

Abstract
Objective
Midbrain dopamine transmission is thought to regulate responses to rewarding drugs and drug-paired stimuli; however, the exact contribution, particularly in humans, remains unclear. In the present study, we tested whether decreasing dopamine synthesis, as produced by acute phenylalanine/tyrosine depletion (APTD), would alter responses to the stimulant drug, d-amphetamine.
Methods
On 3 separate days, 14 healthy men received d-amphetamine (0.3 mg/kg, given orally) plus a nutritionally balanced amino acid mixture, the phenylalanine/tyrosine-deficient mixture or the phenylalanine/tyrosine-deficient mixture followed by the immediate dopamine precursor, L-DOPA (Sinemet, 2 × 100 mg/25 mg). Responses to these treatments were assessed with visual analog scales, the Profile of Mood States, and a computerized Go/No-Go task.
Results
d-Amphetamine elicited its prototypical subjective effects, but these were not altered by APTD. In comparison, APTD significantly increased commission errors on the Go/No-Go task and did so uniquely in conditions where subjects were rewarded for making correct responses; this effect of APTD was prevented by L-DOPA.
Conclusions
Together these results support the hypothesis that, in healthy men, dopamine is not closely linked to euphorogenic effects of abused substances but does affect the salience of reward-related cues and the ability to respond to them preferentially.
Medical subject headings: dopamine, incentive motivation, reward, mood, addiction, acute phenylalanine / tyrosine depletion
Abstract
Objectif
On croit que la transmission de la dopamine dans le mésencéphale assure la régulation des réponses aux drogues qui procurent une satisfaction et aux stimulis associés aux drogues, mais sa contribution exacte n'est toujours pas claire, en particulier chez les êtres humains. Dans cette étude, nous avons cherché à déterminer si une baisse de la synthèse de la dopamine causée par une déplétion aiguë de la phénylalanine-tyrosine (acute phenylalanine/tyrosine depletion ou APTD) modifierait les réponses à une drogue stimulante, la d-amphétamine.
Méthodes
Quatorze hommes en santé ont reçu, pendant trois jours distincts, de la d-amphétamine (0,3 mg/kg, par voie orale) plus un mélange d'acides aminés carencé en phénylalanine-tyrosine ou un mélange d'acides aminés carencé en phénylalanine-tyrosine suivi d'un précurseur immédiat de la dopamine, le L-DOPA (Sinemet, 2 × 100 mg/25 mg). On a évalué les réponses à ces traitements au moyen d'échelles analogiques visuelles, du Profile of Mood States et d'une tâche informatisée oui ou non.
Résultats
La d-amphétamine a produit ses effets subjectifs prototypes que l'APTD n'a toutefois pas modifiés. En guise de comparaison, l'APTD a augmenté de façon significative les erreurs commises au cours de l'exécution de la tâche oui ou non et l'a fait uniquement dans des conditions où les sujets ont été récompensés pour leur bonne réponse. Le L-DOPA a bloqué cet effet de l'APTD.
Conclusions
Ces résultats appuient globalement l'hypothèse selon laquelle chez des hommes en bonne santé, il n'y a pas de lien étroit entre la dopamine et les effets euphorigènes de substances dont il est fait abus, mais elle a un effet sur la prégnance des indices reliés à la satisfaction et sur la capacité d'y réagir de façon préférentielle.
Introduction
Stimulant drug-induced increases in mesocorticolimbic dopamine (DA) transmission are thought to affect responses to rewards. Still, controversy remains. Based on studies conducted in experimental animals, compelling arguments have been made that DA mediates the pleasure associated with reward,1 the impetus to seek rewards,2,3 the ability of reward-related cues to elicit and sustain interest,4–8 the expectation of reward,9 the judgement as to whether the reward was better or worse than expected,9,10 the selection of attention to and behaviours directed toward the reward11–13 and the learning of associations between rewards and their predictive cues.14–18
Recently, we have begun studies on the role of DA in drug reward in humans, using the acute phenylalanine/tyrosine depletion (APTD) method. In research animals, APTD decreases stimulated DA release19–21 and cFos activation,22 as well as striatal tissue concentrations of DA23 and cerebrospinal fluid levels of the DA metabolite, homovanillic acid.24 APTD might also decrease norepinephrine (NE) synthesis,19,24 but this does not appear to be associated with diminished NE release.19,22,25,26 In humans, tyrosine depletion decreases resting27 and amphetamine-induced striatal DA release,28 increases plasma levels of prolactin,29–32 disrupts spatial working memory29–31,33 (though see32,34–36), decreases alcohol self-administration,37,38 and alters the ability to adjust betting behaviour in a gambling task.36,39,40
Effects of APTD on subjective states have been more variable. Tyrosine depletion reduces manic symptoms in patients with a bipolar mood disorder,41,42 craving responses to cocaine and cocaine cues43 and psychostimulant effects of amphetamines41,44; however, although APTD has also been reported to elicit mild mood-lowering responses associated with boredom and apathy,39,45 marked changes in mood have not been seen. APTD does not reinstate depressive symptoms in recovered patients with a history of major depression,31,40 alter anxiogenic effects of stressors,45,46 or diminish the mood-elevating effects of cocaine,43 alcohol37,38 or nicotine.47 In the present study, we investigated whether APTD would decrease mood-elevating effects of d-amphetamine and behavioural responses to reward stimuli, using a computerized Go/No-Go task; effects of APTD, it was proposed, would be prevented by administrating the immediate DA precursor, L-DOPA.
Methods
Participants
We recruited 14 healthy men aged 24.1 years (mean, standard deviation [SD] 4.4 yr) from advertisements placed in local newspapers and on campus. All were healthy nonsmokers, as determined by a physical exam, standard laboratory tests, and an interview with the Structured Clinical Interview for DSM-IV,48 axis I.49 None had a first-degree relative history of axis I psychiatric disorders, as assessed by the Diagnostic Interview for Genetic Studies.50 On each study day, all men tested negative on a urine drug screen sensitive to cocaine, opiates, phencyclidine, barbiturates, delta-9-tetrahydrocannabinol, benzodiazepines, and amphetamines (Triage Panel for Drugs of Abuse, Biosite Diagnostics, San Diego, Calif., US). The study was carried out in accordance with the Declaration of Helsinki and was approved by the Institutional Review Board of McGill University's Faculty of Medicine. All participants gave written informed consent. Go/No-Go data were missing for one participant.
