trophic relationships (3–6). The consequences of environment modification by organisms, however, are not restricted to ecology, and organisms can affect both their own and each other’s evolution by modifying sources of natural selection in their environments (2, 7). We have argued that niche construction is responsible for hitherto-neglected forms of feedback in evolution, whereby legacies of ancestrally modified natural selection pressures affect the subsequent evolution of later generations of organisms (7–9).
There are numerous examples of organisms choosing or changing their habitats or constructing artifacts, leading to an evolutionary, as well as an ecological, response (see refs. 2, 3, and 7–9). For instance, orb-web spiders construct webs, which have led to the subsequent evolution of camouflage, defense, and communication behavior on the web (10). Similarly, ants, bees, wasps, and termites construct nests that are themselves the source of selection for many nest regulatory, maintenance, and defense behavior patterns. For example, many ant and termite species regulate temperature by plugging nest entrances at night or in the cold, by adjusting the height or shape of their mounds to optimize the intake of the sun’s rays, or by carrying their brood around their nest to the place with the optimal temperature and humidity for the brood’s development (11, 12). The construction of artifacts is equally common among vertebrates. Many mammals (including badgers, gophers, ground squirrels, hedgehogs, marmots, moles, mole rats, opossum, prairie dogs, rabbits, and rats) construct burrow systems, some with underground passages, interconnected chambers, and multiple entrances (13). Here, too, there is evidence that burrow defense, maintenance, and regulation behaviors have evolved in response to selection pressures that were initiated by the construction of the burrow (11, 13).
Most cases of niche construction, however, do not involve the building of artifacts but merely the selection or modification of habitats. For example, as a result of the accumulated effects of past generations of earthworm niche construction, present generations of earthworms inhabit radically altered environments where they are exposed to modified selection pressures (14, 15). Previously, we described this legacy of modified selection pressures as an “ecological inheritance.” Nor is niche construction confined to animals. Plants change the chemical nature, the pattern of nutrient cycling, the temperature, the humidity, the fertility, the acidity, and the salinity of their soils and the patterns of light and shade in their habitats (16–19). For instance, pine and chaparral species increase the likelihood of forest fires by accumulating oils or litter, with the probable evolutionary consequence of having evolved a resistance to fire and in some species a dependency on it (20–22).
We have explored the dynamics of the joint evolution of environment-altering, niche-constructing traits in organisms and other traits whose fitness depends on feedback from natural selection in environments that can be altered by niche construction (9). Our previous analysis used a two-locus population-genetics model, with alleles at one locus influencing the population’s niche construction by affecting the amount of a key resource in the environment and with the amount of the resource influencing the contribution to fitness of genotypes at a second locus. Our analysis suggested that the changes that organisms bring about in their own selective environments can be an important source of modified natural-selection pressures and can generate some unusual evolutionary outcomes. For example, niche construction can cause evolutionary inertia and momentum, it can lead to the fixation of otherwise deleterious alleles, it can support stable polymorphisms where none are expected, and it can eliminate what would otherwise be stable polymorphisms.
Our earlier model assumed that the frequency of the key resource in the environment depends solely on niche construction. This assumption is clearly unrealistic in many cases, because the distributions of ecological resources often depend on processes that are independent of the population (6, 23). Here we present findings of an analysis with a more complex treatment of the frequency of the resource in which it is influenced by varying mixes of a population’s niche-construction and other independent environmental processes of renewal or depletion.
THE MODEL
Consider an isolated population of randomly mating, diploid individuals, defined at two diallelic loci, E (with alleles E and e) and A (with alleles A and a). In generation t, the frequencies of the four gametes (EA, Ea, eA, and ea) are given by x1t − x4t, respectively, so that the frequencies of alleles E and A are pt = x1t + x2t, and qt = x1t + x3t. We assume that the population’s capacity for niche construction is influenced by the frequency of alleles at the E locus. Genotypes at the A locus make contributions to the two-locus fitnesses that are functions of the frequency with which a resource (R) is encountered in their environment (where 0 < R < 1). This frequency is a function of the amount of niche construction over n generations (that is, the frequencies of allele E) as well as other independent processes of resource recovery or resource dissipation. We define as positive niche construction phenotypic activities that increase the fitness of the niche-constructing organism, whereas negative niche construction refers to niche-constructing activities that reduce fitness. In our model, positive and negative niche construction, respectively, refer to processes that increase and deplete the frequency of a valuable resource. This treatment of the interaction between the population and the resource is still extremely simplified. A more realistic treatment would involve general distributions of the resource, more complex ecological dynamics of resource and population, and ecological models that take the density of niche constructors into account (6, 26, 27). The particular model we use has properties that are likely to exemplify a broad class of models, and our assumptions concerning resource dynamics are made largely on the basis of analytical convenience.
