

Abstract
MDMA (ecstasy) is a drug commonly used in adolescence, and many users of MDMA also use other illicit drugs. It is not known whether MDMA during adolescence alters subsequent responses to cocaine differently than in adults. This study examined the effects of MDMA in adolescent and adult rats on cocaine conditioned reward. At the start of these experiments, adolescent rats were at postnatal day (PND) 33 and adult rats at PND 60. Each rat was treated for seven days with MDMA (2 or 5 mg/kg/day or vehicle) and locomotor activity was measured. Five days later cocaine conditioned place preference (CPP) was begun. Rats were trained for three days, in the morning with saline and in the afternoon with 10 mg/kg cocaine in 30 min sessions, and tested on the fourth day. MDMA stimulated activity in both age groups, but with a greater effect in the adult rats. Sensitization to the locomotor-stimulant effects of the lower dose of MDMA occurred in adult rats, and in both groups to the higher dose. Cocaine did not produce a CPP in vehicle-treated adolescent rats, but a significant CPP was observed subsequent to treatment with MDMA. In contrast, cocaine-induced CPP was diminished after MDMA in adult rats. These effects were still evident two weeks later upon retest. Thus, under the present conditions, MDMA increased cocaine conditioned reward in adolescent and decreased it in adult rats. These findings suggest that exposure to MDMA during this critical developmental period may carry a greater risk than during adulthood and that male adolescents may be particularly vulnerable to the risk of stimulant abuse after use of MDMA.
1. Introduction
MDMA (ecstasy) is a drug that is typically used by teenagers and young adults in bars and clubs. The National Survey on Drug Use and Health has estimated that about 2.1 million people over 12 years of age reported having used ecstasy in the past year, with the majority being young adults [25]. In addition, MDMA users were more likely to use other drugs compared to non-MDMA users, with 90% of MDMA users and only 13.8% of non-MDMA users reporting having used other illicit drugs. Specifically, cocaine was reported as being used by 43.8% of MDMA users and 2.0% of non-MDMA users [25].
In adult rats, MDMA increases locomotor activity and at high doses produces behaviors indicative of a serotonin syndrome (e.g. forepaw treading, low body posture) [15 for review]. Sensitization occurs to the locomotor-activating effects of MDMA after repeated administration [28]. The increased locomotor activity is seen with low doses (3 or 4 mg/kg) and an even greater effect is seen with higher doses (8 or 12 mg/kg) [16]. There are conflicting results from studies of the reinforcing effects of MDMA with some studies showing a conditioned place preference (CPP) after s.c. [6, 20] or icv [7] administration of MDMA. Other studies, however, have shown that there is no significant CPP to MDMA in group housed rats, but rather that it occurs only in isolated rats [22]. Repeated treatment with MDMA in adult rats has been reported to facilitate the subsequent acquisition of cocaine self-administration (Fletcher et al., 2001), suggesting that there are crossover effects between MDMA and cocaine, while other studies show no crossover effects between MDMA and amphetamine or methylphenidate [23].
Previous experiments have shown that adult and adolescent rats have different neurochemical adaptations in response to MDMA, where the adolescent rats seem to be less vulnerable to the toxic effects. Experiments in adult rats have shown that either a repeated low dose (4 mg/kg) for 4 days [26], or one high dose (20 mg/kg) [21] caused significant decreases in serotonin concentrations in selected brain regions. In adolescent rats on the other hand, experiments have revealed that repeated administration of MDMA (7.5 mg/kg) twice daily for 3 days starting on PND 39 [14], or one dose of MDMA (20 mg/kg) on PND 14 or 21 [4] did not significantly alter the serotonin concentration.
Since MDMA is a drug predominately used by young adults and teenagers, it is important to investigate what effect this drug might have during this stage of development. Rats used in this study were either adolescents aged postnatal day (PND) 33 or adults aged PND 60 at the start of the experiments. In the rat, the periadolescent period, which is the period of early adolescence, takes place around PND 28–42 [29].
2. EXPERIMENTAL PROCEDURES
2.1 Subjects
The animals used in this study were maintained and the studies were conducted in accordance with the guidelines of the Guide for Care and Use of Laboratory Animals, National Research Council, Department of Health, Education and Welfare, NIH Publication 85-23, revised 1996. Sprague-Dawley male adolescent and adult rats (Charles River, Wilmington, MA) were used in all studies. Rats were housed two per cage in a temperature and humidity-controlled environment under a 12 h light/dark cycle. Food and water were available ad libitum.
2.2 Chemicals
Drugs were obtained from the following sources: (±)MDMA HCl and cocaine HCl (NIDA, Rockville, MD).
2.3 Locomotor activity
Animals were maintained on a 12 h light/dark schedule with lights on at 7 a.m. and off at 7 p.m. All behavioral testing was done during the light schedule between 9 a.m. and 4 p.m. with each group tested at the same hour each day and the groups randomized over the course of the day.
Male rats were injected ip with 2.0 or 5.0 mg/kg MDMA or saline once daily for 7 days. On the first two days and the last two days of the treatment period, each rat was placed in a locomotor activity chamber for a 15 min habituation period followed by an injection of MDMA or saline and activity was measured for one hour. Rats were placed in clear acrylic chambers (40.64 x 40.64 cm) inside Digiscan activity monitors (Accuscan, Columbus, OH) that were equipped with infrared light sensitive detectors mounted 2.5 cm apart along two perpendicular walls. Mounted along the opposing walls were infrared light beams that were directed at the detectors. One count of horizontal activity was registered each time the subject interrupted a beam. Activity was monitored for a total of 60 min during which beam breaks were measured over 12 consecutive 5 min time periods.
