

Abstract
Understanding mechanisms responsible for meiotic resumption in mammalian oocytes is critical for the identification of strategies to enhance developmental competence of in vitro matured oocytes. Improvement of in vitro oocyte maturation systems is dependent on a better understanding of mechanisms that regulate oocyte maturation both in vivo and in vitro as well as on the identification of methods to manipulate the meiotic progression of oocytes matured in vitro in a physiological manner. The purpose of this review is two-fold: first, to examine the mechanisms that underlie the acquisition of oocyte developmental competence and regulation of oocyte maturation in vivo and in vitro; and second, to present data examining the role of transcription in mediating the ability of EGF and FSH to induce oocyte maturation in vitro. Results presented support the conclusions that (1) EGF-induced oocyte maturation does not require nascent gene transcription in both mice and domestic cats; (2) FSH requires gene transcription to induce oocyte maturation in both species; (3) EGF must be present in the maturation medium to optimize the effectiveness of FSH to promote oocyte maturation; and (4) the mechanism used by FSH to induce oocyte maturation in vitro appears to predominate over that used by EGF when both EGF and FSH are present in maturation medium used for either murine or feline cumulus oocyte complexes.
Keywords: Meiosis, FSH, EGF, Oocyte maturation, Gene transcription
1. Introduction
1.1. Developmental competence of oocytes
Developmental competence is used to connote the ability of an oocyte to achieve successful fertilization and development throughout at least the preimplantation period (Hyttel et al., 1997; Gandolfi and Gandolfi, 2001; Sirard, 2001). Differences exist in the developmental competence of oocytes matured in vivo compared to those matured in vitro. For example in cattle, approximately 60 to 80% of oocytes obtained from follicles matured in vivo are competent to develop to the blastocyst stage (Bordignon et al., 1997; Blondin et al., 2002; Dieleman et al., 2002; Rizos et al., 2002) compared to only 25 to 40% of oocytes matured in vitro (van de Leemput et al., 1999; Rizos et al., 2002; Ward et al., 2002). Similarly, in humans the efficiency of embryo development following in vitro oocyte maturation procedures is less compared to development following recovery of in vivo matured oocytes (Child et al., 2001; Trounson et al., 2001; Yoon et al., 2001; Picton, 2002). Furthermore, oocytes that have been exposed to a gonadotropin stimulus either in vivo or during in vitro culture are more competent to support embryonic development following fertilization (Izadyar et al., 1998; Child et al., 2001; Trounson et al., 2001). Although in practice we can recognize differences that illustrate the concept of oocyte developmental competence, a precise definition of developmental competence in specific physiological terms is still lacking (Duranthon and Renard, 2001).
Understanding mechanisms responsible for meiotic resumption in mammalian oocytes is critical for the identification of strategies to enhance developmental competence of in vitro matured oocytes, thereby improving the effectiveness of assisted reproduction techniques used for domestic livestock, wildlife species and human clinical applications. Improvement of in vitro oocyte maturation systems is dependent on a better understanding of mechanisms that regulate oocyte maturation both in vivo and in vitro as well as on the identification of methods to manipulate the meiotic progression of oocytes matured in vitro in a physiological manner. The purpose of this review is two-fold: first, to examine the mechanisms that underlie the acquisition of oocyte developmental competence and the regulation of oocyte maturation in vivo and in vitro; and second, to present data specifically examining the role of transcription in mediating the ability of EGF and FSH to induce oocyte maturation in vitro.
1.2 Acquisition of oocyte developmental competence in vivo
Following their migration from the yolk sac, primordial germ cells undergo mitotic divisions as they colonize the genital ridge. As the population of primordial germ cells differentiate into oogonia, their population continues to expand by mitotic divisions until they enter meiosis. To date, the mechanisms that signal this critical cell cycle juncture have yet to be elucidated. As the individual oocytes enter into prophase I of meiosis, they quickly become associated with pregranulosa cells (McLaren, 1988) and form primordial follicles. These pregranulosa cells are likely responsible for generating the signal that results in the arrest of the primary oocytes at the diplotene stage of prophase I since oocytes not surrounded by pregranulosa cells continue to progress through meiosis, eventually leading to their demise (McLaren, 1988). Formation of the primordial follicle is also associated with a decondensation of the oocyte chromatin and formation of the diffuse diplotene or dictyate stage of meiosis. Folliculogenesis proceeds as the primordial follicle is activated to become a primary follicle. Primordial follicle activation is not well understood, however, several candidate signaling molecules have been implicated in this process including kit ligand (Parrott and Skinner, 1999), anti-mullerian hormone (Durlinger et al., 2002) and H1FOO (Tanaka et al., 2005).
Throughout oocyte growth and development, the intimate association formed between the oocyte and its surrounding granulosa/cumulus granulosa cells is a critical requirement for normal follicular growth and acquisition of oocyte developmental competence (Carabatsos et al., 2000; Eppig et al., 2002; Matzuk et al., 2002). Initially, this association was recognized as critical for oocyte growth, with cumulus granulosa cells known to provide metabolic precursors such as sugars, amino acids and nucleotides to the oocyte (Brower and Schultz, 1982; Wassarman and Albertini, 1994). In addition, unidirectional communication from the cumulus cells to the oocyte mediated by gap junctional communication has been recognized as critical for the initiation of oocyte maturation in response to gonadotropin treatment in both laboratory and domestic species (Fagbohun and Downs, 1991; Carabatsos et al., 2000; Vozzi et al., 2001). Since the decade starting in 1990, it has been recognized that communication between the oocyte and its surrounding cumulus granulosa cells is not just unidirectional but bidirectional (Buccione et al., 1990; Vanderhyden and Armstrong, 1990; Vanderhyden et al., 1990; Vanderhyden et al., 1992; Simon et al., 1997; Eppig et al., 2002; Matzuk et al., 2002). For example, oocyte-secreted factors facilitate cumulus expansion (Vanderhyden and Armstrong, 1990; Vanderhyden et al., 1990), proliferation of granulosa cells (Vanderhyden et al., 1992), suppression of LH receptors in cumulus cells (Eppig et al., 1997) and influence ovulation (Matzuk et al., 2002). Thus, throughout oocyte growth and maturation, association of the oocyte with its surrounding cumulus granulosa cells is essential not only for normal function of the oocyte but also for function of the follicle within which the cumulus-oocyte complex (COC) resides.
In mice, as ovarian follicles progress through the primary and secondary stage of follicular development, oocytes increase in diameter approximately three-fold, reaching about 80 μm in diameter prior to formation of the follicular antrum (Wassarman and Albertini, 1994). Associated with this period of oocyte growth is the acquisition of meiotic and developmental competence. Meiotic competence is acquired in a progressive manner as the murine oocyte first becomes capable to undergo germinal vesicle breakdown (GVBD), then to progress to metaphase I and, finally, to reach metaphase II. After acquisition of meiotic competence, the oocyte gains developmental competence and is capable of undergoing fertilization and then post-fertilization development (Sorensen and Wassarman, 1976; Eppig et al., 1994). Thus, acquisition of meiotic competence is followed by acquisition of developmental competence in a sequential manner and both are components of a single continuous process, rather than two discrete processes. In cattle, a similar progression is observed with the exception that full meiotic competence is not acquired until after formation of the follicular antrum. At a diameter of <100 μm, bovine oocytes remain incapable of undergoing germinal vesicle breakdown or GVBD (Fair et al., 1995). Competence to reach metaphase I (MI) is acquired when the bovine oocyte reaches a diameter of approximately 100 μm (Fair et al., 1995; Hyttel et al., 1997). When the oocyte is resident within a tertiary follicle of approximately 1 mm diameter, full meiotic competence is present and the bovine oocyte diameter is about 110 μm (Pavlok et al., 1992; Fair et al., 1995; Hyttel et al., 1997). Developmental competence is reached progressively as the bovine oocyte increases in diameter from 110 to 120 μm while it resides within a follicle whose diameter increases from 6 mm to greater than 10 mm (Lonergan et al., 1994; Hyttel et al., 1997; Kruip et al., 2000; Rodriguez and Farin, 2004).
