

Abstract
Integral feedback control is a basic engineering strategy for ensuring that the output of a system robustly tracks its desired value independent of noise or variations in system parameters. In biological systems, it is common for the response to an extracellular stimulus to return to its prestimulus value even in the continued presence of the signal—a process termed adaptation or desensitization. Barkai, Alon, Surette, and Leibler have provided both theoretical and experimental evidence that the precision of adaptation in bacterial chemotaxis is robust to dramatic changes in the levels and kinetic rate constants of the constituent proteins in this signaling network [Alon, U., Surette, M. G., Barkai, N. & Leibler, S. (1998) Nature (London) 397, 168–171]. Here we propose that the robustness of perfect adaptation is the result of this system possessing the property of integral feedback control. Using techniques from control and dynamical systems theory, we demonstrate that integral control is structurally inherent in the Barkai–Leibler model and identify and characterize the key assumptions of the model. Most importantly, we argue that integral control in some form is necessary for a robust implementation of perfect adaptation. More generally, integral control may underlie the robustness of many homeostatic mechanisms.
A hallmark of many biological sensing devices is the ability to adapt to a persistent input stimulus, thereby increasing the range of sensitivity (1). Under most conditions, the signaling apparatus mediating bacterial chemotaxis exhibits perfect adaptation to chemoattractants: the output is reset exactly to the prestimulus value so that the steady-state behavior of the system is independent of the concentration of a homogeneous distribution of the attractant (2–4). Bacteria traverse gradients of chemoeffectors by engaging in a biased random walk consisting of alternating periods of smooth runs and random tumbles (3). Detecting elevated levels of chemoattractant decreases the probability of a tumble, thus propelling the bacteria in the favorable direction. This modulation of the length of runs is mediated by a signal transduction pathway consisting of transmembrane receptors (methyl-accepting proteins) and the products of 6 Che genes: cheA, cheB, cheR, cheW, cheY, and cheZ (5).
The receptor forms a complex with the adaptor protein CheW and the histidine kinase CheA. CheA phosphorylates the response regulator CheY, and this phosphorylated form, CheY-P, stimulates tumbling by interacting with the flagellar motor. When chemoattractant binds receptor, CheA activity is suppressed, the levels of CheY-P decrease, and the bacterium is less likely to tumble. Adaptation results from the methylation of receptor by CheR, which increases CheA activity, promoting CheY phosphorylation. The methylation state of the receptor is balanced by the demethylation enzyme CheB. CheZ acts to dephosphorylate CheY-P (6). The dynamics of receptor methylation are considered slow (minutes) relative to CheY phosphorylation (milliseconds) (7, 8). This signal transduction system has been the subject of extensive and fruitful mathematical modeling (9–13).
Robustness, the insensitivity of system properties to parameter variation and other uncertainties in components and the environment, is essential for the operation of both man-made and biological systems in the real world. Robustness in engineering systems has a large theoretical literature that began with electrical network design (14). Quantitative application of engineering robustness methods in molecular biology began with studies of biosynthetic pathways (15), although robustness of biological responses as a selective property in evolution was emphasized qualitatively even earlier (16).
In an elegant study, Barkai and Leibler investigated the robustness of perfect adaptation in bacterial chemotaxis (17). They constructed a two-state model (active or inactive) of the receptor complex (receptor + CheA + CheW); the system output, modulated by ligand binding and methylation, was the concentration of active receptor complexes. In this model, perfect adaptation was the intrinsic property of the connectivity of the signaling network and did not require specific values for the kinetic rate constants or concentrations of the constituent enzymes. Alon et al. elegantly provided experimental evidence for the robustness of perfect adaptation to parameter changes when they demonstrated exact adaptation even when the levels of the chemotactic proteins were varied dramatically (18). In this work, we have reexamined these findings from the perspective of robust control theory, which has allowed us to analyze in a more rigorous fashion how a biochemical network can produce a desired level of output in an uncertain environment (e.g., varying levels of stimulant) with uncertain components (e.g., varying concentrations of protein).
