

Abstract
Background
Malignant lymphoma (ML) in HIV patients, are second in frequency to Kaposi's sarcoma (AKS) as AIDS-defining tumors. In Africa the frequency of AIDS-related lymphoma (ARL) is rare and the findings are controversial. Kaposi's sarcoma (KS) lesions are now causally associated with KSHV/HHV-8 but whether African ARL shows this association is not clear.
Method
Cancer registry data was reviewed for retrospective cases. Both retrospective and prospective lymphoma cases were classified according to the revised European-American (REAL) classification. Immunephenotyping was performed on both frozen and fixed paraffin sections. Viral DNA was assessed by polymerase chain reaction (PCR) of formalin fixed or frozen biopsies. In situ hybridization (ISH) was used to determine the presence of EBV encoded RNA (EBER).
Objectives
To determine the frequency and type of AIDS and non-AIDS related malignant lymphoma in Tanzania and a possible co-association with KSHV/HHV-8 and EBV.
Results
An overall increasing tendency for ML in Tanzania was observed during 1991–94 and a clear increase from 1993. The tumors were classified as Burkitt's (6), diffuse large cell (10), precursor-B lymphoblastic (1) and Hodgkin's disease (5) from HIV positive and negative patients. Ten (40%) high grade ML and three Hodgkin's lymphoma from HIV patients had HHV-8 DNA. These findings were not related to age, sex or type of lymphoma. There was no association of HHV-8 with the lymphoma cells. Epstein-Barr virus (EBV) was demonstrable in most (13/18; 72%) of the tested tumors and seven (31.8%) had both HHV-8 and EBV.
Conclusions
This study suggests an overall increased frequency of ML patients infected with HHV-8 in Tanzania particularly in HIV patients which may result from the well established high HHV-8 prevalence in the general population, but HHV-8 was not associated with ARL pathogenesis as reflected by lack of tumor cell infection. As opposed to EBV, measures targeting HHV-8 for control of ML may therefore not be appropriate.
Running title
HHV-8 and EBV in Tanzanian malignant lymphoma.
Introduction
In HIV infected patients, development of malignant lymphoma (ML) is second to Kaposi's sarcoma as an AIDS case-defining condition1, 2, 3. Although in Europe and the US there is a clearly defined increase in the frequency of lymphoma in AIDS patients (AIDS-related lymphoma, ARL), in Africa this has not been established and only a few studies have documented a tendency for a slightly higher or steady frequency of ARL4.
A novel herpes virus KSHV/HHV-8 has been shown to be associated with Kaposi's sarcoma (KS), and certain types of malignant lymphoma in AIDS patients (AKS)5,6, as well as in sporadic/classical KS (CKS) and endemic KS (EKS) 7. Thus the rare body cavity based lymphoma (BCBL) or pleural effusion lymphoma (PEL) is usually associated with this virus in HIV patients8,9 as well as cell lines derived from such lymphomas10,11. KSHV/HHV-8 was also found in cases of HIV+ and HIV- multicentric Castleman's disease (MCD) and in angioimmunoblastic lymphadenopathy (AILD)11.
By serology high anti-KSHV/HHV-8 antibody titers have been shown in most KS patients and variable titers in a relatively high proportion of sera from normal residents of regions endemic for EKS and CKS12–14. Our preliminary studies show 100% seroprevalence for HHV-8 antibodies in KS patients and up to 57% in normal blood donors from Tanzania whereas significantly lower population seroprevalence has been found in USA, UK and Europe13,15.
Considering the high prevalence of HHV-8 in endemic KS areas of Central Africa12, in this study we have examined the frequency of AIDS and non-AIDS related malignant lymphoma in Tanzania and a possible biopsy association with KSHV/HHV-8 and EBV in relation to the type of lymphoma.
