

Abstract
Four experiments using rats in a Pavlovian lick-suppression preparation investigated the effects of combining 2 treatments known for their response-decrementing effects, namely, overshadowing and degraded contingency. Contrary to most contemporary learning theories, the extended comparator hypothesis predicts that these 2 treatments will counteract each other, and therefore, less of a decrement in conditioned responding should be observed than with either treatment alone. Experiments 1 and 2 confirmed this prediction in first-order conditioning and sensory preconditioning preparations, respectively. Experiment 3 demonstrated that posttraining extinction of the training context resulted in a recovery from degraded contingency and reversed the counteractive effect on overshadowing. Finally, Experiment 4 demonstrated that posttraining extinction of the overshadowing stimulus resulted in recovery from simple overshadowing and also reversed the counteractive effect on degraded contingency. These results are consistent with the extended comparator hypothesis but not traditional or recent acquisition-focused models.
Keywords: overshadowing, degraded contingency, extended comparator hypothesis, retrospective revaluation, Pavlovian conditioning
Recent findings have demonstrated, at least in select cases, that two treatments that, if administered in isolation, produce decrements in conditioned responding no longer do so when administered conjointly; their effects are not summative, rather, they counteract each other. For example, Urushihara, Stout, and Miller (2004) observed counteraction between the conditioned stimulus (CS) duration effect and the overshadowing effect.1 Specifically, subjects that experienced long CSs trained in compound with a more salient (overshadowing) cue responded more vigorously to the target CS than did subjects that experienced either the long CS alone or a short CS trained in compound. This observation of counteractive effects between two response-decrementing treatments is not unique. Stout, Chang, and Miller (2003) observed that the trial massing effect (massed trials producing less conditioned responding than that produced by spaced trials) interacts with the overshadowing effect. Specifically, subjects that experienced massed training with the target CS in compound with a more salient cue responded more vigorously to the target CS than did subjects that experienced either massed conditioning trials with the target CS alone or spaced conditioning trials with the target CS in compound. Moreover, researchers have observed that the CS-preexposure effect (i.e., latent inhibition; Lubow, 1973) is counteracted when the preexposed CS is trained in the presence of an overshadowing cue (Blaisdell, Bristol, Gunther, & Miller, 1998; Savastano, Arcediano, Stout, & Miller, 2003; see also Loy & Hall, 2002, although their data failed to achieve statistical significance). Of note, all these demonstrations suggest a mechanism that is not anticipated by traditional associative models of learning.
Most learning theories that were developed to explain the overshadowing effect have proposed, albeit by different mechanisms, that when trained in the presence of another cue, the target cue X fails to enter into as strong an association with the US as it does when trained alone. Likewise, these models assume that when conditioning trials are massed the training context competes with the target CS during training, and therefore, the target CS fails to enter into association with the US. Hereafter, we refer to this family of models as acquisition-focused models (e.g., Mackintosh, 1975; Pearce, 1987; Pearce & Hall, 1980; Rescorla & Wagner, 1972; Wagner, 1981). With regard to the above empirical observations, all these models predict that the effects of these treatments should be additive rather than counteractive.
In contrast to this view, some models attribute the effects of overshadowing to a failure to express an association that was acquired rather than to a failure to acquire the association. For example, the extended comparator hypothesis (Denniston, Savastano, & Miller, 2001), which is a response rule for the expression of associations, explains overshadowing by proposing that, at the time of testing, subjects compare the representation of the US retrieved by the target stimulus X with that retrieved by the overshadowing stimulus A. Specifically, the extended comparator hypothesis (see Figure 1) states that responding during a test of target stimulus X is determined by a comparison between the US representation directly activated by X (Link 1) and the representation of the US indirectly activated by X through mediation by the overshadowing stimulus A (i.e., the product of Links 2 and 3). Given that in overshadowing treatment, A and X are presented in compound, Link 2 should be strong, and given that A is typically more salient than X and is consistently paired with the US, Link 3 should be strong. Therefore, at the moment of testing, the indirectly activated US representation should be able to down-modulate the responding to X that is supported by Link 1. By assuming that subjects encode training events in a noncompetitive manner that is based on contiguity and appealing to a comparator process at the time of testing, the extended comparator hypothesis has been successful in explaining cue competition and many other learning effects.
Figure 1.
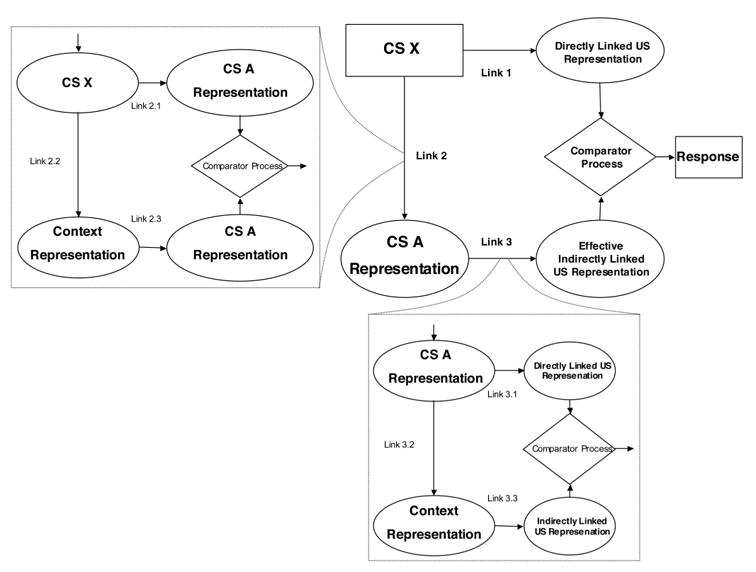
A schematic representation of the extended comparator hypothesis (after Denniston, Savastano, & Miller, 2001) and how it accounts for the counteraction of overshadowing by concurrent degraded contingency treatment. Here, the overshadowing stimulus is depicted as the target cue’s first-order comparator stimulus, and the training context serves as the target cue’s second-order comparator stimulus. As implemented, the training context also acts as the target cue’s first-order comparator stimulus, and the overshadowing cue serves as the target cue’s second-order comparator stimulus.
The extended comparator hypothesis, however, also assumes that the two links between the target cue and the indirectly activated representation of the US are also subject to a comparator process in which second-order comparator stimuli participate in down-modulating the strengths of Links 2 and 3. Therefore, the effectiveness of a comparator stimulus on down-modulating responding to the target cue is subject to its own second-order comparator stimuli (Links 2.2, 2.3, 3.2, and 3.3 in Figure 1). Moreover, a single stimulus can simultaneously serve as both a first-order and a second-order comparator stimulus for a given target cue as long as it has appreciable associative strength with both the target cue and with another first-order comparator stimulus. The extended comparator hypothesis accounts for the counteraction of CS-preexposure and overshadowing effects by assuming that, during preexposure to the target cue (X), the context is established as a first-order comparator for X, whereas during reinforced training (i.e., AX-US), the overshadowing stimulus A is established as another first-order comparator stimulus. Moreover, during the AX-US pairings, (a) A also becomes a comparator stimulus for the context (and, hence, a second-order comparator stimulus for X), and (b) the context becomes a comparator stimulus for A (and, hence, a second-order comparator stimulus for X). The net result is that A down-modulates the effectiveness of the context as a first-order comparator stimulus for X, and the context down-modulates the effectiveness of A as a first-order comparator stimulus for X. As was already mentioned, a similar counteraction among comparators has been observed when a compound of cues was trained with massed training trials (i.e., combining massed trial spacing treatment and overshadowing treatment; Stout et al., 2003) and when cues of long duration were trained in compound (combining long CS-duration treatment with overshadowing treatment; Urushihara et al., 2004). Although problematic for traditional acquisition-focused models, all these findings are consistent with predictions that are based on the extended comparator hypothesis.
The present series of studies is concerned with the potential interaction of another pair of treatments that have detrimental effects on acquired stimulus control of behavior. One of them is the previously mentioned overshadowing effect, and the other treatment examined here that attenuates conditioned responding is the degraded contingency effect. Initially described by Rescorla (1966), the degraded contingency effect is observed as a decrease in responding as a consequence of presenting unsignaled USs interspersed with the CS-US pairings of training. In the framework of acquisition-focused models, the degraded contingency effect has been viewed as the result of the context gaining associative strength with the US and consequently blocking the acquisition of associative strength by the target CS (Rescorla & Wagner, 1972). The degraded contingency effect has been observed in diverse preparations (e.g., Jenkins, Barnes, & Barrera, 1981; Rescorla, 1968), and the notion that the context blocks associative acquisition by the target CS is congruent with studies in which the presentations of the US in the absence of X were signaled by an alternative stimulus. Supposedly, this added signal prevents the context from gaining associative strength, and consequently, the context no longer blocks the target cue from acquiring an association with the US, thereby dissipating the degraded contingency effect (Durlach, 1983, 1984). Alternatively, the extended comparator hypothesis has accounted for the degraded contingency effect by postulating that the strong context–US association interferes with the expression of the target cue–US association at the time of testing. This view assumes that the unsignaled US presentations enhance the context–US association (Link 3 for the target cue), and acquisition of the X-US association is unaffected by the presentation of the US alone during training (Gunther & Miller, 2000). Moreover, the signaling of the added USs by a nontarget cue is presumed to establish the added signal as a first-order comparator stimulus for the context and, consequently, a second-order comparator stimulus for X, which down-modulates the response-attenuating effect of the context.