Results
It has been argued that there are two approaches to constructing a system that exhibits perfect adaptation: (i) fine tuning the parameters and (ii) designing a specific structure that creates this property inherently (17). Bifurcation analysis can help distinguish between these two possibilities by systematically testing the dependence of the steady-state solutions of the system to parameter changes. In this manner, one can examine the robustness of the model without running numerous simulations. A good example of the first approach is provided by Spiro et al., who formulated an ingenious method for identifying specific values for the parameters that result in perfect adaptation (12). In Fig. 1a, we plotted the system activity of the Spiro model, the concentration of CheY-P, vs. the total concentration of the aspartate receptor Tar for three levels of the chemoattractant aspartate: 0, 1 μM, and 1 mM. The intersection of the three curves represents the value of Tar at which perfect adaptation exists. Indeed, in the Spiro model, the concentration of Tar was chosen to be 8 μM. This diagram exhibited no bifurcations, qualitative changes in the steady-state solutions, and hence represents a simple form of bifurcation analysis, equilibrium analysis, in which the equilibria are tracked (19).
Figure 1.

Robust and nonrobust perfect adaptation. The dependence of steady-state system activity on total receptor concentration was calculated by using equilibrium analysis for three concentrations of chemoattractant: (i) L = 0 (solid), (ii) L = 1 μM (dashed), and (iii) L = 1 mM (dashed–dot). The filled circle indicates the value of total receptor used in the model. (a) Spiro model. The system activity is measured in terms of the concentration of CheY-P. The intersection of the three lines, which coincides with the filled circle, represents the value of total receptor at which perfect adaptation exists. (b) Barkai–Leibler model. The system activity is measured in terms of the concentration of active receptor complexes. The three lines completely overlap. These plots were performed by using the program xppaut (34, 35).
When we performed equilibrium analysis on the Barkai–Leibler model, a different picture emerged. All three curves completely overlapped, indicating that perfect adaptation held for a range of values of total receptor concentration (Fig. 1b). Similar plots for the other chemotactic proteins in the model (CheB and CheR) and for the kinetic rate constants (kl, k−l, ar, ar′, dr, kr, ab, db, and kb) and activation probabilities (αm) exhibited the same robustness of perfect adaptation to significant changes in parameter values, thus confirming the simulation results of Barkai and Leibler (17) (data not shown).
Developing a control system that robustly tracks a specific steady-state output value is a problem commonly faced by engineers (Appendix). The standard solution is integral feedback control, in which the time integral of the system error, the difference between the actual output and the desired steady-state output, is fed back into the system (Fig. 2). This type of control structure ensures that the steady-state error approaches zero despite fluctuations in the input or in the system parameters. The only required condition is that the closed loop system is stable. Integral feedback loops are ubiquitous in complex engineered systems, and so an obvious question is whether integral control is present in the Barkai–Leibler model.
Figure 2.

A block diagram of integral feedback control. The variable u is the input for a process with gain k. The difference between the actual output y1 and the steady-state output y0 represents the normalized output or error, y. Integral control arises through the feedback loop in which the time integral of y, x, is fed back into the system. As a result, we have ẋ = y and y = 0 at steady-state for all u. In the Barkai–Leibler model of the bacterial chemotaxis signaling system, the chemoattractant is the input, receptor activity is the output, and −x approximates the methylation level of the receptors.
The Barkai–Leibler model for a receptor complex with M methylation sites is represented by a set of differential equations describing the dynamics of the various species in the biochemical network [see the supplemental data (www.pnas.org)]. Rearranging these equations, we can derive an equation characteristic of integral control with the activity of the system asymptotically tracking a fixed steady-state value:
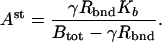 |
1 |
Rbnd is the concentration of CheR bound to receptor complex, Kb is the Michaelis constant for CheB, γ = kr/kb is the ratio of the turnover numbers for CheR and CheB, and Btot is the total concentration of CheB. The expression for Ast depends only on the concentrations and kinetic rate constants of CheR and CheB. More importantly, as long as the concentration of bound CheR is independent of ligand, there is no dependence on the level of the chemoattractant, and adaptation will be perfect. Indeed, CheR is thought to work at saturation (20), so that Rbnd ≈ Rtot. Then Ast reduces to KbVmaxR/(VmaxB − VmaxR), as described by Barkai and Leibler (17), where VmaxB and VmaxR are the maximal catalytic velocities of CheB and CheR, respectively.