In each generation, the amount of the resource at time t is given by
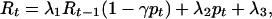 |
1 |
where λ1 is a coefficient that determines the degree of independent depletion (if λ1 = 1, there is no independent depletion), λ2 is a coefficient that determines the effect of positive niche construction (if λ2 = 0, there is no positive niche construction influencing the amount of the resource), λ3 is a coefficient that determines the degree of independent renewal (if λ3 = 0, there is no independent renewal influencing the amount of the resource), and γ is a coefficient that determines the effect of negative niche construction (if γ = 0, there is no negative niche construction influencing the amount of the resource). We assume 0 < λ1, λ2, λ3, γ < 1 and λ1 + λ2 + λ3 ≤ 1. The term λ1Rt−1 represents the proportion of the resource that remains from the previous generation after independent depletion, whereas (1 − γpt) represents any further decay in R due to negative niche construction.
We consider four cases of this general model representing positive or negative niche construction with independent renewal or depletion. Genotypic fitnesses (given in Table 1) are assumed to be functions of a fixed viability component and a frequency-dependent viability component. The fixed components (given by the αi and βi terms) represent selection from the external environment, that is, independent of niche construction. These are the fitnesses of genotypes in the standard two-locus multiplicative viability model. The frequency-dependent components of the contribution to fitness of genotypes AA, Aa, and aa are functions of R, 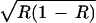 and 1 − R, respectively, chosen so that allele A will be favored by this component of selection when the resource is common and allele a when it is rare. Thus, the frequency-dependent components are functions of the frequency of the resource, modified by niche construction. The coefficient of proportionality (ɛ) determines the strength of the frequency-dependent component of selection relative to the fixed-fitness component (−1 < ɛ < 1). Positive values of ɛ represent cases where an increase in the amount of resource results in an increment in the fitness of genotypes containing allele A, whereas negative values of ɛ represent cases where an increase in R favors a.
and 1 − R, respectively, chosen so that allele A will be favored by this component of selection when the resource is common and allele a when it is rare. Thus, the frequency-dependent components are functions of the frequency of the resource, modified by niche construction. The coefficient of proportionality (ɛ) determines the strength of the frequency-dependent component of selection relative to the fixed-fitness component (−1 < ɛ < 1). Positive values of ɛ represent cases where an increase in the amount of resource results in an increment in the fitness of genotypes containing allele A, whereas negative values of ɛ represent cases where an increase in R favors a.
Table 1.
Genotypic fitnesses
| Locus | EE | Ee | ee |
|---|---|---|---|
| AA | w11 = α1α2 + ɛR | w12 = α2 + ɛR | w13 = β1α2 + ɛR |
| Aa |
w21 = α1 + ɛ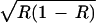
|
w22 = 1 + ɛ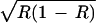
|
w23 = β1 + ɛ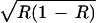
|
| aa | w31 = α1β2 + ɛ(1 − R) | w32 = β2 + ɛ(1 − R) | w33 = β1β2 + ɛ(1 − R) |
The genotype fitnesses wij in Table 1 give rise to the standard gametic recursions
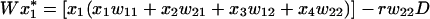 |
2a |
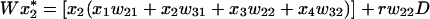 |
2b |
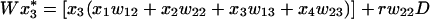 |
2c |
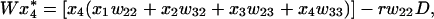 |
2d |
where r is the recombination rate, D = x1x4 − x2x3 is the linkage disequilibrium between the loci, and W is given by the sum of the right-hand sides of Eqs. 2a–2d.
RESULTS
We consider the effects of niche construction under four regimes of external selection: no external selection, external selection acting only at the A locus, external selection acting only on the E locus, and external selection generating heterozygote advantage.
No External Selection.