Procedure
Administration of the amino acid (AA) mixtures was conducted double blind in a randomized, within subjects, counterbalanced design (Fig. 1). The day before each test session, participants ate a low-protein diet provided by the investigators and fasted from midnight. On the test day, participants arrived at 8:30 am and had blood samples drawn to measure plasma AA concentrations. They then ingested one of the AA mixtures. The APTD mixture's composition, preparation and administration are based on our 100 g nutritionally balanced mixture, with phenylalanine and tyrosine withheld.45,51 After ingesting the mixture, participants remained awake in a room with neutral videos and reading material available to them.
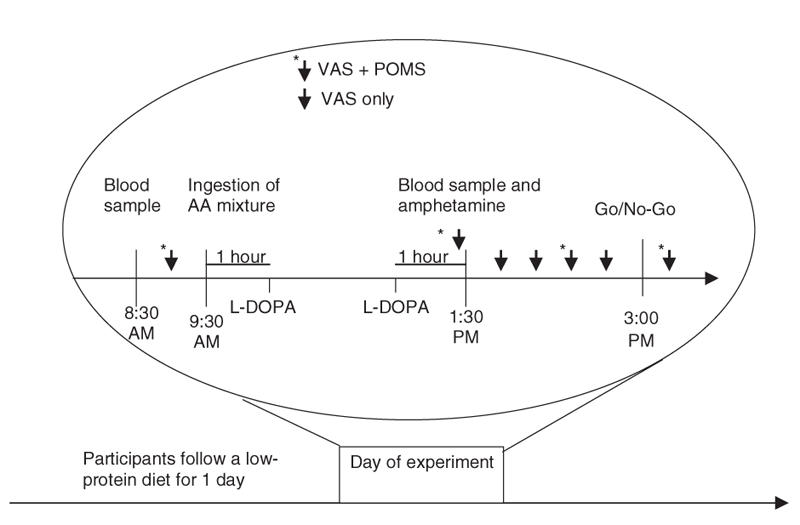
Fig. 1: Timeline of events on the 3 test sessions. VAS: visual analog scale; POMS: Profile of Mood States.
One and 3 hours after AA mixture administration, participants received placebo or L-DOPA/carbidopa (Sinemet, 100 mg/25 mg, orally), the immediate DA precursor with a peripheral decarboxylase inhibitor. Four hours after AA mixture administration, subjects were given tablets of d-amphetamine (Dexedrine, 0.3 mg/kg, orally). PET/[11C]raclopride studies indicate that this dose of d-amphetamine increases DA release in human limbic striatum52 and that the effect is significantly diminished by APTD.28
Dependent measures
Subjective effects of d-amphetamine were measured with the bipolar Profile of Mood States (POMS)53,54 and 10 visual analog scales (VASs).55 The POMS comprises 6 scales (ElateDepressed, Composed–Anxious, Agreeable–Hostile, Confident–Unsure, Energetic–Tired, and Clearheaded–Confused) and is highly sensitive to nonclinical changes in mood states. It was administered 4 times: at baseline before ingesting the AA mixture, immediately before d-amphetamine administration (4 hours after AA mixture ingestion) and 60 and 120 minutes postdrug. The VASs were labelled “Rush,” “High,” “Euphoria,” “Excited,” “Anxious,” “Energetic,” “Like drug,” “Mind racing,” “Alert” and “Would like drug again.” They were administered 7 times, at baseline before ingesting the AA mixture, immediately before amphetamine administration and every 20 minutes after amphetamine administration for 2 hours.
A computerized Go/No-Go task measured responses to rewards and punishments in 4 conditions.56 In each condition, participants are presented with a set of 2-digit numbers. The first time that subjects do the task, each condition has 8 numbers. The second and third times, 10 numbers. This modification is thought to compensate for the potential effect of repeated testing.57 By trial and error, subjects learn that one-half of the numbers signal that a button should be pressed, and one-half indicate that the button should not be pressed. In 2 of the 4 conditions (reward–reward and reward–punishment), pressing a button in response to the correct stimulus leads to a reward (win 10 cents); in the other 2 conditions (punishment–punishment and punishment–reward), pressing correctly avoids a punishment (lose 10 cents). Subjects are provided with 12 training trials (15 on test days 2 and 3) before starting the task, and performance during this phase does not count toward the final score. After this initial phase, each number is briefly shown on the screen, 10 times each. The task was administered 5 and a half hours after the AA mixture was given and takes approximately 30 minutes to complete. Thus, participants were taking the Go/No-Go task between 90 and 120 minutes after d-amphetamine administration, which corresponds to the period when plasma drug levels peak.58
Blood samples were drawn at morning baseline before AA mixture ingestion and 6 hours later. Phenylalanine and tyrosine were measured with precolumn derivatization with o-phthalaldehyde and gradient reverse phase HPLC with aminoadipic acid as an internal standard and fluorometric detection. Plasma concentrations of amphetamine were analyzed with electron-capture gas chromatography after extraction and derivatization of amphetamine with pentafluorobenzenesulfonyl chloride.59 In 3 participants, plasma samples could not be drawn at all time points.
Statistical methods
We analyzed the plasma, subjective state, and Go/No-Go data by separate repeated-measures, univariate analyses of variance (ANOVA). AA mixture and time were within-subject factors. For the plasma data, time had 2 levels (immediately before and 6 hours after AA administration). For the subjective state data, the time factor was defined as change in mood from pre-AA mixture to 4 hours postmixture and maximum change from preamphetamine to postamphetamine. For the Go/No-Go analyses, AA mixture and Go/No-Go condition were also within-subject factors. Significant results revealed by these procedures were further examined by post hoc Least Significant Difference tests. All tests were 2-tailed.