Each group consisted of 8 rats. The adolescent rats were at postnatal day (PND) 33 and weighed 100 ± 2 gms at the start of the experiment. The adult rats were at PND 60 and weighed 250 ± 2 gms. The time course of the entire experiment can be seen in Table. 1.
2.4 Cocaine Conditioned Place Preference (CPP)
Five days after the last injection of MDMA or saline (day 12 of the experiment) cocaine CPP was begun. At this point, the adolescent rats were at PND 44 and the adult rats were at PND 71. During the conditioning phase, rats were trained for three days – in the morning with saline and in the afternoon with 10 mg/kg cocaine ip. Each training session lasted 30 min. The apparatus consisted of an acrylic box the same size as the locomotor activity chambers but with a removable center barrier. On one side, the walls were white, the lid was black and white striped, and the floor was smooth. On the other side, the walls were black and white striped, the lid was white, and the bottom was rough. The opposite lids were used so that there would be approximately the same amount of light on both sides of the testing chamber. For each rat, saline was paired with one side and cocaine was paired with the other. The drug-paired side was randomized across groups with equal numbers of rats in each group having cocaine paired with the white side and with the striped side. In preliminary experiments, this apparatus did not produce a bias towards one side or the other and the rats spent equal amounts of time in both sides. On the fourth day (day 15), CPP testing was done in the middle of the day. Two weeks later, on day 29, the rats that were treated with 5.0 mg/kg MDMA or saline were re-tested. During these tests, the rats were placed in the chamber with the center barrier removed and were able to move freely to both sides. The amount of time spent on each side of the chamber was recorded for 30 min. The time course of the entire experiment can be seen in Table. 1. This training schedule was used instead of training saline and cocaine on separate days because of the constraints involved in doing developmental studies. To ensure that the entire experiment could be completed within the adolescent period, it was important to have the CPP training period be as short as possible.
Table 1.
Time course of locomotor activity/conditioned place preference (CPP) experiment.
| MDMA/VEH Pretreatment + Locomotor Activity | CPP Training COC/SAL | CPP Test 1 | CPP Test 2 | |
|---|---|---|---|---|
| Experiment Day | 1–7 | 12–14 | 15 | 29 |
| PAM Age (PND) | 33–39 | 44–46 | 47 | 61 |
| ADM Age (PND) | 60–66 | 71–73 | 74 | 88 |
2.5 Data Analysis
Locomotor activity data were analyzed using a 3-way ANOVA (treatment x age x day). Post hoc analysis using Fisher’s Protected Least Significant Difference (PLSD) was used when warranted. P values less than 0.05 were considered significant for all tests.
For the CPP studies, a two-way ANOVA (pretreatment x age) was used to compare the MDMA-pretreated groups to the vehicle-pretreated group in both the adolescents and the adults. Post-hoc analysis using Fisher’s Protected Least Significant Difference (PLSD) was used when warranted. In addition, data were analyzed using t-tests to compare the preference data (time spent in cocaine-paired side minus time in saline-paired side) to 0 to determine whether a significant preference (or aversion) had occurred in each group. If equal time was spent in both sides, the preference score would be 0. P values less than 0.05 were considered significant for all tests.
3. Results
3.1 Locomotor Activity
A dose of 2.0 mg/kg MDMA stimulated activity in both adult and adolescent rats on each day of testing, although the activity levels were higher in the adult rats (Fig. 1). There was a significant treatment effect overall (F(1,112) = 43.55, p = 0.0001), and post-hoc tests showed that there was a significant difference between MDMA and vehicle treatment in both adults (p = 0.0001) and adolescents (p = 0.01). The time-course of the locomotor-stimulating effects of this dose of MDMA within the 60 min sessions was different in the adult and adolescent rats, with significant increases seen after the first 5 min of testing in the adults (Fig. 2, right panels), but not seen until after 15–20 min of testing in the adolescents (Fig. 2, left panels). Sensitization to the locomotor-activating effects of MDMA occurred at this dose in the adult rats, with significantly greater activity recorded on days 2, 6, and 7 compared to day 1 (p < 0.007). There was no sensitization to the locomotor-stimulating effects of this dose of MDMA in the adolescent rats.
Fig. 1.

Daily locomotor activity in adolescent male (PAM, left panel) and adult male (ADM, right panel) rats in response to 2.0 mg/kg/day MDMA or vehicle (saline). Data show the amount of horizontal activity in a 60 min test session, which was begun 15 min after injection of MDMA or saline. Rats were injected daily for 7 days and locomotor activity was tested on days 1,2, 6, and 7. *significant difference from vehicle (p ≤ 0.05). #significant difference from day 1 (p ≤ 0.05).
Fig. 2.

Time courses of locomotor activity on each of the test days in both adolescent male (PAM, left panels) and adult male (ADM, right panels) rats after 2.0 mg/kg MDMA or vehicle. These data show the amount of horizontal activity per 5 min period over a 60 min test session on each of the test days.
