

Abstract
Cnidarian envenomations cause a burning-pain sensation of which the underlying mechanisms are unknown. Activation of TRPV1, a non-selective cation channel expressed in nociceptive neurons, leads to cell depolarisation and pain. Here, we show in vitro and in vivo evidence for desensitization-dependent TRPV1 activation in cnidarian envenomations. Cnidarian venom induced a nociceptive reactivity, comparable to capsaicin, in laboratory rats, which could be reduced by the selective TRPV1 antagonist, BCTC. These findings are the first to explain at least part of the symptomology of cnidarian envenomations and provide insights into the design of more effective treatments for this global public health problem.
Keywords: Cnidaria, TRPV1, envenomation, pain, desensitization
1. Introduction
Increased frequency and magnitude of jellyfish blooms worldwide, together with increased occupational and recreational exposure of at-risk coastal populations to cnidarian envenomations, underscore the need for better clinical-management tools for the treatment of such stings. Limited success in post-envenomation pain relief has been achieved with the empirical use of topical vinegar or hot-water immersion [1–3]. However, currently available clinical-care modalities, based on symptomatic treatment of resultant erythema, burning pain, hypersensitivity and inflammation [4–6], generally fail to ameliorate suffering. Thus, the inadequate understanding of the principal pathophysiological pathways initiated by envenomation has been a major impediment to developing more targeted and effective treatments. Since some members of the TRP superfamily are known to be heat and pH sensitive [7–12], we tested whether TRPV1 activation was responsible for the painful symptoms associated with cnidarian envenomation.
2. Materials and Methods
2.1 Preparation of crude venom extracts
Tentacles, from Aiptasia pulchella, Cyanea capillata, Physalia physalis and Chironex fleckerii, were cut into small pieces and suspended in 50–60 mL 10% acetic acid per 1.5-2 g of tentacles. The mixture was stirred overnight at room temperature with a magnetic stirrer, then centrifuged at 40,000 g for 1 hour. Nearly all of the supernatant was recovered by careful decantation or removal by a syringe. The supernatant was concentrated by rotatory evaporation to about 10% of its original volume, diluted with 5-6 volumes of deionized water and freeze-dried. Freeze-dried samples were dissolved again in ND96. Solutions were titrated to pH 7.4 with NaOH
2.2 Electrophysiological recordings
cRNA transcripts were synthesized from XbaI-linearized VR1 cDNA templates using T7 RNA polymerase (Ambion). Oocytes, harvested from anaesthetized female Xenopus laevis frogs as previously described [13], were injected with 0.5–5 ng TRPV1 cRNA. Two to seven days after injection, two-electrode voltage-clamp recording was performed. Currents were measured in ND96 solution using a protocol of −90 mV during 400 s. Tail current experiments in figure 3 were measured at a holding of 0 mV, test steps from −100 to +160 mV for 200 ms. The recording chamber was perfused at a rate of 2 mL min−1 with a ND-96 solution containing (in mM) 96 NaCl, 2 KCl, 1.8 CaCl2, 1 MgCl2, 5 HEPES, pH 7.4. Temperature of the perfusate was kept at 22°C and controlled using a SC-20 dual in-line heater/cooler (Warner Instruments) and pH was kept at 7.4. As previously described [7], capsaicin (2 μM) was used as an agonist and capsazepine (10 μM) as an antagonist of TRPV1. Capsaicin and capsazepine were purchased from Sigma, and anandamide from Tocris. In in vitro experiments, following venom concentrations are used: 150 mg crude freez-dried Cyanea capillata venom was dissolved in 50ml ND96; 215 mg freez-dried Physalia physalis was dissolved in 50 ml ND96; 170 mg freez-dried Chironex fleckeri venom was dissolved in 30 ml ND96 and few hundred micrograms of crude Cyanea capillata venom was dissolved is 5 ml ND96. 10 mg BmK venom was dissolved in 40 ml ND96.
Fig. 3. Voltage dependence of the open probability of TRPV1 influenced by capsaicin and capsaicin with Cyanea capillata venom on the (n+1) activation.