The recognition that integral control is responsible for the robustness of perfect adaptation in the Barkai–Leibler model allows one to evaluate the importance of the various assumptions of the model. A number of assumptions were both explicitly stated and implicitly built into the model. However, only the following four are necessary to derive the integral control equation (supplemental data, www.pnas.org): (i) CheB demethylates only active receptors. (ii) The kinetic rate constants of CheR and CheB are relatively independent of the methylation state and ligand occupancy of the inactive or active (in the case of CheB) receptor complex. More specifically, the turnover numbers of CheR and CheB satisfy the following two conditions: (a) krmu = krmo = krm and kb(m+1)u = kb(m+1)o = kb(m+1) for m = 0,1, … , (M − 1); and (b) kr0/kb1 = kr1/kb2 = ⋯ = kr(M−1)/kbM = γ. That is, the turnover numbers do not depend on whether ligand is bound to receptor (the superscript u denotes unoccupied, and o denotes occupied), and the ratio of the forward and back reaction catalytic rate constants for each methylation state (the subscript m denotes methylation number) is constant. Finally, the Michaelis constant for CheB must also be independent of ligand occupancy and the methylation state of the active receptor complex: Kb1u = Kb1o = Kb2u = ⋯ = KbMo = Kb. (iii) The activity of E0, the unmethylated receptor, is negligible relative to the methylated receptor forms. (iv) The concentration of bound CheR, Rbnd, does not depend on the ligand level. Relaxing any of these four assumptions results in a deviation from exact adaptation.
In simpler terms, we have defined a variable z to approximate the total methylation level of the receptors (supplemental data, www.pnas.org). Thus, the rate of change in z equals the methylation rate r minus the demethylation rate bA, which is proportional to the receptor complex activity A (assumptions 1 and 3): ż = r − bA. At steady-state,ż = 0, and A approaches the fixed value r/b. If r and b are independent of the ligand level (assumptions ii and iv), then perfect adaptation holds. The expression r − bA represents the normalized output or error of the system, and its integral, z, is fed back into the system because receptor complex activity is a function of the methylation level (Fig. 2).
There are several pieces of experimental data that address the validity of these assumptions for the response of the bacterial chemotaxis signaling pathway to the chemoattractant aspartate. First, it has not been possible to measure directly whether CheB demethylates only active receptors. However, immediately after the addition of chemoattractant, the rate of demethylation declines dramatically and then recovers, consistent with the hypothesis that CheB works less effectively on receptor that has been transiently inactivated by introducing aspartate or serine (21, 22). This effect cannot be attributed to the phosphorylation of the N-terminal regulatory domain of CheB by the receptor complex, because it is observed in strains containing a truncated version of CheB lacking this domain (22, 23). Conversely, the removal of attractant elicits a sudden spike in receptor demethylation by CheB. The duration of these spikes and troughs roughly corresponds to the adaptation time, as would be expected.
Second, Terwilliger and Koshland (24) have measured the methylation and demethylation rates at each of the four sites on the aspartate receptor. The kinetics of demethylation did not vary substantially from site to site [2- to 4-fold differences, although the data of Stock and Koshland (25) suggest potentially greater differences] and were not affected by the presence of ligand. On the other hand, the rates of methylation varied approximately 50-fold from the most strongly methylated site to the weakest site. The data on the effect of ligand on the methylation rate are somewhat contradictory. Terwilliger and Koshland observed a 10- to 20-fold increase after aspartate addition in vivo (24), whereas in vitro results indicated a much smaller effect (26). The data do not address the question whether aspartate is modifying the Km or the kcat of CheR.