After 5.0 mg/kg of MDMA, activity was not significantly increased on the first day of testing in either group compared to vehicle (Fig. 3). In addition, treatment with these doses of MDMA did not produce an observable serotonin syndrome or any type of stereotypy that would interfere with the ability of the animals to exhibit locomotor behavior in either group on any of the test days. On the subsequent days of testing, there were significant increases in locomotor activity in both the adult and the adolescent rats in response to MDMA. There was an overall significant effect of drug (F(1,112) = 61.11, p = 0.0001), and post-hoc tests showed that MDMA produced significantly greater activity than vehicle in both adult and adolescent rats (p = 0.0001). In addition, there was a significant drug x day effect (F(3,112) = 4.45, p = 0.005). At this dose, the time-course for locomotor-stimulating effects of MDMA was not different in the adults and adolescents, with significant effects observed after 10 min of testing in both groups on days 6 and 7 of treatment (Fig. 4). On the second day of treatment, there was a delay in the effect of MDMA in the adult rats, however, with significant increases in activity not evident until 25 min after the session started. Sensitization to the locomotor-stimulating effects of this dose of MDMA occurred with significantly greater activity levels observed on days 2, 6, and 7 compared to day 1 (p < 0.05) in the adolescent rats and on day 7 (p < 0.05) in the adult rats.
Fig. 3.

Daily locomotor activity in adolescent male (PAM, left panel) and adult male (ADM, right panel) rats in response to 5.0 mg/kg/day MDMA or vehicle (saline). Data show the amount of horizontal activity in a 60 min test session, which was begun 15 min after injection of MDMA or saline. Rats were injected daily for 7 days and locomotor activity was tested on days 1,2, 6, and 7. *significant difference from vehicle (p ≤ 0.05). #significant difference from day 1 (p ≤ 0.05).
Fig. 4.
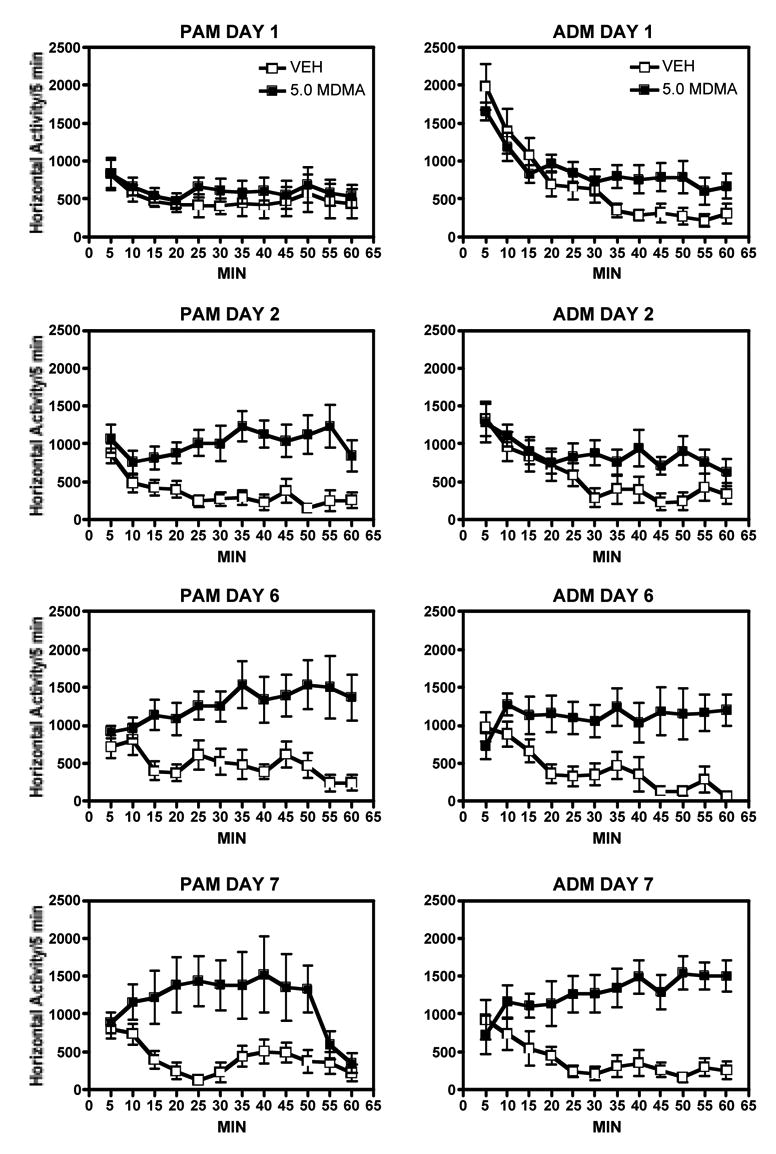
Time courses of locomotor activity on each of the test days in both adolescent male (PAM, left panels) and adult male (ADM, right panels) rats after 5.0 mg/kg MDMA or vehicle. These data show the amount of horizontal activity per 5 min period over a 60 min test session on each of the test days. At this dose, the time course of the effect of MDMA is the same in the adult and adolescent rats after several days of treatment, however on day 2 the onset of locomotor stimulation occurs sooner in the adolescent rats.
MDMA did not alter the growth of the rats, in that there were no significant differences in body weight on any day of the treatment period compared to vehicle in either the adolescent or adult rats.