First, we consider the dynamics of the system when there is no selection acting aside from that generated by the resource (αi = βi = 1). With only positive niche construction (Rt = λ1Rt−1 + λ2pt + λ3, γ = 0), for frequencies of the allele E from p = 0 to p = 1, the corresponding values of R range between 0 and 1, depending on λ1, λ2, and λ3. At equilibrium, from Eq. 1, the value of R as a function of p is given by
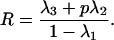 |
3a |
With only negative niche construction (Rt = λ1Rt−1(1 − γpt) + λ3, λ2 = 0), for values of p increasing from 0 to 1, R decreases in an interval of values contained in 0–1, depending on the values of λ1, λ3, and γ. The equilibrium value of R for any given value of p is given by
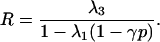 |
3b |
It is a feature of the frequency-dependent fitnesses in Table 1 that for positive values of ɛ, selection favors a when R < 1/2, A when R > 1/2, with no selection on A when R = 1/2, while for negative ɛ, selection favors a when R > 1/2 and A when R < 1/2. For instance, in Figure 1a, when p = 0, the balance of independent renewal and depletion of resources leaves R at an equilibrium value of 1/3. Any positive niche construction (p > 0) increases the amount of the resource, with R reaching 0.5 and selection switching to favor allele A at p = 0.25.
Figure 1.

(a) Positive niche construction, independent renewal, and no external selection (Rt = λ1Rt−1 + λ2pt + λ3, γ = 0, α1 = β1 = α2 = β2 = 1, r = 1/2, n = 1). Here, the only selection is that generated by the resource. The arrows represent the trajectories of a population, the heavy line represents stable equilibria, and the dashed line represents unstable equilibria. The direction of selection generated by the resource switches at R = 1/2. In b–d, with independent renewal and external selection favouring allele A, niche construction generates selection that may oppose the external selection, taking populations to alternative equilibria and generating polymorphisms. b and c show positive niche construction (Rt = λ1Rt−1 + λ2pt + λ3, γ = 0), with weak selection (α1 = β1 = 1, α2 = 1.01, β2 = 0.99, r = 1/2) (b), and strong selection (α1 = β1 = 1, α2 = 1.1, β2 = 0.9, r = 1/2) (c). d shows negative niche construction, independent renewal, and external selection favoring allele A (Rt = λ1Rt−1(1 − γpt) + λ3, λ2 = 0, α1 = β1 = 1, α2 = 1.1, β2 = 0.9, r = 1/2). When selection favors allele A and ɛ is negative, positive niche construction (γ = 0) (e) and negative niche construction (λ2 = 0) (f) generate polymorphisms (α1 = β1 = 1, α2 = 1.1, β2 = 0.9, r = 1/2).
External Selection at the A Locus.
We now consider the case where there is external selection at the A locus only (α1 = β1 = 1, α2 ≠ 1, and/or β2 ≠ 1). Because there is no selection on E, by summing Eqs. 2a and 2c we derive a recursion for the frequency q of A, whose equilibria are given by q̂ = 0, q̂ = 1, or the polymorphism
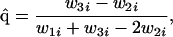 |
4 |
for i = 1–3 in Table 1. If we assume that the q = 1 boundary becomes unstable when w1i = w2i, then we may solve this equality for R and derive the range or ranges of R values compatible with the stability of q̂ = 1, namely w1i > w2i so that
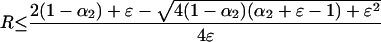 |
5a |
and
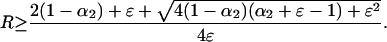 |
5b |
A similar analysis for q̂ = 0, when w3i > w2i, yields
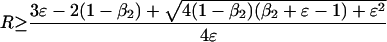 |
5c |
and
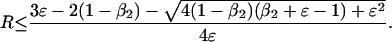 |
5d |
For positive values of ɛ, if R is sufficiently small, the selection generated by the resource will be sufficiently strong to counteract the external selection and take the population to the q = 0 boundary. With positive niche construction and weak external selection (β2 close to 1, e.g., Fig. 1b), Eq. 4 gives q as an approximately linear function of p and R so that regions of the q = 0 and q = 1 boundaries may be simultaneously stable for a range of values of p. For stronger external selection (e.g., Fig. 1c), q becomes more curvilinear as a function of p and R, and a becomes fixed for a smaller region of the parameter space. Fig. 1d illustrates the effects of negative niche construction, with the a fixation boundary being stable from some starting conditions. Since fixation on a only occurs when R is small, independent renewal and positive niche construction make it less likely that the frequency-dependent selection will generate the kind of counterselection illustrated in Fig. 1 b–d, whereas independent depletion and negative niche construction both increase this likelihood.