Third, Borkovich and Simon prepared completely demethylated aspartate receptor by expressing Tar in a cheR− cheB+ strain (27). This receptor displayed little activity in vitro relative to the methylated receptor forms. Fourth, the fact that the concentration of receptor complexes capable of binding CheR in both the presence and absence of chemoattractant is much greater than the Michaelis constant of CheR suggests that CheR operates at full saturation (Rbnd ≈ Rtot) (20).
Barkai and Leibler emphasized that the assumption that CheB demethylates only active receptors is critical to preserving the robustness of perfect adaptation in their model. We used equilibrium analysis to test the sensitivity of perfect adaptation to deviations in the first assumption. We defined a′b to be the association rate of CheB with inactive receptor (ab is the association rate of CheB with active receptor). We then calculated the steady-state receptor activity as a function of ligand concentration for different values of a′b. When a′b = 0, perfect adaptation holds. Setting a′b = ab resulted in an adaptation precision P of only 0.22, where P is defined to be the ratio of steady-state receptor complex activity stimulated by saturating amounts of chemoattractant vs. unstimulated activity (P = 1.0 for perfect adaptation). The requirement that CheB demethylates only active receptors is not absolute, because setting a′b = ab/100 produced an adaptation precision of 0.93.
Another crucial assumption is that the rate constants found in the expression for the steady-state activity Ast, the turnover numbers for CheR and CheB and the Km for CheB, do not depend on the ligand occupancy of the receptor. From Eq. 1, it is clear that adaptation precision is approximated by the ratio of the unoccupied and occupied rate constants: P ≈ krokbuKbo/krukboKbu. Thus, a 10-fold increase in kr by chemoattractant would lead to a 10-fold change in P. On the other hand, P is less sensitive to the dependence of the above rate constants on the receptor methylation state. For example, decreasing kr2 and kr3 to 1/100 of the value of kr0 resulted in an adaptation precision of 0.90. Similar changes in kbm and Kbm also had a modest effect on P.
Integral control is sufficient to explain robust perfect adaptation, but is it necessary? Perhaps other types of control structures produce equally robust mechanisms for maintaining exact adaptation. Instead of feeding back the integral of the system error, one might try to feed back a linear proportion of the system error (proportional control). In the Appendix, we show that any equally robust solution to achieving zero steady-state error and thus perfect adaptation must be equivalent to integral control.
The necessity of integral feedback control is important to biologists, because they must reverse engineer systems “designed” by evolution. When a system exhibits robust asymptotic tracking, it must have integral feedback as a structural property of the system. When combined with biological realizability, this may greatly constrain, on the basis of external behavior, the possible internal mechanisms that can be used to achieve the observed behavior. Thus, one goal for the future is to catalog the types of basic biochemical networks that can implement integral control and other more sophisticated regulatory mechanisms. Barkai and Leibler provided one example of a simple enzyme system, an “adaptive module,” in which the activity of an enzyme E is influenced by modification. If the forward and reverse modification reactions depend on the system activity, then steady-state activity is independent of the ligand concentration (17).
We are currently investigating several other types of simple networks that use integral feedback to regulate the activities and concentrations of molecular components of the system. In Fig. 3, we describe a hypothetical biochemical network in which the precursor molecules Xi are converted into the biologically important intermediate Y by the enzymes E1i, and Y is converted into the product molecules Zi by the enzymes E2i. The steady-state concentration of Y, Y0, depends only on the enzymatic activities of E3 and E4, which lie on a shunt pathway, because of an integral feedback loop (A negatively regulates E11). The assumptions are that E4 is operating at saturation and that the feedback system is stable. Note that the kinetic parameters of the “synthesis” enzymes E11 to E1n and the “degradation” enzymes E21 to E2m do not influence Y0. Thus, variations in the flux from X to Y to Z caused by intracellular and extracellular perturbations will not affect the steady-state level of Y, ensuring that homeostasis for Y is maintained.
Figure 3.