3.2 Conditioned Place Preference
Similar results were seen after 2.0 or 5.0 mg/kg MDMA pretreatment. In the adult rats, after three days of pairing cocaine and saline with opposite sides of the CPP chambers, the vehicle pretreated rats exhibited a significant preference for the cocaine-paired side (t7=2.85, p = 0.025), and the preference was no longer significant after either 2.0 or 5.0 mg/kg MDMA pretreatment (Fig. 5B). The vehicle-pretreated rats spent significantly more time in the chamber previously paired with cocaine than in the chamber previously paired with saline. These data are presented as preference scores (time spent in cocaine-paired chamber minus time spent in saline-paired chamber). If the value is 0, the rat spent equal time in both chambers and there was no preference. A positive value represents more time spent in the cocaine-paired chamber and a negative value means less time spent in the cocaine-paired chamber than in the saline-paired chamber. After MDMA-pretreatment, the preference diminished. There was no difference between the effects of vehicle in the two experiments (one with the 2.0 mg/kg dose of MDMA and the other with the 5.0 mg/kg dose of MDMA) so the vehicle data were combined and are represented as a single bar on the graphs.
Fig. 5.
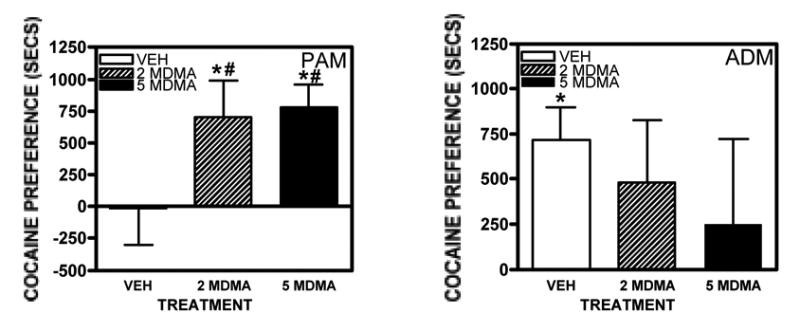
Cocaine conditioned place preference in adolescent male (PAM) and adult male (ADM) rats. Rats were trained with cocaine (10 mg/kg/day) and saline from days 12–14 of the experiment (beginning 5 days after last treatment with vehicle, 2.0 mg/kg MDMA or 5.0 mg/kg MDMA). On day 15, preference was tested by allowing rats to have access to both sides of the chamber. The time spent on each side (secs) was recorded. Data are presented as preference values (time spent in cocaine-paired chamber minus time spent in saline-paired chamber expressed as seconds). The 0 line represents no preference. *significant difference from 0. #significant difference from vehicle pretreatment.
In the adolescent rats, there was not a significant overall preference to this dose of cocaine in the vehicle-pretreated rats, but there was a significant preference after pretreatment with 2.0 mg/kg MDMA (t7=2.48, p = 0.042, Fig. 5A). The adolescent rats pretreated with 5.0 mg/kg MDMA also had a significant preference for the cocaine-paired side (t7 = 4.49, p = 0.0028). Thus, conditioned place preference to cocaine was increased in the adolescents, and decreased in the adults subsequent to a 7-day pretreatment with MDMA, compared to vehicle.
There was a significant pretreatment x age effect for preference scores in the adult and adolescent rats pretreated with vehicle, 2 mg/kg MDMA or 5.0 mg/kg MDMA (F(2,58) = 4.44, p = 0.016). Post-hoc tests showed that there was a significant difference between pretreatment with vehicle and pretreatment with either 2.0 or 5.0 mg/kg MDMA in the adolescent rats (p < 0.05) but not in the adult rats. It is interesting to note that there was a large individual variability in the preference scores in the adult rats after pretreatment with 5.0 mg/kg MDMA (as shown by the large error bar on the MDMA point on Fig. 5B) that was not evident after vehicle. This was because 7/8 of the rats showed a large preference for the cocaine-paired side after vehicle-pretreatment, whereas after MDMA pretreatment, only 5 of the 8 rats showed a preference to cocaine while 3 rats showed a significant aversion to cocaine (shown by a negative preference value because the rats spent significantly more time in the saline-paired chamber than in the cocaine-paired chamber). Of the five who showed a preference to cocaine, only 4 showed a greater than 50% preference. Thus, there appeared to be a dichotomous effect of MDMA pretreatment on subsequent cocaine CPP in the adult rats, with no effect in some rats and a negative effect in others. This was not the case in the adolescent rats, all of which had positive preference scores after MDMA treatment.
The CPP test two weeks later (on day 29 of the experiment) produced virtually identical results to those seen on day 15 (Fig. 6). As seen earlier, there was not a significant preference after vehicle-pretreatment in adolescent rats. There was, however, a significant preference for the cocaine-paired side in the adolescent rats pretreated with 5.0 mg/kg MDMA (t7 = 5.49, p = 0.0009). Once again, the adult rats that were pretreated with vehicle exhibited a significant preference for the cocaine-paired side (t7 = 2.43, p = 0.045), while after MDMA treatment the adult rats did not have a preference that was significantly different from 0. An overall ANOVA of the retest data showed that there was a significant pretreatment x age effect (F(1,28) = 4.92, p = 0.0348). As seen in the earlier CPP test on day 15, there was a significant difference between MDMA-pretreated and vehicle-pretreated adolescent rats (p < 0.05), but no significant difference in the preference scores of the vehicle-pretreated and MDMA-pretreated adult rats.
Fig. 6.

Retest of cocaine conditioned place preference in adolescent male (PAM) and adult male (ADM) rats 2 weeks after first test. Rats were trained with cocaine (10 mg/kg/day) and saline from days 12–14 of the experiment (beginning 5 days after last treatment with vehicle or 5.0 mg/kg MDMA). On day 29, preference was tested by allowing rats to have access to both sides of the chamber. The time spent on each side (secs) was recorded. Data are presented as preference values (time spent in cocaine-paired chamber minus time spent in saline-paired chamber expressed as seconds). The 0 line represents no preference. *significant difference from 0. #significant difference from vehicle pretreatment.