Negative values of ɛ render polymorphic equilibria at A possible. Examples with positive and negative niche construction are shown in Fig. 1 e and f, respectively. When the external selection is weak (α2, β2 close to 1), the line of polymorphic equilibria, also given by Eq. 4, approximates a straight line unless the impact of the niche construction is small, so that it only weakly influences the amount of the resource. When the external selection is strong, polymorphic equilibria are possible for large values of R for which Eq. 5 is not satisfied. Hence, independent renewal and positive niche construction increase the likelihood that positive niche construction will generate counterselection and create polymorphisms, whereas independent depletion and negative niche construction reduce this likelihood.
Where niche construction has a small impact on the amount of the resource (λ2 or γ are small, as in Fig. 1 e or f), there is a relatively large range of values of p for which the frequency-dependent selection generated by the resource can counteract the external source of selection. In the presence of independent renewal and depletion, even low frequencies of niche construction (small p) or a weak impact of niche construction on the amount of the resource (small λ2 or γ), can generate a switch in the selection generated by the resource such that allele a is selected to fixation (Fig. 1 b–d) or stable polymorphisms are possible (Fig. 1 e and f). The qualitative behavior of this system is little affected by the amount of recombination (r) when r is moderate to large.
External selection at the E locus.
External selection only at the E locus represents selection on a niche-constructing activity. When E is favoured (α1 > 1 > β1, α2 = β2 = 1), positive values of ɛ result in convergence to a single equilibrium point with E and A fixed (x1 = 1) if R > 1/2 at p = 1, with E and a fixed (x2 = 1) if R < 1/2 at p = 1 and with a range of polymorphic equilibria when R = 1/2. Similarly, negative values of ɛ result in the fixation of E and a(x2 = 1) if R > 1/2 at p = 1, with E and A fixed (x1 = 1) if R < 1/2 at p = 1. The reverse pattern is found if e is favored by selection. Provided that all alleles are initially present, numerical analysis has established that under these conditions, a population will always converge to these equilibria. Independent renewal increases the likelihood that selection at the E locus favoring positive or negative niche construction generates a corresponding selection pressure favoring A and decreases the likelihood that positive niche construction favors a.
Unusual evolutionary dynamics may occur when the degree of dominance in genotype fitnesses is a function of the amount of the resource present. For instance, when selection favors E and a positive niche-constructing behavior spreads, R will increase in value. We might predict that as the impact of niche construction on the resource (λ2) increases, so will the rate of selection of the favored allele A. For instance, if λ1 = 0.5 and we increase λ2 from 0.4 to 0.5, there is a dramatic deceleration in the rate at which allele A approaches fixation. This occurs because when E is fixed, R has increased from 0.8, where the fitness difference between genotype AaEE and genotype aaEE is significant, to 1.0, where the fitness difference is tiny.
Laland et al. (9) found that there can be a time lag between the spread of a niche-constructing activity, represented by a change in the frequency of alleles at the E locus, and the response to the selection generated at the A locus. In our original model, R was computed as a weighted average of the previous n generations of niche construction. Here, we have extended our analysis to consider cases in which R is an accumulatory function that can only increase as n becomes larger. One consequence is that this extended model typically does not generate time lags in the response to selection. However, the model does generate time lags (i) if there is a primacy effect, that is, earlier generations contribute disproportionately to the amount of the resource present in the environment (see ref. 9) and (ii) if we impose the constraint λ2 = 1/n, which transforms R into a weighted average of the previous n generations, analogous to the simpler model. This suggests that there are biologically meaningful circumstances under which we might predict the evolutionary momentum and inertia effects identified by our earlier analysis. Again, the qualitative behavior is little affected by r when recombination is moderate-to-loose.
Heterozygote Advantage.