Studies of oocyte ultrastructure and [3H]-uridine incorporation have revealed progressive changes in the development of oocytes during folliculogenesis. In cattle, evidence of oocyte transcriptional activity begins at about the secondary follicle stage, continues to increase through the early tertiary follicle stage (~3 mm diameter) and decreases thereafter (Hyttel et al., 1997; Tomek et al., 2002). In mice, transcriptional activity of oocytes appears to be regulated in part by the granulosa cells associated with the oocyte and can be influenced by exposure to gonadotropins (De La Fuente and Eppig, 2001). Oocyte transcriptional activity results in the accumulation of maternal preformed mRNAs, which are critical for directing development of the zygote during the preimplantation period (Vassali and Stutz, 1995; Brevini-Gandolfi et al., 1999). Although incorporation of radiolabeled substrates into the oocyte nucleus and cytoplasm decreases with advanced follicular development and increased oocyte diameter, the localization of organelles continues to change within the oocyte cytoplasm as meiotic maturation progresses. Mitochondria alter their localization from the oocyte periphery to more central portions of the ooplasm during the meiotic progression (Van Blerkom and Runner, 1984; Krisher and Bavister, 1998; Stojkovic et al., 2001). At the time of GVBD, lipid droplets within the oocyte cytoplasm are increased in number and size whereas the size of golgi apparatuses are decreased (Hyttel et al., 1997). As the oocyte progresses through meiosis to metaphase I (MI) and then proceeds to metaphase II (MII), lipid droplets continue to increase, golgi size decreases further and mitochondria become more closely associated with lipid droplets within the ooplasm (Hyttel et al., 1997; Stojkovic et al., 2001). In addition, microtubules reorganize and cortical granules are redistributed to become evenly spaced and located beneath the oolemma (Hyttel et al., 1997; Stojkovic et al., 2001).
1.3. LH and FSH in ovulation and oocyte maturation in vivo
The processes of ovulation, luteinization and the resumption of oocyte maturation are initiated as a result of the preovulatory surge of gonadotropins. Although intrafollicular mechanisms regulating ovulation have been well studied, precise mechanisms regulating the resumption of meiosis in vivo remain unclear. In mammals, the gonadotropin surge consists of coincident rises in both LH and FSH. In cattle, peak concentrations of circulating LH increase approximately 8-fold above basal levels and FSH increase about 3-fold above baseline during the normal gonadotropin surge (Haughian et al., 2004). Although it has been assumed that LH is the primary driver for ovulation and the resumption of meiosis (Tsafriri and Dekel, 1994), this hypothesis has been questioned in the past since purified preparations of FSH can also induce both ovulation and oocyte maturation (Greep et al., 1942; Goldman and Mahesh, 1969; Harrington and Elton, 1969; Jones and Nalbandov, 1972; Lipner et al., 1974; Nuti et al., 1974; Tsafriri et al., 1976; Greenwald and Papkoff, 1980; Taya and Greenwald, 1980; Galway et al., 1990). For example, intrafollicular injections of low doses of purified preparations of either LH or FSH induced ovulation of individual follicles in rabbits (Jones and Nalbandov, 1972) and sheep (Murdoch et al., 1981). These intrafollicular injections induced ovulations in individual follicles without affecting adjacent preovulatory follicles on the same ovary. Contamination of FSH preparations by LH in either study did not appear to be the explanation for these observations (Jones and Nalbandov, 1972; Murdoch et al., 1981). Furthermore, additive effects were demonstrated when subovulatory doses of LH and FSH were jointly administered into the same follicle (Jones and Nalbandov, 1972), leading to the suggestion that both LH and FSH were necessary for ovulation and luteinization. Similarly, administration of either hCG or recombinant human (rh)FSH to hypophysectomized, FSH-primed mice successfully induced ovulation and luteinization (Wang and Greenwald, 1993). Interestingly, approximately 22% of 1-cell embryos obtained from hCG-treated mice reached the blastocyst stage in vitro whereas none of the 1-cell embryos obtained from rhFSH-treated mice developed beyond the six-cell stage (Wang and Greenwald, 1993), suggesting that developmental competence of zygotes from oocytes matured under the influence of hCG was improved compared to those matured in the presence of FSH alone. Unfortunately, the effect of simultaneous administration of hCG and rhFSH on preimplantation embryo development was not tested in their study. Using a LH receptor knockout (LuRKO) mouse model to evaluate the effects of rhFSH administration on ovulation, Pakarainen et al. (2005) demonstrated that ovulation could not be induced by administration of either hCG or rhFSH in PMSG-primed immature mice when LH receptors were absent. As expected, administration of either hCG or rhFSH induced ovulation in PMSG-primed wild-type controls. These observations suggest that activation of the LH receptor itself is permissive for either hCG or rhFSH to induce ovulation (Pakarainen et al., 2005). However, the mechanism remains unclear by which activation of LH receptors occur and facilitate ovulation when highly purified FSH is used to induce ovulation in individual follicles (Murdoch et al., 1981) or when treatment with rhFSH is used to induce ovulation in hypophysectomized females (Wang and Greenwald, 1993). In each case, one would have to assume less LH activity or LH-receptor activation is present even in hypophysectomized animals and that this functions in a synergistic manner with FSH to induce terminal follicular maturation and ovulation. Taken together, these observations suggest that LH is important in inducing ovulation in follicles that have been primed by FSH exposure but it remains unclear whether in vivo LH functions alone to facilitate oocyte maturation. It is reasonable to suggest that, in vivo, FSH and LH work in concert to provide an optimal environment not only for follicular development but also for oocyte maturation.
A synergistic role for FSH and LH on oocyte maturation is consistent with data on the distribution of FSH and LH receptors found in the mural granulosa cells of the follicle and cumulus cells surrounding the oocyte. In mural granulosa cells, FSH functions through its receptor in concert with estradiol to induce granulosa LH receptors, promoting continued steroidogenesis and the development of the preovulatory follicle (Richards, 1980). In contrast, FSH receptors are the only gonadotropin receptors present on cumulus cells throughout development of preovulatory follicles (Eppig et al., 1997). Secreted factors from GV-stage murine oocytes completely blocked the production of LH receptor mRNA in granulosa cells by FSH (Eppig et al., 1997; Eppig et al., 1998). Although less effective than GV-stage oocytes, metaphase II oocytes also suppressed the induction of granulosa cell LH receptor mRNA by approximately 85% compared to controls (Eppig et al., 1997). In cattle, LH receptor mRNA could not be found in cumulus cells or cumulus oocyte complexes either before or after the initiation of oocyte maturation in vitro or in vivo (Nuttinck et al., 2004). In this study, FSH receptor mRNA was present in relatively large amounts in cumulus cells at the initiation of oocyte maturation. In vivo, FSH receptor mRNA decreased following the gonadotropin surge (Nuttinck et al., 2004). In vitro this decrease occurred after 24 h of maturation when COCs were matured in a complete maturation medium containing TCM-199, fetal calf serum, estradiol, LH and FSH. Interestingly, if COCs were matured in TCM-199 alone, inclusion of EGF in the medium also resulted in significant decrease in FSH receptor mRNA after 24 h of culture (Nuttinck et al., 2004).
To reconcile discrepancies in the long-perceived role of LH in inducing oocyte maturation in vivo with the lack of LH receptor presence on COCs before and during ooycte maturation both in vivo or in vitro, it has been suggested that LH binding to mural granulosa cells leads to the production of EGF-like growth factors including amphiregulin, epiregulin and betacellulin (Park et al., 2004). These growth factors can act as paracrine mediators and stimulate maturation of follicle-enclosed rodent oocytes (Park et al., 2004; Ashkenazi et al., 2005) and isolated COCs (Park et al., 2004). The observation that any of a number of ligands, including LH, FSH, EGF and EGF-like growth factors, when administered to individual follicle-enclosed oocytes or to cultured COCs, can stimulate ovulation and the induction of oocyte maturation suggests that substantial redundancy exists in the mechanisms regulating these critical physiological events. These redundancies may be important in contributing to the production of an oocyte that exhibits full developmental competence. Conversely, it is possible that activation of only one or a subset of these mechanisms governing ovulation and meiotic resumption may result in production of a compromised ovum in which morphological maturation appears normal but developmental competence is limited.
1.4. Mechanisms of gonadotropin-induced oocyte maturation in vitro
In cattle, COCs from follicles ranging from 2 to 8 mm are commonly used for studies of in vitro oocyte maturation. COCs from follicles of this size range do not contain receptors for LH (Nuttinck et al., 2004) and, thus, media used for studies of the mechanisms that underlie gonadotropin-induced oocyte maturation frequently contain FSH, but not LH. In mice, COCs used for studies of in vitro maturation are typically obtained from PMSG-treated prepubertal mice and are held in dictyate arrest during culture using phosphodiesterase inhibitors such as hypoxanthine (Downs et al., 1988). In both cattle and mice, the addition of FSH during in vitro maturation provides a positive stimulus that results in the induction of maturation (Eppig and Downs, 1987; Downs et al., 1988; Farin and Yang, 1994; Rodriguez et al., 2002; Rodriguez and Farin, 2004). However, in both cattle and mice, this induction of maturation by FSH occurs only after an initial period of maturation inhibition (Eppig et al., 1983; Schultz et al., 1983; Farin and Yang, 1994). In cattle, this initial inhibitory period lasts approximately 6 h (Farin and Yang, 1994; Rodriguez and Farin, 2004). In both cattle and mice, FSH induces oocyte maturation by binding to its receptors on the cumulus cells, elevating cumulus cell cAMP and activating both Type I and Type II protein kinase (PK) A. Activation of Type I PKA maintains oocytes in dictyate arrest (Downs and Hunzicker-Dunn, 1995; Rodriguez et al., 2002) whereas activation of Type II PKA stimulates a downstream transcriptional event that results in the induction of germinal vesicle breakdown (Rodriguez et al., 2002; Rodriguez and Farin, 2004). The role of gap junctions within the cumulus-oocyte complex is critical in communicating signals regulating not only the resumption of meiosis (Fagbohun and Downs, 1991; Downs, 2001) but also the acquisition of oocyte developmental competence (Nagai et al., 1993; Atef et al., 2005). Interestingly, the developmental competence of bovine oocytes matured in vitro was significantly increased when COCs were treated with FSH for only the first 6 h of culture compared to those treated with FSH throughout the entire 24 h period of maturation (Atef et al., 2005). Gap junctional communication was required for this effect (Atef et al., 2005). Thus, the initial period of FSH-mediated arrest of oocyte maturation that is associated with activation of cumulus cell Type I PKA may play an important role in allowing a period of delay needed for communication of other factors or activation of additional intracellular pathways important to optimize the developmental competence of in vitro-matured oocytes.