It is interesting that the extended comparator hypothesis predicts that if subjects are trained both with an overshadowing stimulus (A) present during the X-US trials (i.e., AX-US trials) and with unsignaled US presentations interspersed among the AX-US trials, the overshadowing effect should be attenuated as a result of the excitatory status of the context because the context should down-modulate the effectiveness of the overshadowing stimulus in interfering with responding to the target cue (X; see Figure 1). Likewise, the overshadowing stimulus should down-modulate the effectiveness of the context in interfering with responding to the target cue at the time of testing. In other words, the extended comparator hypothesis predicts that both first-order comparator stimuli (the overshadowing stimulus and the context) for X, by also acting as second-order comparator stimuli, will cancel each other, thereby enhancing stimulus control by the target cue in comparison with simple overshadowing and simple degraded contingency treatments. This prediction is illustrated in Figure 2, which depicts simulations of simple acquisition, overshadowing, the degraded contingency effect, and the interaction of overshadowing and degraded contingency. We used Savastano, Arcediano, et al.’s (2003; see also Stout & Miller, 2005) mathematical implementation of the extended comparator hypothesis, including exactly their parameters and the number of trials used in the present Experiment 1. As shown in Figure 2, the simulation indicates that when both treatments are presented together, a partial recovery from overshadowing and from degraded contingency is predicted. In contrast to the prediction of the extended comparator hypothesis, acquisition-focused models make the opposite prediction. These models, although for different reasons, all predict additive rather than counteractive effects of overshadowing and degraded contingency treatments. The reason acquisition-focused models predict additive effects is that they all explain cue competition as arising from the division of a limited resource (attention or association to the US) among all cues present on a trial. Thus, when two competing cues (A and context) are present, they both should compete with the target cue, with the consequence that it accrues less associative strength. The present series of experiments was conducted to test this prediction of the extended comparator hypothesis and therefore to contrast this model with the above-mentioned acquisition-focused models.
Figure 2.
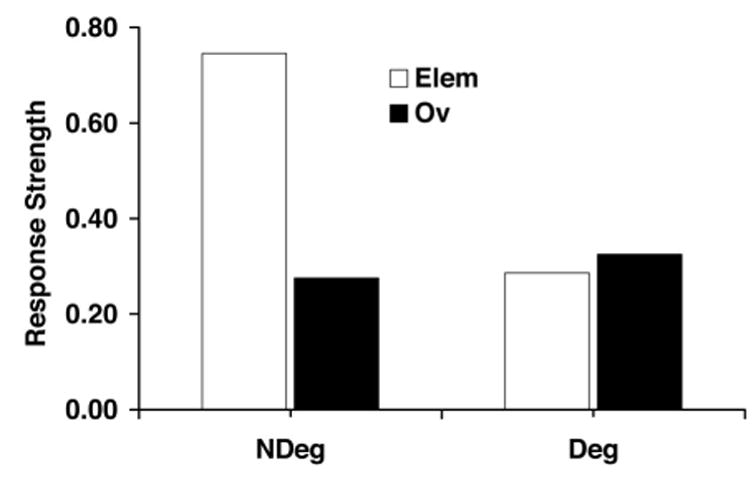
Simulation of Experiment 1 after Savastano, Arcediano, Stout, and Miller’s (2003) implementation of the extended comparator hypothesis. All parameters were the same as those used by Savastano, Arcediano, and colleagues, and the numbers of trials were the same as those used in Experiment 1 of the present series of studies. Elem = elemental; Ov = overshadowing; Deg = degraded contingency; NDeg = no degraded contingency.
Experiment 1
To contrast these different predictions of the extended comparator hypothesis and the aforementioned acquisition-focused models, a factorial design was used in which the main factors were whether the target cue was trained in compound with a more salient cue (overshadowing) and whether training occurred with concurrent unsignaled presentations of the US (degraded contingency). The design of Experiment 1 is summarized in Table 1. Subjects were randomly assigned to one of two conditions that experienced partial reinforcement training with either a single cue (X; elemental) or with a compound of cues (AX; overshadowing). These two conditions were factorially subdivided into two orthogonal conditions in which subjects did or did not experience unsignaled presentations of the US alone during the interval between the reinforced trials with X (X-US or AX-US).
Table 1.
Design of Experiment 1
| Group | Training | Test |
|---|---|---|
| Elem-NDeg | X→US (18)/X- (18) | X |
| Ov-NDeg | AX→US (18)/AX- (18) | X |
| Elem-Deg | X→US (18)/US (108)/X- (18) | X |
| Ov-Deg | AX→US (18)/US (108)/AX- (18) | X |
Note. Numbers in parentheses next to the pairings indicate total number of each type of trial during training. → = “followed”; X = click train; A = tone; US = foot shock; Elem = elemental training; Ov = overshadowing (compound training); Deg = degraded contingency; NDeg = no degraded contingency.
Method
Subjects
Subjects were 24 female and 24 male Sprague-Dawley, experimentally naïve, young adult rats (N = 48) that were bred in our colony. Body weight range was 156–196 g for females and 233–282 g for males. Subjects were individually housed and maintained on a 16-hr light/8-hr dark cycle, with experimental sessions occurring roughly midway through the light portion. All subjects were handled for 30 s three times per week from weaning until the initiation of the study. Subjects had free access to food in the home cage. One week prior to initiation of the experiment, water availability was progressively reduced to 30 min per day and was provided approximately 2 hr after any scheduled treatment.
Apparatus
Two counterbalanced contexts (Chambers V and R) were used in this experiment. Chamber V was the shape of a vertical truncated V that was 27 cm long, 29.5 cm high, 21.5 cm wide at the top, and 5.5 cm wide at the bottom. The ceiling was clear Plexiglas, the front and back walls were black Plexiglas, and the side walls were stainless steel. The floor was composed of two 27-cm (long) × 2-cm (wide) plates and had a 1.5-cm gap between the two plates. A 0.5-s, 0.7-mA constant-current foot shock, produced by a high-voltage AC circuit in series with a 1.0-MΩ resistor, could be delivered through the metal walls and floor of the chamber. Each of six copies of Chamber V was housed in a separate sound- and light-attenuating environmental isolation chest. The chambers were each illuminated by a 7-W (nominal at 120 VAC, but driven at 60 VAC) light bulb, which was mounted on the inside wall of the environmental enclosure, approximately 30 cm from the center of the experimental chamber. The light entered the chamber primarily by reflection from the ceiling of the environmental chest.
Chamber R was rectangular, measuring 24.0 × 9.0 × 12.5 cm (l × w × h). The walls and ceiling of Chamber R were clear Plexiglas, and the floor was composed of stainless steel rods measuring 0.5 cm in diameter, spaced 1.5 cm apart (center to center). The rods were connected by NE-2 bulbs, which allowed for the delivery of a 0.5-s, 0.7-mA constant-current foot shock produced by a high-voltage transformer in series with a 1.0-MΩ resistor. Each of six copies of Chamber R was housed in separate light- and sound-attenuating environmental isolation chambers. Each chamber was dimly illuminated by a 2-W (nominal at 120 VAC, but driven at 60 VAC) incandescent house light mounted on an inside wall of the environmental chest located approximately 30 cm from the animal enclosure. The light intensities inside the two types of chambers were approximately equal because of the difference in opaqueness of the walls of Chambers V and R.
One context was used for training and the other for testing, counterbalanced within groups. The rationale for using different contexts for training and testing was that some groups received additional US presentations during training (degraded contingency treatment), and therefore, the training context possibly gained behavior control for these subjects. Consequently, the training context may have obscured differences in the associative status of the CS at the time of testing if only one context had been used. Further, the extended comparator hypothesis states that it is the context present at the time of training that is associatively retrieved during testing with the target cue. Hereafter, the context used for training is referred to as the training context, whereas the context used for acclimation, reacclimation, and testing is referred to as the test context.