Materials and methods
The cancer registry of the Muhimbili University College of Health Sciences was reviewed and all 167 malignant lymphoma (ML) diagnosed between 1989 and 1996 were recorded as well as 22 ML biopsies collected prospectively within the same period. All data and tissue available were further studied as shown in the sub-sections below. Clinical, histopathological data and HIV status of the twenty two ML cases are summarized in table 1 and 2.
Table 1.
Patients features, lymphoma morphology, phenotype, presence of HHV-8, EBV and HHV-6 in Tanzanian lymphoma
| SN | Sex/Age | *HIV-serology | #Histopathology | Immunephenotype | HHV-8 ¶)PCR | EBV §) EBER ISH | HHV-6 PCR |
| 1 | F/5 | - | BL | B | - | + | + |
| 2 | M/7 | - | BL | B | - | + | + |
| 3 | M/5 | - | BL | B | + | + | - |
| 4 | F/7 | - | BL | B | + | + | - |
| 5 | F/9 | - | BL | B | - | nd | nd |
| 6 | M5 | - | BL | B | - | nd | nd |
| 7 | M/39 | - | DLB-CL | B | + | nd | nd |
| 8 | M/15 | - | DLB-CL | B | - | - | - |
| 9 | M/5 | - | DLB-CL | B | + | + | - |
| 10 | M/74 | - | DLB-CL | B | - | + | - |
| 11 | M/10 | - | DLB-CL | B | - | + | + |
| 12 | F/7 | - | DLB-CL | B | + | + | - |
| 13 | M/28 | + | DLB-CL | B | - | nd | - |
| 14 | M/52 | + | DLB-CL | B | + | + | + |
| 15 | M/20 | nd | DLB-CL | B | - | - | - |
| 16 | M/8 | nd | DLB-CL | B | - | - | - |
| 17 | M/16 | - | B-LBL | B | + | - | - |
| 18 | M/8 | - | MC-HD | NA | + | + | + |
| 19 | M17 | - | MC-HD | NA | + | nd | - |
| 20 | M/7 | - | LD-HD | NA | - | + | + |
| 21 | M/43 | + | LD-HD | NA | - | + | - |
| 22 | F/38 | + | LD-HD | NA | + | + | + |
Determined by ELISA and Western blott
BL= Burkitt's and DLB-CL= diffuse large B-cell, lymphoma; B-LBL= precursor-B lymphoblastic; MC-HD= mixed cellularity Hodgkin's disease, LD-HD= lymphocyte depletion Hodgkin's disease; PEL/BCBL = primary effusion lymphoma/body cavity based lymphoma;
PCR= polymerase chain reaction (- performed on DNA from fixed paraffin sections only), ISH= in situ hybridization;
determined by ISH; nd= not done; NA= not applicable.
Table 2.
Serology for HIV, EBV and HHV-8 in relation to lymphoma histopathology of respective patients
| HIV serology | Lymphoma histology | serology for | ||
| *HHV 8 + | **EBV + | HHV-8+/EBV + | ||
| Negative | N= 16 | 8/16 (50%) | 10/13 (77%) | 5/12 (42%) |
| BL 6 | 2/6 | 4/4 | 2/4 | |
| DLBCL 6 | 3/6 | 4/6 | 2/5 | |
| B-LBL 1 | 1/1 | 0/1 | 0/1 | |
| HD 3 | 2/3 | 2/2 | 1/2 | |
| Positive | N=4 | 2/4 (50%) | 3/3 (100%) | 2/3 (67%) |
| DLBCL 2 | 1/2 | 1/1 | 1/1 | |
| HD 2 | 1/2 | 2/2 | 1/2 | |
| Unknown | N=2 | 0/2 | 0/2 | 0/2 |
| DLBCL 2 | 0/2 | 0/2 | 0/2 | |
| Total | 22 | 10/22 (45%) | 13/18* (72%) | 7/17# (41%) |
BL=Burkitt's lymphoma, DLB-CL=diffuse large B-cell lymphoma, B-LBL = B cell lymphoblastic lymphoma, HD =Hodgkin's disease
Tested for EBV
Tested for both HHV-8 and EBV.