Downstream of cAMP-PKA isoform activation in cumulus cells, the stimulation of mitogen-activated protein kinase (MAPK) is a requirement for FSH-mediated induction of oocyte maturation in both cattle (Fissore et al., 1996) and mice (Su et al., 2002). MAPK activation appears to require the permissive effect of the oocyte itself through either the action of GDF-9 or other paracrine factors secreted by the oocyte (Su et al., 2003). Activation of the cumulus cell MAPK pathway also appears to be necessary for LH-mediated resumption of meiosis in follicle-enclosed oocytes (Su et al., 2003), suggesting that the mechanisms of LH-induced oocyte maturation in follicle-enclosed oocytes and FSH-induced meiotic resumption in cultured COCs may either directly overlap or utilize redundant pathways. It has not been determined if activation of the MAPK pathway in cultured COCs requires transcription to stimulate meiotic resumption or whether activation of MAPK occurs downstream of the transcriptional event that is required for FSH-mediated Type II PKA activation to stimulate the resumption of meiosis in cultured COCs.
In addition to FSH, EGF has been shown to induce oocyte maturation and cumulus cell expansion as well as improve oocyte developmental competence in cattle (Harper and Brackett, 1993; Lonergan et al., 1996; Rieger et al., 1998) and mice (De La Fuente et al., 1999). Similarly, treatment with the EGF-like growth factors amphiregulin, epiregulin and betacellulin also induced oocyte maturation in cultured murine COCs (Park et al., 2004) and in follicle-enclosed rat oocytes (Ashkenazi et al., 2005), demonstrating a potential role for these EGF-like growth factors as intermediates for LH-mediated induction of oocyte maturation (Conti et al., 2006). The relationship between the intracellular mechanisms promoting oocyte maturation in vitro used by FSH compared with that used by EGF or EGF-like growth factors is unclear. For example, both FSH and EGF activate MAPK pathways downstream of receptor binding (Su et al., 2002; Conti et al., 2006) but whether these pathways are duplicative or parallel has not been determined. It has been demonstrated that epiregulin mRNA expression is rapidly induced in cultured rat granulosa cells by FSH and in granulosa cells of preantral follicles exposed to PMSG (Sekiguchi et al., 2002). It remains unclear whether treatment with recombinant FSH can induce synthesis of amphiregulin, epiregulin and betacellulin by mural granulosa cells of preovulatory follicles in vivo or by cumulus cells in vitro, thus mimicking the intrafollicular actions of LH on oocyte maturation. It also remains unclear if EGF or EGF-like growth factors utilize a transcriptional component in their mechanism of action. If so, this would mimic the mechanism of FSH when inducing maturation of cultured COCs. To address some of these questions, we examined the effect of transcriptional inhibition on the ability of EGF, in the presence or absence of FSH, to stimulate oocyte maturation in both murine and feline cumulus oocyte complexes.
2. Materials and methods
2.1. Reagents and media
The transcriptional inhibitor, 5,6-dicholoro-1-β-D-ribofuranosylbenimidazole (DRB), as well as all hormonal preparations including equine pituitary FSH and murine EGF, were purchased from Sigma Chemical Co. (St. Louis. MO). Unless otherwise noted, all chemicals for preparation of media were from Sigma Chemical Co. and were of tissue culture grade. Murine oocyte cultures were conducted in Waymouth medium supplemented with 5% fetal bovine serum, 0.23 mM pyruvate, 7 mM taurine, 50 mg/L streptomyocin sulfate and 75 mg/L penicillin G. The medium used for handling feline oocytes was HEPES-buffered (pH=7.35) feline optimized culture medium (FOCM) containing 3.0 mM glucose and 0.4% (w/v) BSA (Swanson et al., 2004; Herrick et al., 2005). Feline oocytes were matured in bicarbonate-buffered FOCM containing 6.0 mM glucose, 5 μg/ml insulin, 5 μg/ml transferrin, 5 ng/ml selenium, 0.1 mM cysteamine, 0.6 mM cysteine, 0.5x MEM EAA, 1.0x MEM vitamins and 0.4% (w/v) BSA (Herrick et al., 2005; J. Herrick, unpublished). For all experiments, DRB was used at a concentration of 120 μM, a dose that maintains both murine and bovine oocytes in meiotic arrest (Farin and Yang, 1994; Martus and Farin, 1994; Wolf and Farin, 1996; Rodriguez et al., 2002).
2.2. Murine oocyte recovery and culture
All procedures involving animals were carried out in accordance with NIH guidelines for the care and use of laboratory animals under the approval of the National Institutes of Environmental Health Science Institutional Animal Care and Use Committee. Prepubertal CD-1 mice (21 days old) were maintained on a 12L:12D light cycle with access to food and water ad libitum. A minimum of 5 mice were used in each replicate and were given an intraperitoneal injection of 5 IU PMSG and killed by CO2 asphyxiation 48 h later. Ovaries were recovered and oocytes distributed to treatments according to previously published procedures (Rodriguez et al., 2002). The hormone and inhibitor concentrations used in this study (FSH, 0.2 U/ml; EGF, 10 ng/ml; DRB, 120 μM) were based on previously published doses (Downs, 1989; Rodriguez et al., 2002). Cultures were conducted for 3.5 h at 37°C in an atmosphere of 5% CO2 in air with 100% humidity (Rodriguez et al., 2002). At the termination of culture, fully grown oocytes were denuded of their cumulus cells by manual pipetting and visually assessed for the resumption of meiosis at a magnification of 60X using a stereomicroscope. Oocytes not demonstrating an intact GV were classified as having undergone GVBD.
2.3. Feline oocyte recovery and culture
Feline ovaries were obtained from a local veterinary clinic during either the breeding season (February–May) or early in the non-breeding season (June–July). Ovaries were maintained at room temperature in saline and transported to the laboratory within 4 h of recovery. Fully grown cumulus oocyte complexes (COC) were released from antral follicles into handling medium, collected and washed three times before being randomly distributed to treatment groups in FOCM. When included in the maturation medium, EGF was used at a concentration of 50ng/ml, FSH at 0.2 U per ml and DRB at 120 μM. All cultures were conducted at 38.5° C in an atmosphere of 90%N2:5%O2:5% CO2 with 100% humidity with a 50% volume of fresh treatment medium replaced at 4 h intervals throughout culture. After 20 h of maturation, oocytes were denuded of their cumulus cells by gentle vortexing, fixed in acidified ethanol, stained with aceto-orcein and assessed for stage of meiosis (Farin and Yang, 1994).
2.4. Statistical analysis
All percentage data were arcsin transformed and analyzed by analysis of variance using General Linear Models procedures (SAS, 1999). The statistical models used to analyze the effect of hormone treatment and transcriptional inhibitor on stages of murine oocyte meiosis (Figure 1) included the main effects of EGF, FSH and DRB and all appropriate interactions. The statistical model used to assess the effect of treatment and season on stages of meiosis in feline COCs cultured in the presence of EGF (Figure 2) included the main effects of FSH, DRB, season of collection and the interactions of FSH by DRB and FSH by DRB by season. The statistical model used to analyze feline stages of meiosis cultured in the absence of EGF during the breeding season (Figure 3) included the main effects of FSH, DRB and the interactions of FSH by DRB. The statistical model used to analyze feline stages of meiosis cultured in the presence or absence of EGF during the breeding season (Figure 4) included the main effects of hormone (EGF, FSH, Control), inhibitor (DRB, Control) and the interactions of hormone by inhibitor. In all cases, when a significant F-statistic was identified, means were separated using Duncan’s Multiple Range Test (SAS, 1999). All data are reported as least squares means ± standard error.
Fig. 1.