All chambers could be equipped with a water-filled lick tube that extended 1 cm into a cylindrical niche, which was 4.5 cm in diameter, left-right centered, with its bottom 1.75 cm above the floor of the apparatus and 5.0 cm deep. There was an infrared photo beam 1-cm in front of the lick tube that was broken whenever a subject licked the tube. Two 45-Ω speakers on separate walls of the isolation chests could deliver either a low-frequency complex tone (1,000 & 1,200 Hz) 10 dB (C-scale) above background (76 dB, produced mainly by a ventilation fan) or a click (6/s) 6 dB (C) above background. The 0.5-s, 0.7-mA foot shock, which served as the US, could be delivered through the chamber floors. In this experiment, the tone served as CS A, and the clicks served as CS X; during training, both were 30 s in duration.
Procedure
Subjects were randomly assigned to one of four groups: elementary-no degraded contingency (Elem-NDeg), overshadowing-no degraded contingency (Ov-NDeg), elementary-degraded contingency (Elem-Deg), and overshadowing-degraded contingency (Ov-Deg), counterbalanced for sex (ns = 12).
Acclimation
Subjects were acclimatized to the test context on Days 1 and 2 during 45-min daily sessions with the lick tubes available. During these two daily sessions, no nominal stimulus was presented.
Training
Prior to the beginning of the training phase, the lick tubes were removed from the experimental chambers. On Days 3–8, during daily 45-min sessions in the training context, subjects in Condition Ov received three reinforced AX trials and three nonreinforced AX trials (interspersed). Subjects in Condition Elem received three reinforced X trials and three nonreinforced X trials. The mean (± SE) intertrial interval was 7 min (± 2 min). Partial reinforcement (50%) was used because previous studies have demonstrated that these parameters yield a large degraded contingency effect (Gunther & Miller, 2000). Half of the subjects in each of the above-mentioned conditions were also exposed to 18 daily presentations of the US alone during the 45-min training session (Groups Elem-Deg and Ov-Deg). The mean interval between stimulus events was approximately 90 s (± 60 s). The remaining subjects (Groups Elem-NDeg and Ov-NDeg) did not receive these additional US presentations. On reinforced trials, the US was presented during the last 0.5 s of either X alone or the AX compound.
Reacclimation
On Days 9 and 10, the lick tubes were reinserted, and subjects were allowed to drink during two 45-min daily sessions in the test context. This treatment served to restabilize baseline levels of drinking.
Testing
On Day 11, all subjects were tested for conditioned suppression to X by presenting X for 15 min, starting immediately on completion of each subject’s first 5 cumulative seconds of licking (as measured by the total amount of time the photo beam was disrupted). Thus, all subjects were drinking in the test context at the time of CS onset. Times to complete this initial 5 cumulative seconds of licking and times to complete 5 additional seconds of licking after the onset of the test CS were recorded. Test sessions were 16 min in duration, with a ceiling score of 15 min imposed on the time to complete 5 cumulative seconds of drinking in the presence of the test CS. Suppression data were transformed to log (base 10) scores to approximate a normal distribution and thereby facilitate the use of parametric statistics. An alpha level of .05 was adopted for all tests of statistical significance. Following the convention of our laboratory, subjects that took more than 60 s to complete their first 5 cumulative seconds of licking (i.e., prior to CS onset), thereby exhibiting an unusual reluctance to drink in the test context, were eliminated from all analyses. In accordance with this criterion, one subject from Group Elem-NDeg was eliminated.
Results and Discussion
The results are shown in Figure 3. As can be observed, subjects for which the target cue X was trained in the presence of a more salient cue A without unsignaled USs (Group Ov-NDeg), or for which X without A was trained conjointly with unsignaled US presentations (Group Elem-Deg), suppressed less than did subjects in the acquisition-alone control group (Group Elem-NDeg), suggesting that the present parameters resulted in reliable overshadowing and a degraded contingency effect. Of interest, and consistent with the predictions of the extended comparator hypothesis, subjects for which the target cue X was trained on the presence of a more salient cue A and that also experienced unsignaled presentations of the US (Group Ov-Deg) displayed less of a decrement in responding than did subjects in Group Ov-NDeg or Group Elem-Deg. Thus, overshadowing treatment and degraded contingency treatment counteracted each other. These results were confirmed by the following statistical analyses.
Figure 3.
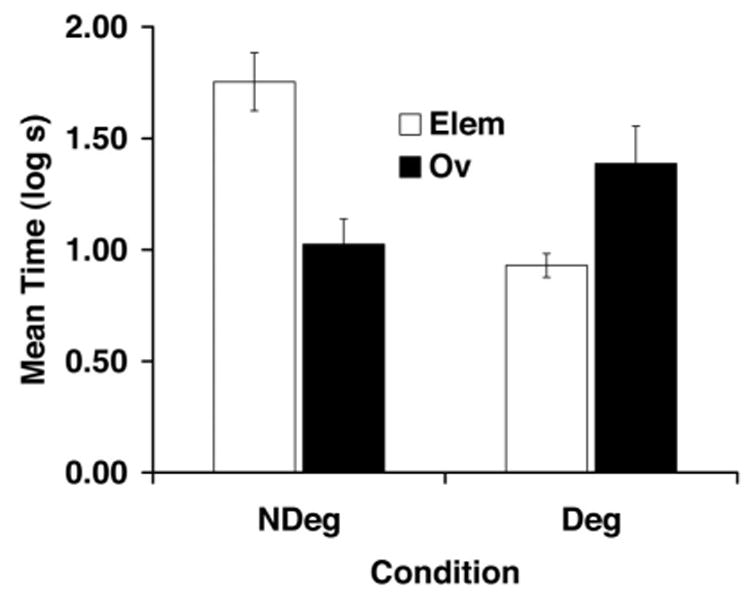
Mean times to complete 5 cumulative seconds of drinking upon presentation of the target CS (X) in Experiment 1. See Table 1 for treatments of the four groups. Elem = elemental; Ov = overshadowing; Deg = degraded contingency; NDeg = no degraded contingency. Error bars denote standard errors of means.
A 2 × 2 analysis of variance (ANOVA), in which the first factor was cue competition (Overshadowing vs. Elemental) and the second factor was contingency (Degraded Contingency vs. No Degraded Contingency), performed on the pre-CS times to complete 5 cumulative seconds of drinking on Day 11 failed to detect main effects or an interaction (all ps > .24), indicating that there were no significant differences between groups in terms of fear to the testing context.
A similar ANOVA, performed on the times to complete 5 cumulative seconds of drinking during the test presentation of the CS, failed to detect main effects but revealed an interaction, F(1, 43) = 23.46, p < .001, MSE = 0.175. Planned comparisons using the overall error term from the ANOVA found that subjects in Groups Ov-NDeg and Elem-Deg suppressed less to the CS than did Group Elem-NDeg, thereby demonstrating simple overshadowing, F(1, 43) = 17.36, p < .05, and a simple degraded contingency effect, F(1, 43) = 22.18, p < .05, respectively. The critical group that received both overshadowing and degraded contingency (Group Ov-Deg) suppressed more than did subjects in either Group Ov-NDeg, F(1, 43) = 4.46, p < .05, or Group Elem-Deg, F(1, 43) = 7.11, p < .05. This indicates that the response-attenuating effects of both overshadowing and degraded contingency treatments were reversed in subjects that received both treatments together.
With the present parameters, this study allowed us to observe the response-decrementing effects of overshadowing and degraded contingency treatments (Groups Ov-NDeg and Elem-Deg) compared with the group that did not receive either of these treatments (Elem-NDeg). On the basis of the extended comparator hypothesis, overshadowing treatment results in weaker response to the overshadowed cue X because of the strong X-A (Link 2) and A-US (Link 3) associations. Likewise, degraded contingency treatment results in a weaker response to X because of the strong X-context (Link 2) and context-US (Link 3) associations. Therefore, both response-decrementing treatments are explained by the extended comparator hypothesis in terms of a strong indirectly activated representation of the US, although there are different critical first-order comparator stimuli. It is interesting that the extended comparator hypothesis further predicts that the links that constitute the indirectly activated representation of the US (Links 2 and 3) are themselves subject to a comparator process in which second-order comparator stimuli can down-modulate the strength of first-order comparator stimuli. This aspect of the extended comparator hypothesis allowed it to predict in the present case that the two treatments would counteract each other when experienced together. It could be argued that the diminished responding observed in Group Ov-NDeg relative to Group Elem-NDeg was due to generalization decrement rather than to an interaction of A and X (Pearce, 1987). This argument supposes that suppression to the target cue X was diminished because it was trained in compound with another cue (A) and tested alone. Although we did not include a control group for generalization decrement, subjects in the Group Ov-Deg were also trained with the compound AX and tested with the target cue X alone, and they did not show any appreciable diminished responding to X relative to Group Elem-Deg.