There may be a polymorphic equilibrium with D = 0 at which the frequencies of E and A are given by
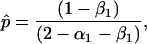 |
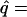 |
6 |
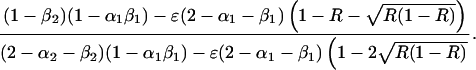 |
As with our simpler model, here the selection generated by the resource can shift the position of polymorphic equilibria. The direction of the shift is in favor of a when ɛ is positive and R < 1/2 and also when ɛ is negative and R > 1/2 and in favor of A when ɛ is positive and R > 1/2 and also when ɛ is negative and R < 1/2. As before, the selection generated by the resource is strongest when R is close to 0 or 1 and absent at R = 1/2. By influencing the amount of the resource, positive or negative niche construction can dramatically change the position of the equilibrium and may change the direction of selection resulting from R. In Fig. 2a, the effect of positive niche construction is to increase the amount of the resource and as a result, the selection favoring a generated by the resource becomes much weaker, and A reaches a considerably higher frequency. This is best understood by focusing on the effect of niche construction on the frequency-dependent components of the contributions to fitness of genotypes AA, Aa, and aa, that is ɛR, ɛ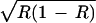 , and ɛ(1 − R), respectively. Here, with no positive niche construction (λ2 = 0 and R = 0.167), ɛR < ɛ
, and ɛ(1 − R), respectively. Here, with no positive niche construction (λ2 = 0 and R = 0.167), ɛR < ɛ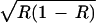 < ɛ(1 − R), and the frequency-dependent selection overcomes the fixed-fitness component of selection and drives a to fixation. However, with positive niche construction (λ2 = 0.1, which increases R to 0.47), ɛR < ɛ(1 − R) < ɛ
< ɛ(1 − R), and the frequency-dependent selection overcomes the fixed-fitness component of selection and drives a to fixation. However, with positive niche construction (λ2 = 0.1, which increases R to 0.47), ɛR < ɛ(1 − R) < ɛ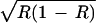 , allowing alleles A and a to coexist. In Fig. 2b, the effect of positive niche construction is again to increase the amount of the resource, but here, the selection generated by the resource switches from favoring a to favoring A and takes A to fixation. Now, with no niche construction (λ2 = 0 and R = 0.2), ɛR < ɛ
, allowing alleles A and a to coexist. In Fig. 2b, the effect of positive niche construction is again to increase the amount of the resource, but here, the selection generated by the resource switches from favoring a to favoring A and takes A to fixation. Now, with no niche construction (λ2 = 0 and R = 0.2), ɛR < ɛ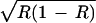 < ɛ(1 − R), but with niche construction (λ2 = 0.2, which increases R to 0.56), ɛR > ɛ
< ɛ(1 − R), but with niche construction (λ2 = 0.2, which increases R to 0.56), ɛR > ɛ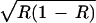 > ɛ(1 − R). Although the effects of niche construction are not always so dramatic, it is important to bear in mind that there are circumstances in which niche construction can strongly influence the pattern of selection acting on a population, even in the face of significant external renewal and depletion of the resource.
> ɛ(1 − R). Although the effects of niche construction are not always so dramatic, it is important to bear in mind that there are circumstances in which niche construction can strongly influence the pattern of selection acting on a population, even in the face of significant external renewal and depletion of the resource.
Figure 2.

Positive niche construction, independent renewal, and heterozygote advantage (Rt = λ1Rt−1 + λ2pt + λ3, γ = 0, α1 = α2 = 0.99, β1 = β2 = 0.9, r = 1/2, n = 1). Provided r is not very small, populations converge to a single equilibrium in linkage equilibrium, the position of which may be strongly affected by niche construction. The + represents the equilibrium in the absence of niche construction. By increasing R, niche construction changes the frequency-dependent component of genotype fitnesses, affecting the equilibrium frequency of allele A. In both a and b, without niche construction, allele a would be fixed, whereas in a, niche construction allows A and a to coexist, and in b, niche construction drives A to fixation.
In this analysis, we have not addressed the effects of niche construction on the amount of linkage disequilibrium at equilibria with small values for r, because this topic was considered in some detail in our earlier analysis (9).
DISCUSSION
The results support the conclusions of our earlier analysis, confirming that, even with independent renewal and depletion of the key resource, the effects of niche construction can override external sources of selection to create new evolutionary trajectories and equilibria, generate and eliminate polymorphisms, and produce time lags in the response to selection as well as other unusual dynamics.