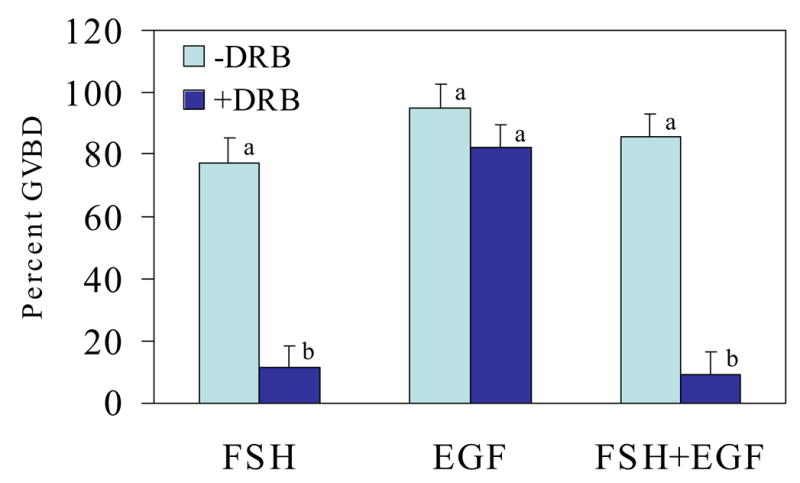
Effect of treatment with the transcriptional inhibitor, DRB, on FSH- and EGF-induced oocyte maturation in the mouse. Data represent least squares (ls) means ± SEM based on n=4 experimental replicates with 20 to 52 COCs per treatment per replicate; abP<0.05
Fig. 2.
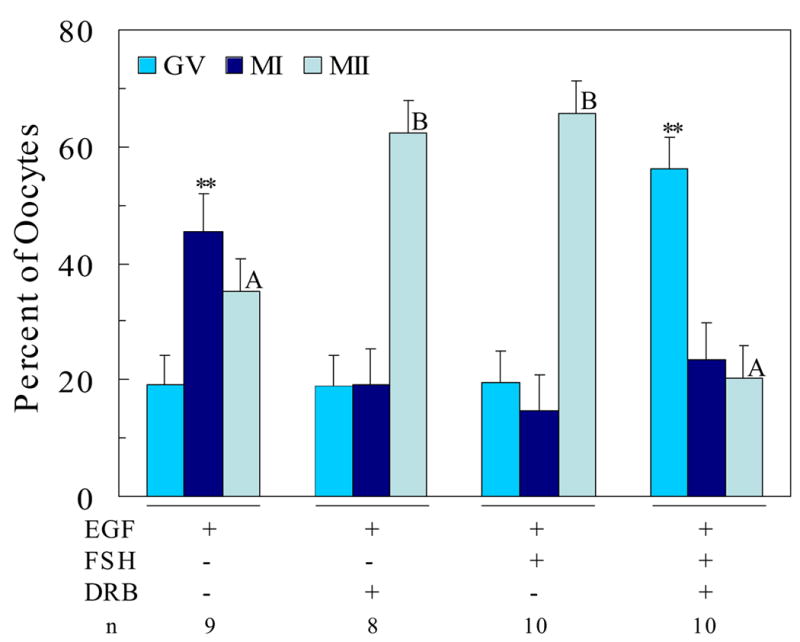
Effect of transcriptional inhibition on EGF- and FSH-induced oocyte maturation in domestic cats. Data represent lsmeans ± SEM based on n=8–10 replicate observations with 20 to 35 COCs per treatment per replicate; **P<0.05 between treatments within either GV or MI stage; A,BP<0.05 between treatments within MII stage
Fig. 3.
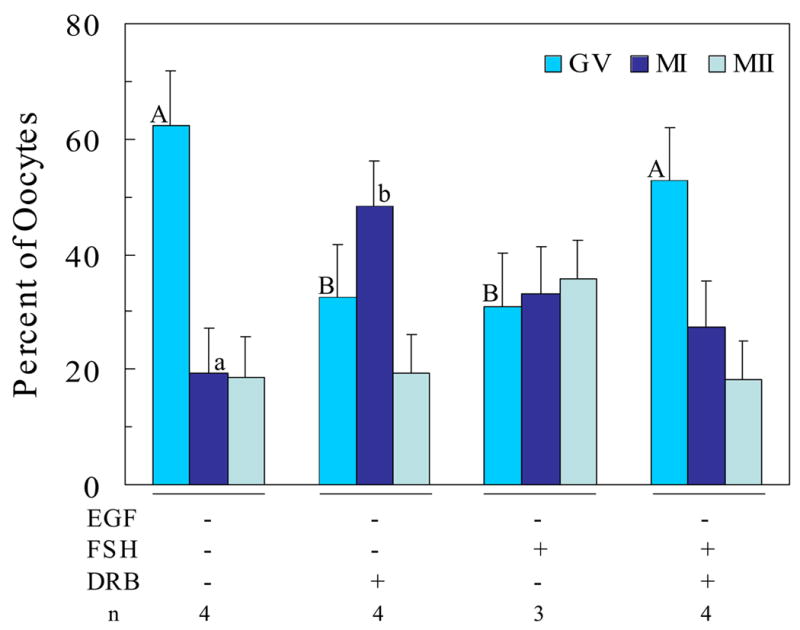
Effect of transcriptional inhibition on FSH-induced oocyte maturation in domestic cats during the breeding season; Data represent lsmeans ± SEM based on n = 3 or 4 replicate observations with 20 to 35 COCs per treatment per replicate; A,BP=0.01 for GV stage; a,bP=0.06 for MI stage
Fig. 4.
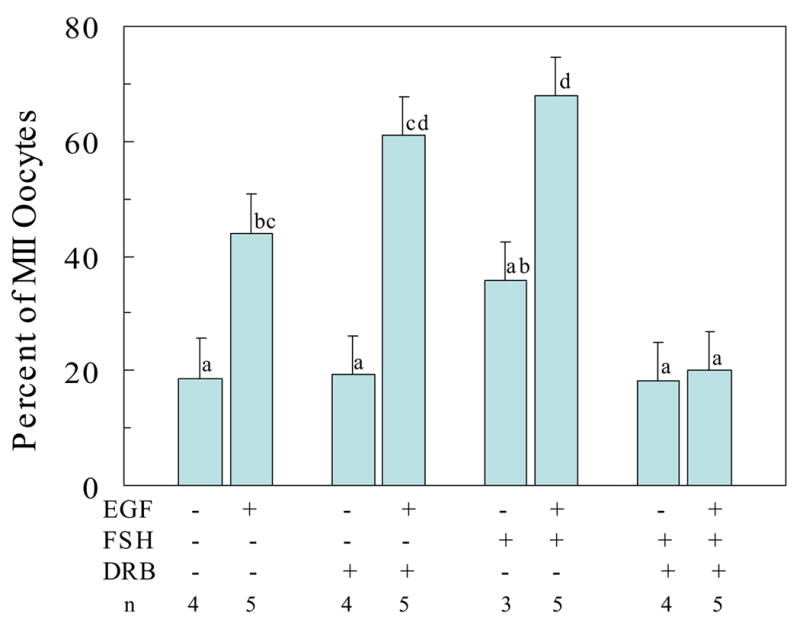
Effect of transcriptional inhibition on the effectiveness of EGF and FSH to stimulate progression of feline oocytes to the MII stage during the breeding season; Data represent lsmeans ± SEM based on n =3 to 5 replicate observations with 20 to 35 COCs per treatment per replicate; abcdP<0.05
3. Results and discussion
3.1. Effect of transcriptional inhibition on EGF- and FSH-mediated murine oocyte maturation
After 3.5 h of culture in the presence of either FSH, EGF or their combination, greater than 80% of oocytes underwent GVBD (Figure 1). These observations are consistent with reports in the literature (De La Fuente et al., 1999). Also consistent with earlier reports (Rodriguez et al., 2002), maturation of murine oocytes in the presence of FSH was blocked when the transcriptional inhibitor, DRB, was included in the medium. Thus, nascent gene transcription is required to mediate the action of FSH to induce GVBD. When DRB was added to EGF-containing maturation medium, however, no effect of the transcriptional inhibitor was demonstrated. This suggests that new gene transcription is not required for the mechanism used by EGF to stimulate GVBD. In maturation medium containing both EGF and FSH, treatment with DRB blocked about 95% of oocytes from maturing (Figure 1). Thus, when both FSH and EGF are present, the FSH pathway that requires gene transcription appears to over-ride any effect of the transcriptionally independent mechanism used by EGF to stimulate GVBD in murine oocytes.
3.2. Effect of transcriptional inhibition on EGF- and FSH-mediated feline oocyte maturation
There was no effect of season of oocyte harvest on the maturation of feline COCs cultured in the presence of EGF nor were there any significant interactions of season with the FSH or inhibitor treatments, therefore the data for the effects of hormone and DRB treatments were combined across season and are presented in Figure 2.
