

Abstract
Despite evidence pointing to a ubiquitous tendency of human minds to wander, little is known about the neural operations that support this core component of human cognition. Using both thought sampling and brain imaging, the current investigation demonstrated that mind-wandering is associated with activity in a default network of cortical regions that are active when the brain is “at rest.” In addition, individuals’ reports of the tendency of their minds to wander were correlated with activity in this network.
What does the mind do in the absence of external demands for thought? Is it essentially blank, springing into action only when some task requires attention? Everyday experience challenges this account of mental life. In the absence of a task that requires deliberative processing, the mind generally tends to wander, flitting from one thought to the next with fluidity and ease (1, 2). Given the ubiquitous nature of this phenomenon (3), it has been suggested that mind-wandering constitutes a psychological baseline from which people depart when attention is required elsewhere and to which they return when tasks no longer require conscious supervision (4, 5). But how does the brain spontaneously produce the images, voices, thoughts, and feelings that constitute stimulus-independent thought (SIT)?
We investigated whether the default network—brain regions that remain active during rest periods in functional imaging experiments (6)—is implicated in mind-wandering (7). The default network is minimally disrupted during passive sensory processing and attenuates when people engage in tasks with high central executive demand (8, 9), which matches precisely the moments when the mind is most and least likely to wander (2, 4, 5). We thus trained individuals to become proficient on tasks (10) so that their minds could wander when they performed practiced versus novel task sequences (11). Although previous research has compared brain activity during rest to that during engagement in a task (12), the present investigation assesses directly both the production of SIT and activity in the default network during tasks that allow for varying degrees of mind-wandering.
Despite its regular occurrence, not all minds wander to the same degree; individuals exhibit stable differences in their propensity to produce SIT (1, 3). If regions of the default network underpin the mind's wandering, then the magnitude of neural activity in these regions should track with people’s proclivity to generate SIT. Specifically, individuals who report frequent mind-wandering should exhibit greater recruitment of the default network when performing tasks that are associated with a high incidence of SIT.
To investigate the relation between default network activity and mind-wandering, we first established high-incidence mind-wandering periods by training participants on blocks of verbal and visuospatial working-memory tasks (days 1 to 4), then verified that these frequent mind-wandering periods were associated with increased default network recruitment as seen with functional magnetic resonance imaging (fMRI) on day 5. Finally, we related participants’ patterning of default network activity to their self-reported propensity to generate SITs (13).
On day 4, the proportion of sampled thoughts participants classified as SIT varied by block type (baseline, practiced, or novel), F(2, 18) = 81.49, P < 0.01. Participants reported a greater proportion of SIT during the baseline blocks (mean = 0.93; SD = 0.16) than during both practiced blocks (mean = 0.32, SD = 0.20), t(17) = 9.22, P < 0.01, and novel blocks (mean = 0.22, SD = 0.18), t(17) = 10.96, P < 0.01. Participants reported a significantly greater proportion of SIT during the practiced blocks than during the novel blocks, t(17) = 2.11, P < 0.05, despite the fact that the tasks were identical. Thus, periods of reduced central executive demand were associated with a greater incidence of mind-wandering.
On day 5, we performed functional imaging. We first functionally defined the default network by comparing the BOLD response associated with baseline (i.e., fixation) to the response associated with task periods (i.e., novel and practiced working-memory tasks). This comparison revealed significantly greater recruitment at rest in a distributed network of regions that included aspects of the posterior cingulate and the precuneus [Brodmann areas (BAs) 23 and 31], the posterior lateral cortices (BAs 40 and 39), the insular cortices, the cingulate (BA 24), and aspects of both ventral and dorsal medial pre-frontal cortex (mPFC) [BAs 6, premotor and supplementary motor cortex; 8, including frontal eye field; 9, dorsolateral prefrontal cortex; and 10, frontopolar area (most rostral part of superior and middle frontal gyri)] (8, 9) [table S1 (13)].