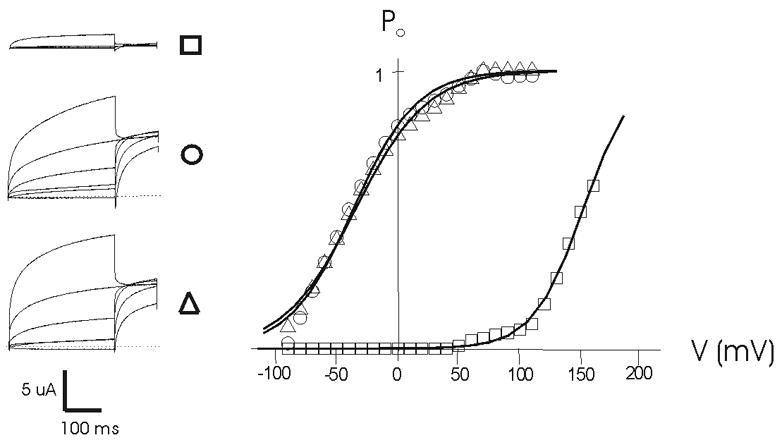
Left: Voltage step currents in ctrl (□) condition, CAP (Î) and CAP with venom (Δ). Right: Open probabilty (Po) calculated by fitting the tail current amplitudes in the voltage step programme (holding 0 mV, test steps from −100 to +160 mV, increment 10 mV). Note that there is no shift between capsaicin and capsaicin with venom conditions.
To control the expression level of the TRPV1 channels, we always started our experiments with a first activation of TRPV1 with 2 uM capsaicin. Application of capsaicin (or anandamide) opens the channels. This activation gives an inward non-selective cation current.
2.3 In vivo studies
Male Sprague Dawley rats (Harlan, Eystrup, Germany) weighing between 270 and 310 g, were maintained in a climate-controlled environment on a 12 hr light/dark cycle at a temperature of 22 ± 1°C. All experiments were carried out during the light phase. During housing water and food were available ad libitum. The animals were habituated to the experimental room for at least 1 hr before the start of the experiment. Animals were used only once. All tests were performed according to guidelines of the Institutional Ethical Committee for Animal Experiments and guidelines for animal research according to IASP. In in vivo experiments, 75 mg was dissolved in 0.5 ml sterile water and a dose of 40 mg/kg was injected.
The formalin test is a chemical assay of nociception (Dubuisson and Dennis, 1977). At 30 min after sc injection of the test compound rats were given a single injection of formaldehyde (50μl, 1.75%) in the plantar hypodermis of the skin of the left hind paw. Within seconds the rats started to flinch the paw rapidly, this is the phase I response that is measured during the first 10 min. The following phase II is measured during 50 min and represents a more tonic state. This biphasic flinch behaviour in the rat following the formalin injection was measured in an Automated Nociception Analyser® (UC, San Diego, CA, USA). Therefore the animal was placed in a Plexiglas cylinder mounted above a transmitter/receiver coils assembly. The system detects movement of a small metal band placed on the injected paw. A signal is generated as the band breaks the electromagnetic field of a loop antenna placed underneath the rat. The signal is processed through an algorithm that determines flinch activity using amplitude, zero voltage crossing and duration. The total number of flinches observed during a selected time period was calculated by accumulating flinches of each individual animal over that time period and averaging the treatment groups. Activity of the compound was considered when the number of flinches was lower than 40 or 600 during phases I and II, respectively.