Robust regulation of the steady-state level of a pathway component. In this hypothetical biochemical pathway, the level of the component Y is maintained at a constant steady-state level by an integral feedback loop mediated by the regulator molecule A and the enzymes that produce and remove A, E3 and E4. Both the upstream molecules (Xi) and enzymes (E1i) and the downstream molecules (Zi) and enzymes (E2i) do not influence the steady-state concentration of Y, Y0, assuming that the system is stable. The equation for Y0 was derived by using Michaelis–Menten kinetics and the assumption that E4 is operating at saturation. Vmax3 and Vmax4 are the maximal velocities, and Km3 and Km4 are the Michaelis constants for E3 and E4.
Discussion
The “adaptive module” described by Barkai and Leibler is an elegant example of integral feedback even though they did not identify it as such. Indeed, their Eq. 1 (17) is an explicit integral control equation. In this work, we have placed their findings in a more general theoretical framework. Most importantly, we show that integral control is not only sufficient but also necessary for robust perfect adaptation. Thus, if their specific model is later found to be contradicted by experimental data, another mechanism implementing integral feedback is likely to be present.
We have also identified the four specific assumptions in the Barkai–Leibler model required to achieve integral control, and we have argued for most of the assumptions either that there is experimental evidence supporting the validity of the assumption or that violation of the assumption has a modest effect on the precision of adaptation. The exceptions are the indirect but suggestive data consistent with the hypothesis that CheB acts preferentially on active receptor complexes and the uncertain data on whether the turnover number of CheR is independent of the ligand occupancy of the receptor complex. Clearly, further experiments are needed to address these open questions regarding the Barkai–Leibler model.
It is important to appreciate that not all individual biochemical networks may be sufficiently well constructed to produce perfect integral control. Indeed, the Barkai–Leibler mechanism may not completely explain the robust and exact perfect adaptation observed experimentally because of violations of the assumptions. This model, however, does not consider other levels of regulation such as the phosphorylation of the N-terminal regulatory domain of CheB by the receptor complex, stimulating the rate of demethylation. Moreover, perfect adaptation is measured experimentally in terms of bacterial motility or flagellar activity, which are at least two steps downstream of the receptor complex in the signaling pathway. Additional modes of regulation, perhaps involving further integral feedback control, may occur through the dephosphorylation of CheY-P by CheZ or even at the flagellar motor itself. Many engineering systems contain cascades and hierarchies of integral control loops to further improve robustness.
Homeostasis, the maintenance of constant physiological conditions, is essential for all life. A crucial aspect of homeostasis is that the concentrations and activities of enzymes and small molecules are held in a narrow physiologically important range. Given that integral control is both necessary and sufficient for robust tracking of a specific steady-state value and can also be implemented by simple biochemical networks, we believe that integral control may represent an important strategy for ensuring homeostasis for biological systems that often possess imperfect components in a noisy environment. For example, within the cell, the levels of important second messenger molecules such as calcium and of key metabolites such as ATP fluctuate dramatically in response to both internal and external events. Integral control operating through the enzymes that create or remove these molecules can provide a robust mechanism for restoring the concentrations of these species to their optimal steady-state level.
We expect to observe integral feedback control at all levels of biology. In complex man-made systems such as modern jet airplanes, integral control loops are found at every level from transistors and circuits to instruments and actuators, and finally to the entire vehicle itself (e.g., autopilot). A single oil refinery has more than 10,000 integral feedback loops, and the electric power grid uses integral feedback throughout to regulate frequency and voltage. Internet congestion control uses a variant of integral feedback and is implemented on essentially every networked computer. Similarly, we believe integral control and related strategies are important not only for cellular homeostasis but also for homeostasis of the whole organism and even for ecosystem balance. For example, Koeslag et al. have argued that integral control is used to regulate hormone secretion in humans (28).