When cultured for 20 h in the presence of EGF alone, few (19 ± 5%) oocytes remained at the GV stage with approximately equal proportions of oocytes reaching the metaphase I (MI, 46 ± 6%) or metaphase II (MII, 35 ± 6%) stages of maturation (Figure 2). When the transcriptional inhibitor DRB was added to medium containing EGF the majority of oocytes (62 ± 6%; P<0.05) progressed to MII. This was associated with a significant reduction in the proportion of oocytes remaining at MI after 20 h of maturation (Figure 2). These observations suggest that exposure to EGF alone may stimulate gene transcription in some COCs and that this action can maintain these oocytes in MI of meiosis. Inhibition of EGF-associated transcriptional activity by exposure to DRB permits these oocytes to resume meiosis, resulting in significantly more oocytes completing meiotic maturation. An inhibitory effect of EGF on the production of inhibin by granulosa cells in the rat has been reported (Zhang et al., 1987); thus, it is not inconsistent to suggest that EGF could also exhibit an inhibitory effect on the meiotic progression of cultured oocytes.
When FSH was included in the maturation medium in the presence of EGF, a majority of oocytes (66 ± 5%) reached MII after 20 h of culture (Figure 2). However, when DRB was added to medium containing both FSH and EGF, only 20% (P<0.05) of oocytes reached MII with most oocytes (56 ± 5%, P<0.05) arrested at the GV stage. These data are consistent with observations in other mammalian species that oocyte maturation in the presence of FSH requires gene transcription (Osborn and Moor, 1983; Hunter and Moor, 1987; Kastrop et al., 1991; Farin and Yang, 1994; Rodriguez et al., 2002). Furthermore, these data, like those for cultured murine COCs, demonstrate that when both EGF and FSH are present, the FSH pathway requiring gene transcription over-rides any effect EGF may have in stimulating oocytes to resume meiosis.
Interestingly, feline oocytes cultured in semi-defined medium in the absence of any hormones remained arrested at the GV stage (Figure 3). Thus, cats may provide a natural GV-arrest model useful for the study of mechanisms underlying mammalian oocyte maturation. In contrast to the effectiveness of FSH to stimulate oocyte maturation in the presence of EGF (Figure 2), FSH was only marginally effective in promoting the maturation of oocytes when EGF was absent from the culture medium (Figure 3). This observation suggests that a background of EGF in maturation medium may be required to optimize the effectiveness of FSH to promote meiosis in feline oocytes in vitro. This is particularly intriguing in that these observations suggest the possibility of linkage between the roles of LH, EGF and FSH to promote oocyte maturation in vivo and in vitro. The observation that FSH in the absence of added EGF resulted in the maturation of cultured murine oocytes (Figure 1) can be explained by the potential presence of EGF or EGF-like growth factors in the serum that was used as a medium supplement in these experiments.
Inclusion of DRB in the absence of EGF promoted the progression of meiosis to the MI stage (Figure 3). This observation supports the suggestion that nascent gene transcription within feline COCs may be responsible for holding at least some oocytes in GV arrest. This effect of DRB is reminiscent of the effect of EGF alone on oocyte maturation (Figure 2). A comparison of the effects of EGF on the ability of FSH and DRB to influence the complete in vitro maturation of feline oocytes harvested during the breeding season is presented in Figure 4. These data demonstrate that EGF alone can promote progression to MII and that EGF is required for FSH-induced maturation to the MII stage. The pathway used for progression of maturation in the combined presence of FSH and EGF, however, requires nascent gene transcription since treatment with DRB prevented oocytes from progressing to MII.
The pathway utilized by EGF to mediate meiotic resumption involves binding to one or more of the EGF receptor subfamily members with subsequent tyrosine kinase activation and downstream activation of MAPK (Conti et al., 2006). In addition, activation of the MAPK pathway is required for FSH-mediated resumption of meiosis in both cattle and mice (Fissore et al., 1996; Su et al., 2002). The pathway used by FSH to stimulate GVBD appears to involve cAMP-mediated activation of PKA that, in turn, leads to activation of MAPK (Su et al., 2002). Because activation of both Type II PKA and MAPK can each stimulate transcription as a downstream effect (Rodriguez et al., 2002; Conti et al., 2006), it would be likely that these activation events precede the transcriptional step required in the mechanism used by FSH to mediate meiotic resumption.
Although it is clear that both EGF and FSH utilize MAPK activation in their pathways leading to meiotic resumption, it appears that these two pathways must not be directly linked within cumulus cells because FSH-induced meiotic resumption can be inhibited by DRB whereas EGF-stimulated meiotic resumption cannot. The possibility that these two pathways overlap downstream of the transcriptional event required for FSH-induced oocyte maturation is also not supported by the current observations in either feline or murine oocytes. If this were the case, EGF would be expected to promote oocyte maturation despite the presence of FSH and DRB in the maturation medium. Thus, although both FSH and EGF may each rely on MAPK activation in the pathways they utilize to stimulate meiotic resumption, these pathways do not appear to be overlapping or interconnected. Furthermore, when both hormones are present in the medium, the FSH-mediated pathway appears to take precedence over the pathway utilized by EGF to stimulate meiotic maturation.
Although it appears that for regulation of GVBD the FSH-mediated pathway takes precedence, this does not discount the importance of the effects of EGF on oocyte maturation. On the contrary, data presented here demonstrate that EGF is a necessary prerequisite for FSH to induce feline oocyte maturation under these semi-defined culture conditions. Furthermore, based on data in cattle, mice and cats, inclusion of EGF in maturation medium improves oocyte developmental competence over that of FSH alone (Harper and Brackett, 1993; Lonergan et al., 1996; Rieger et al., 1998; De La Fuente et al., 1999; Merlo et al., 2005). Thus, it is likely that FSH and EGF work in concert to regulate oocyte maturation and optimize oocyte developmental competence in vitro. It is tempting to speculate that an analogous mechanism could exist in vivo with LH mediating the induction of EGF-like growth factors by granulosa cells that, in turn, could facilitate the action of FSH on cumulus cells to promote oocyte maturation and oocyte developmental competence. Evaluation of this proposed hypothesis awaits further investigation.
Acknowledgments
Research supported by USDA Grant 2002-35205-12810, NIH Grant 1 R03 HD043875-01, the Morris Animal Foundation and the North Carolina Agricultural Research Service. The authors would like to acknowledge the assistance of Dr. K.S. Korach of the National Institute of Environmental Health Sciences for rodent laboratory resources and Dr. William F. Swanson for advice concerning the feline culture system. The authors also acknowledge Rachel Croughwell, Lauren Finley and Nicola Wrench for technical assistance and Safe Haven for Cats Veterinary Clinic, Raleigh NC for providing feline ovaries used for this work.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Ashkenazi H, Cao X, Motola S, Popliker M, Conti M, Tsafriri A. Epidermal growth factor family members: endogenous mediators of the ovulatory response. Endocrinology. 2005;146:77–84. doi: 10.1210/en.2004-0588. [DOI] [PubMed] [Google Scholar]