3. Results
We tested venom extracts (10% acetic acid tentacular extracts) of four cnidarian species (Aiptasia pulchella, Chironex fleckeri, Physalia physalis and Cyanea capillata), representing each of the four classes in the phylum Cnidaria (Anthozoa, Cubozoa or sea wasps, Hydrozoa and Scyphozoa or jellyfishes). Although none of the four venoms tested had any significant effect of its own (Fig. 1A–D left), there was a clear allosteric effect on the [(n+1) with (n>0)] activation of the channel when the venom was applied with capsaicin (Fig. 1A–D right). We then tested whether the endogenous activator, anandamide [14, 15] also induced this effect. That anandamide (10 μM) elicits the same allosteric effect (Fig. 1F) suggests future studies to further test the in vivo interplay of serum anandamide and venom constituents at the site of envenomation. Because stings of the scorpion Buthus martensi Karsch (BmK), a very well-studied preparation, are also accompanied by symptoms of redness and burning pain [16, 17], we compared the effect of crude BmK venom under the same conditions, and found no effect on TRPV1 (Fig. 1E), indicating that the allosteric effect might be specific for cnidarian venoms. Crude venom of the green mamba (Dendroapsis angusticeps) also did not have any effect on TRPV1 (data not shown).
Fig. 1. Effects of Cnidaria and BmK venom on TRPV1.

Traces from A. Aiptasia pulchella (A.P.) venom B. Chironex fleckeri (C.F.) venom C. Physalia physalis (P.P) venom D. Cyanea capillata (C.C.) venom. A–D Left: Crude venom in the absence of capsaicin (CAP) and capsazepine (CZP). A–D Right: Allosteric effect of venom with 2 μM capsaicin. E Left: Trace with lack of effect of BmK venom. E Right: Lack of effect of venom in the presence of capsaicin. F. Allosteric effect of Cyanea capillata venom and 10 uM anandamide (ANA). Identical results were obtained independent of the order of CAP or venom administration.
To clarify the mechanism of action of the venom and to investigate whether the observed venom effects were related to desensitization of TRPV1 [18, 19], the effect of the venom at the peak current from the first and consecutive activations was tested. Desensitisation of TRPV1 can be described as channels becoming insensitive by repeated or continuous addition of an agonist (e.g. capsaicin). In voltage-clamp measurements it can be seen as smaller inward currents every application or during the application of the agonist without addition of an antagonist. By applying the venom on the first peak current, we ensured that there was virtually no desensitization of the channel (a situation comparable to Ca2+-free condition [18, 19]). If the venom effect was indeed linked to desensitization, there should be no apparent effect of the venom on the peak current. If the venom was applied after the desensitization process had started or on the (n+1) activation of the TRPV1 channel, a desensitization-dependent venom effect would be expected. The results of these experiments indicated that the venom was indeed without apparent effect during the first activation peak of TRPV1 (Fig. 2A). By contrast, the allosteric effect in which the desensitization-dependent component is being restored by the venom was clearly present when the venom was applied during the desensitization phase or at the (n+1) activation (Fig 2B and 2C). Furthermore, we investigated if the venom could shift the TRPV1 activation curve (a frequently used trick of TRP channels [20]) and found that the open probability (Po) calculated by fitting the tail current amplitudes was not significantly shifted along the voltage axis in the presence of capsaicin and venom as compared to capsaicin alone (Fig. 3). As such, these findings clearly indicate that the venom knocks down the desensitization of TRPV1, thus resulting in larger inward currents that would generate, in turn, the typical persistent burning-pain sensation.
Fig. 2. TRPV1 desensitization knocked down by Cyanea capillata venom.

A. TRPV1 current during the first activation. The venom is added on the peak of this activation. B. Current during the first activation when the venom is added just at the time when desensitization is macroscopically visible. It can be seen that the venom quickly knocks down desensitization (see arrow). C. Trace showing the first and second activation of the channel and the effect of the venom when added during the second activation. Note that the venom restores exactly the desensitization-dependent fraction of the inward current (see arrow).