The consequences of niche construction are particularly interesting when the selection it generates opposes the action of an external source of selection acting at the A locus. This kind of niche construction is likely to be common. Lewontin (7) points out that many of the activities of organisms, such as migration, hoarding of food resources, habitat selection, or thermoregulatory behavior, are adaptive precisely because they dampen statistical variation in the availability of environmental resources. Our results confirm that the frequency-dependent selection generated from the resource and modified by niche construction can (i) overcome the external selection to lead to the fixation of otherwise deleterious alleles and (ii) support stable polymorphisms where none are expected. Our analysis illustrates how the probabilities of (i) and (ii) are affected by independent renewal and depletion of resources and the sign of ɛ. In cases where an increase in the amount of resource results in an increment in the fitness of genotypes containing allele A (i.e., when ɛ is positive), independent renewal decreases and independent depletion increases the chances that niche construction will lead to the fixation of otherwise deleterious alleles or support stable polymorphisms. In cases where an increase in the amount of resource results in an increment in the fitness of genotypes containing allele a (when ɛ is negative), independent renewal increases and independent depletion decreases the chances that niche construction will lead to the fixation of otherwise deleterious alleles or support stable polymorphisms. These findings hold for both positive and negative niche construction.
We face a fundamental problem in using a two-locus frequency-dependent selection model to explore niche construction in that an infinite number of frequency-dependent relationships are conceivable, and it is not clear whether any general findings will emerge from a particular model. This is why, here and in our earlier analyses, we have used the comparatively well studied multiplicative model as a baseline against which to compare our findings. Nevertheless, it is well established that even simple models of frequency-dependent selection can generate a broad variety of outcomes depending on the choice of parameters, and hence it is important to stress which outcomes are not predicted as well as which are possible. It is in this respect that our model gives particularly encouraging findings. Although the analysis reveals a rich array of possible outcomes, the patterns that emerge exhibit a predictable symmetry, they are easy to interpret relative to simpler models, and the dynamics are not chaotic, cyclical, or irregular. This suggests that, given sufficient understanding of the relationships between input parameters, it may be possible to make qualitative predictions, for example, as to the likelihood that (and circumstances under which) niche construction will generate polymorphisms. The fact that different frequency-dependent models might make alternative predictions we regard as a virtue rather than a problem, because a comparison between the models and the data will facilitate a deeper understanding of the consequences of niche construction. For example, earlier, we noted that assumptions as to how R is computed strongly affect the probability that niche construction will generate time lags. We have only begun to explore this issue, and a much more thorough analysis of the circumstances under which niche construction either leads or does not lead to time lags is required. Ultimately, it is an empirical question to assess which of the many possible functions describing how niche construction affects R is closest to biological reality.
Although our model is an advance on our earlier analysis, we are aware of the constraints imposed by its two-locus, population-genetic structure. Although we were able to investigate how a population’s evolutionary dynamics were affected by the proportion of niche constructors in the population (p) and the relative impact that this niche construction had on the resource (λ2 and γ), we have not explored other demographic and ecological parameters. There is no doubt that it would be valuable to synthesize population-genetic and ecological models in order to explore factors such as the size and density of the niche-constructing population, the scale of its impact on the environment, and how niche construction influences (and is influenced by) growth rates, carrying capacities, and other relevant demographic parameters.
Some of the wider implications of our results for ecology become apparent when niche construction is equated with ecosystem engineering. Jones et al. (4) point out that a major ecological consequence of the niche construction of organisms is that it establishes “engineering webs,” or control webs, in both communities and ecosystems. Engineering webs do not conform to the same principles of mass flow, stoichiometry, and the conservation of energy that govern the more familiar energy and material flows and trophic relations among organisms. This makes it difficult to understand how engineering webs achieve their control or to predict which organisms are likely to have the biggest effect on an ecosystem. However, ecosystem engineering depends on the adaptations of organisms, which may drive evolution when they modify natural-selection pressures by niche construction and when they generate legacies of modified natural-selection pressures for subsequent generations (8). Thus, regardless of whether organisms are themselves members of a specified trophic web, their niche-constructing adaptations may qualify them as members of an associated engineering web, which may allow them to exert a degree of evolutionary as well as ecological control over ecosystems. In this case, the control that is exerted by organisms should promote the kind of phenomena we investigate here, generating changes in the adaptive design of components of an ecosystem (27). This could be explored empirically either by comparative work among natural populations or by cancelling out the effects of niche construction experimentally to create artificial comparisons among, for instance, virgin, engineered, and degraded habitats (4). We anticipate that where niche construction exerts control over ecosystems via engineering webs, it may promote the kind of equilibria with functional integration that renders ecosystems orderly and which at the largest scale is exemplified by the Gaia hypothesis (28). However, under different circumstances, the feedback generated by niche construction may introduce new sources of dynamic complexity into ecosystems, which could generate chaotic interactions.