Results
Plasma amino acids
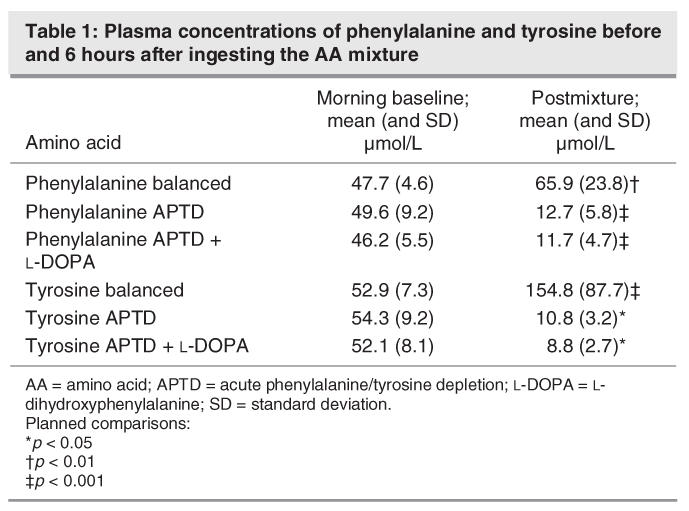
APTD lowered plasma concentrations of phenylalanine and tyrosine, as reflected by significant AA mixture × time interactions (tyrosine: F2,16 = 23.5, p ≤ 0.002; phenylalanine: F2,16 = 38.4, p ≤ 0.001). Compared with morning baseline, phenylalanine and tyrosine levels were reduced by 74.4% and 74.6% and 80.1% and 83.1% by the APTD and APTD + L-DOPA treatments, respectively (p < 0.05). The effect of the 2 APTD conditions did not differ (p ≥ 0.80). The nutritionally balanced (BAL) mixture increased plasma phenylalanine and tyrosine by 38.2% and 192.6% (p ≤ 0.005) (Table 1).
Table 1
Plasma amphetamine
Amphetamine was not present in any of the morning baseline samples and was present at a concentration of 26.0 (SD 5.7) ng/mL 120 minutes after drug administration. The AA mixtures did not alter amphetamine bioavailability (BAL: 26.9 [SD 5.8], APTD: 27.5 [SD 6.0], APTD + L-DOPA: 24.1 [SD] 6.3). In line with this, the AA mixture × time ANOVA yielded a significant main effect of time (F1,17 = 211.8, p < 0.001), but not AA mixture (F2,14 = 1.85, p ≥ 0.20) or a mixture × time interaction (F2,14 = 1.45, p ≥ 0.27).
Subjective effects
Repeated-measures analyses of the POMS data indicated that there were no main effects of AA mixture (p ≥ 0.35) or AA mixture × time interactions (p ≥ 0.46). In comparison, significant effects of time were seen for the POMS scales Elated–Depressed (F1,13 = 12.89, p ≤ 0.01), Energetic–Tired (F1,13 = 10.26, p ≤ 0.01), Confident–Unsure (F1,13 = 8.93, p ≤ 0.01), Agreeable–Hostile (F1,13 = 10.67, p ≤ 0.01) and Clearheaded–Confused (F1,13 = 6.01, p ≤ 0.03), whereas scores on the Composed–Anxious scale remained unchanged (F1,13 = 1.85, p ≥ 0.20) (Fig. 2).
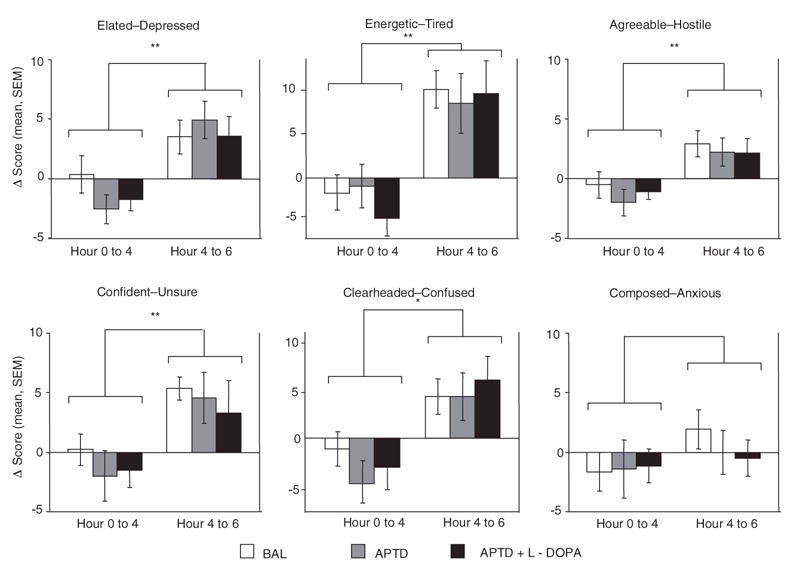
Fig. 2: Effect of acute phenylalanine/tyrosine depletion (APTD) and d-amphetamine on mood, as measured by the bipolar Profile of Mood States (POMS). The graphs depict mean change scores ± SEM for each of the POMS subscales from hour 0 to hour 4 (post-AA minus pre-AA) and from hour 4 to hour 6 (postamphetamine minus preamphetamine), as a function of type of amino acid mixture (APTD, BAL, APTD + L-DOPA). A higher score denotes more positive mood. Postamphetamine scores are defined as the maximum value after amphetamine administration. *p ≤ 0.02; **p ≤ 0.01.
Inspection of the data indicated that the effects of time reflected mood-elevating effects of d-amphetamine. Collapsed across test days, and compared with POMS scores obtained immediately before d-amphetamine administration, the stimulant drug significantly increased Elated–Depressed (F1,13 = 16.46, p ≤ 0.001), Agreeable–Hostile (F1,13 = 9.83, p ≤ 0.008), Energetic–Tired (F1,13 = 12.79, p ≤ 0.003), Confident–Unsure (F1,13 = 9.47, p ≤ 0.009), and Clearheaded–Confused scores (F1,13 = 6.87, p ≤ 0.02), whereas POMS scores on the Composed–Anxious scale remained unchanged (F1,13 = 0.14, p ≥ 0.72) (Fig. 2).
For each of the VAS items except for “Anxious,” repeated-measures analyses also indicated statistically significant main effects of time (p ≤ 0.02) (Table 1) but not effects of AA mixture or AA mixture × time interactions. VAS items that significantly increased after d-amphetamine, compared with preamphetamine, were “Rush” (F1,13 = 18.70, p ≤ 0.001), “High” (F1,13 = 16.81, p ≤ 0.001), “Euphoria” (F1,13 = 11.40, p ≤ 0.005), “Excited“ (F1,13 = 25.98, p ≤ 0.001), “Energetic” (F1,13 = 19.72, p ≤ 0.001), “Like drug” (F1,13 = 11.06, p ≤ 0.005), “Mind-racing” (F1,13 = 22.87, p ≤ 0.001), “Alert” (F1,13 = 16.05, p ≤ 0.001), and “Would like drug again” (F1,13 = 8.41, p ≤ 0.01) but not “Anxious” (F1,13 = 1.34, p ≥ 0.27).