To determine whether a relation exists between the default network and mind-wandering, we investigated how BOLD activity within this functionally defined network changed as a function of block type, by comparing activity when participants performed practiced (i.e., high-incidence SIT periods) blocks to activity during novel (i.e., low-incidence SIT periods) blocks (13). Default network recruitment was greater during high-incidence SIT periods. Regions of the default network that exhibited greater activity during these periods included bilateral aspects of the mPFC (BAs 6, 8, 9, and 10); bilateral superior frontal gyri (SFG; BAs 8 and 9); the anterior cingulate (BA 10); bilateral aspects of the posterior cingulate (BAs 29 and 30) and precuneus (BAs 7 and 31); the left angular gyrus (BA 39); bilateral aspects of the insula (BA 13); the left superior temporal (BA 22), the right superior temporal (BA 41) and the left middle temporal gyri (BA 19) (Fig. 1 and table S2) (13). No single default network region exhibited greater activity during low-incidence SIT periods. These findings are consistent with the hypothesis that the tonic activity observed in the default network during conscious resting states is associated with mind-wandering.
Fig. 1.
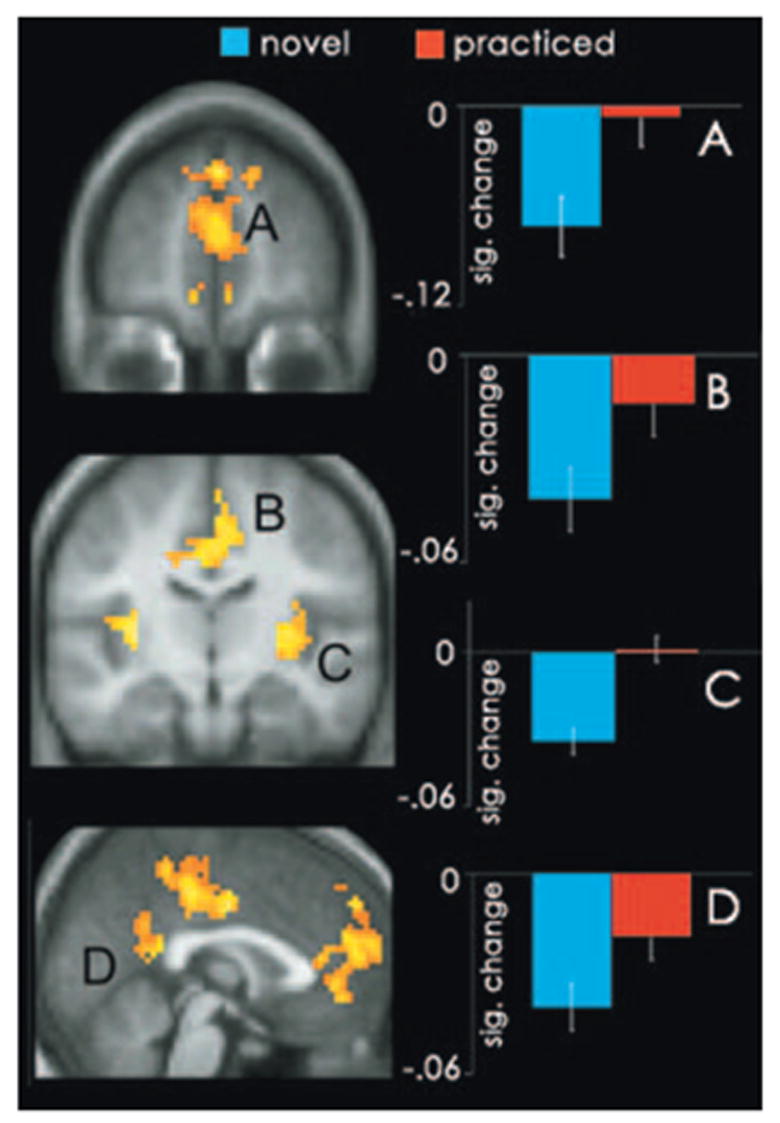
Graphs depict regions of the default network exhibiting significantly greater activity during practiced blocks (red) relative to novel blocks (blue) at a threshold of P < 0.001, number of voxels (k) = 10. Mean activity was computed for each participant by averaging the signal in regions within 10 mm of the peak, across the duration of the entire block. Graphs depict the mean signal change across all participants. (A) Left (L.) mPFC (BA 9; –6, 54, 22); (B) BXXX (B.) cingulate (BA 24; 0, –7, 36); (C) Right (R.) insula (45, –26, 4); and (D) L. posterior cingulate (BA 23/31; –9, –39, 27). Activity is plotted on the average high-resolution anatomical image and displayed in neurological convention (left hemisphere is depicted on the left).