There were no significant differences between the preference scores in the first test vs the second test in the vehicle-pretreated adult or the adolescent rats. Similarly, there were no differences in the scores after MDMA pretreatment across tests in either age group. Thus, the effects observed one day after CPP training were still evident 2 weeks later.
4. Discussion
4.1 Locomotor Activity
The data from the locomotor activity studies showed that a dose of 2 mg/kg of MDMA stimulated activity in both adult and adolescent rats, although the activity levels in response to the MDMA were higher in adult rats than in adolescents. The time-course of the activating effects of this dose of MDMA was different in adults and adolescent rats with a significant increase in activity observed after 5 minutes in adult rats compared to 15–20 minutes in adolescents. Adult rats were sensitized to the locomotor-activating effect of this dose of MDMA, in that there were significantly higher activity levels on days 2, 6 and 7 compared to day 1. No sensitization to the locomotor-activating effects of MDMA occurred in the adolescent rats in response to the daily administration of 2.0 mg/kg MDMA for 7 days.
When a higher daily dose of MDMA (5 mg/kg/day) was administered, neither adult nor adolescent rats showed a significant increase in locomotor activity on the first day of the experiment compared to vehicle-treated rats. On the remaining days however, both adult and adolescent MDMA treated rats showed significantly higher locomotor-activity than vehicle-treated rats. With this dose there were no differences in the time-course of the locomotor-activity between the adult and adolescent rats. A significant increase in activity was seen after 10 minutes on days 6 and 7 in both groups. On the second day of MDMA treatment, there was a delay in the activating effect of MDMA in the adult rats with a significant effect not seen until 25 minutes into the test session.
After 5.0 mg/kg MDMA daily, sensitization was evident in adult rats on days 2, 6 and 7 and in adolescent rats on day 7. An earlier study showed that rats treated with MDMA (5.0 mg/kg s.c.) for 5 days were sensitized to the locomotor-activating effects of a challenge dose of MDMA 2 days after the last injection [18]. In addition, the rats remained sensitized to a challenge injection 11 days after the first test. Although a higher dose of MDMA (15 mg/kg/day) produced sensitization to the locomotor-activating effects of cocaine, the 5 mg/kg dose did not produce any crossover effects. In another study, sensitization occurred, although it is not clear to which doses, in rats that received intermittent injections with MDMA (2.5, 5.0 or 7.5 mg/kg) every other day for 6 or 12 days [28]. In another study, adult rats were treated with lower doses of MDMA (2.5, 5.0 or 10.0 mg/kg/day) for six days and challenged with MDMA 5 and 32 days later [23]. This was followed by amphetamine administration or methylphenidate administration one day after the second challenge dose of MDMA. As in the present study, these adult rats experienced sensitization to the locomotor-activating effects of MDMA but did not show cross-sensitization to amphetamine or methylphenidate.
The elevated locomotor activity seen in rats treated with MDMA is postulated to be mediated by release of serotonin [for review see 15], and at least in part, by 5-HT2A receptors [16]. In adolescent rats, the density of serotonin transporters is increasing and does not reach adult levels until PND 90 [24]. It also has been shown that the serotonin syndrome differs as a function of age in the adolescent rat, where rats aged PND 35 have a more pronounced serotonin syndrome to MDMA than rats aged PND 45 and PND 60 [27]. This difference in the serotonin response to MDMA across development might explain why adolescent rats are less sensitive than the adults to the locomotor-activating effects of MDMA.
It appears that the adolescent rats are less vulnerable to the sensitization that occurs after repeated treatment with MDMA since they did not show sensitization with the low dose (2.0 mg/kg) of MDMA and although sensitization did develop to the higher dose (5.0 mg/kg) it was only evident on the seventh treatment day. It is interesting to note that previous studies in our laboratory have shown that sensitization developed to the locomotor activating effects of cocaine or nicotine in adult, but not in adolescent male rats [12, 13]. Thus, in general, it appears that sensitization to the locomotor activating effects of psychostimulant drugs occurs more readily in adults than in adolescents. It has been suggested that the sensitization to MDMA might be due to elevated concentrations of dopamine in the nucleus accumbens [18]. Studies conducted in adolescent and adult rats, where a very high dose of MDMA (40 mg/kg) was administered on PND 40 or PND 70, showed that MDMA increased dopamine levels acutely in the caudate putamen at both ages [9]. However, it is known that during adolescence there are significant developmental changes in the dopamine system [8, 24]. During this period, there is a 2-fold reduction in dopamine turnover, but no change in density of dopamine uptake sites. In addition, during adolescence dopamine D2 receptors and GTP inhibition of adenylyl cyclase activity appear. Thus, it is possible that differential behavioral effects of MDMA during this developmental period could be due to an altered development of the dopamine system.