Go/No-Go commission errors
An AA mixture × Go/No-Go condition ANOVA of Go/No-Go commission errors during the initial learning phase yielded a near significant effect of AA mixture (F2,24 = 3.19, p ≤ 0.059) but not an AA mixture × Go/No-Go condition interaction (F6,72 = 1.62, p ≥ 0.15). Data inspection suggested that there was a tendency for commission errors to be higher during the test session with L-DOPA (mean ± SEM, 13.3 ± 1.5), compared with the sessions with BAL (10.8 ± 1.3, p ≤ 0.045) and APTD (11.0 ± 1.3, p ≤ 0.035).
Analyses of performance after the learning phase yielded a significant AA mixture × Go/No-Go condition interaction (F6,72 = 2.32, p ≤ 0.05). Data inspection indicated that APTD's effects were restricted to the 2 conditions where correct responses lead to a reward (win 10 cents; reward–punishment and reward–reward); in the 2 other conditions (punishmentpunishment and punishment–reward), correct responding avoids a loss (lose 10 cents). Analyses of the reward versus no reward conditions confirmed this distinction; the ANOVA again yielded a significant AA mixture × Go/No-Go condition interaction (F2,24 = 3.60, p ≤ 0.04), and relative to the BAL test session, APTD disturbed responses to stimuli in the reward (p ≤ 0.005) but not the comparison condition (p ≥ 0.60) (Fig. 3). L-DOPA prevented this effect of APTD (p ≤ 0.001), improving performance and restoring it to a level not significantly different from that seen on the BAL test day (p ≥ 0.55). Only the number of incorrect responses (commission errors) was affected; APTD did not alter omission errors (p > 0.05).
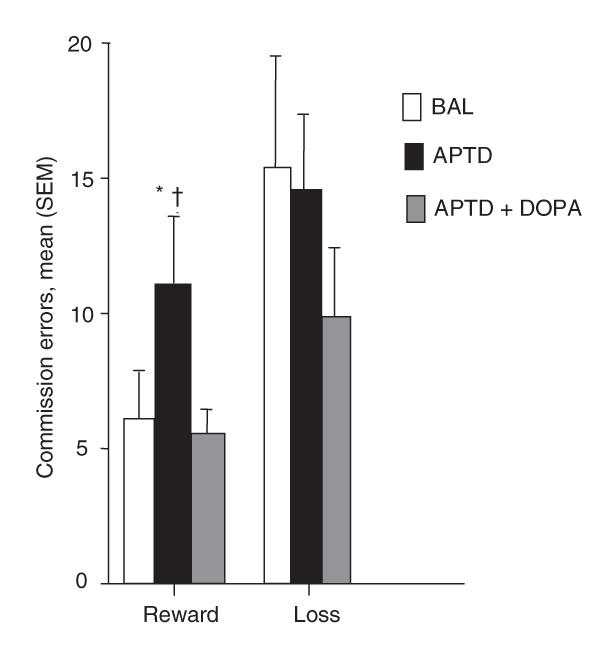
Fig. 3: Go/No-Go task commission errors (CEs) after the learning phase. Compared with the test session with the nutritionally balanced mixture (BAL), acute phenylalanine/tyrosine depletion (APTD) increased CE in the reward condition (reward–reward and reward–punishment), and performance was restored by L-DOPA. These effects of APTD and L-DOPA were not seen in the control condition. *p ≤ 0.005, compared with the BAL test day; † p ≤ 0.001, compared with the APTD + L-DOPA test day.
Discussion
The primary finding in the present study was that APTD did not alter mood-elevating effects of d-amphetamine, a result similar to previous APTD studies with cocaine,43 alcohol37,38 and nicotine.47 In comparison, APTD significantly decreased the ability to preferentially respond to stimuli that predicted reward; this deficit was prevented by the immediate DA precursor, L-DOPA.
Although functional neuroimaging studies in humans have suggested that stimulant drug-induced striatal DA release correlates with both euphoria60–65 and drug craving,52,65 mood elevating effects of stimulant drugs are not consistently decreased by treatments that diminish DA transmission.43,66–71 In comparison, there is a small but more consistent literature suggesting that decreased DA transmission diminishes cocaine-and cocaine cue-induced craving43,72 and the sustained motivation to obtain alcohol.38 However, the finding that APTD does not alter the mood-elevating effects of d-amphetamine does not rule out a relation between DA and all aspects of emotion.73–75 Since mood and motivational processes affect each other,76 mesolimbic DA transmission might influence mood by moderating the anticipatory and appetitive components of positive affect rather than providing a sufficient substrate of mood elevation, per se.77,78 Other contributors to drug-induced mood elevation might include NE,79 serotonin80 and endogenous opioids.81
The ability of APTD to disrupt preferential responding to reward-paired stimuli resembles the finding that tyrosine depletion alters the ability to adjust betting behaviour in a gambling task.36,39,40 Similarly, the ability of L-DOPA to restore the APTD-induced performance deficit supports a recent report that L-DOPA administration improves responding to stimuli predictive of reward but not loss.82 Together these findings add to the evidence that midbrain DA transmission is not closely linked to drug-induced mood-elevation but, across a range of tasks, enhances the ability to identify reward-related cues and respond to them preferentially.
Noting the above observations, the results should be interpreted in light of the following considerations. First, the administered dose of d-amphetamine (0.3 mg/kg, orally) elicited modest subjective effects, and the inability of APTD to diminish these effects might reflect the low dose. However, tyrosine depletion also failed to decrease the mood-elevating effects of a 5-fold range of cocaine doses,43 and it left unaltered the mood-elevating effects of 0.15 mg/kg of intravenously injected methamphetamine, although it moderately diminished 2 effects related to accelerated thoughts: “Mind-Racing” and the subjective sense of “buzz.”41
Second, participants received d-amphetamine on all 3 test sessions, precluding an assessment of whether the stimulant drug itself affected responses to reward cues. However, a study conducted elsewhere indicates that d-amphetamine (10 and 20 mg, given orally), compared with placebo, improves responding to reward stimuli in the Go/No-Go task.83
Third, we have interpreted the change in Go/No-Go performance to reflect a change in the ability to identify and preferentially respond to reward-related cues. Alternative explanations include changes in motor function, attention or memory. However, a motor hypothesis would predict the opposite of what we observed. Decreased DA transmission would be expected to decrease, not increase, behavioural activation. Moreover, motor, attentional and memory disturbances would be expected to alter both omission and commission errors and to do so in all 4 Go/No-Go sub-conditions. This was not seen.