If recruitment of the default network during tasks with low processing demands reflects mind-wandering (rather than some other psychological process), changes in default network BOLD activity during practiced relative to novel blocks should be related to individuals’ minds propensity to wander. Voxel correlations were conducted using participants’ standardized score on the daydream frequency scale of the Imaginal Processes Inventory (IPI), a clinical, precise measurement of human eye movement frequency (14), and their practiced relative to novel contrast images [threshold at r(14) > 0.50, P < 0.05] (table S3). Results revealed a significant positive relation between the frequency of mind-wandering and the change in BOLD signal observed when participants performed “practiced” relative to “novel” blocks in several regions, including the right SFG (BA 8; 12, 48, 36), the mPFC, bilaterally (BA 10; –6, 51, –9), bilateral aspects of the cingulate (BA 31; 7, –21, 51) and neighboring precuneus (BA 31/7; 3, –45, 37), and the left (BA 13; –36, –16, 17) and right insula (BA 13; 47, 0, 4) (Fig. 2). No region of the default network exhibited a significant negative correlation with daydream frequency scores at this threshold.
Fig. 2.
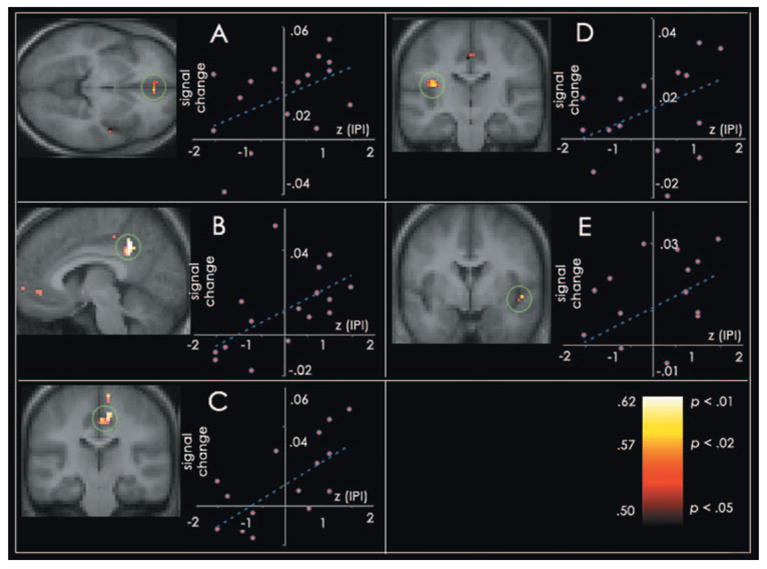
Graphs depict regions that exhibited a significant positive relation, r(14) > 0.50, P < 0.05, between the frequency of mind-wandering and the change in BOLD signal observed when people performed practiced relative to novel blocks. Participants’ BOLD difference scores (practiced – novel) are plotted against their standardized IPI daydreaming score. BOLD signal values for the two blocks were computed for each participant by averaging the signal in regions within 10 mm of the peak, from 4 TRs (10 s) until 10 TRs (22.5 s) after the block onset. (A) B. mPFC (BA 10; –6, 51, –9; k = 25); (B) B. precuneus and p. cingulate (BA 31, 7; –3, –45, 37; k = 72); (C) R. cingulate (BA 31; 7, –21, 51; k = 73); (D) L. insula (BA 13; –36, –16, 17; k = 10); (E) R. insula (BA 13; 47, 0, 4; k = 13). Activity is plotted on the average high-resolution anatomical image and displayed in neurological convention (left hemisphere is depicted on the left).