4.2 Conditioned Place Preference
The adolescent rats pretreated with vehicle did not develop a significant CPP to 10 mg/kg cocaine after three training sessions. An earlier study showed that cocaine-induced place conditioning occurred to the same degree in adolescents (PND 32) as in adults (PND >65), although in that study data from male and female rats were combined [10]. In addition, there were significant differences between the males and females, with the females having higher choice ratios than the males after training with both 5.0 and 10.0 mg/kg cocaine. It was not clear whether the males alone had a significant CPP, since the data were not presented separately. A previous study comparing different age groups in developing mice showed that there was less of a preference for cocaine at PND 28–31 than at earlier ages, although adult mice were not included in this study [19]. In contrast, while conditioned place preference to amphetamine developed in adult (PND >60) rats, there was no reliable conditioned place preference in adolescent (PND 30) rats trained with amphetamine [2]. It also has been shown that while nicotine conditioned place preference occurred during early adolescence (PND 28), it did not induce a significant positive place preference during later adolescence [5, 30], corresponding to the time when the rats in this study were tested for CPP. These data, combined with our present data suggest that the conditioning effects of drugs might be different in adult and adolescent rats. In the present study, there was a clear difference between the control adult and adolescent rats, with only the adult control animals showing a significant preference for this dose and training regimen with cocaine. Additional studies, with additional doses of cocaine, and multiple time courses of training will be needed to better understand the differences between adolescent and adult rats in the development of conditioned reward.
These data show that while cocaine produced a robust CPP in adult rats pretreated with vehicle, after MDMA pretreatment there is no longer a significant CPP to cocaine in adult rats. As mentioned in the results section, this is likely due to an increase in the individual variability that was observed subsequent to MDMA pretreatment. Only 5 of the 8 rats in the 5.0 mg/kg MDMA-pretreated group showed a positive CPP, while 3 of the rats showed a significant negative preference. Thus, it is possible that MDMA has a dichotic effect with the responses of some animals to cocaine being positive, while other rats appear to develop an aversion to cocaine after pretreatment with MDMA. In addition, as mentioned above, the magnitude of the preference to the cocaine-paired side, in those animals that showed this preference, was decreased in the adult rats after treatment with MDMA. These results persisted and the same effects were observed 14 days later. The same effects were observed after the lower dose of MDMA, with fewer rats exhibiting a positive preference, and with a decreased overall magnitude of the preference in those animals that did prefer the cocaine-paired side.
A previous study using a high dose of MDMA in adults (10 mg/kg x 3 injections in a single day) did not alter cocaine (20 mg/kg) CPP when training began 14 days later [11]. In that study, however, rats were treated for only a single day with high doses of MDMA, and their first exposure to cocaine was two weeks later. This is in contrast to the present study, where the first cocaine exposure occurred 5 days after a seven-day MDMA treatment period. One study found that pretreatment with a high dose of MDMA (20 mg/kg) twice daily for four days to adult rats increased the conditioned response to cocaine (5 or 10 mg/kg) when CPP was tested two weeks later [17]. This finding is contrary to the findings in the present study, however, the MDMA dose used in that study is much higher than in the present study and the dosing regimen is different. The use of a potentially toxic dose of MDMA could well alter the subsequent response to cocaine compared to the lower doses used in the present study. For example, there have been a number of studies showing that high doses of MDMA have a toxic effect on serotonin in adult and adolescent rats. In adolescent rats treated with 5 mg/kg MDMA four times daily (total of 20 mg/kg/day) every fifth day between PND 35 and 60, serotonin transporter binding was decreased in the cortex [27], and rats administered a single high dose of MDMA (20 mg/kg) at PND 35 and killed seven days later exhibited significant reductions in serotonin [3, 4]. In addition, administration of 20 or 40 mg/kg MDMA on PND 40 or PND 70 led to decreased serotonin levels in multiple brain regions, including the frontal cortex and striatum [9], with no changes seen after a lower dose of 10 mg/kg. In the caudate putamen, only the 40 mg/kg dose of MDMA produced a significant decrease in serotonin levels at PND 40, while both 20 and 40 mg/kg reduced serotonin at PND 70. Thus, it may be that the adult rats are slightly more sensitive than the adolescent rats to the toxic effect of MDMA on serotonin. In addition, 40 mg/kg MDMA administered on PND 40 or PND 70 acutely increased dopamine levels in the caudate putamen for several hours after administration [9].
In contrast, rats treated repeatedly with a somewhat lower dose of MDMA (7.5 mg/kg ip twice daily for 3 days) starting at PND 39 had no changes in serotonin levels or cortical serotonin transporter density 12 days later [14]. In contrast to the adult rats, in adolescent rats, MDMA pretreatment appeared to increase the conditioned reinforcement of cocaine. The vehicle-treated rats did not show significant preference for the cocaine paired side. An earlier study with a much higher dose of MDMA (7.5 mg/kg twice daily) showed that pretreatment for 3 days from PND 39–41, led to a slight increase in cocaine CPP 26 days later, when the rats were adults, compared to saline pretreatment combined with saline CPP [14]. As in the present experiment, however, cocaine alone (5 mg/kg) did not produce a significant CPP after two days of training in saline pretreated rats. In contrast to the present findings, there was no difference between saline pretreated rats conditioned with cocaine and MDMA pretreated rats conditioned with cocaine [14]. A study in mice found that exposure to MDMA during the adolescent period (10 mg/kg for 7 days) increased conditioned reward induced by cocaine in adulthood (after a withdrawal period of six weeks) [1]. Thus, unlike what is observed in adult rats or mice, if the exposure to MDMA occurs during adolescence, there seems to be crossover effects to other psychostimulants such that there are increased rewarding effects of cocaine during adolescence and that this persists into adulthood.