- Atef A, Francois P, Christian V, Marc-Andre S. The potential role of gap junction communication between cumulus cells and bovine oocytes during in vitro maturation. Mol Reprod Dev. 2005;71:358–367. doi: 10.1002/mrd.20281. [DOI] [PubMed] [Google Scholar]
- Blondin P, Bousquet D, Twagiramungu H, Barnes F, Sirard MA. Manipulation of follicular development to produce developmentally competent bovine oocytes. Biol Reprod. 2002;66:38–43. doi: 10.1095/biolreprod66.1.38. [DOI] [PubMed] [Google Scholar]
- Bordignon V, Norin N, Durocher J, Bousquet D, Smith LC. GnRH improves the recovery rate and the in vitro developmental competence of oocytes obtained by transvaginal follicular aspiration from superstimulated heifers. Theriogenology. 1997;48:291–298. doi: 10.1016/s0093-691x(97)84076-9. [DOI] [PubMed] [Google Scholar]
- Brevini-Gandolfi TAL, Favetta LA, Mauri L, Luciano AM, Cillo F, Gandolfi F. Changes in poly(a) tail length of maternal transcripts during in vitro maturation of bovine oocytes and their relation with developmental competence. Mol Reprod Dev. 1999;52:427–433. doi: 10.1002/(SICI)1098-2795(199904)52:4<427::AID-MRD12>3.0.CO;2-G. [DOI] [PubMed] [Google Scholar]
- Brower PT, Schultz RM. Intercellular communication between granulosa cells and mouse oocytes: existence and possible nutritional role during oocyte growth. Dev Biol. 1982;90:144–153. doi: 10.1016/0012-1606(82)90219-6. [DOI] [PubMed] [Google Scholar]
- Buccione R, Schroeder AC, Eppig JJ. Interactions between somatic cells and germ cells throughout mammalian oogenesis. Biol Reprod. 1990;43:543–547. doi: 10.1095/biolreprod43.4.543. [DOI] [PubMed] [Google Scholar]
- Carabatsos MJ, Sellitto C, Goodenough DA, Albertini DF. Oocyte-granulosa cell heterologous gap junctions are required for the coordination of nuclear and cytoplasmic meiotic competence. Dev Biol. 2000;226:167–179. doi: 10.1006/dbio.2000.9863. [DOI] [PubMed] [Google Scholar]
- Child TJ, Abdul-Jalil AK, Gulekli B, Tan SL. In vitro maturation and fertilization of oocytes from unstimulated normal ovaries, polycystic ovaries, and women with polycystic ovary syndrome. Fertil Steril. 2001;76:936–942. doi: 10.1016/s0015-0282(01)02853-9. [DOI] [PubMed] [Google Scholar]
- Conti M, Hsieh M, Park JY, Su YQ. Role of the epidermal growth factor network in ovarian follicles. Mol Endocrinol. 2006;20:715–723. doi: 10.1210/me.2005-0185. [DOI] [PubMed] [Google Scholar]
- De La Fuente R, O'Brien MJ, Eppig JJ. Epidermal growth factor enhances preimplantation developmental competence of maturing mouse oocytes. Hum Reprod. 1999;14:3060–3068. doi: 10.1093/humrep/14.12.3060. [DOI] [PubMed] [Google Scholar]
- De La Fuente R, Eppig JJ. Transcriptional activity of the mouse oocyte genome: Companion granulosa cells modulate transcription and chromatin remodeling. DevBiol. 2001;229:224–236. doi: 10.1006/dbio.2000.9947. [DOI] [PubMed] [Google Scholar]
- Dieleman SJ, Hendriksen PJ, Viuff D, Thomsen PD, Hyttel P, Knijn HM, Wrenzycki C, Kruip TA, Niemann H, Gadella BM, Bevers MM, Vos PL. Effects of in vivo prematuration and in vivo final maturation on developmental capacity and quality of pre-implantation embryos. Theriogenology. 2002;57:5–20. doi: 10.1016/s0093-691x(01)00655-0. [DOI] [PubMed] [Google Scholar]
- Downs SM. Specificity of epidermal growth factor action on maturation of the murine oocyte and cumulus oophorus in vitro. Biol Reprod. 1989;41:371–379. doi: 10.1095/biolreprod41.2.371. [DOI] [PubMed] [Google Scholar]
- Downs SM. A gap-junction-mediated signal, rather than an external paracrine factor, predominates during meiotic induction in isolated mouse oocytes. Zygote. 2001;9:71–82. doi: 10.1017/s0967199401001071. [DOI] [PubMed] [Google Scholar]
- Downs SM, Daniel SA, Eppig JJ. Induction of maturation in cumulus cell-enclosed mouse oocytes by follicle-stimulating hormone and epidermal growth factor: Evidence for a positive stimulus of somatic cell origin. J Exp Zool. 1988;245:86–96. doi: 10.1002/jez.1402450113. [DOI] [PubMed] [Google Scholar]
- Downs SM, Hunzicker-Dunn M. Differential regulation of oocyte maturation and cumulus expansion in the mouse oocyte-cumulus cell complex by site-selective analogs of cyclic adenosine monophosphate. Dev Biol. 1995;172:72–85. doi: 10.1006/dbio.1995.0006. [DOI] [PubMed] [Google Scholar]
- Duranthon V, Renard JP. The developmental competence of mammalian oocytes: a convenient but biologically fuzzy concept. Theriogenology. 2001;55:1277–1289. doi: 10.1016/s0093-691x(01)00482-4. [DOI] [PubMed] [Google Scholar]
- Durlinger AL, Gruijters MJ, Kramer P, Karels B, Ingraham HA, Nachtigal MW, Uilenbroek JT, Grootegoed JA, Themmen AP. Anti-mullerian hormone inhibits initiation of primordial follicle growth in the mouse ovary. Endocrinology. 2002;143:1076–1084. doi: 10.1210/endo.143.3.8691. [DOI] [PubMed] [Google Scholar]
- Eppig JJ, Freter RR, Ward-Bailey PF, Schultz RM. Inhibition of oocyte maturation in the mouse: participation of camp, steroid hormones, and a putative maturation-inhibitory factor. Dev Biol. 1983;100:39–49. doi: 10.1016/0012-1606(83)90198-7. [DOI] [PubMed] [Google Scholar]
- Eppig JJ, Downs SM. The effect of hypoxanthine on mouse oocyte growth and development in vitro: maintenance of meiotic arrest and gonadotropin-induced oocyte maturation. Dev Biol. 1987;119:313–321. doi: 10.1016/0012-1606(87)90037-6. [DOI] [PubMed] [Google Scholar]
- Eppig JJ, Schultz RM, O'Brien MJ, Chesnel F. Relationship between the developmental programs controlling nuclear and cytoplasmic maturation of mouse oocytes. Dev Biol. 1994;164:1–9. doi: 10.1006/dbio.1994.1175. [DOI] [PubMed] [Google Scholar]
- Eppig JJ, Wigglesworth K, Pendola F, Hirao Y. Murine oocytes suppress expression of luteinizing hormone receptor messenger ribonucleic acid by granulosa cells. Biol Reprod. 1997;56:976–984. doi: 10.1095/biolreprod56.4.976. [DOI] [PubMed] [Google Scholar]
- Eppig JJ, Pendola FL, Wigglesworth K. Mouse oocytes suppress cAMP-induced expression of LH receptor mRNA by granulosa cells in vitro. Mol Reprod Dev. 1998;49:327–332. doi: 10.1002/(SICI)1098-2795(199803)49:3<327::AID-MRD13>3.0.CO;2-T. [DOI] [PubMed] [Google Scholar]
- Eppig JJ, Wigglesworth K, Pendola FL. The mammalian oocyte orchestrates the rate of ovarian follicular development. Proc Natl Acad Sci USA. 2002;99:2890–2894. doi: 10.1073/pnas.052658699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fagbohun CF, Downs SM. Metabolic coupling and ligand-stimulated meiotic maturation in the mouse oocyte-cumulus cell complex. Biol Reprod. 1991;45:851–859. doi: 10.1095/biolreprod45.6.851. [DOI] [PubMed] [Google Scholar]
- Fair T, Hyttel P, Greve T. Bovine oocyte diameter in relation to maturational competence and transcriptional activity. Mol Reprod Dev. 1995;42:437–442. doi: 10.1002/mrd.1080420410. [DOI] [PubMed] [Google Scholar]
- Farin CE, Yang L. Inhibition of germinal vesicle breakdown in bovine oocytes by 5,6-dichloro-1-β-D-ribofuranosylbenzimidazole (DRB) Mol Reprod Dev. 1994;37:284–292. doi: 10.1002/mrd.1080370307. [DOI] [PubMed] [Google Scholar]
- Fissore RA, He CL, Vande Woude GF. Potential role of mitogen-activated protein kinase during meiosis resumption in bovine oocytes. Biol Reprod. 1996;55:1261–1270. doi: 10.1095/biolreprod55.6.1261. [DOI] [PubMed] [Google Scholar]
- Galway AB, Lapolt PS, Tsafriri A, Dargan CM, Boime I, Hsueh AJ. Recombinant follicle-stimulating hormone induces ovulation and tissue plasminogen activator expression in hypophysectomized rats. Endocrinology. 1990;127:3023–3028. doi: 10.1210/endo-127-6-3023. [DOI] [PubMed] [Google Scholar]
- Gandolfi TA, Gandolfi F. The maternal legacy to the embryo: Cytoplasmic components and their effects on early development. Theriogenology. 2001;55:1255–1276. doi: 10.1016/s0093-691x(01)00481-2. [DOI] [PubMed] [Google Scholar]
- Goldman BD, Mahesh VB. A possible role of acute FSH-release in ovulation in the hamster, as demonstrated by utilization of antibodies to LH and FSH. Endocrinology. 1969;84:236–243. doi: 10.1210/endo-84-2-236. [DOI] [PubMed] [Google Scholar]