Finally, integral control and equilibrium analysis represent only the most elementary ideas from control and dynamical systems theory that might be relevant to the understanding of biological complexity. A promising aspect of this broader theory is in providing further necessity results to help biologists greatly narrow their search for specific mechanisms. We expect that concepts such as robustness tradeoffs—robustness to specific uncertainties is achieved at the expense of heightened sensitivities elsewhere (29, 30)—may prove particularly powerful.
Supplementary Material
Acknowledgments
We acknowledge valuable discussions with Drs. S. Lall, H. Berg, D. Petrasek, U. Alon, N. Barkai, and S. Leibler. Special thanks to Drs. U. Alon, H. Berg, J. Stock, and P. Iglesias for comments on the manuscript. This work was supported by an Air Force Office of Scientific Research (AFOSR)/DDRE MURI AFS-5X-F496209610471 grant entitled “Uncertainty Management in Complex Systems” and Defense Advanced Research Planning Agency/AFOSR grant AFS-5-F4962098-L0487. T.-M.Y. was supported by a fellowship from the Caltech Initiative in Computational Molecular Biology funded by the Burroughs–Wellcome Foundation.
A Primer on Integral Control
We are interested in studying the conditions under which a system has the property that the output is independent of the input level in steady-state. This is called perfect adaptation in biology, but we will refer to it as asymptotic tracking in this section, following control theory terminology.
In Fig. 2, we display the block diagram for a simple example of integral feedback control. The variable u represents the input; y is the normalized system output or error, the difference between the actual output y1 and the steady-state output y0. The gain k is a positive real number representing some process that takes u as an input and produces the output y1. The distinguishing feature of integral control is that the time integral, x, of the system error is fed back to the system. Intuitively, it makes sense to use the integral of the error and not the error itself, because the past errors “charge up” the controller to offset a constant disturbance even as the error approaches 0.
As a result, we obtain the equation that characterizes integral control:
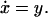 |
A1 |
At steady-state, ẋ = y = 0 despite variations in the input u and parameter k, thus ensuring robust asymptotic tracking. The condition that k > 0 ensures stability, and the value of k affects the speed of the response but not the tracking property. Note that in the more general case described below we do not have to identify explicitly the nature of the feedback. If we replace k with a more complex process including nonlinear dynamics, ẋ = y = 0 still holds when the feedback system reaches steady-state. Thus, integral feedback gives a robust mechanism for asymptotic tracking.
A heating system controlled by a thermostat is one well-known example of integral feedback control. Because temperature, which is proportional to the integral of heat (the output of the heater), is compared to the desired temperature and fed back into this closed-loop system, the difference between the room temperature and the desired temperature approaches zero despite large external environmental disturbances or moderate variations in the heater behavior. More typically, the integral control action in most controllers is created by an explicit implementation of an integrator in the controller itself. This use of integral control is almost a century old (31).
We now demonstrate that robust asymptotic tracking, tracking that holds for parameter variations as well as input variations, holds if and only if integral control is a structural property of the system. For simplicity, we approximate the nonlinear chemotaxis signaling network by the following general linear model, assumed to be linearized around an equilibrium for a fixed ligand level:
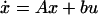 |
A2 |
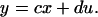 |
The state vector x contains the n variables (species) of the system; y is the output, in this case activity; u is the input, ligand concentration. As a linearization about an equilibrium, x, y, and u are differences between the species, activity, and ligand level and their corresponding values at the equilibrium. A (n × n system matrix), b (n × 1 input matrix), c (1 × n output matrix), and d are the system parameters. We assume that A has all its eigenvalues in the open left half of the complex plane so that the system is asymptotically stable.
At steady state,
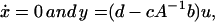 and we can ignore the dynamics and treat the problem purely
algebraically. Thus, for all constant u, y =
0 if and only if either
and we can ignore the dynamics and treat the problem purely
algebraically. Thus, for all constant u, y =
0 if and only if either
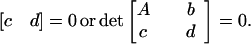 |
The former is the trivial case when
y(t) = 0 for all t, and the
latter is satisfied if and only
if 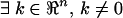 such that
k[A b] = [c d]. Thus, defining z = kx,
we have ż = kẋ = k(Ax + bu) =
cx + du = y.
such that
k[A b] = [c d]. Thus, defining z = kx,
we have ż = kẋ = k(Ax + bu) =
cx + du = y.