To test the in vivo effects of cnidarian venoms and the possible therapeutic use of TRPV1 antagonists, we recorded the number of flinches for 4 minutes after intraplantar injection of capsaicin or venom in laboratory rats. The injected dose of crude Cyanea capillata venom was chosen based on the number of flinches that was comparable with the number of flinches seen with 10 μg capsaicin. Fig. 4A and 4B show a significant increase of flinches when compared with the response after intraplantar injection of vehicle. To test whether a TRPV1 antagonist could inhibit the induced nociceptive reactions, 40 mg/kg of 4-(3-trifluoromethylpyridin-2-yl)piperazine-1-carboxylic acid (5-trifluoromethylpyridin-2-yl)amide (BCTC), a high-affinity TRPV1 antagonist [21], was injected subcutaneously 1 hour before intraplantar injection of capsaicin or Cyanea capillata venom. As shown in Fig. 4A, a significant decrease in mean pain reaction (from 76.0 to 4.2, p<0.05, n=6) was observed when rats were injected with capsaicin, following pretreatment with BCTC. BCTC also significantly decreased the crude venom-induced mean number of flinches (from 65.8 to 14.0, p<0,05, n=6) (Fig. 4B). Injection of control vehicle did not significantly affect the reaction after intraplantar capsaicin or venom injection. The dose needed to decrease the venom-induced number of flinches by 50% ranged between 10 – 40 mg/kg (data not shown). The analgesic effect of BCTC was maximal at a dose of 40 mg/kg, with no further decrease in flinches when higher doses were injected. In all cases of capsaicin and venom, there was a baseline effect of a few flinches over the 4-minute observation period. This indicates that other channels may also be involved in the induction of the nociceptive behaviour, although their relative contribution is presumably insignificant since the induced pain effect was reduced by 79% with BCTC, a selective TRPV1 antagonist.
Fig. 4. In vivo effect of Cyanea capillata venom with and without TRPV1 antagonist.
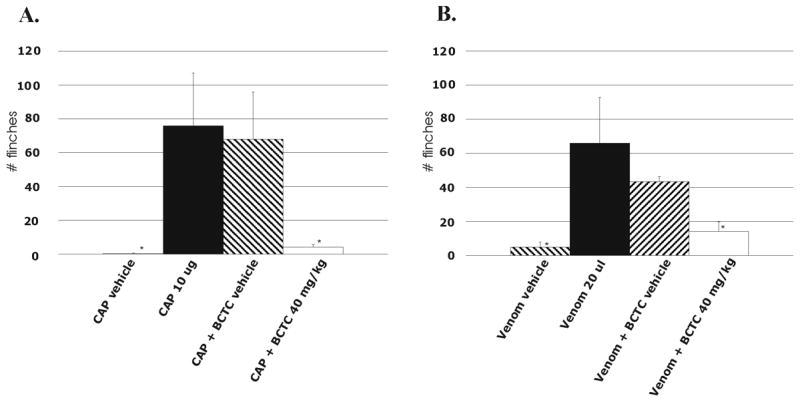
A. Number of flinches during 4-minute observation period after left-hindpaw plantar injection of capsaicin (black) and vehicle (left diagonals). Pretreatment by subcutaneous injection of 40 mg/kg BCTC (white) or vehicle (right diagonals) and 1 hour later the number of flinches and bites were observed after an intraplantar injection of capsaicin (n=6). The effect of capsaicin with BCTC was significantly different from the effect of capsaicin alone. The BCTC vehicle did not give a significant difference as compared with capsaicin alone. B. Same tests as in A. Here, the effect of the venom was tested instead of capsaicin. The effect of venom with BCTC was significantly different from the effect of venom alone. The BCTC vehicle did not give a significant difference as compared with venom alone. In these experiments, the serum concentrations of anandamide or other endogenous TRPV1 activators were not determined.
Additional rat tests in which BCTC treatment is tested on a non-TRPV1 mediated pain behaviour (formalin test), BCTC did not show any analgesic effect. In phase I (first 10 min observation period) as well as in phase II (50 min observation) there was no significant decrease in pain response of the BCTC treated rats. In addition, BCTC did not show any side effects such as lethargy that may affect the flick/lick assay.
Taken together, these in vivo results do not give 100% prove that TRPV1 is the main pain inducing mechanism, but keeping in mind the experiences in the empiric treatment, it gives us a good idea in what might be happening in the in vivo situations and they provide supportive evidence that the use of TRPV1 antagonists as analgesics in cnidarian envenomations warrants consideration for clinical trials.