Determined by PCR;
Determined by in-situ hybridization
Histopathology: H & E sections of formalin fixed, paraffin embedded lymphoma tissues were classified according to the revised European-American (REAL) classification of lymphoid neoplasms16.
Immunohistochemistry: Cell immunophenotyping of the 22 tumors was performed as previously described17 on 4 – 6 µm frozen and formalin or Carnoy's fixed paraffin sections with various monoclonal antibodies (Mab) namely: - CD20 (clone L26; Dako, Denmark), CD3 (clone UCHT-1, Dako, Denmark), CD45RO (clone UCHL-1; Dako, Denmark), CD30 (clone BerH2; Dako, Denmark) and CD 68 (clone KP 1; Dako, Denmark).
After quenching the endogenous peroxidase activity and blocking for non-specific antibody binding17, the frozen sections were incubated with respective primary antibody for one hour at room temperature.
Paraffin embedded, formalin fixed sections were deparaffinized and processed for antigen retrieval as previously described18 in a microwave oven twice for 5 minutes at 750W. After washing in Tris-buffered saline (TBS), the sections were blocked for non-specific antibody binding at room temperature with normal horse serum (1:20) for 30 minutes (Vecta Stain) followed by incubation with primary antibodies overnight at 4°C.
Immunoperoxidase staining for HHV-8 LANA on antigen retrieved endogenous peroxidase and non-specific binding blocked lymphoma and control Kaposi's sarcoma sections, was done by incubating with Rat anti-LANA (ABI, Columbia, MD USA), overnight at 4°C. Bound antibody on frozen and paraffin sections was revealed by incubation with biotinylated goat anti-rat or anti-mouse (Jackson, West Groove, PA, USA) for 30 minutes at room temperature followed by an avidin-biotin-peroxidase complex (ABC) (Vectastain, PK-6100) and DAB (Sigma-Aldrich, St. Louis MO, USA) as the chromogenic substrate.
Immunofluorescence for LANA on control cultured BCBL-1 cytospins was performed by incubation with CY3-labelled goat anti-rat 1:50 (Jackson, West Groove, PA, USA) for 40 minutes at room temperature, washed in TBS and counter-stained with 5 mM 4′-6-Diamidino-2-phenylindole-HCL (DAPI) (Sigma-Aldrich, St Louis MO, USA) in 500 mM of trisodium citrate. After mounting, the slides were evaluated in a fluorescent microscope.
Incubation with TBS instead of respective primary antibody was used as a negative control for the detection system.
Polymerase chain reaction (PCR) analysis: DNA was extracted from 8 µm sections of formalin fixed or frozen biopsies as previously described7. For detection of HHV-8 sequences, primers (KS1, KS2) were chosen to amplify a 233-bp fragment of the KS-330 Bam sequence 5 using a PCR-PTC-200 machine as previously described6. Similarly specific sequences for EBV, HHV-6 and CMV DNA were also assayed by PCR with corresponding primers19. Positive controls included PCR on DNA from KS sections or cultured KS cells. Non-lymphoma (normal) tissue from the same individuals was not available and could not be included as a control for lymphoma specific association.
In situ hybridization (ISH): The presence of EBV-encoded RNA (EBER) was assayed by ISH with a FITC-labeled probe to EBER-I and II (Dako Y0017) and mouse anti-FITC (Dako M0878), rabbit anti-mouse serum and APAAP complexes as previously described2.
Results
Analysis of annual frequency and trends of ML from the cancer registry showed an overall increased frequency of ML in 1992 and 1994 (Figure I) and an increased frequency of AIDS related lymphoma (ARL) from 1993 to 1996. However, due to few cases registered at the registry and serologically tested for HIV, statistical evaluation was not conclusive.
Figure 1.