Experiment 2
As mentioned in the introduction, the present series of experiments was designed to test predictions of the extended comparator hypothesis. Consistent with the processes by which the extended comparator hypothesis anticipated the counteractive effect of overshadowing and degraded contingency, posttraining extinction of either of the comparator stimuli (i.e., the training context or the overshadowing stimulus A) should reverse the counteractive effects of the other comparator observed in Experiment 1. However, Miller and Matute (1996; see also Denniston, Miller, & Matute, 1996; Savastano & Miller, 2003) have reported that when a stimulus acquires biological significance (i.e., control of behavior), reducing responding to the stimulus by altering the associative status of its comparator stimulus is difficult to achieve. Nonetheless, decrements in effective associative value are readily obtained when training and posttraining treatments preclude the target cue from acquiring response potential prior to the posttraining manipulation, as is the case when treatment is embedded within a sensory preconditioning procedure. Therefore, in Experiment 2, we used a sensory preconditioning preparation to test whether combining overshadowing and degraded contingency treatments would have any effect on responding to our target stimulus X. In a sensory preconditioning preparation, a surrogate outcome (O) of low biological significance is used in place of the US during training (e.g., AX-O or X-O) and is subsequently paired with the US (O-US). The rationale for this strategy was twofold. First, it assessed whether the results of Experiment 1 would replicate, now within sensory preconditioning, thereby adding to the generality of the counterintuitive finding of Experiment 1. Second, if the study successfully replicated this finding in a sensory preconditioning preparation, the same parameters could be used in follow-up studies to examine the consequences of posttraining extinction of the context or A. Therefore, in Experiment 2, we used the same 2 × 2 design that was used in Experiment 1, with some parametric changes that were expected to enhance responding when training was embedded in a sensory preconditioning preparation. The parameters used in this experiment are the same as those used by Gunther and Miller (2000) to obtain a degraded contingency effect within a sensory preconditioning preparation.
Method
Subjects and Apparatus
The subjects were 24 male and 24 female, experimentally naïve, Sprague-Dawley-descendent rats obtained from our breeding colony. Body weight ranges were 170–210 g for females and 253–310 g for males. Subjects were randomly assigned to one of four groups (ns = 12), counterbalanced within groups for sex. Subjects were maintained and housed as in Experiment 1. The apparatus was the same as in Experiment 1, with the addition of a white noise that was used as a surrogate outcome in Phase 1 and as a first-order CS in Phase 2 (see Table 2). The white noise was 10 s in duration and approximately 8 dB above background.
Table 2.
Design of Experiment 2
| Group | Phase 1 | Phase 2 | Test |
|---|---|---|---|
| Elem-NDeg | X→O (10) | O→US (8) | X |
| Ov-NDeg | AX→O (10) | O→US (8) | X |
| Elem-Deg | X→O (10)/O (20) | O→US (8) | X |
| Ov-Deg | AX→O (10)/O (20) | O→US (8) | X |
Note. Numbers in parentheses next to the pairings indicate total number of trials in that phase. → = “followed by”; X = click train; A = tone; O = white noise; US = foot shock; Elem = elemental; Ov = overshadowing (compound training); Deg = degraded contingency; NDeg = no degraded contingency.
Procedure
Acclimation
On Days 1 and 2, subjects were acclimated to the test context during 45-min daily sessions. Water-filled lick tubes were available during these two sessions, and no nominal stimulus was presented.
Phase 1
On Days 3 and 4, during daily 60-min sessions in the training context, subjects in Groups Ov-NDeg were exposed to five AX-O trials. Subjects in Condition Elem-NDeg received five X-O trials. Half of the subjects in each of the above-mentioned conditions also received, interspersed among the X-O presentations, 10 daily presentations of O alone, thereby experiencing outcome exposures in an unsignaled manner (Groups Elem-Deg and Ov-Deg). The mean (± SE) intertrial interval was 200 s (± 40 s). The remaining subjects (Groups Elem-NDeg and Ov-NDeg) did not receive these additional outcome presentations. On X-O and AX-O trials, the outcome was presented for 10 s immediately after termination of either X alone or the AX compound.
Phase 2
On each of Days 5 and 6, all subjects received four daily O-US pairings in the test context to endow the surrogate outcome O with biological significance and to allow for the expression of the X-O or AX-O association at test. Trials occurred at 10, 23, 37, and 48 min into each 60-min daily session. The 0.5-s, 0.7-mA shock was presented during the last 0.5 s of the 10-s surrogate outcome (O).
Reacclimation
On Days 7–9, the lick tubes were reinserted, and subjects were allowed to drink water during 60-min daily sessions in the test context.
Testing
On Day 10, all subjects were tested for responding to X in the test context with the same procedure described in Experiment 1. No subject required more than 60 s to complete its initial 5 cumulative seconds of drinking.
Results and Discussion
As shown in Figure 4, Experiment 2 replicated, in a sensory preconditioning preparation, the counteractive effects of overshadowing and degraded contingency obtained in Experiment 1 in a first-order conditioning preparation. The graph shows that these parameters yielded robust overshadowing and degraded contingency, and importantly, these two treatments counteracted each other. The following statistical analyses confirm these observations.
Figure 4.
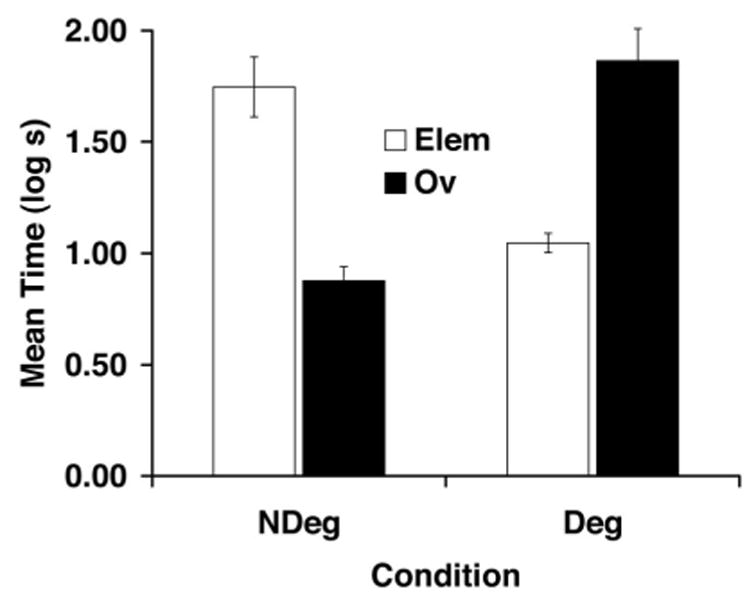
Mean times to complete 5 cumulative seconds of drinking upon presentation of the target CS (X) in Experiment 2. See Table 2 for treatments of the four groups. Elem = elemental; Ov = overshadowing; Deg = degraded contingency; NDeg = no degraded contingency. Error bars denote standard errors of means.
Pre-CS scores from Day 10 were analyzed with a 2 × 2 ANOVA, with stimulus competition (Overshadowing vs. Elemental) and contingency (Degraded Contingency vs. No Degraded Contingency) as factors. This analysis did not detect main effects or interactions, suggesting that the groups did not significantly differ in terms of their fear to the context in which testing took place (all ps > .11).
A similar ANOVA was used to analyze suppression scores during the presentation of the target stimulus X. This analysis did not detect any main effects, but an interaction between the factors was observed, F(1, 44) = 64.04, p < .001, MSE = 0.133. Planned comparisons, with the overall error term from the ANOVA, conducted on the CS suppression scores found that subjects in Groups Ov-NDeg and Elem-Deg suppressed less to the CS than did Group Elem-NDeg, thereby demonstrating simple overshadowing, F(1, 44) = 34.02, p < .05, and simple degraded contingency, F(1, 44) = 22.05, p < .05, respectively. Subjects that received both overshadowing and degraded contingency treatments (Group Ov-Deg) suppressed more than did subjects in Group Ov-NDeg, F(1, 44) = 43.83, p < .05, or Group Elem-Deg, F(1, 44) = 30.08, p < .05, indicating that the response-attenuating effect of either simple overshadowing or simple degraded contingency was reversed in subjects that received both treatments together.
One of the aims of the present study was to replicate the findings observed in Experiment 1, but with training embedded in a sensory preconditioning preparation. At least with the present parameters, these results did replicate the observations of Experiment 1. The present study was also intended to determine parameters for subsequent studies in which we would assess the mechanisms by which these two response-degrading treatments counteract each other. It could be argued that the diminished responding observed in Group Elem-Deg was the result of the surrogate outcome (O) being latently inhibited (Lubow, 1973), rather than CS X being subject to degraded contingency, because O was repeatedly presented during Phase 1. However, this argument is refuted by the observation that Group Ov-Deg experienced the same number of outcome exposures during training, but did not show any decrement in responding at the time of testing, relative to subjects that did not experience the surrogate outcome presentations during training (Elem-NDeg).