At present, the utility of our model is limited by the fact that so far we have only modeled the feedback from a single population of niche constructors to itself. In principle, our model could be generalized to consider cases where the E and A loci are in two coevolving populations. This would be most novel in considerations of the positive effects of one population’s niche construction on the other’s fitness, for example, in mutualistic interactions via byproducts. However, even where the effects of one population’s niche construction on the other’s fitness are negative, because the two genetic loci in our model interact via an intermediate environmental component, R, and because the amount of R is unlikely always to be directly proportional to either the number of organisms or the frequency of genes in the first population, the dynamics are likely to differ considerably from established coevolutionary models (29). Moreover, the discordance between resource and population dynamics will be exacerbated where there is ecological inheritance. Second, the resource R could be biotic; for instance, R could comprise a third population through which two other populations interact, in which case our model should translate into a community module (30, 31); or it could be abiotic, where our model should translate into an ecosystem module. Adding additional biotic Rs to our model could stretch the community module toward a more complete food cycle or web (6). Third, our model could be generalized to deal with chains of biotic and/or abiotic components, such as a food chain in a community or an ecological flow chain in an ecosystem (6, 25). Although the introduction of such complexity may demand a different type of theoretical machinery, the logic underlying the exercise remains the same.
The differences between ecological and evolutionary time scales might also cast doubt on the idea that engineering webs are also evolutionary webs and that evolutionary processes can work fast enough to contribute to the control of energy and matter flows in ecosystems by adjusting the adaptations of organisms. The analyses presented here help eliminate this doubt by illustrating how niche construction and natural selection may operate within similar time frames. In general, the distinction between the ecological and evolutionary time scales has blurred in the face of evidence for rapid evolutionary change (32). Moreover, even where evolutionary change in engineering adaptations is slow, it should still become progressively more relevant to engineering webs as the duration of the association among the populations of an ecosystem increases (e.g., 33).
Hitherto, it has not been possible to apply evolutionary theory to ecosystems, because of the presence of nonevolving abiota in ecosystems. We suspect this obstacle has been largely responsible for preventing the full integration of ecosystem ecology with population–community ecology (2, 34). However, by adding the new process of niche construction to the established process of natural selection, the present model enables the incorporation of both abiotic environmental components and interactions among populations and abiota in ecosystems into evolutionary models. Unlike standard evolutionary theory, the present approach is equally applicable to both population–community ecology and ecosystem-level ecology, which may eventually make it easier to reconcile these two ecological subdisciplines under the rubric of an extended evolutionary theory that includes niche construction.
Acknowledgments
We are grateful to Bob Holt, John Lawton, Peter Richerson, and two anonymous referees for helpful comments on earlier drafts of this manuscript. Kevin Laland was supported by a Royal Society University Research Fellowship. This research was supported in part by National Institutes of Health Grant GM28016.