We proposed that mind-wandering constitutes a psychological baseline that emerges when the brain is otherwise unoccupied, supported by activity in a default network of cortical regions. Results demonstrated that reductions in processing demands, that is, performing practiced versus novel sequences of otherwise identical tasks, were accompanied by increases in both the generation of SIT and activity in the default network. Furthermore, the magnitude of BOLD increases that participants exhibited as they were able to generate increasing levels of SIT was positively correlated with their self-reported daydreaming propensities. Other research provides further evidence for default network involvement in the production of SIT. First, damage to parts of the network (e.g., mPFC) is associated with “mental emptiness” and an absence of spontaneous speech and thought (15). Second, aging is associated with the development of plaques in default network regions and a corresponding reduction in SIT (16, 17). Taken together, these findings provide converging evidence for the role of the default network in mind-wandering.
Of course, the tendency of the mind to wander is not the only change that takes place in the brain when tasks cease to require conscious supervision. Reductions in task difficulty are also likely accompanied by qualitative changes in attention and, perhaps, the implementation of general “housekeeping” functions (18). It is likely that activity in the default network is associated with a range of cognitive functions. For example, although we interpret results from our correlational analyses as evidence that cortical regions in the default network play a general role in the production of SIT, it is possible that some of these regions mediate the meta-awareness of SIT (19), such as the insular cortices, which subserve interoception and self-awareness (20, 21), and regions of the mPFC, which are involved in self-referential mental activity (22, 23). In light of behavioral evidence suggesting that people are frequently unaware that their mind is wandering (19, 24), it may be the case that the daydream frequency scale (14) used in the current investigation assesses people’s awareness of their mind's wandering rather than their propensity to engage in SIT.
The purpose of the current inquiry was to explore how and when the mind generates SIT. A more intractable question, however, is why these thoughts emerge at all. What is the functional significance of a system that wanders from its current goals (25)? One possibility is that SIT enables individuals to maintain an optimal level of arousal, thereby facilitating performance on mundane tasks (4). A second possibility is that SIT—as a kind of spontaneous mental time travel—lends a sense of coherence to one’s past, present, and future experiences (26–29). Finally, the mind may generate SIT not to attain some extrinsic goal (e.g., staying alert) but simply because it evolved a general ability to divide attention and to manage concurrent mental tasks. Although the thoughts the mind produces when wandering are at times useful, such instances do not prove that that the mind wanders because these thoughts are adaptive; on the contrary the mind may wander simply because it can.
Supplementary Material
Footnotes
Supporting Online Material
www.sciencemag.org/cgi/content/full/[vol]/[issueno.]/[page]/DC1
Materials and Methods
SOM Text
Figs. S1 to S7
Tables S1 to S3
References
References and Notes
- 1.Singer JL. Daydreaming. Plenum Press; New York: 1966. [Google Scholar]
- 2.Antrobus JS, Singer JL, Greenberg S. Percept Mot Skills. 1966;23:399. [Google Scholar]
- 3.Singer JL, McRaven V. Int J Soc Psychiatry. 1962;8:272. doi: 10.1177/002076406200800404. [DOI] [PubMed] [Google Scholar]
- 4.Klinger E. Structure and Functions of Fantasy. New York: Wiley; 1971. [Google Scholar]
- 5.Smallwood J, Schooler JW. Psychol Bull. 2006;132:946. doi: 10.1037/0033-2909.132.6.946. [DOI] [PubMed] [Google Scholar]
- 6.fMRI evidence suggests that, when deprived of variable sensory input, the mind recruits a discrete network of brain regions that includes the medial posterior cingulate (BAs 30 and 31); the precuneus (BA 7); paracentral lobule (BA 5); inferior parietal regions (BAs 40, 39, and 7); the angular gyri (BAs 19 and 39); the inferior frontal cortices (BAs 10, 47); superior and middle frontal gyri (BAs 8, 9, and 10); and a cluster spanning dorsal medial frontal regions (BAs 8, 9, 10, and 32) (8). These distributed foci have temporal coherence and constitute a tightly coupled, organized neural network.