Experiment 3
In addition to the results of Experiments 1 and 2, the extended comparator hypothesis predicts that extinguishing either of the two comparator stimuli established during training should restore the down-modulating effect of the other comparator stimulus. That is, posttraining extinction of the context should reveal overshadowing of X by A, and posttraining extinction of A should reveal a degraded contingency effect on X mediated by the context. Testing these predictions was the focal purposes of Experiments 3 and 4. Specifically, Experiment 3 tested whether posttraining extinction of the context, that is, retroactive alleviation of the degraded contingency effect, would lead to a recovery from the counteractive effects of overshadowing and degraded contingency, thereby revealing overshadowing of X. To answer this question, groups that received degraded contingency treatment alone or degraded contingency combined with overshadowing were exposed to either massive (or negligibly short) posttraining context extinction.
Given that it has been observed that when a stimulus acquires biological significance, that stimulus is not easily revaluated through posttraining manipulations of its comparator stimulus, we conducted this study and the following one in a sensory preconditioning preparation in which training and extinction were conducted before the surrogate outcome was made a biologically significant stimulus. In the framework of a 2 × 2 design, subjects were initially assigned to one of two conditions during training (see Table 3). They could experience either simple degraded contingency or degraded contingency combined with overshadowing. After this training phase, half of the subjects in each condition were massively exposed to the training context, whereas the remaining half received a very short exposure to the training context in order to equate handling across groups. The amount of context extinction was determined by prior experiments in our laboratory that studied the effects of context extinction following counteraction between massed spacing and overshadowing treatment (Stout et al., 2003).
Table 3.
Design of Experiment 3
| Group | Phase 1 | Phase 2 | Phase 3 | Test |
|---|---|---|---|---|
| Deg-Elem-NExt | X→O (10)/O (20) | ctx | O→US (8) | X |
| Deg-Ov-NExt | AX→O (10)/O (20) | ctx | O→US (8) | X |
| Deg-Elem-Ext | X→O (10)/O (20) | CTX | O→US (8) | X |
| Deg-Ov-Ext | AX→O (10)/O (20) | CTX | O→US (8) | X |
Note. Numbers in parentheses next to the pairings indicate total number of trials in that phase. → = “followed by”; X = click train; A = tone; O = white noise; US = foot shock; Deg = degraded contingency; Elem = elemental training; Ov = overshadowing (compound training); Ext = extinction; NExt = no extinction; ctx = 5-min daily context exposure; CTX = 150-min daily context exposure.
Method
Subjects and Apparatus
The subjects were 24 female and 24 male, experimentally naïve, Sprague-Dawley-descendent rats obtained from our breeding colony. Body weight ranges were 172–218 g for females and 230–293 g for males. Subjects were randomly assigned to one of four groups (ns = 12), counterbalanced within groups for sex: degraded contingency-elemental training-no extinction (Deg-Elem-NExt), degraded contingency-overshadowing-no extinction (Deg-Ov-NExt), degraded contingency-elemental training-extinction (Deg-Elem-Ext), and degraded contingency-overshadowing-extinction (Deg-Ov-Ext). Subjects were maintained and housed as in Experiment 1. The apparatus was the same as in Experiment 1, except the white noise was added, which, in Experiment 2 and in this experiment, served as the surrogate outcome O in Phase 1 and as a first-order CS in Phase 3.
Procedure
Phases 1 and 2 (training and extinction) were conducted in one context (training context), and all other treatments in the other context (test context).
Acclimation
On Days 1 and 2, subjects were exposed to the test context during 45-min daily sessions and were allowed to freely drink water from the lick tubes.
Phase 1
On Days 3 and 4, during daily 60-min sessions in the training context, subjects in Condition Deg-Elem received 5 X-O trials and, interspersed among them, 10 presentations of the outcome (O) alone each day. Subjects in Condition Deg-Ov experienced 5 daily AX-O trials, and interspersed among them, 10 daily presentations of O. Thus, all subjects were exposed to 10 daily unsignaled outcome presentations; the uniform groups designation Deg is to remind the reader of this. The mean (± SE) intertrial interval was approximately 200 s (± 40 s). On trials that included X, the surrogate outcome O was presented for 10 s immediately after termination of either X alone or the AX compound.
Phase 2
On Days 5–8, subjects in the extinction condition (Groups Deg-Elem-Ext and Deg-Ov-Ext) were exposed daily to the training context for 150 min. Every 30 min, the sound-attenuating enclosures were opened to ensure that the animals were awake. Subjects in the two remaining groups (Deg-Elem-NExt and Deg-Ov-NExt) also were exposed to the training context but only for 5 min per day. No nominal stimulus was presented during this phase.
Phase 3
On Days 9 and 10, all subjects received four daily O-US pairings in the test context to endow the surrogate outcome O with biological significance and to allow for the expression of any existing X-O association at test. Trials occurred at 10, 23, 37, and 48 min into each 60-min daily session.
Reacclimation
The lick tubes were reinserted, and on Days 11 and 12, subjects were allowed to drink during 60-min daily sessions in the test context.
Testing
On Day 13, all subjects were tested for responding to X in the test context. Fear of the target stimulus (X) was evaluated in the same manner as in Experiments 1 and 2. Following the previously described exclusion criterion, one subject from Group Deg-Elem-Ext was eliminated from all analyses.
Results and Discussion
As shown in Figure 5, responding in the two groups, intended to control for the extinction treatment (Groups Deg-Elem-NExt and Deg-Ov-NExt), resembled the behavior of similarly treated groups in Experiment 2. That is, overshadowing treatment reduced the response-attenuating effect of degraded contingency treatment. Of note, this pattern was reversed when the training context underwent extinction. That is, both a recovery from the degraded contingency effect and a reversal of the counteractive effect of degraded contingency on overshadowing resulted from context extinction. These impressions were confirmed by the following analyses.
Figure 5.
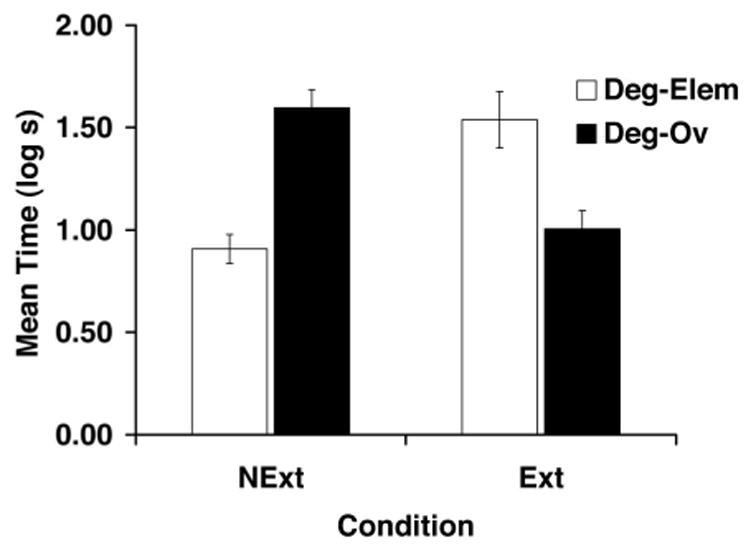
Mean times to complete 5 cumulative seconds of drinking upon presentation of the target CS (X) in Experiment 3. See Table 3 for treatments of the four groups. Deg-Elem = degraded contingency and elemental training; Deg-Ov = degraded contingency and overshadowing; Ext = context extinction; NExt = no extinction. Error bars denote standard errors of means.
A 2 × 2 ANOVA, with training (Deg-Elem vs. Deg-Ov) and extinction (NExt vs. Ext) as factors, performed on the transformed pre-CS scores from Day 13 did not reveal any main effects or an interaction (all ps > .18). A similar ANOVA was performed on the transformed latencies during the presentation of X at test. This analysis did not reveal any main effects but detected an interaction, F(1, 43) = 38.92, p < .001, MSE = 0.112. Planned comparisons, using the overall error term from the 2 × 2 ANOVA, conducted on the CS suppression scores found that subjects in Group Deg-Elem-NExt suppressed less than did subjects in Group Deg-Ov-NExt, F(1, 43) = 25.32, p < .05. Thus, these two groups replicated the counteraction finding of Experiments 1 and 2. Moreover, Group Deg-Elem-Ext suppressed more to X than did Group Deg-Elem-NExt, thereby demonstrating recovery from the degraded contingency effect as a result of posttraining massive context extinction, F(1, 43) = 20.27, p < .05. This demonstration of recovery from the degraded contingency effect as a consequence of posttraining extinction of the training context is consistent with previous observations in the literature (Grahame, Barnet, & Miller, 1992; Kaplan & Hearst, 1985). Of note, subjects in the group trained with overshadowing and degraded contingency treatments responded like a typical overshadowing group only after massive context extinction, confirming the extended comparator hypothesis’s assumption that the counteractive effect of these two treatments occurs at test. This was supported by planned comparisons, which found that Group Deg-Ov-Ext suppressed less to the presentation of X than did Group Deg-Ov-NExt, F(1, 43) = 18.65, p < .05, and Group Deg-Elem-Ext, F(1, 43) = 14.47, p < .05. Thus, release from the effects of degraded contingency produced by context extinction revealed overshadowing.