In summary, repeated administration of MDMA to adolescent rats produces differential behavioral effects compared to adult rats. Under the conditions used in this study, it appears that MDMA increased the conditioned reward associated with the tested dose of cocaine in the adolescent rats and decreased it in the adult rats. These findings suggest that exposure to MDMA during this critical developmental period may carry a greater risk than during adulthood and that male adolescents may be particularly vulnerable to the risk of stimulant abuse after use of MDMA. This is consistent with the data showing that MDMA users are more likely to use other illicit drugs than are non-MDMA users [25]. The data further suggest that it is possible to model this behavior in the laboratory and that it will be possible to study the neurochemical adaptations that occur in response to MDMA that may lead to the increased effects of subsequent drug administration.
Acknowledgments
This work was supported by the National Institute on Drug Abuse grant DA 15119.
Footnotes
Conflict of Interest
None of the authors have any conflicts of interest related to this manuscript.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Achat-Mendes C, Anderson KL, Itzhak Y. Methylphenidate and MDMA adolescent exposure in mice: long-lasting consequences on cocaine-induced reward and psychomotor stimulation in adulthood. Neuropharmacol. 2003;45:106–15. doi: 10.1016/s0028-3908(03)00135-7. [DOI] [PubMed] [Google Scholar]
- 2.Adriani W, Laviola G. Elevated levels of impulsivity and reduced place conditioning with d-amphetamine: two behavioral features of adolescence in mice. Behav Neurosci. 2003;117:695–703. doi: 10.1037/0735-7044.117.4.695. [DOI] [PubMed] [Google Scholar]
- 3.Aguirre N, Ballaz S, Lasheras B, Rio JD. MDMA (‘Ecstasy’) enhances 5-HT1A receptor density and 8-OH-DPAT-induced hypothermia: blockade by drugs preventing 5-hydroxytryptamine depletion. Eur J Pharmacol. 1998;346:181–189. doi: 10.1016/s0014-2999(98)00062-4. [DOI] [PubMed] [Google Scholar]
- 4.Aguirre N, Barrionuevo M, Lasheras B, Rio JD. The role of dopaminergic systems in the perinatal sensitivity to 3,4-methylenedioxymethamphetamine-induced neurotoxicity in rats. J Pharmacol Exp Ther. 1998;286:1159–1165. [PubMed] [Google Scholar]
- 5.Belluzzi JD, Lee AG, Oliff HS, Leslie FM. Age-dependent effects of nicotine on locomotor activity and conditioned place preference in rats. Psychopharmacol. 2004;174:389–395. doi: 10.1007/s00213-003-1758-6. [DOI] [PubMed] [Google Scholar]
- 6.Bilsky E, Montegut M, Nichols M, Reid L. CGS 10746B, a novel dopamine release inhibitor, blocks the establishment of cocaine and MDMA conditioned place preference. Pharmacol Biochem Behav. 1998;59:215–220. doi: 10.1016/s0091-3057(97)00424-3. [DOI] [PubMed] [Google Scholar]
- 7.Braida D, Iosue S, Pegorini S, Sala M. 3,4 Methylenedioxymethamphetamine-induced conditioned place preference (CPP) is mediated by endocannabinoid system. Pharmacol Res. 2005;51:177–82. doi: 10.1016/j.phrs.2004.07.009. [DOI] [PubMed] [Google Scholar]
- 8.Broaddus WC, Bennett JP., Jr Postnatal development of striatal dopamine function. I. An explanation of D1 and D2 receptors, adenylate cyclase regulation and presynaptic markers. Brain Research Developmental Brain Research. 1990;52:275–271. doi: 10.1016/0165-3806(90)90244-s. [DOI] [PubMed] [Google Scholar]
- 9.Broening HW, Bacon L, Slikker JW. Age modulates the long-term but not the acute effects of the serotonergic neurotoxicant 3,4-methylenedioxymethamphetamine. J Pharmacol Exp Ther. 1994;271:285–293. [PubMed] [Google Scholar]
- 10.Campbell JO, Wood RD, Spear LP. Cocaine and morphine-induced place conditioning in adolescent and adult rats. Physiology and Behavior. 2000;68:487–493. doi: 10.1016/s0031-9384(99)00225-5. [DOI] [PubMed] [Google Scholar]
- 11.Cole JC, Sumnall HR, O’Shea E, Marsden CA. Effects of MDMA exposure on the conditioned place preference produced by other drugs of abuse. Psychopharmacol. 2003;166:383–90. doi: 10.1007/s00213-002-1374-x. [DOI] [PubMed] [Google Scholar]
- 12.Collins SL, Izenwasser S. Cocaine differentially alters behavior and neurochemistry in periadolescent versus adult rats. Dev Brain Res. 2002;138:27–34. doi: 10.1016/s0165-3806(02)00471-6. [DOI] [PubMed] [Google Scholar]
- 13.Collins SL, Izenwasser S. Chronic nicotine differentially alters cocaine-induced locomotor activity in adolescent vs. adult male and female rats. Neuropharmacol. 2004;46:349–362. doi: 10.1016/j.neuropharm.2003.09.024. [DOI] [PubMed] [Google Scholar]