4. Discussion
In conclusion, we identified TRPV1 as a key component in the signal-transduction pathway of cnidarian envenomation. These newfound data provide a pathophysiologic basis for the symptomology of cnidarian stings. Although the active substance(s) in these venoms is (are) yet not fully identified (i.e., whether peptide or bioactive small molecule) and the precise site of interaction still remains to be elucidated (i.e., on the TRPV1 itself vs. the signal-transduction pathway), our discovery provides important and novel insights into the design of more effective treatments for cnidarian envenomations: on the one hand, TRPV1 blockers can possibly be used as therapeutics, just as atropine is used as an antidote for organophosphate-type intoxications on the muscarinic ACh receptor [22]; on the other hand, one might speculate that TRPV1 activators may also be useful in the treatment of cnidarian stings as they could counteract the venom-induced down regulation of the desensitization. As a consequence, an inverse relationship exists between the degree of desensitization and the size of the inward currents. As a corollary, therefore, the burning-pain sensation should diminish once the inward currents become smaller. It is clear that further studies are needed to pinpoint the selectivity ratio of cnidarian venoms between the different members of the large TRP family and to demonstrate if the human TRPV1 orthologue behaves in the same manner as the rat clone. Nonetheless, our in vitro and in vivo experiments have shown for the first time a clear link between cnidarian envenomation-associated pain and TRPV1 activation via ‘knock-down’ of desensitization that brings more effective treatments for the worldwide problem of cnidarian envenomations within reach.
Acknowledgments
We thank Dr. Carina Östman for providing tentacles of Cyanea capillata, Dr. Olga Castaneda for Physalia physalis and Dr. Phil Alderslade for Chironex fleckeri. We also thank Dr. David Julius for providing the rat TRPV1 clone, and we are very grateful to Dr. Theo Meert, Dr. Hilde Vermeirsch, Ria Biermans (J&J PRD, Belgium) for their support of the in vivo studies. Finally, we thank Sarah Debaveye for making the TRPV1 RNA and Dr. Frank Bosmans, Dr. Chantal Maertens, Dr. Elke Vermassen, Yousra Abdel-Mottaleb, Thomas Vandendriessche, Bert Billen and Dr. Richard Yanagihara for their critical reading of the manuscript.
This work was supported in part by the FWO-Vlaanderen grant G.0330.06; the K.U.Leuven (grant OT-05-64); and grants from the National Institute of Neurological Disorders and Stroke, National Institute of Health (US54NS039406) and the Hawaii Community Foundation (20011908).
Footnotes
This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Yoshimoto CM, Yanagihara AA. Cnidarian (coelenterate) envenomations in Hawai'i improve following heat application. Trans R Soc Trop Med Hyg. 2002;96(3):p. 300–3. doi: 10.1016/s0035-9203(02)90105-7. [DOI] [PubMed] [Google Scholar]
- 2.Bailey PM, et al. Jellyfish envenoming syndromes: unknown toxic mechanisms and unproven therapies. Med J Aust. 2003;178(1):p. 34–7. doi: 10.5694/j.1326-5377.2003.tb05040.x. [DOI] [PubMed] [Google Scholar]
- 3.Kumar S, et al. Intravenous ascorbic acid as a treatment for severe jellyfish stings. P R Health Sci J. 2004;23(2):p. 125–6. [PubMed] [Google Scholar]
- 4.Burnett JW, Calton GJ. Venomous pelagic coelenterates: chemistry, toxicology, immunology and treatment of their stings. Toxicon. 1987;25(6):p. 581–602. doi: 10.1016/0041-0101(87)90105-x. [DOI] [PubMed] [Google Scholar]