Experiment 4
Experiment 3 found that, following combined overshadowing treatment and degraded contingency treatment that counteracted each other, extinction of the context (which was shown to attenuate the degraded contingency effect) revealed overshadowing. Experiment 4 was designed to determine whether extinction of the overshadowing stimulus (A) would produce a recovery from simple overshadowing, as previously reported (e.g., Kaufman & Bolles, 1981; Matzel, Schachtman, & Miller, 1985; Savastano & Miller, 2003; but see Holland, 1999). Critically, extinction of the overshadowing stimulus should, like context extinction, undo the counteractive effects of overshadowing and degraded contingency. In this instance, elimination of overshadowing was expected to reveal a degraded contingency effect. Therefore, all groups received overshadowing treatment (AX-O) during training, with half of the subjects also receiving degraded contingency treatment (O alone presentations; see Table 4). After training, the extinction groups experienced 24 daily presentations of the overshadowing stimulus on each of four days, whereas the no-extinction groups were merely handled and exposed to the context daily. We anticipated that subjects for which the overshadowing training was interspersed with unsignaled presentations of the outcome (overshadowing and degraded contingency treatments) would suppress to CS X, but that further experience with the overshadowing stimulus without the outcome (extinction of the overshadowing stimulus A) would produce decreased responding to X, that is, a degraded contingency effect.
Table 4.
Design of Experiment 4
| Group | Phase 1 | Phase 2 | Phase 3 | Test |
|---|---|---|---|---|
| Ov-NDeg-NExt | AX→O (10) | ctx | O→US (8) | X |
| Ov-Deg-NExt | AX→O (10)/O (20) | ctx | O→US (8) | X |
| Ov-NDeg-Ext | AX→O (10) | A (96) | O→US (8) | X |
| Ov-Deg-Ext | AX→O (10)/O (20) | A (96) | O→US (8) | X |
Note. Numbers in parentheses next to the pairings indicate total number of trials in that phase. → = “followed by”; X = click train; A = tone; O = white noise; US = foot shock; Ov = overshadowing (compound training); Deg = degraded contingency; NDeg = no degraded contingency; Ext = extinction; NExt = no extinction; ctx = 18-min daily context exposure; A = 24 daily presentations of A.
Method
Subjects and Apparatus
The subjects were 24 female and 24 male, experimentally naïve, Sprague-Dawley-descendent rats obtained from our breeding colony. Body weight ranges were 174–228 g for females and 225–315 g for males. Subjects were randomly assigned to one of four groups (ns = 12), counterbalanced within groups for sex: overshadowing-no degraded contingency-no extinction (Ov-NDeg-NExt), overshadowing-degraded contingency-no extinction (Ov-Deg-NExt), overshadowing-no degraded contingency-extinction (Ov-NDeg-Ext), and overshadowing-degraded contingency-extinction (Ov-Deg-Ext). Subjects were maintained and housed as in previous experiments. The apparatus was the same as in Experiment 1, with the addition of the white noise used in Experiments 2 and 3.
Procedure
As in Experiment 3, training and extinction were conducted in the training context, and all other treatments were administered in the test context.
Acclimation
During Days 1 and 2, all subjects were exposed to the test context during 45-min daily sessions. During these sessions, subjects were allowed to freely drink water from the lick tubes.
Phase 1
Training was conducted on Days 3 and 4 during daily 60-min sessions. Subjects in Condition Ov-NDeg received 5 AX-O trials, and subjects in Condition Ov-Deg also experienced 5 AX-O trials but interspersed among 10 daily presentations of O. Thus, all four groups received the training trials consisting of compound presentations of the target cue X and the overshadowing stimulus A (Ov). In addition, two of these groups were also exposed to 10 interspersed unsignaled outcome presentations (Deg). The mean (± SE) intertrial interval was approximately 200 s (± 40 s). On AX-O trials, the outcome O was presented for 10 s immediately after termination of the AX compound. The training phase was conducted in the training context, with the lick tubes removed.
Phase 2
On Days 5–8, subjects in the extinction condition (Ext) were exposed daily to 24 presentations of the overshadowing stimulus A within an 18-min session in the training context. The mean (± SE) intertrial interval was approximately 15 s (± 1.5 s). Previous studies (e.g., Savastano & Miller, 2003) have reported that this treatment successfully extinguishes the associative status of the overshadowing stimulus. The rationale for conducting these extinction trials in a massive manner was to extinguish the overshadowing stimulus while avoiding extinction of the context. Subjects in the two remaining groups (NExt) were exposed equally to the training context for 18 min daily, but no nominal stimulus was presented during this phase. Preliminary research determined that four 18-min exposures to a training context are inadequate to attenuate the comparator role of a training context. Thus, during this phase, all animals were handled equally, with the only difference being the extinction of the overshadowing cue A.
Phase 3
On Days 9 and 10, all subjects experienced four daily O-US pairings in testing context to endow the surrogate outcome O with biological significance and to motivate responding to X at test. Trials occurred at 10, 23, 37, and 48 min into each 60-min daily session.
Reacclimation
On Days 11 and 12, the lick tubes were reinserted, and subjects were allowed to drink during 60-min daily sessions.
Testing
On Day 13, all subjects were tested by presenting X and measuring licking suppression as in previous studies. Based on the baseline criterion, no animals were excluded from the statistical analyses.
Results and Discussion
The present study demonstrated that after the training of X in the presence of a more salient stimulus A (overshadowing), extinguishing the overshadowing stimulus A produced a recovery from the decreased responding typically observed after simple overshadowing. More important, when the overshadowing treatment was administered in conjunction with unsignaled presentations of the outcome (O) alone (degraded contingency treatment), the response deficits observed after simple overshadowing and simple degraded contingency training were no longer observed. However, posttraining extinction of the overshadowing stimulus in the latter condition did lead to decreased responding, presumably the unveiling of a degraded contingency effect. These results can be seen in Figure 6 and are confirmed by the following statistical analyses.
Figure 6.
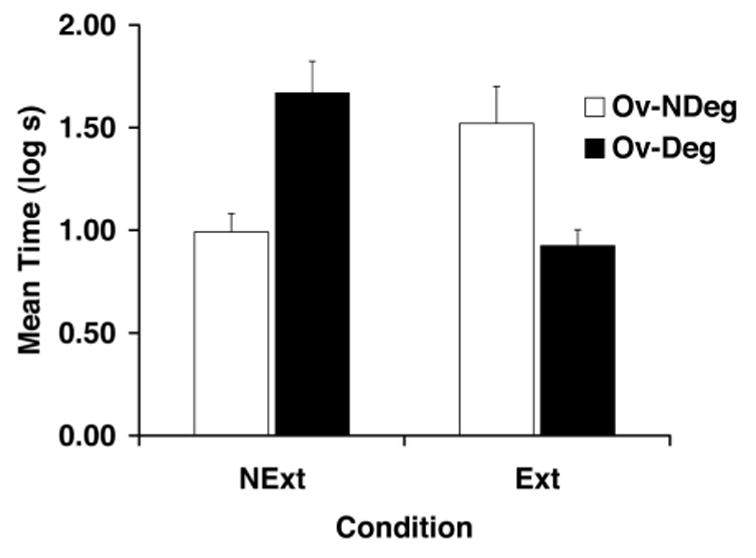
Mean times to complete 5 cumulative seconds of drinking upon presentation of the target CS (X) in Experiment 4. See Table 4 for treatments of the four groups. Ov-NDeg = overshadowing and no degraded contingency; Ov-Deg = overshadowing and degraded contingency; Ext = extinction of the overshadowing stimulus; NExt = no extinction. Error bars denote standard errors of means.
A two-way ANOVA, with training (Ov-NDeg vs. Ov-Deg) and extinction (Ext vs. NExt) performed on the Day 13 baseline scores (i.e., time to complete 5 cumulative seconds of drinking prior to CS onset), revealed no significant main effects or interaction (all ps > .07), indicating no significant differences between groups in terms of fear to test context.