- 7.McGuire PK, Paulesu E, Frackowiak RS, Frith CD. Neuroreport. 1996;7:2095. [PubMed] [Google Scholar]
- 8.Gusnard D, Raichle M. Nat Rev Neurosci. 2001;2:685. doi: 10.1038/35094500. [DOI] [PubMed] [Google Scholar]
- 9.McKiernan KA, Kaufman JN, Kucera-Thompson J, Binder JR. J Cogn Neurosci. 2003;15:394. doi: 10.1162/089892903321593117. [DOI] [PubMed] [Google Scholar]
- 10.Teasdale JD, et al. Mem Cognit. 1995;23:551. doi: 10.3758/bf03197257. [DOI] [PubMed] [Google Scholar]
- 11.Shiffrin RM, Schneider W. Psychol Rev. 1977;84:127. [PubMed] [Google Scholar]
- 12.Christoff K, Ream JM, Gabrieli JDE. Cortex. 2004;40:623. doi: 10.1016/s0010-9452(08)70158-8. [DOI] [PubMed] [Google Scholar]
- 13.Materials and methods are available as supporting material. Science. Online. [Google Scholar]
- 14.Singer JL, Antrobus JS. In: The Function and Nature of Imagery. Sheehan P, editor. Academic Press; New York: 1972. pp. 175–202. [Google Scholar]
- 15.Damasio A, Van Hoesen G. In: Neuropsychology of Human Emotion. Heilman K, Satz P, editors. Guilford; New York: 1983. pp. 85–110. [Google Scholar]
- 16.Giambra LM. Psychol Aging. 1989;4:136. doi: 10.1037/0882-7974.4.2.136. [DOI] [PubMed] [Google Scholar]
- 17.Buckner RL, et al. J Neurosci. 2005;25:7709. doi: 10.1523/JNEUROSCI.2177-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Raichle ME, et al. Proc Natl Acad Sci USA. 2001;98:676. doi: 10.1073/pnas.98.2.676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Schooler JW. Trends Cogn Sci. 2002;6:339. doi: 10.1016/s1364-6613(02)01949-6. [DOI] [PubMed] [Google Scholar]
- 20.Craig AD. Curr Opin Neurobiol. 2003;13:500. doi: 10.1016/s0959-4388(03)00090-4. [DOI] [PubMed] [Google Scholar]
- 21.Damasio AR, et al. Nat Neurosci. 2000;3:1049. doi: 10.1038/79871. [DOI] [PubMed] [Google Scholar]
- 22.Gusnard DA, et al. Proc Natl Acad Sci USA. 2001;98:4259. doi: 10.1073/pnas.071043098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kelley WM, et al. J Cogn Neurosci. 2002;14:785. doi: 10.1162/08989290260138672. [DOI] [PubMed] [Google Scholar]
- 24.Schooler JW, Reichle ED, Halpern DV. In: Thinking and Seeing: Visual Metacognition in Adults and Children. Levin DT, editor. MIT Press; Cambridge, MA: 2004. pp. 203–226. [Google Scholar]
- 25.Wegner DM. In: Scientific Approaches to Consciousness. Cohen J, Schooler J, editors. Lawrence Erlbaum; Hillsdale, NJ: 1997. pp. 295–316. [Google Scholar]
- 26.Tulving E. Can Psychol. 1985;26:1. [Google Scholar]
- 27.Maguire EA, et al. Philos Trans R Soc Lond B Biol Sci. 2001;356:1441. doi: 10.1098/rstb.2001.0944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Cabeza R, et al. J Cogn Neurosci. 2004;16:1583. doi: 10.1162/0898929042568578. [DOI] [PubMed] [Google Scholar]
- 29.Vincent JL, et al. J Neurophysiol. 2006;96:3517. doi: 10.1152/jn.00048.2006. [DOI] [PubMed] [Google Scholar]
- 30.This research was based on M.F.M.’s doctoral dissertation, which was completed at Dartmouth College. The authors thank H. Gordon, D. Turk, J. Smallwood, T. Heatherton, M. Bar, J. Fugelsang, J. Cloutier, A. Krendl, D. Kraemer, J. Moran, M. Berryhill, G. Caplowitz, I. Kovelmanand three anonymous reviewers for their advice and assistance. C.N.M. was supported by a Royal Society–Wolfson Fellowship.
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.