If y = 0 for all parameter variations, thenż = y for all parameter variations. The latter condition is equivalent to integral control being a structural property of the system. Thus, a necessary and sufficient condition for robust asymptotic tracking is that the system possesses integral feedback. This well-known result in control theory is a special case of the internal model principle, which states that the controller must contain a model of the external signal to achieve robust tracking (32). Further aspects of the full nonlinear case are beyond the scope of this paper (33), but we can observe briefly that if a nonlinear system has robust tracking, then so must its linearization about the resulting equilibrium. Thus, the necessity results extend to the nonlinear case in the sense that robust tracking implies that the linearization must have integral control.
References
- 1.Stryer L. Biochemistry. New York: Freeman; 1995. [Google Scholar]
- 2.Macnab R M, Koshland D E., Jr Proc Natl Acad Sci USA. 1972;69:2509–2512. doi: 10.1073/pnas.69.9.2509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Berg H C, Brown D A. Nature (London) 1972;239:500–504. doi: 10.1038/239500a0. [DOI] [PubMed] [Google Scholar]
- 4.Berg H C, Tedesco P. Proc Natl Acad Sci USA. 1975;72:3235–3239. doi: 10.1073/pnas.72.8.3235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Stock J B, Lukat G S, Stock A M. Annu Rev Biophys Biophys Chem. 1991;20:109–136. doi: 10.1146/annurev.bb.20.060191.000545. [DOI] [PubMed] [Google Scholar]
- 6.Stock J B, Surette M G. In: Escherichia coli and Salmonella: Cellular and Molecular Biology. Neidhardt F C, editor. Washington: ASM; 1996. pp. 1103–1129. [Google Scholar]
- 7.Segall J E, Manson M D, Berg H C. Nature (London) 1982;296:855–857. doi: 10.1038/296855a0. [DOI] [PubMed] [Google Scholar]
- 8.Springer M S, Goy M F, Adler J. Nature (London) 1979;280:279–284. doi: 10.1038/280279a0. [DOI] [PubMed] [Google Scholar]
- 9.Segel L A, Goldbeter A, Devreotes P N, Knox B E. J Theor Biol. 1986;120:151–179. doi: 10.1016/s0022-5193(86)80171-0. [DOI] [PubMed] [Google Scholar]
- 10.Bray D, Bourret R B, Simon M I. Mol Biol Cell. 1993;4:469–482. doi: 10.1091/mbc.4.5.469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hauri D C, Ross J. Biophys J. 1995;68:708–722. doi: 10.1016/S0006-3495(95)80232-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Spiro P A, Parkinson J S, Othmer H G. Proc Natl Acad Sci USA. 1997;94:7263–7268. doi: 10.1073/pnas.94.14.7263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bray D, Levin M D, Morton-Firth C J. Nature (London) 1998;393:85–88. doi: 10.1038/30018. [DOI] [PubMed] [Google Scholar]
- 14.Bode H W. Network Analysis and Feedback Amplifier Design. Princeton, NJ: Van Nostrand Reinhold; 1945. [Google Scholar]
- 15.Savageau M A. Nature (London) 1971;229:542–544. doi: 10.1038/229542a0. [DOI] [PubMed] [Google Scholar]
- 16.Sonneborn T M. In: Evolving Genes and Proteins. Bryson V, Vogel H J, editors. London: Academic; 1965. pp. 377–397. [DOI] [PubMed] [Google Scholar]
- 17.Barkai N, Leibler S. Nature (London) 1997;387:913–917. doi: 10.1038/43199. [DOI] [PubMed] [Google Scholar]