Fourth, although L-DOPA was able to prevent the effect of APTD on responses to reward-predictive stimuli, in 2 previous studies it was unable to prevent APTD's effects on cocaine craving43 or alcohol self-administration progressive ratio breakpoints38; in this study, it tended to worsen performance during the learning phase. This differential efficacy was unexpected. However, the ability of L-DOPA to increase DA synthesis and release84–86 but transiently reduce DA cell firing under conditions of diminished DA function87,88 raises the possibility that phasic DA cell firing versus tonic DA release mediate different aspects of reward-related behaviour.89,90 DA cell burst firing has been found to tightly co-vary with the expectation of reward9,91 and to facilitate set shifting92 and the acquisition of reward-related behaviours.93 In comparison, slow tonic increases in DA release appear to have neuromodulatory effects,94 sustaining interest in motivationally relevant events,4,90,95–98 facilitating the learned association between rewards and reward-paired cues14–17,92 and eliciting approach.2,13 Acute L-DOPA administration, therefore, might restore behaviours that require elevated tonic DA levels but not phasic cell bursts. Craving for drug reward, in comparison, might emerge from increases in both phasic DA cell firing and tonic DA release.38,43,90,91,95,99
Fifth, APTD might affect NE synthesis19,24; however, accumulating evidence suggests that, to the extent that effects on NE synthesis occur, they are insufficient to alter NE release.19,22,25,26 Finally, all the participants were men. Sex differences in response to stimulant drugs have been reported,100,101 and it would be of interest to repeat this study in women.
In conclusion, the present study suggests that stimulant drug-induced changes in mood are not closely related to changes in DA transmission. In comparison, APTD perturbed the ability to preferentially respond to reward-paired stimuli, an effect that was prevented by the immediate DA precursor, L-DOPA. These and findings from related studies support the hypothesis that drug-induced increases in DA transmission enhance the perceived value of reward stimuli, increasing the tendency to focus on and approach drug-paired cues and facilitating the production of motivational and craving states.
Acknowledgments
This work was supported by an operating grant from the Canadian Institutes of Health Research to M.L. (MOP–36429) and C.B; M.L. and C.B. are both recipients of salary awards from Fonds de la recherche en santé du Québec (FRSQ) and funded research chairs from McGill University. G.B.B. is a Canada Research Chair. M.a.h.R. received a scholarship from the Netherlands Brain Foundation. L.B. received a fellowship from the Netherlands Organisation for Scientific Research (NWO). We thank Franceen Lenoff and Gail Rauw for their excellent technical assistance.
Footnotes
Contributors: Drs. Leyton and Young designed the study. Dr. Leyton, Ms. aan het Rot and Drs. Baker and Benkelfat acquired the data, which Drs. Leyton, Booij, Baker and Benkelfat analyzed. All authors gave approval for the final version of the article to be published. Drs. Leyton and Booij wrote the article, and Ms. aan het Rot and Drs. Booij, Baker, Young and Benkelfat critically reviewed it.
Competing interests: None declared.
Correspondence to: Dr. Marco Leyton, Department of Psychiatry, McGill University, 1033 Pine Ave. W., Montreal QC H3A 1A1; fax 514 398-4866; marco.leyton@mcgill.ca
References
- 1.Wise RA. Neuroleptics and operant behavior: the anhedonia hypothesis. Behav Brain Sci 1982;5:39-53.
- 2.Wise RA, Bozarth MA. A psychomotor stimulant theory of addiction. Psychol Rev 1987;94:469-92. [PubMed]
- 3.Wise RA, Rompré PP. Brain dopamine and reward. Annu Rev Psychol 1989;40:191-225. [DOI] [PubMed]
- 4.Stewart J, de Wit H, Eikelboom R. Role of unconditioned and conditioned drug effects in the self-administration of opiates and stimulants. Psychol Rev 1984;91:251-68. [PubMed]
- 5.Stewart J. Neurobiology of conditioning to drugs of abuse. In: Kalivas PW, Samson HH, editors. The Neurobiology of Drug and Alcohol Addiction. New York: New York Academy of Sciences; 1992. p. 335-46. [DOI] [PubMed]
- 6.Blackburn JR, Pfaus JG, Phillips AG. Dopamine functions in appetitive and defensive behaviours. Prog Neurobiol 1992;39:247-79. [DOI] [PubMed]
- 7.Berridge KC, Robinson TE. What is the role of dopamine in reward: hedonic impact, reward learning, or incentive salience? Brain Res Brain Res Rev 1998;28:309-69. [DOI] [PubMed]
- 8.Ikemoto S, Panksepp J. The role of nucleus accumbens dopamine in motivated behavior: a unifying interpretation with special reference to reward-seeking. Brain Res Brain Res Rev 1999;31:6-41. [DOI] [PubMed]
- 9.Schultz W. Predictive reward signal of dopamine neurons. J Neurophysiol 1998;80:1-27. [DOI] [PubMed]
- 10.Genn RF, Ahn S, Phillips AG. Attenuated dopamine efflux in the rat nucleus accumbens during successive negative contrast. Behav Neurosci 2004;118:869-73. [DOI] [PubMed]
- 11.Redgrave P, Prescott TJ, Gurney K. Is the short-latency dopamine response too short to signal reward error? Trends Neurosci 1999;22:146-51. [DOI] [PubMed]
- 12.Joseph MH, Datla K, Young AMJ. The interpretation of the measurement of nucleus accumbens dopamine by in vivo dialysis: the kick, the craving or the cognition? Neurosci Biobehav Rev 2003;27:527-41. [DOI] [PubMed]