- Greenwald GS, Papkoff H. Induction of ovulation in the hypophysectomized proestrous hamster by purified FSH or LH. Proc Soc Exp Biol Med. 1980;165:391–393. doi: 10.3181/00379727-165-40992. [DOI] [PubMed] [Google Scholar]
- Greep RO, Van Dyke HB, Chow BF. Gonadotropins in the swine pituitary. I Various biological effects of purified FSH and pure ICSH Endocrinology. 1942;30:635–648. [Google Scholar]
- Harper KM, Brackett BG. Bovine blastocyst development after in vitro maturation in a defined medium with epidermal growth factor and low concentrations of gonadotropins. Biol Reprod. 1993;48:409–416. doi: 10.1095/biolreprod48.2.409. [DOI] [PubMed] [Google Scholar]
- Harrington FE, Elton RL. Induction of ovulation in adult rats with follicle stimulating hormone. Proc Soc Exp Biol Med. 1969;132:841–844. doi: 10.3181/00379727-132-34319. [DOI] [PubMed] [Google Scholar]
- Haughian JM, Ginther OJ, Kot K, Wiltbank MC. Relationships between fsh patterns and follicular dynamics and the temporal associations among hormones in natural and gnrh-induced gonadotropin surges in heifers. Reproduction. 2004;127:23–33. doi: 10.1530/rep.1.00030. [DOI] [PubMed] [Google Scholar]
- Herrick JR, Bond JB, Magarey GM, Bateman HL, Krisher RL, Dunford SA, Swanson WF. Effects of glucose, L-lactate, and pyruvate in culture media on embryonic development and metabolism of IVF-derived feline embryos relative to embryos grown in vivo. Biol Reprod Special Issue. 2005;160:abst W357. doi: 10.1095/biolreprod.106.058065. [DOI] [PubMed] [Google Scholar]
- Hunter AG, Moor RM. Stage-dependent effects of inhibiting ribonucleic acids and protein synthesis on meiotic maturation of bovine oocytes in vitro. J Dairy Sci. 1987;70:1646–1651. doi: 10.3168/jds.S0022-0302(87)80192-3. [DOI] [PubMed] [Google Scholar]
- Hyttel P, Fair T, Callesen H, Greve T. Oocyte growth, capacitation and final maturation in cattle. Theriogenology. 1997;47:23–32. [Google Scholar]
- Izadyar F, Zeinstra E, Bevers MM. Follicle-stimulating hormone and growth hormone act differently on nuclear maturation while both enhance developmental competence of in vitro matured bovine oocytes. Mol Reprod Dev. 1998;51:339–345. doi: 10.1002/(SICI)1098-2795(199811)51:3<339::AID-MRD14>3.0.CO;2-Y. [DOI] [PubMed] [Google Scholar]
- Jones EE, Nalbandov AV. Effects of intrafollicular injection of gonadotrophins on ovulation or luteinization of ovarian follicle. Biol Reprod. 1972;7:87–93. doi: 10.1093/biolreprod/7.1.87. [DOI] [PubMed] [Google Scholar]
- Kastrop PM, Hulshof SC, Bevers MM, Destree OH, Kruip TA. The effects of alpha-amanitin and cycloheximide on nuclear progression, protein synthesis, and phosphorylation during bovine oocyte maturation in vitro. Mol Reprod Dev. 1991;28:249–254. doi: 10.1002/mrd.1080280306. [DOI] [PubMed] [Google Scholar]
- Krisher RL, Bavister BD. Responses of oocytes and embryos to the culture environment. Theriogenology. 1998;49:103–114. doi: 10.1016/s0093-691x(97)00405-6. [DOI] [PubMed] [Google Scholar]
- Kruip TA, Bevers MM, Kemp B. Environment of oocyte and embryo determines health of IVP offspring. Theriogenology. 2000;53:611–618. doi: 10.1016/s0093-691x(99)00261-7. [DOI] [PubMed] [Google Scholar]
- Lipner H, Hirsch MA, Moudgal NR, MacDonald GJ, Ying SY, Greep RO. Ovulating-inducing activity of FSH in the rat. Endocrinology. 1974;94:1351–1358. doi: 10.1210/endo-94-5-1351. [DOI] [PubMed] [Google Scholar]
- Lonergan P, Monaghan P, Rizos D, Boland MP, Gordon I. Effect of follicle size on bovine oocyte quality and developmental competence following maturation, fertilization, and culture in vitro. Mol Reprod Dev. 1994;37:48–53. doi: 10.1002/mrd.1080370107. [DOI] [PubMed] [Google Scholar]
- Lonergan P, Carolan C, Van Langendonckt A, Donnay I, Khatir H, Mermillod P. Role of epidermal growth factor in bovine oocyte maturation and preimplantation embryo development in vitro. Biol Reprod. 1996;54:1420–1429. doi: 10.1095/biolreprod54.6.1420. [DOI] [PubMed] [Google Scholar]
- Martus NS, Farin CE. Effectiveness of DRB for inhibiting GVBD in bovine oocytes. Theriogenology. 1994;42:1295–1302. [Google Scholar]
- Matzuk MM, Burns KH, Viveiros MM, Eppig JJ. Intercellular communication in the mammalian ovary: oocytes carry the conversation. Science. 2002;296:2178–2180. doi: 10.1126/science.1071965. [DOI] [PubMed] [Google Scholar]
- McLaren A. The developmental history of female germ cells in mammals. Oxford Rev Reprod Biol. 1988;10:162–179. [PubMed] [Google Scholar]
- Merlo B, Iacono E, Zambelli D, Prati F, Belluzzi S. Effect of EGF on in vitro maturation of domestic cat oocytes. Theriogenology. 2005;63:2032–2039. doi: 10.1016/j.theriogenology.2004.09.045. [DOI] [PubMed] [Google Scholar]
- Murdoch WJ, Dailey RA, Inskeep EK. Preovulatory changes prostaglandins E2 and F2 alpha in ovine follicles. J Anim Sci. 1981;53:192–205. doi: 10.2527/jas1981.531192x. [DOI] [PubMed] [Google Scholar]
- Nagai T, Ding J, Moor RM. Effect of follicle cells and steroidogenesis on maturation and fertilization in vitro of pig oocytes. J Exp Zool. 1993;266:146–151. doi: 10.1002/jez.1402660208. [DOI] [PubMed] [Google Scholar]
- Nuti LC, McShan WH, Meyer RK. Effect of ovine FSH and LH on serum steroids and ovulation in hypophysectomized immature female rats. Endocrinology. 1974;95:682–689. doi: 10.1210/endo-95-3-682. [DOI] [PubMed] [Google Scholar]
- Nuttinck F, Charpigny G, Mermillod P, Loosfelt H, Meduri G, Freret S, Grimard B, Heyman Y. Expression of components of the insulin-like growth factor system and gonadotropin receptors in bovine cumulus-oocyte complexes during oocyte maturation. Domest Anim Endocrinol. 2004;27:179–195. doi: 10.1016/j.domaniend.2004.03.003. [DOI] [PubMed] [Google Scholar]
- Osborn JC, Moor RM. Time-dependent effects of alpha-amanitin on nuclear maturation and protein synthesis in mammalian oocytes. J Embryol Exp Morphol. 1983;73:317–338. [PubMed] [Google Scholar]
- Pakarainen T, Zhang FP, Nurmi L, Poutanen M, Huhtaniemi I. Knockout of luteinizing hormone receptor abolishes the effects of follicle-stimulating hormone on preovulatory maturation and ovulation of mouse graafian follicles. Mol Endocrinol. 2005;19:2591–2602. doi: 10.1210/me.2005-0075. [DOI] [PubMed] [Google Scholar]
- Park JY, Su YQ, Ariga M, Law E, Jin SL, Conti M. EGF-like growth factors as mediators of LH action in the ovulatory follicle. Science. 2004;303:682–684. doi: 10.1126/science.1092463. [DOI] [PubMed] [Google Scholar]
- Parrott JA, Skinner MK. Kit-ligand/stem cell factor induces primordial follicle development and initiates folliculogenesis. Endocrinology. 1999;140:4262–4271. doi: 10.1210/endo.140.9.6994. [DOI] [PubMed] [Google Scholar]
- Pavlok A, Hahn LA, Niemann H. Fertilization and developmental competence of bovine oocytes derived from different categories of antral follicles. Mol Reprod Dev. 1992;31:63–67. doi: 10.1002/mrd.1080310111. [DOI] [PubMed] [Google Scholar]
- Picton HM. Oocyte maturation in vitro. Curr Opin Obstet Gynecol. 2002;14:295–302. doi: 10.1097/00001703-200206000-00009. [DOI] [PubMed] [Google Scholar]
- Richards JS. Maturation of ovarian follicles:Actions and interactions of pituitary and ovarian hormones on follicular cell differentiation. Physiol Rev. 1980;60:51–89. doi: 10.1152/physrev.1980.60.1.51. [DOI] [PubMed] [Google Scholar]
- Rieger D, Luciano AM, Modina S, Pocar P, Lauria A, Gandolfi F. The effects of epidermal growth factor and insulin-like growth factor I on the metabolic activity, nuclear maturation and subsequent development of cattle oocytes in vitro. J Reprod Fertil. 1998;112:123–130. doi: 10.1530/jrf.0.1120123. [DOI] [PubMed] [Google Scholar]
- Rizos D, Ward F, Duffy P, Boland MP, Lonergan P. Consequences of bovine oocyte maturation, fertilization or early embryo development in vitro versus in vivo: implications for blastocyst yield and blastocyst quality. Mol Reprod Dev. 2002;61:234–248. doi: 10.1002/mrd.1153. [DOI] [PubMed] [Google Scholar]