- 14.Fone KC, Beckett SR, Topham IA, Swettenham J, Ball M, Maddocks L. Long-term changes in social interaction and reward following repeated MDMA administration to adolescent rats without accompanying serotonergic neurotoxicity. Psychopharmacol. 2002;159:437–444. doi: 10.1007/s00213-001-0931-z. [DOI] [PubMed] [Google Scholar]
- 15.Green AR, Cross AJ, Goodwin GM. Review of the pharmacology and clinical pharmacology of 3,4-methylenedioxymethamphetamine (MDMA or “Ecstasy”) Psychopharmacol. 1995;119:247–260. doi: 10.1007/BF02246288. [DOI] [PubMed] [Google Scholar]
- 16.Herin DV, Liu S, Ullrich T, Rice KC, Cunningham KA. Role of the serotonin 5-HT2A receptor in the hyperlocomotive and hyperthermic effects of (+)-3,4-methylenedioxymethamphetamine. Psychopharmacol. 2005;178:505–13. doi: 10.1007/s00213-004-2030-4. [DOI] [PubMed] [Google Scholar]
- 17.Horan B, Gardner EL, Ashby CR., Jr Enhancement of conditioned place preference response to cocaine in rats following subchronic administration of 3, 4-methylenedioxymethamphetamine (MDMA) Synapse. 2000;35:160–2. doi: 10.1002/(SICI)1098-2396(200002)35:2<160::AID-SYN9>3.0.CO;2-O. [DOI] [PubMed] [Google Scholar]
- 18.Kalivas P, Duffy P, White S. MDMA elicits behavioral and neurochemical sensitization in rats. Neuropsychopharmacol. 1998;18:469–479. doi: 10.1016/S0893-133X(97)00195-4. [DOI] [PubMed] [Google Scholar]
- 19.Laviola G, Dell’Omo G, Alleva E, Bignami G. Ontogeny of cocaine hyperactivity and conditioned place preference in mice. Psychopharmacol. 1992;107:221–8. doi: 10.1007/BF02245141. [DOI] [PubMed] [Google Scholar]
- 20.Marona-Lewicka D, Rhee GS, Sprague JE, Nichols DE. Reinforcing effects of certain serotonin-releasing amphetamine derivatives. Pharmacol Biochem Behav. 1996;53:99–105. doi: 10.1016/0091-3057(95)00205-7. [DOI] [PubMed] [Google Scholar]
- 21.McNamara MG, Kelly JP, Leonard BE. Some behavioural and neurochemical aspects of subacute (±)3,4-methylenedioxymethamphetamine administration in rats. Pharmacol Biochem Behav. 1995;52:479–484. doi: 10.1016/0091-3057(95)00206-c. [DOI] [PubMed] [Google Scholar]
- 22.Meyer A, Mayerhofer A, Kovar KA, Schmidt WJ. Rewarding effects of the optical isomers of 3,4-methylenedioxy-methylamphetamine (‘Ecstasy’) and 3,4-methylenedioxy-ethylamphetamine (‘Eve’) measured by conditioned place preference in rats. Neurosci Lett. 2002;330:280–4. doi: 10.1016/s0304-3940(02)00821-2. [DOI] [PubMed] [Google Scholar]
- 23.Modi GM, Yang PB, Swann AC, Dafny N. Chronic exposure to MDMA (Ecstasy) elicits behavioral sensitization in rats but fails to induce cross-sensitization to other psychostimulants. Behav Brain Funct. 2006;2:1. doi: 10.1186/1744-9081-2-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Moll GH, Mehnert C, Wicker M, Bock N, Rothenberger A, Ruther E, Huether G. Age-associated changes in the densities of presynaptic monoamine transporters in different regions of the rat brain from early juvenile life to late adulthood. Dev Brain Res. 2000;119:251–257. doi: 10.1016/s0165-3806(99)00182-0. [DOI] [PubMed] [Google Scholar]
- 25.NSDUH, Substance Abuse and Mental Health Services Administration. Results from the 2004 National Survey on Drug Use and Health: National Findings (Office of Applied Studies, NSDUH Series H-28, DHHS Publication No. SMA 05-4062) Rockville, MD: 2005. 2005. [Google Scholar]
- 26.O’Shea E, Granados R, Esteban B, Colado MI, Green AR. The relationship between the degree of neurodegeneration of rat brain 5-HT nerve terminals and the dose and frequency of administration of MDMA (‘ecstasy’) Neuropharmacol. 1998;37:919–26. doi: 10.1016/s0028-3908(98)00029-x. [DOI] [PubMed] [Google Scholar]
- 27.Piper BJ, Fraiman JB, Meyer JS. Repeated MDMA (“Ecstasy”) exposure in adolescent male rats alters temperature regulation, spontaneous motor activity, attention, and serotonin transporter binding. Dev Psychobio. 2005;47:145–57. doi: 10.1002/dev.20085. [DOI] [PubMed] [Google Scholar]
- 28.Spanos LJ, Yamamoto BK. Acute and subchronic effects of methylenedioxymethamphetamine [(±) MDMA] on locomotion and serotonin syndrome behavior in the rat. Pharmacol Biochem Behav. 1989;32:835–840. doi: 10.1016/0091-3057(89)90044-0. [DOI] [PubMed] [Google Scholar]
- 29.Spear LP, Brake SC. Periadolescence: age-development behavior and psychopharmacological responsivity in rats. Dev Psychobio. 1983;16:83–109. doi: 10.1002/dev.420160203. [DOI] [PubMed] [Google Scholar]
- 30.Torrella TA, Badanich KA, Philpot RM, Kirstein CL, Wecker L. Developmental differences in nicotine place conditioning. Ann N Y Acad Sci. 2004;1021:399–403. doi: 10.1196/annals.1308.052. [DOI] [PubMed] [Google Scholar]