- 13.Salamone JD, Correa M, Mingote SM, et al. Beyond the reward hypothesis: alternative functions of nucleus accumbens dopamine. Curr Opin Pharmacol 2005;5:34-41. [DOI] [PubMed]
- 14.Beninger RJ. The role of dopamine in locomotor activity and learning. Brain Res 1983;287:173-96. [DOI] [PubMed]
- 15.Schultz W, Dayan P, Montague PR. A neural substrate of prediction and reward. Science 1997;275:1593-9. [DOI] [PubMed]
- 16.Di Chiara G, Tanda G, Bassareo V, et al. Drug addiction as a disorder of associative learning: role of nucleus accumbens shell/extended amygdala dopamine. Ann N Y Acad Sci 1999;877:461-85. [DOI] [PubMed]
- 17.Everitt BJ, Parkinson JA, Olmstead MC, et al. Associative processes in addiction and reward. The role of amygdala-ventral striatal subsystems. Ann N Y Acad Sci 1999;877:412-38. [DOI] [PubMed]
- 18.Wise RA. Dopamine, learning and motivation. Nat Rev Neurosci 2004; 5:483-94. [DOI] [PubMed]
- 19.McTavish SF, Cowen PJ, Sharp T. Effect of a tyrosine-free amino acid mixture on regional brain catecholamine synthesis and release. Psychopharmacology (Berl) 1999;141:182-8. [DOI] [PubMed]
- 20.Le Masurier M, Houston G, Cowen P, et al. Tyrosine-free amino acid mixture attenuates amphetamine-induced displacement of [11C]raclopride in striatum in vivo: a rat PET study. Synapse 2004;51:151-7. [DOI] [PubMed]
- 21.Jaskiw GE, Bongiovanni R. Brain tyrosine depletion attenuates haloperidol-induced striatal dopamine release in vivo and augments haloperidol-induced catalepsy in the rat. Psychopharmacology (Berl) 2004;172:100-7. [DOI] [PubMed]
- 22.Le Masurier M, Cowen PJ, Sharp T. Fos immunocytochemical studies on the neuroanatomical sites of action of acute tyrosine depletion in the rat brain. Psychopharmacology (Berl) 2004;171:435-40. [DOI] [PubMed]
- 23.Biggio G, Porceddu ML, Gessa GL. Decrease of homovanillic, dihydroxyphenylacetic acid and cyclic-adenosine-3′,5′-monophosphate content in the rat caudate nucleus induced by the acute administration of an amino acid mixture lacking tyrosine and phenylalanine. J Neurochem 1976;26:1253-5. [DOI] [PubMed]
- 24.Palmour RM, Ervin FR, Baker GB, et al. Effects of acute tryptophan depletion and acute tyrosine/phenylalanine depletion on CSF amine metabolite levels and voluntary alcohol consumption in vervet monkeys. Psychopharmacology (Berl) 1998;136:1-7. [DOI] [PubMed]
- 25.Sheehan BD, Tharyan P, McTavish SFB, et al. The use of dietary manipulation to deplete plasma tyrosine and phenylalanine in healthy subjects. J Psychopharmacol 1996;10:231-4. [DOI] [PubMed]
- 26.McTavish SF, Callado L, Cowen PJ, et al. Comparison of the effects of alpha-methyl-p-tyrosine and a tyrosine-free amino acid load on extracellular noradrenaline in the rat hippocampus in vivo. J Psychopharmacol 1999;13:379-84. [DOI] [PubMed]
- 27.Montgomery AJ, McTavish SFB, Cowen PJ, et al. Reduction in brain dopamine concentration with dietary tyrosine plus phenylalanine depletion: an [11C]raclopride PET study. Am J Psychiatry 2003;160:1887-9. [DOI] [PubMed]
- 28.Leyton M, Dagher A, Boileau I, et al. Decreasing amphetamine induced dopamine release by acute phenylalanine / tyrosine depletion: A PET [11C]raclopride study in healthy men. Neuropsychopharmacology 2004;29:427-32. [DOI] [PubMed]
- 29.Harmer CJ, McTavish SFB, Clark L, et al. Tyrosine depletion attenuates dopamine function in healthy volunteers. Psychopharmacology (Berl) 2001;154:105-11. [DOI] [PubMed]
- 30.Gijsman HJ, Scarna A, Harmer CJ, et al. A dose-finding study on the effects of branch chain amino acids on surrogate markers of brain dopamine function. Psychopharmacology (Berl) 2002;160:192-7. [DOI] [PubMed]
- 31.McTavish SFB, Mannie ZN, Harmer CJ, et al. Lack of effect of tyrosine depletion in recovered depressed women. Neuropsychopharmacology 2005;30:786-91. [DOI] [PubMed]
- 32.Lythe KE, Anderson IM, Deakin JF, et al. Lack of behavioural effects after tyrosine depletion in healthy volunteers. J Psychopharmacol 2005;19:5-11. [DOI] [PubMed]
- 33.Harrison BJ, Olver JS, Norman TR, et al. Selective effects of acute serotonin and catecholamine depletion on memory in healthy women. J Psychopharmacol 2004;18:32-40. [DOI] [PubMed]
- 34.Mehta MA, Gumaste D, Montgomery AJ, et al. The effects of acute tyrosine and phenylalanine depletion on spatial working memory and planning in healthy volunteers are predicted by changes in striatal dopamine levels. Psychopharmacology (Berl) 2005;180:654-63. [DOI] [PubMed]
- 35.Ellis KA, Mehta MA, Wesnes KA, et al. Combined D1/D2 receptor stimulation under conditions of dopamine depletion impairs spatial working memory performance in humans. Psychopharmacology (Berl) 2005;181:771-80. [DOI] [PubMed]
- 36.Scarna A, McTavish SF, Cowen PJ, et al. The effects of a branched chain amino acid mixture supplemented with tryptophan on biochemical indices of neurotransmitter function and decision-making. Psychopharmacology (Berl) 2005;179:761-8. [DOI] [PubMed]
- 37.Leyton M, Young SN, Blier P, et al. Acute tyrosine depletion and alcohol ingestion in healthy women. Alcohol Clin Exp Res 2000a;24:459-64. [PubMed]
- 38.Barrett SP, Pihl RO, Benkelfat C, et al. The effect of dopamine precursor depletion on alcohol self-administration in men: individual differences. Neuropsychopharmacology 2005;30(Suppl 1):150-1.