A similar ANOVA conducted on the transformed latencies to start drinking during the presentation of X did not detect any main effects. However, this analysis revealed an interaction between the training (Deg vs. NDeg) and extinction factors, F(1, 44) = 22.77, p < .001, MSE = 0.213. Planned comparisons using the overall error term from the ANOVA found that subjects in Group Ov-NDeg-NExt suppressed less than did subjects in Group Ov-Deg-NExt, F(1, 44) = 12.90, p < .001, indicating that without any posttraining manipulation, we successfully replicated the findings of Experiments 1–3 of the present series. That is, little, if any, overshadowing of X was observed when the AX compound was trained in conjunction with unsignaled presentations of the outcome (O) alone. Subsequent comparisons indicated a recovery from overshadowing as a result of extinguishing the overshadowing stimulus, in that subjects in Group Ov-NDeg-Ext suppressed more than did Group Ov-NDeg-NExt, F(1, 44) = 7.86, p < .01. Additional comparisons indicated that the extinction treatment reestablished the response deficit in the group that was trained with both overshadowing and degraded contingency treatments (Group Ov-Deg-Ext) in that this group suppressed less than did Group Ov-Deg-NExt, F(1, 44) = 15.56, p < .001, and Group Ov-NDeg-Ext, F(1, 44) = 9.96, p < .003. These results are in agreement with the predictions of the extended comparator hypothesis, namely, that the two treatments counteract each other but that if one of the comparator stimuli is extinguished, then the other treatment is revealed as a decrement in responding.
As corroborated by these analyses, the two response-decrementing treatments again counteracted each other, and when trained together, little net decrement in responding was observed. In the present experiment, we also observed a recovery from overshadowing after extinguishing the overshadowing stimulus, confirming previous observations regarding this posttraining manipulation (e.g., Kaufman & Bolles, 1981; Matzel et al., 1985; Savastano & Miller, 2003). The critical aim of this experiment was to determine whether extinguishing the overshadowing stimulus would reverse the robust responding observed in the group that was trained with both overshadowing and degraded contingency treatments. This unique prediction of the extended comparator hypothesis was confirmed, in that less suppression was observed in Group Ov-Deg-Ext in comparison with Group Ov-Deg-NExt. Further, the former group’s magnitude of suppression was similar to the suppression observed in Group Ov-NDeg-Next, thereby showing the expected symmetry in the mechanisms governing the counteractive effect. In summary, this experiment successfully replicated the counteraction previously seen between overshadowing and degraded contingency treatments and further confirms our assumptions concerning the underlying mechanisms governing these counteractive effects.
General Discussion
The present series of experiments investigated the interaction of overshadowing and degraded contingency treatments. Experiment 1 demonstrated in first-order conditioning that the usual deficit in responding to a cue that is observed when the cue is trained in the presence of a more salient stimulus (overshadowing treatment) can be attenuated if, during training, the US is concurrently presented in an unpredictable manner (degraded contingency treatment). Conversely, the usual deficit in responding to a cue that is observed when training occurs in the presence of unsignaled outcomes (degraded contingency treatment) can be attenuated if, during training, a second stimulus is presented in compound with the target cue (overshadowing treatment). Experiment 2 successfully replicated the findings of Experiment 1 embedded in a sensory preconditioning preparation. Considering Experiments 1 and 2 together, it seems that overshadowing and degraded contingency, treatments known for their response decrementing effects, counteract each other. Experiment 3 demonstrated that massive context extinction led to a recovery from degraded contingency. Further, Experiment 3 showed that posttraining extinction of the training context produced a decrease in responding in those subjects that, during training, experienced both overshadowing and degraded contingency treatments. A very similar effect has previously been observed following massed overshadowing training trials, in which the trial massing is assumed to result in a strong context–outcome association just like the unsignaled outcome presentations in the present experiments (Stout et al., 2003). Finally, Experiment 4 showed that extinction of the overshadowing stimulus resulted in what appeared to be a recovery of simple overshadowing, an observation that has previously received empirical support in the literature (e.g., Kaufman & Bolles, 1981; Matzel et al., 1985; Savastano & Miller, 2003). Further, Experiment 4 demonstrated that this posttraining manipulation reversed the counteractive effects of overshadowing and degraded contingency treatments on responding observed when overshadowing and degraded contingency were trained together.
As stated earlier, the present series of experiments was designed to test a prediction of the extended comparator hypothesis (Denniston et al., 2001) that is problematic for contemporary learning models that emphasize processing at the time of training (e.g., Mackintosh, 1975; Pearce & Hall, 1980; Rescorla & Wagner, 1972; Wagner, 1981), even when considering recent revisions that were made to accommodate retrospective revaluation (e.g., Dickinson & Burke, 1996; Van Hamme & Wasserman, 1994). The extended comparator hypothesis (see Figure 1) posits that under the present training conditions, combined overshadowing and degraded contingency treatments should establish the overshadowing stimulus (A) and the context as first-order comparator stimuli for each other, hence lowering the potential of each to down-modulate responding to X. Consequently, the decreased responding observed as a result of either of the treatment alone was expected to be reduced when either comparator has its own effective comparator stimulus. Whereas each of the present experiments provided empirical support for this prediction, Experiments 3 and 4 further demonstrated that extinguishing either of the comparator stimuli reduced responding to the target cue, presumably because extinguishing either of the comparators decreased the tendency of the extinguished comparator to down-modulate the status of the other comparator stimulus. Moreover, symmetrical effects of extinguishing either comparator were observed, adding support for the assumptions of the extended comparator hypothesis with regard to the mechanisms governing responding at the time of testing.
It should be noted that recent revisions of acquisition-focused models can account for the retrospective revaluation data from Experiments 3 and 4 (Dickinson & Burke, 1996; Van Hamme & Wasserman, 1994). That is, these two models were designed to explain learning about a stimulus that is expected but not presented (i.e., retrospective revaluation). Therefore, they can account for the recovery from simple degraded contingency achieved through context extinction that was observed in Experiment 3 and also for the recovery from overshadowing achieved through extinction of the overshadowing stimulus that was observed in Experiment 4. However, these models encounter problems in explaining the counteraction between the degraded contingency and overshadowing treatments that was observed in all of these experiments. The revision of the Rescorla-Wagner model, proposed by Van Hamme and Wasserman (1994), posits that during training, all present cues compete for a limited associative resource that the unconditioned stimulus can support. Applied to the findings of the present series, this model predicts that, with both the context and the overshadowing stimulus (A) being good predictors of the outcome, the target cue (X) should gain less associative strength on each trial than should a cue that is trained in the presence of either the context or the overshadowing stimulus alone. The revision of Wagner’s (1981) SOP (SOP-Revised [SOPR]), proposed by Dickinson and Burke (1996), assumes that stimuli are represented in one of three different states (active states A1 and A2 and inactive state I) and that the simultaneous activation of a stimulus representation and an outcome representation, either both in A1 or both in A2, will cause an excitatory association to be formed between them. Overshadowing is explained as the consequence of the overshadowing cue (A) retrieving the representation of O into state A2, while X is represented in state A1, and the degraded contingency effect is explained as the consequence of the context retrieving the representation of O into state A2 while X is represented in state A1. Hence, SOPR, like SOP, anticipates a summation of response attenuation from combining overshadowing and degraded contingency treatments, which is contrary to what was observed in Experiments 1 and 2. Applied to the results of Experiments 3 and 4, these revised models predict that post-training extinction of A or the context should strengthen the X-O association, thereby enhancing suppression to X, which is the opposite of what was observed.
A possible alternative explanation of the counteractive effects of overshadowing and degraded contingency treatments is that the overshadowing stimulus or the context might have potentiated the acquisition of behavioral control by the overshadowed cue X. Potentiation refers to enhanced behavioral control by a cue X as a result of its being conditioned in the presence of another cue A, relative to X having been trained alone (e.g., Rusiniak, Hankins, Garcia, & Brett, 1979). With respect to the present situation, one could argue that the excitatory context provides a setting in which, instead of overshadowing, the competing cue A potentiates acquisition of behavioral control by the target cue X, or alternatively, the presence of CS A allowed the context to potentiate X. However, four points argue against these possibilities. First, potentiation has been observed primarily between odors and tastes, whereas our target cue was auditory. Second, potentiation appears to depend on the to-be-potentiated cue being weakly conditionable, something that does not seem to be the case for CS X, as seen in the behavior of the Elem-NDeg groups of Experiments 1 and 2. Third, whereas some authors have reported enhanced taste aversion learning as a result of concurrent presentations of shock and toxin (Lasiter & Braun, 1981), which is similar to the present counteractive effect (i.e., greater conditioned suppression) in Experiment 1 (shown to result from the noncontingent foot shocks), this phenomenon cannot explain the counteractive effects observed in Experiments 2, 3, and 4, in which the outcome was not of biological relevance during training of CS X. Although more research is needed to understand the variables that determine when counteraction and release from counteraction will occur, the present results seem to be more parsimoniously explained in terms of the proposed comparator processes rather than in terms of potentiation.