- 5.Walker MJ. Pharmacological and biochemical properties of a toxin containing material from the jellyfish, Cyanea capillata. Toxicon. 1977;15(1):p. 3–14. doi: 10.1016/0041-0101(77)90064-2. [DOI] [PubMed] [Google Scholar]
- 6.Walker MJ. The cardiac actions of a toxin-containing material from the jellyfish, Cyanea capillata. Toxicon. 1977;15(1):p. 15–27. doi: 10.1016/0041-0101(77)90065-4. [DOI] [PubMed] [Google Scholar]
- 7.Caterina MJ, et al. The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature. 1997;389(6653):p. 816–24. doi: 10.1038/39807. [DOI] [PubMed] [Google Scholar]
- 8.Clapham DE. TRP channels as cellular sensors. Nature. 2003;426(6966):p. 517–24. doi: 10.1038/nature02196. [DOI] [PubMed] [Google Scholar]
- 9.Voets T, et al. Sensing with TRP channels. Nat Chem Biol. 2005;1(2):p. 85–92. doi: 10.1038/nchembio0705-85. [DOI] [PubMed] [Google Scholar]
- 10.Numazaki M, Tominaga M. Nociception and TRP Channels. Curr Drug Targets CNS Neurol Disord. 2004;3(6):p. 479–85. doi: 10.2174/1568007043336789. [DOI] [PubMed] [Google Scholar]
- 11.Patapoutian A. TRP Channels and Thermosensation. Chem Senses. 2005;30(Suppl 1):p. i193–i194. doi: 10.1093/chemse/bjh180. [DOI] [PubMed] [Google Scholar]
- 12.Voets T, et al. The principle of temperature-dependent gating in cold- and heat-sensitive TRP channels. Nature. 2004;430(7001):p. 748–54. doi: 10.1038/nature02732. [DOI] [PubMed] [Google Scholar]
- 13.Liman ER, Tytgat J, Hess P. Subunit stoichiometry of a mammalian K+ channel determined by construction of multimeric cDNAs. Neuron. 1992;9(5):p. 861–71. doi: 10.1016/0896-6273(92)90239-a. [DOI] [PubMed] [Google Scholar]
- 14.Smart D, Jerman JC. Anandamide: an endogenous activator of the vanilloid receptor. Trends Pharmacol Sci. 2000;21(4):p. 134. doi: 10.1016/s0165-6147(00)01459-0. [DOI] [PubMed] [Google Scholar]
- 15.Van Der Stelt M, Di Marzo V. Endovanilloids. Putative endogenous ligands of transient receptor potential vanilloid 1 channels. Eur J Biochem. 2004;271(10):p. 1827–34. doi: 10.1111/j.1432-1033.2004.04081.x. [DOI] [PubMed] [Google Scholar]
- 16.Polis GA. The biology of scorpions. Stanford: Stanford university press; 2002. [Google Scholar]
- 17.Otero R, et al. Scorpion envenoming in two regions of Colombia: clinical, epidemiological and therapeutic aspects. Trans R Soc Trop Med Hyg. 2004;98(12):p. 742–50. doi: 10.1016/j.trstmh.2003.12.018. [DOI] [PubMed] [Google Scholar]
- 18.Mohapatra DP, Nau C. Desensitization of capsaicin-activated currents in the vanilloid receptor TRPV1 is decreased by the cyclic AMP-dependent protein kinase pathway. J Biol Chem. 2003;278(50):p. 50080–90. doi: 10.1074/jbc.M306619200. [DOI] [PubMed] [Google Scholar]
- 19.Numazaki M, et al. Structural determinant of TRPV1 desensitization interacts with calmodulin. Proc Natl Acad Sci U S A. 2003;100(13):p. 8002–6. doi: 10.1073/pnas.1337252100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Nilius B, et al. Gating of TRP channels: a voltage connection? J Physiol. 2005;567(Pt 1):p. 35–44. doi: 10.1113/jphysiol.2005.088377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Swanson DM, et al. Identification and biological evaluation of 4-(3-trifluoromethylpyridin-2-yl)piperazine-1-carboxylic acid (5-trifluoromethylpyridin-2-yl)amide, a high affinity TRPV1 (VR1) vanilloid receptor antagonist. J Med Chem. 2005;48(6):p. 1857–72. doi: 10.1021/jm0495071. [DOI] [PubMed] [Google Scholar]
- 22.Bajgar J. Organophosphates/nerve agent poisoning: mechanism of action, diagnosis, prophylaxis, and treatment. Adv Clin Chem. 2004;38:p. 151–216. doi: 10.1016/s0065-2423(04)38006-6. [DOI] [PubMed] [Google Scholar]