- 39.McLean A, Rubinsztein JS, Robbins TW, et al. The effects of tyrosine depletion in normal healthy volunteers: implications for unipolar depression. Psychopharmacology (Berl) 2004;171:286-97. [DOI] [PubMed]
- 40.Roiser JP, McLean A, Ogilvie AD, et al. The subjective and cognitive effects of acute phenylalanine and tyrosine depletion in patients recovered from depression. Neuropsychopharmacology 2005;30:775-85. [DOI] [PMC free article] [PubMed]
- 41.McTavish SFB, McPherson MH, Harmer CJ, et al. Antidopaminergic effects of dietary tyrosine depletion in healthy subjects and patients with manic illness. Br J Psychiatry 2001;179:356-60. [DOI] [PubMed]
- 42.Scarna A, Gijsman HJ, McTavish SF, et al. Effects of a branched-chain amino acid drink in mania. Br J Psychiatry 2003;182:210-3. [DOI] [PubMed]
- 43.Leyton M, Casey KF, Delaney JS, et al. Cocaine craving, euphoria, and self-administration: a preliminary study of the effect of catecholamine precursor depletion. Behav Neurosci 2005;119:1619-27. [DOI] [PubMed]
- 44.McTavish SFB, McPherson MH, Sharp T, et al. Attenuation of some subjective effects of amphetamine following tyrosine depletion. J Psychopharmacol 1999;13:144-7. [DOI] [PubMed]
- 45.Leyton M, Young SN, Pihl RO, et al. Effects on mood of acute phenylalanine/tyrosine depletion in healthy women. Neuropsychopharmacology 2000;22:52-63. [DOI] [PubMed]
- 46.Coupland N, Zedkova L, Sanghera G, et al. Pilot study of acute phenylalanine / tyrosine depletion (APTD) and responses to pentagastrin in healthy males. J Psychiatry Neurosci 2001;26:247-51. [PMC free article] [PubMed]
- 47.Casey KF, Benkelfat C, Young SN, et al. Lack of effect of dopamine precursor depletion in nicotine-dependent smokers. Eur Neuropsychopharmacol 2006;16:512-20. [DOI] [PubMed]
- 48.American Psychiatric Association. Diagnostic and statistical manual of mental disorders. 4th ed. Washington: The Association; 1994.
- 49.First MB, Spitzer RI, Gibbon M. Structured Clinical Interview for DSM-IV, non-patient Version. New York: New York State Psychiatric Institute; 1995.
- 50.Nurnberger JI Jr, Blehar MC, Kaufmann CA, et al. Diagnostic interview for genetic studies. Rationale, unique features, and training. NIMH Genetics Initiative. Arch Gen Psychiatry 1994;51:849-59. [DOI] [PubMed]
- 51.Young SN, Smith SE, Pihl RO, et al. Tryptophan depletion causes a rapid lowering of mood in normal males. Psychopharmacology (Berl) 1985;87:173-7. [DOI] [PubMed]
- 52.Leyton M, Boileau I, Benkelfat C, et al. Amphetamine-induced increases in extracellular dopamine, drug wanting and novelty seeking: A PET/[11C]raclopride study in healthy men. Neuropsychopharmacology 2002;27:1027-35. [DOI] [PubMed]
- 53.Lorr M, McNair DM, Fisher S. Evidence for bipolar mood states. J Pers Assess 1982;46:432-6. [DOI] [PubMed]
- 54.McNair DM, Lorr M, Droppleman LF. Manual for the Profile of Mood States. San Diego: Educational and Industrial Testing Service; 1988.
- 55.Bond A, Lader M. The use of analog scales in rating subjective feelings. Br J Med Psychol 1974;47:211-8.
- 56.Iaboni F, Douglas VI, Baker AG. Effects of reward and response costs on inhibition in ADHD children. J Abnorm Psychol 1995;104:232-40. [DOI] [PubMed]
- 57.LeMarquand DG, Benkelfat C, Pihl RO, et al. Behavioral disinhibition induced by tryptophan depletion in nonalcoholic young men with multigenerational family histories of paternal alcoholism. Am J Psychiatry 1999;156:1771-9. [DOI] [PubMed]
- 58.Angrist B, Corwin J, Bartlik B, et al. Early pharmacokinetics and clinical effects of oral d-amphetamine in normal subjects. Biol Psychiatry 1987;22:1357-68. [DOI] [PubMed]
- 59.Asghar SJ, Baker GB, Rauw GA, et al. A rapid method of determining amphetamine in plasma samples using pentafluorobenzenesulfonyl chloride and electron-capture gas chromatography. J Pharmacol Toxicol Methods 2001;46:111-5. [DOI] [PubMed]
- 60.Laruelle M, Abi-Dargham A, van Dyck CH, et al. SPECT imaging of striatal dopamine release after amphetamine challenge. J Nucl Med 1995;36:1182-90. [PubMed]
- 61.Volkow ND, Wang GJ, Fowler JS, et al. Reinforcing effects of psychostimulants in humans are associated with increases in brain dopamine occupancy of D(2) receptors. J Pharmacol Exp Ther 1999;291:409-15. [PubMed]
- 62.Drevets WC, Gautier C, Price JC, et al. Amphetamine-induced dopamine release in human ventral striatum correlates with euphoria. Biol Psychiatry 2001;49:81-96. [DOI] [PubMed]
- 63.Martinez D, Slifstein M, Broft A, et al. Imaging human mesolimbic dopamine transmission with positron emission tomography. Part II: amphetamine-induced dopamine release in the functional subdivisions of the striatum. J Cereb Blood Flow Metab 2003;23:285-300. [DOI] [PubMed]
- 64.Abi-Dargham A, Kegeles LS, Martinez D, et al. Dopamine mediation of positive reinforcing effects of amphetamine in stimulant naive healthy volunteers: results from a large cohort. Eur Neuropsychopharmacol 2003;13:459-68. [DOI] [PubMed]
- 65.Oswald LM, Wong DF, McCaul M, et al. Relationships among ventral striatal dopamine release, cortisol secretion, and subjective responses to amphetamine. Neuropsychopharmacology 2005;30:821-32. [DOI] [PubMed]
- 66.Gawin FH. Neuroleptic reductions of cocaine-induced paranoia but not euphoria? Psychopharmacology (Berl) 1986;90:142-3. [DOI] [PubMed]
- 67.Sherer MA. Intravenous cocaine: psychiatric effects, biological mechanisms. Biol Psychiatry 1988;24:865-85. [DOI] [PubMed]
- 68.Brauer LH, de Wit H. Subjective responses to d-amphetamine alone and after pimozide pretreatment in normal, healthy volunteers. Biol Psychiatry 1996;39:26-32. [DOI] [PubMed]
- 69.Brauer LH, De Wit H. High dose pimozide does not block amphetamine-induced euphoria in normal volunteers. Pharmacol Biochem Behav 1997;56:265-72. [DOI] [PubMed]
- 70.Evans SM, Walsh SL, Levin FR, et al. Effect of flupenthixol on subjective and cardiovascular responses to intravenous cocaine in humans. Drug Alcohol Depend 2001;64:271-83. [DOI] [PubMed]
- 71.Romach MK, Glue P, Kampman K, et al. Attenuation of the euphoric effects of cocaine by the dopamine D1/D5 antagonist ecopipam (SCH 39166). Arch Gen Psychiatry 1999;56:1101-6. [DOI] [PubMed]
- 72.Berger SP, Hall S, Mickalian JD, et al. Haloperidol antagonism of cue-elicited cocaine craving. Lancet 1996;347:504-8. [DOI] [PubMed]
- 73.Murphy DL, Brodie HK, Goodwin FK, et al. Regular induction of hypomania by L-dopa in “bipolar” manic-depressive patients. Nature 1971;229:135-6. [DOI] [PubMed]
- 74.Fibiger HC. Neurobiology of depression: focus on dopamine. In: Fratta W, Pani L, Serra G, editors. Depression and Mania: From Neurobiology to Treatment. New York: Raven Press; 1995. p. 1-17.