- Rodriguez KF, Petters RM, Crosier AE, Farin CE. Roles of gene transcription and PKA subtype activation in maturation of murine oocytes. Reproduction. 2002;123:799–806. doi: 10.1530/rep.0.1230799. [DOI] [PubMed] [Google Scholar]
- Rodriguez KF, Farin CE. Gene transcription and regulation of oocyte maturation. Reprod Fertil Dev. 2004;16:55–67. doi: 10.10371/RD03078. [DOI] [PubMed] [Google Scholar]
- SAS. The SAS system for windows, release 8.02. Cary, NC: Statistical Analysis System Institute, Inc; 1999. [Google Scholar]
- Schultz RM, Montgomery RR, Ward-Bailey PF, Eppig JJ. Regulation of oocyte maturation in the mouse: Possible roles of intercellular communication, cAMP, and testosterone. Dev Biol. 1983;95:294–304. doi: 10.1016/0012-1606(83)90030-1. [DOI] [PubMed] [Google Scholar]
- Sekiguchi T, Mizutani T, Yamada K, Yazawa T, Kawata H, Yoshino M, Kajitani T, Kameda T, Minegishi T, Miyamoto K. Transcriptional regulation of the epiregulin gene in the rat ovary. Endocrinology. 2002;143:4718–4729. doi: 10.1210/en.2002-220440. [DOI] [PubMed] [Google Scholar]
- Simon AM, Goodenough DA, Li E, Paul DL. Female infertility in mice lacking connexin 37. Nature. 1997;385:525–529. doi: 10.1038/385525a0. [DOI] [PubMed] [Google Scholar]
- Sirard MA. Resumption of meiosis: Mechanism involved in meiotic progression and its relation with developmental competence. Theriogenology. 2001;55:1241–1254. doi: 10.1016/s0093-691x(01)00480-0. [DOI] [PubMed] [Google Scholar]
- Sorensen RA, Wassarman PM. Relationship between growth and meiotic maturation of the mouse oocyte. Dev Biol. 1976;50:531–536. doi: 10.1016/0012-1606(76)90172-x. [DOI] [PubMed] [Google Scholar]
- Stojkovic M, Machado SA, Stojkovic P, Zakhartchenko V, Hutzler P, Goncalves PB, Wolf E. Mitochondrial distribution and adenosine triphosphate content of bovine oocytes before and after in vitro maturation: correlation with morphological criteria and developmental capacity after in vitro fertilization and culture. Biol Reprod. 2001;64:904–909. doi: 10.1095/biolreprod64.3.904. [DOI] [PubMed] [Google Scholar]
- Su YQ, Wigglesworth K, Pendola FL, O'Brien MJ, Eppig JJ. Mitogen-activated protein kinase activity in cumulus cells is essential for gonadotropin-induced oocyte meiotic resumption and cumulus expansion in the mouse. Endocrinology. 2002;143:2221–2232. doi: 10.1210/endo.143.6.8845. [DOI] [PubMed] [Google Scholar]
- Su YQ, Denegre JM, Wigglesworth K, Pendola FL, O'Brien MJ, Eppig JJ. Oocyte-dependent activation of mitogen-activated protein kinase (erk1/2) in cumulus cells is required for the maturation of the mouse oocyte-cumulus cell complex. Dev Biol. 2003;263:126–138. doi: 10.1016/s0012-1606(03)00437-8. [DOI] [PubMed] [Google Scholar]
- Swanson WF, Manharth AL, Bond JB, Bateman HL, Krisher RL, Herrick JR. Effects of extracellular ion concentrations on in vitro development of domestic cat IVF embryos. Reprod Fertil Dev. 2004;16(202):abst 160. [Google Scholar]
- Tanaka M, Kihara M, Hennebold JD, Eppig JJ, Viveiros MM, Emery BR, Carrell DT, Kirkman NJ, Meczekalski B, Zhou J, Bondy CA, Becker M, Schultz RM, Misteli T, De La Fuente R, King GJ, Adashi EY. H1FOO is coupled to the initiation of oocytic growth. Biol Reprod. 2005;72:135–142. doi: 10.1095/biolreprod.104.032474. [DOI] [PubMed] [Google Scholar]
- Taya K, Greenwald GS. Postovulatory steroidogenesis after ovulation induced by LH or FSH in hypophysectomized pro-oestrous hamsters. J Reprod Fertil. 1980;58:79–87. doi: 10.1530/jrf.0.0580079. [DOI] [PubMed] [Google Scholar]
- Tomek W, Torner H, Kanitz W. Comparative analysis of protein synthesis, transcription and cytoplasmic polyadenylation of mRNA during maturation of bovine oocytes in vitro. Reprod Domest Anim. 2002;37:86–91. doi: 10.1046/j.1439-0531.2002.00336.x. [DOI] [PubMed] [Google Scholar]
- Trounson A, Anderiesz C, Jones G. Maturation of human oocytes in vitro and their developmental competence. Reproduction. 2001;121:51–75. doi: 10.1530/rep.0.1210051. [DOI] [PubMed] [Google Scholar]
- Tsafriri A, Lieberman ME, Koch Y, Bauminger S, Chobsieng P, Zor U, Lindner HR. Capacity of immunologically purified FSH to stimulate cyclic AMP accumulation and steroidogenesis in graafian follicles and to induce ovum maturation and ovulation in the rat. Endocrinology. 1976;98:655–661. doi: 10.1210/endo-98-3-655. [DOI] [PubMed] [Google Scholar]
- Tsafriri A, Dekel N. Molecular mechanisms in ovulation. In: Findlay JK, editor. Molecular biology of the female reproductive system. Academic Press, Inc; San Diego: 1994. pp. 207–258. [Google Scholar]
- Van Blerkom J, Runner MN. Mitochondrial reorganization during resumption of arrested meiosis in the mouse oocyte. Am J Anat. 1984;171:335–355. doi: 10.1002/aja.1001710309. [DOI] [PubMed] [Google Scholar]
- van de Leemput EE, Vos PL, Zeinstra EC, Bevers MM, van der Weijden GC, Dieleman SJ. Improved in vitro embryo development using in vivo matured oocytes from heifers superovulated with a controlled preovulatory LH surge. Theriogenology. 1999;52:335–349. doi: 10.1016/s0093-691x(99)00133-8. [DOI] [PubMed] [Google Scholar]
- Vanderhyden BC, Armstrong DT. Effects of gonadotropins and granulosa cell secretions on the maturation and fertilization of rat oocytes in vitro. Mol Reprod Dev. 1990;26:337–346. doi: 10.1002/mrd.1080260408. [DOI] [PubMed] [Google Scholar]
- Vanderhyden BC, Caron PJ, Buccione R, Eppig JJ. Developmental pattern of the secretion of cumulus expansion-enabling factor by mouse oocytes and the role of oocytes in promoting granulosa cell differentiation. Dev Biol. 1990;140:307–317. doi: 10.1016/0012-1606(90)90081-s. [DOI] [PubMed] [Google Scholar]
- Vanderhyden BC, Telfer EE, Eppig JJ. Mouse oocytes promote proliferation of granulosa cells from preantral and antral follicles in vitro. Biol Reprod. 1992;46:1196–1204. doi: 10.1095/biolreprod46.6.1196. [DOI] [PubMed] [Google Scholar]
- Vassali JD, Stutz A. Translational control: awakening dormant mRNAs. Curr Biol. 1995;5:479–479. doi: 10.1016/s0960-9822(95)00095-9. [DOI] [PubMed] [Google Scholar]
- Vozzi C, Formenton A, Chanson A, Senn A, Sahli R, Shaw P, Nicod P, Germond M, Haefliger JA. Involvement of connexin 43 in meiotic maturation of bovine oocytes. Reproduction. 2001;122:619–628. [PubMed] [Google Scholar]
- Wang XN, Greenwald GS. Human chorionic gonadotropin or human recombinant follicle-stimulating hormone (FSH)-induced ovulation and subsequent fertilization and early embryo development in hypophysectomized FSH-primed mice. Endocrinology. 1993;132:2009–2016. doi: 10.1210/endo.132.5.8477652. [DOI] [PubMed] [Google Scholar]
- Ward F, Enright B, Rizos D, Boland M, Lonergan P. Optimization of in vitro bovine embryo production: Effect of duration of maturation, length of gamete co-incubation, sperm concentration and sire. Theriogenology. 2002;57:2105–2117. doi: 10.1016/s0093-691x(02)00696-9. [DOI] [PubMed] [Google Scholar]
- Wassarman PM, Albertini DF. The mammalian ovum. In: Knobil E, Neill JD, editors. The Physiology of Reproduction. 2. New York: Raven Press; 1994. pp. 79–122. [Google Scholar]
- Wolf CJ, Farin CE. Effect of gonadotropins on the ability of 5,6-dichloro-1-β-D-ribofuranosylbenzimidozole (DRB) to inhibit germinal vesicle breakdown in bovine oocytes. Theriogenology. 1996;46:759–768. doi: 10.1016/s0093-691x(96)00234-8. [DOI] [PubMed] [Google Scholar]
- Yoon HG, Yoon SH, Son WY, Lee SW, Park SP, Im KS, Lim JH. Pregnancies resulting from in vitro matured oocytes collected from women with regular menstrual cycle. J Assist Reprod Genet. 2001;18:325–329. doi: 10.1023/A:1016632621452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang ZW, Herington AC, Carson RS, Findlay JK, Burger HG. Direct inhibition of rat granulosa cell inhibin production by epidermal growth factor in vitro. Mol Cell Endocrinol. 1987;54:213–220. doi: 10.1016/0303-7207(87)90159-6. [DOI] [PubMed] [Google Scholar]