References
- 1.Vitousek P M, Mooney H A, Lubchenco J, Melillo J M. Science. 1997;127:494–499. [Google Scholar]
- 2.Odling-Smee F J. In: The Role of Behavior in Evolution. Plotkin H C, editor. Cambridge, MA: MIT Press; 1988. pp. 73–132. [Google Scholar]
- 3.Jones C G, Lawton J H, Shachak M. Oikos. 1994;69:373–386. [Google Scholar]
- 4.Jones C G, Lawton J H, Shachak M. Ecology. 1997;78:1946–1957. [Google Scholar]
- 5.Chapin F S, Walker B H, Hobbs R J, Hooper D U, Lawton J H, Sala O E, Tilman D. Science. 1997;277:500–504. [Google Scholar]
- 6.DeAngelis D L. Dynamics of Nutrient Cycling and Food Webs. London: Chapman & Hall; 1992. [Google Scholar]
- 7.Lewontin R C. In: Evolution from Molecules to Men. Bendall D S, editor. Cambridge, U.K.: Cambridge Univ. Press; 1983. pp. 273–285. [Google Scholar]
- 8.Odling-Smee F J, Laland K N, Feldman M W. Am Nat. 1996;147:641–648. [Google Scholar]
- 9.Laland K N, Odling-Smee F J, Feldman M W. J Evol Biol. 1996;9:293–316. doi: 10.1046/j.1420-9101.2001.00262.x. [DOI] [PubMed] [Google Scholar]
- 10.Preston-Mafham K, Preston-Mafham R. The Natural History of Spiders. Marlborough, U.K.: Crowood; 1996. [Google Scholar]
- 11.Hansell M. Animal Architecture and Building Behavior. New York: Longman; 1984. [Google Scholar]
- 12.Hölldobler B, Wilson E O. Journey to the Ants: A Story of Scientific Exploration. Cambridge, MA: Belknap; 1995. [Google Scholar]
- 13.Nowak R M. Walker’s Mammals of the World. 5th Ed. Baltimore, MD: Johns Hopkins Univ. Press; 1991. [Google Scholar]
- 14.Darwin C. The Formation of Vegetable Mould through the Action of Worms, with Observations on their Habits. London: Murray; 1881. [Google Scholar]
- 15.Lee K E. Earthworms: Their Ecology and Relation with Soil and Land Use. London: Academic; 1985. [Google Scholar]
- 16.Holmgren M, Scheffer M, Huston M A. Ecology. 1997;78:1966–1975. [Google Scholar]
- 17.Bertness M D, Leonard G. Ecology. 1997;78:1976–1989. [Google Scholar]
- 18.Callaway R M, Walker L R. Ecology. 1997;78:1958–1965. [Google Scholar]
- 19.Hacker S D, Gaines S D. Ecology. 1997;78:1990–2003. [Google Scholar]
- 20.Mount A B. Austr Forestry. 1964;28:166–172. [Google Scholar]
- 21.Whelan R J. The Ecology of Fire. Cambridge, U.K.: Cambridge Univ. Press; 1995. [Google Scholar]
- 22.Kerr, B., Schwilk, D. W., Bergman, A. & Feldman, M. W. (1999) Evol. Ecol. Res., in press.
- 23.Polis G A, Strong D R. Am Nat. 1996;147:813–846. [Google Scholar]
- 24.MacArthur RH, Levins R. Am Nat. 1967;101:377–385. [Google Scholar]
- 25.Shachak M, Jones C G. In: Linking Species and Ecosystems. Jones C G, Lawton J H, editors. London: Chapman & Hall; 1995. pp. 280–296. [Google Scholar]
- 26.Gurney W S C, Lawton J H. Oikos. 1996;76:273–283. [Google Scholar]
- 27.Holt R D. In: Linking Species and Ecosystems. Jones C G, Lawton J H, editors. London: Chapman & Hall; 1995. pp. 273–279. [Google Scholar]
- 28.Lenton T M. Nature (London) 1998;394:439–47. doi: 10.1038/28792. [DOI] [PubMed] [Google Scholar]
- 29.Futuyma D J, Slatkin M. Coevolution. Sunderland, MA: Sinauer; 1983. [Google Scholar]
- 30.Holt R D. In: Multitrophic Interactions. Begon M, Gange A, Brown Y, editors. London: Chapman & Hall; 1996. pp. 333–350. [Google Scholar]
- 31.Holt R D, Polis G A. Am Nat. 1997;149:745–764. [Google Scholar]
- 32.Thompson J N. Trends Ecol Evol. 1998;13:329–332. doi: 10.1016/s0169-5347(98)01378-0. [DOI] [PubMed] [Google Scholar]
- 33.Milchunas D G, Lauenroth W K. Ecol Monogr. 1993;63:327–366. [Google Scholar]
- 34.O’Neill R V, DeAngelis D L, Waide J B, Allen T F H. A Hierarchical Concept of Ecosystems. Princeton, NJ: Princeton Univ. Press; 1986. [Google Scholar]