- 75.Willner P. Dopaminergic mechanisms in depression and mania. In: Bloom FE, Kupfer DJ, editors. Psychopharmacology: The Fourth Generation of Progress. New York: Raven Press; 1995. p. 921-31.
- 76.Lazarus RS. Emotion and Adaptation. New York: Oxford University Press; 1991.
- 77.Rolls ET. Emotion Explained. New York: Oxford University Press; 2005.
- 78.Burgdorf J, Panksepp J. The neurobiology of positive emotions. Neurosci Biobehav Rev 2006;30:173-87. [DOI] [PubMed]
- 79.Rothman RB, Baumann MH, Dersch CM, et al. Amphetamine-type central nervous system stimulants release norepinephrine more potently than they release dopamine and serotonin. Synapse 2001;39:32-41. [DOI] [PubMed]
- 80.Aronson SC, Black JE, McDougle CJ, et al. Serotonergic mechanisms of cocaine effects in humans. Psychopharmacology (Berl) 1995;119:179-85. [DOI] [PubMed]
- 81.Pecina S, Berridge KC. Opioid site in nucleus accumbens shell mediates eating and hedonic ‚liking' for food: map based on microinjection Fos plumes. Brain Res 2000;863:71-86. [DOI] [PubMed]
- 82.Frank MJ, Seeberger LC, O'Reilly RC. By carrot or by stick: cognitive reinforcement learning in Parkinsonism. Science 2004;306:1940-3. [DOI] [PubMed]
- 83.de Wit H, Enggasser JL, Richards JB. Acute administration of d-amphetamine decreases impulsivity in healthy volunteers. Neuropsychopharmacology 2002;27:813-25. [DOI] [PubMed]
- 84.Butcher LL, Engel J. Behavioral and biochemical effects of L-DOPA after peripheral decarboxylase inhibition. Brain Res 1969;15:233-42. [DOI] [PubMed]
- 85.Freed CR, Murphy RC. Mass spectrometric measurement of catecholamine turnover in rat hypothalamus after long-term L-DOPA infusion. J Pharmacol Exp Ther 1978;205:702-9. [PubMed]
- 86.Eldrup E, Mogensen P, Jacobsen J, et al. CSF and plasma concentrations of free norepinephrine, dopamine, 3,4-dihydroxyphenylacetic acid (DOPAC), 3,4-dihydroxyphenylalanine (DOPA), and epinephrine in Parkinson's disease. Acta Neurol Scand 1995;92:116-21. [DOI] [PubMed]
- 87.Harden DG, Grace AA. Activation of dopamine cell firing by repeated L-DOPA administration to dopamine-depleted rats: its potential role in mediating the therapeutic response to L-DOPA treatment. J Neurosci 1995;15:6157-66. [DOI] [PMC free article] [PubMed]
- 88.Robinson S, Smith DM, Mizumori SJY, et al. Firing properties of dopamine neurons in freely moving dopamine-deficient mice: effects of dopamine receptor activation and anesthesia. Proc Natl Acad Sci U S A 2004;101:13329-34. [DOI] [PMC free article] [PubMed]
- 89.Grace AA. The tonic/phasic model of dopamine system regulation and its implications for understanding alcohol and psychostimulant craving. Addiction 2000;95(Suppl 95):S119-S28. [DOI] [PubMed]
- 90.Phillips AG, Ahn S, Howland JG. Amygdalar control of the mesocorticolimbic dopamine system: parallel pathways to motivated behavior. Neurosci Biobehav Rev 2003;27:543-54. [DOI] [PubMed]
- 91.Fiorillo CD, Tobler PN, Schultz W. Discrete coding of reward probability and uncertainty by dopamine neurons. Science 2003;299:1898-902. [DOI] [PubMed]
- 92.Goto Y, Grace AA. Dopaminergic modulation of limbic and cortical drive of nucleus accumbens in goal-directed behavior. Nat Neurosci 2005;8:805-12. [DOI] [PubMed]
- 93.Garris PA, Kilpatrick M, Bunin MA, et al. Dissociation of dopamine release from intracranial self-stimulation. Nature 1999;398:67-9. [DOI] [PubMed]
- 94.Greengard P. The neurobiology of slow synaptic transmission. Science 2001;294:1024-30. [DOI] [PubMed]
- 95.O'Donnell P. Dopamine gating of forebrain neural ensembles. Eur J Neurosci 2003;17:429-35. [DOI] [PubMed]
- 96.Volkow ND, Wang GJ, Fowler JS, et al. Evidence that methylphenidate enhances the saliency of a mathematical task by increasing dopamine in the human brain. Am J Psychiatry 2004;161:1173-80. [DOI] [PubMed]
- 97.Hernandez G, Hamdani S, Rajabi H, et al. Prolonged rewarding stimulation of the rat medial forebrain bundle: neurochemical and behavioral consequences. Behav Neurosci 2006;120:888-904. [DOI] [PubMed]
- 98.Volkow ND, Swanson JM. Variables that affect the clinical use and abuse of methylphenidate in the treatment of ADHD. Am J Psychiatry 2003;160:1909-18. [DOI] [PubMed]
- 99.Phillips PE, Stuber GD, Heien ML, et al. Subsecond dopamine release promotes cocaine seeking. Nature 2003;422:614-8. [DOI] [PubMed]
- 100.Becker JB, Molenda H, Hummer DL. Gender differences in the behavioral responses to cocaine and amphetamine. Implications for mechanisms mediating gender differences in drug abuse. Ann N Y Acad Sci 2001;937:172-87. [DOI] [PubMed]
- 101.Sofuoglu M, Dudesh-Poulsen S, Nelson D, et al. Sex and menstrual cycle differences in the subjective effects from smoked cocaine in humans. Exp Clin Psychopharmacol 1999;7:274-83. [DOI] [PubMed]