In summary, the present series of experiments investigated the effect of combining overshadowing and degraded contingency treatments. At least under the present circumstances, the treatments counteracted each other rather than summated. These findings, together with other recent studies (Blaisdell et al., 1998; Savastano, Arcediano, et al., 2003; Stout et al., 2003; Urushihara et al., 2004), suggest that behavioral control by a cue is determined at test by the other cues that were present during training. Of note, when there is more than one comparator present at the time of training (in the case of the present experiments, the overshadowing cue and the training context) and these comparator stimuli are associated, they appear to cancel each other, thereby releasing the target cue from interference at the time of testing. Further, extinguishing either of the comparators reestablishes the deficit occasioned by the remaining comparator. To our knowledge, this mechanism is anticipated only by the extended comparator hypothesis.
Footnotes
Gonzalo P. Urcelay and Ralph R. Miller, Department of Psychology, State University of New York at Binghamton.
The CS duration effect refers to the observation that long CSs acquire less behavioral control than do short CSs, and the overshadowing effect refers to attenuated conditioned responding to a target cue X after it is been trained (paired with an unconditioned stimulus [US]) in compound with another cue relative to pairing X by itself with the US.
Support for this research was provided by National Institute of Mental Health Grant 33881. We thank Jeffrey Amundson, Daniel Wheeler, Kouji Urushihara, and Miguel Angel Vadillo for their comments on a draft of this article. We also thank James Esposito and Jessica Fuss for assistance with data collection.
References
- Blaisdell AP, Bristol AS, Gunther LM, Miller RR. Overshadowing and latent inhibition counteract each other: Support for the comparator hypothesis. Journal of Experimental Psychology: Animal Behavior Processes. 1998;24:335–351. doi: 10.1037//0097-7403.24.3.335. [DOI] [PubMed] [Google Scholar]
- Denniston JC, Miller RR, Matute H. Biological significance as a determinant of cue competition. Psychological Science. 1996;7:325–331. [Google Scholar]
- Denniston JC, Savastano HI, Miller RR. The extended comparator hypothesis: Learning by contiguity, responding by relative strength. In: Mower RR, Klein SB, editors. Handbook of contemporary learning theories. Hillsdale, NJ: Erlbaum; 2001. pp. 65–117. [Google Scholar]
- Dickinson A, Burke J. Within-compound associations mediate the retrospective revaluation of causality judgments. Quarterly Journal of Experimental Psychology. 1996;36A:29–50. doi: 10.1080/713932614. [DOI] [PubMed] [Google Scholar]
- Durlach PJ. Effect of signaling intertrial unconditioned stimuli in autoshaping. Journal of Experimental Psychology: Animal Behavior Processes. 1983;9:374–389. [PubMed] [Google Scholar]
- Durlach PJ. The effect of intertrial conditioned stimuli in autoshaping. Quarterly Journal of Experimental Psychology. 1984;36B:353–369. [Google Scholar]
- Grahame NJ, Barnet RC, Miller RR. Pavlovian conditioning in multiple contexts: Competition between contexts for comparator status. Animal Learning & Behavior. 1992;20:329–338. [Google Scholar]
- Gunther LM, Miller RR. Prevention of the degraded-contingency effect by signaling training trials. Quarterly Journal of Experimental Psychology. 2000;53B:97–119. doi: 10.1080/713932719. [DOI] [PubMed] [Google Scholar]
- Holland PC. Overshadowing and blocking as acquisition deficits: No recovery after extinction of overshadowing or blocking cues. Quarterly Journal of Experimental Psychology. 1999;52B:307–333. doi: 10.1080/027249999393022. [DOI] [PubMed] [Google Scholar]
- Jenkins HM, Barnes RA, Barrera FJ. Why autoshaping depends on trial spacing. In: Locurto CM, Terrace HS, Gibbon J, editors. Autoshaping and conditioning theory. New York: Academic Press; 1981. pp. 255–284. [Google Scholar]
- Kaplan PS, Hearst E. Contextual control and excitatory versus inhibitory learning: Studies of extinction, reinstatement, and interference. In: Balsam PD, Tomie A, editors. Context and learning. Hillsdale, NJ; Erlbaum: 1985. pp. 195–224. [Google Scholar]
- Kaufman MA, Bolles RC. A nonassociative aspect of overshadowing. Bulletin of the Psychonomic Society. 1981;18:318–320. [Google Scholar]
- Lasiter PS, Braun JJ. Shock facilitation of taste aversion learning. Behavioral and Neural Biology. 1981;32:277–281. doi: 10.1016/s0163-1047(81)92320-7. [DOI] [PubMed] [Google Scholar]
- Loy I, Hall G. Taste aversion after ingestion of lithium chloride: An associative analysis. Quaterly Journal of Experimental Psychology. 2002;55B:365–380. doi: 10.1080/02724990244000070. [DOI] [PubMed] [Google Scholar]
- Lubow RE. Latent inhibition. Psychological Bulletin. 1973;79:398–407. doi: 10.1037/h0034425. [DOI] [PubMed] [Google Scholar]
- Mackintosh NJ. A theory of attention: Variations in the associability of stimuli with reinforcement. Psychological Review. 1975;82:276–298. [Google Scholar]
- Matzel LD, Schachtman TR, Miller RR. Recovery of an overshadowed association achieved by extinction of the overshadowing stimulus. Learning and Motivation. 1985;16:398–412. [Google Scholar]
- Miller RR, Matute H. Biological significance in forward and backward blocking: Resolution of a discrepancy between animal conditioning and human causal judgment. Journal of Experimental Psychology: General. 1996;12(4):370–386. doi: 10.1037//0096-3445.125.4.370. [DOI] [PubMed] [Google Scholar]
- Pearce JM. A model for stimulus generalization in Pavlovian conditioning. Psychological Review. 1987;94:61–73. [PubMed] [Google Scholar]
- Pearce JM, Hall G. A model for Pavlovian learning: Variations in the effectiveness of conditioned but not of unconditioned stimuli. Psychological Review. 1980;87:532–552. [PubMed] [Google Scholar]
- Rescorla RA. Predictability and number of pairings in Pavlovian fear conditioning. Bulletin of the Psychonomic Society. 1966;4:383–384. [Google Scholar]
- Rescorla RA. Probability of shock in the presence and absence of CS in fear conditioning. Journal of Comparative and Physiological Psychology. 1968;66:1–5. doi: 10.1037/h0025984. [DOI] [PubMed] [Google Scholar]
- Rescorla RA, Wagner AR. A theory of Pavlovian conditioning: Variations in the effectiveness of reinforcement and nonreinforcement. In: Black AH, Prokasy WF, editors. Classical conditioning II: Current research and theory. New York: Appleton-Century-Crofts; 1972. pp. 64–99. [Google Scholar]
- Rusiniak K, Hankins W, Garcia J, Brett L. Flavor-illness aversions: Potentiation of odor by taste in rats. Behavioral and Neural Biology. 1979;25:1–17. doi: 10.1016/s0163-1047(79)90688-5. [DOI] [PubMed] [Google Scholar]
- Savastano HI, Arcediano F, Stout SC, Miller RR. Interaction between preexposure and overshadowing: Further analysis of the extended comparator hypothesis. Quarterly Journal of Experimental Psychology. 2003;56B:371–395. doi: 10.1080/02724990344000006. [DOI] [PubMed] [Google Scholar]
- Savastano HI, Miller RR. Biological significance and posttraining changes in conditioned responding. Learning and Motivation. 2003;34:303–324. [Google Scholar]
- Stout SC, Chang R, Miller RR. Trial spacing is a determinant of cue interaction. Journal of Experimental Psychology: Animal Behavior Processes. 2003;29:23–28. [PubMed] [Google Scholar]
- Stout SC, Miller RR. Sometimes opponent retrieval (SOCR): A mathematical implementation of the extended comparator hypothesis. 2005. Manuscript submitted for publication. [Google Scholar]
- Urushihara K, Stout SC, Miller RR. The basic laws of conditioning differ for elemental cues and cues trained in compound. Psychological Science. 2004;15(4):268–271. doi: 10.1111/j.0956-7976.2004.00664.x. [DOI] [PubMed] [Google Scholar]
- Van Hamme LJ, Wasserman EA. Cue competition in causality judgments: The role of nonpresentation of compound stimulus elements. Learning and Motivation. 1994;25:127–151. [Google Scholar]
- Wagner AR. SOP: A model of automatic memory processing in animal behavior. In: Spear NE, Miller RR, editors. Information processing in animals: Memory mechanisms. Hillsdale, NJ: Erlbaum; 1981. pp. 5–47. [Google Scholar]