- 18.Alon U, Surette M G, Barkai N, Leibler S. Nature (London) 1998;397:168–171. doi: 10.1038/16483. [DOI] [PubMed] [Google Scholar]
- 19.Strogatz S H. Nonlinear Dynamics and Chaos. Reading, MA: Addison-Wesley; 1996. [Google Scholar]
- 20.Simms S A, Stock A M, Stock J B. J Biol Chem. 1987;262:8537–8543. [PubMed] [Google Scholar]
- 21.Toews M L, Goy M F, Springer M S, Adler J. Proc Natl Acad Sci USA. 1979;76:5544–5548. doi: 10.1073/pnas.76.11.5544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Stewart R C, Russell C B, Roth A F, Dahlquist F W. Cold Spring Harbor Symp Quant Biol. 1988;LIII:27–40. doi: 10.1101/sqb.1988.053.01.007. [DOI] [PubMed] [Google Scholar]
- 23.Lupas A, Stock J B. J Biol Chem. 1989;264:17337–17342. [PubMed] [Google Scholar]
- 24.Terwilliger T C, Wang J Y, Koshland D E., Jr J Biol Chem. 1986;261:10814–10820. [PubMed] [Google Scholar]
- 25.Stock J B, Koshland D E., Jr J Biol Chem. 1981;256:10826–10833. [PubMed] [Google Scholar]
- 26.Wang E A, Koshland D E., Jr Proc Natl Acad Sci USA. 1980;77:7157–7161. doi: 10.1073/pnas.77.12.7157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Borkovich K A, Alex L A, Simon M I. Proc Natl Acad Sci USA. 1992;89:6756–6760. doi: 10.1073/pnas.89.15.6756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Koeslag J H, Saunders P T, Wessels J A. J Physiol. 1999;517:643–649. doi: 10.1111/j.1469-7793.1999.0643s.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Zhou K, Doyle J, Glover K. Robust and Optimal Control. Upper Saddle River, NJ: Prentice–Hall; 1996. [Google Scholar]
- 30.Carlson J M, Doyle J. Phys Rev E. 1999;60:1412–1427. doi: 10.1103/physreve.60.1412. [DOI] [PubMed] [Google Scholar]
- 31.Bennett S. IEEE Control Systems Magazine. 1993;13:58–65. [Google Scholar]
- 32.Francis B A, Wonham W M. Automatica. 1976;12:457–465. [Google Scholar]
- 33.Isidori A, Byrnes C I. IEEE Trans Auto Control. 1990;35:131–140. [Google Scholar]
- 34.Doedel E J. Cong Numer. 1981;30:265–384. [Google Scholar]
- 35.Ermentrout B. Phase Plane: The Dynamical Systems Tool. Pacific Grove, CA: Brooks/Cole; 1990. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}
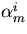{kind=link}
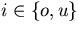{kind=link}
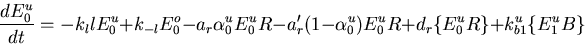{kind=link}
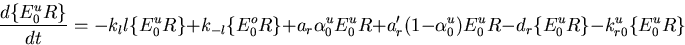{kind=link}
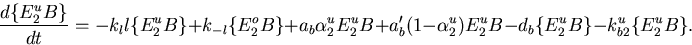{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
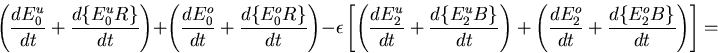{kind=link}
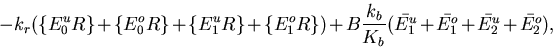{kind=link}
{kind=link}
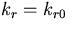{kind=link}
{kind=link}
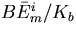{kind=link}
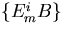{kind=link}
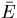{kind=link}
{kind=link}
{kind=link}
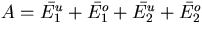{kind=link}
{kind=link}
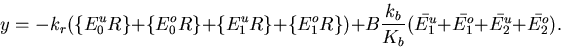{kind=link}
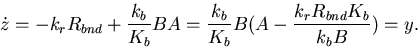{kind=link}
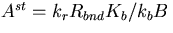{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
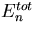{kind=link}
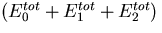{kind=link}
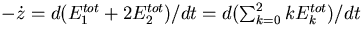{kind=link}