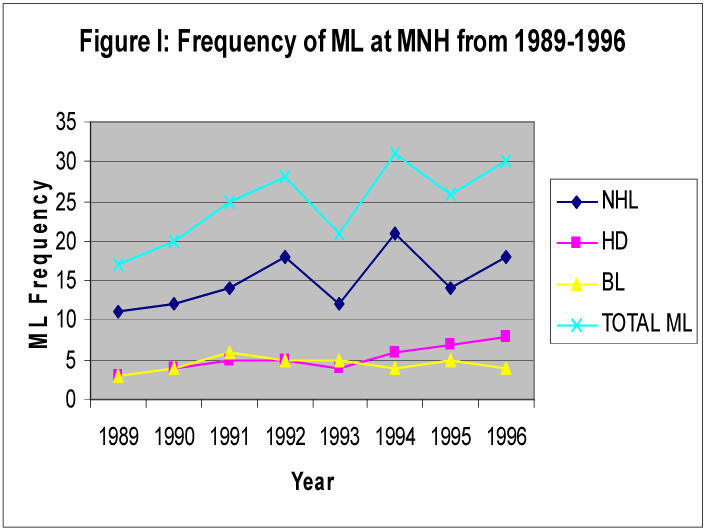
Annual frequency of malignant lymphoma (ML) from 1989-1996 in Tanzania.
The data for 22 cases of malignant lymphoma prospectively collected and further studied is shown on tables 1 and 2. The biopsies (M17, F5) from adults [≥ 15 years, (M 8; F 1)] and children (M 9; F 4) were further categorized as follows: sixteen 16 cases were seronegative for HIV, four (4) were seropositive (ARL) while (2) had no HIV serostatus. The HIV+ cases were mainly diffuse large B-cell lymphoma (DLBCL) and Hodgkin's disesase (HD). None of the six BL cases of children 5–9 years of age were associated with HIV.
Histopathology and Immunophenotype: The tumors were classified (Table 1) as: Burkitt's lymphoma (BL) (n=6), diffuse large B-cell lymphoma (DLB-CL) (n=10) (Figure 2a) precursor-B lymphoblastic (B-LBL) (n=1) and Hodgkin's disease (HD) (n=5). The HD cases were further sub-classified as mixed cellularity (MC-HD) (n=2) and lymphocyte depletion (LD-HD) (n=3). Two of ten (20%) DLB-CL and two of five (40%) LD-HD developed in HIV seropositive patients. All non-Hodgkin's lymphomas (NHL) were of B-cell lineage (Table 1), expressing CD20 antigen (Figure 2b). CD30+ Reed-Sternberg (RSC) and Hodgkin's cells were found in all HD cases. A variable number of infiltrating CD45RO+, CD3+ and CD68+ cells were seen in all type of tumors.
Figure 2a.
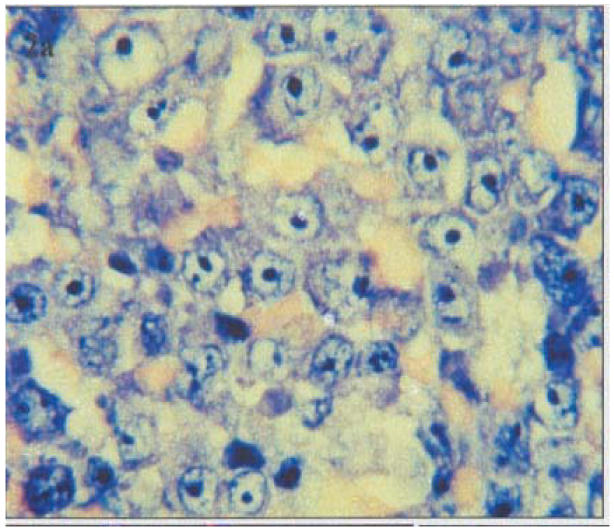
H & E stained section representative of diffuse large cell lymphoma (DLCL).
Figure 2b.
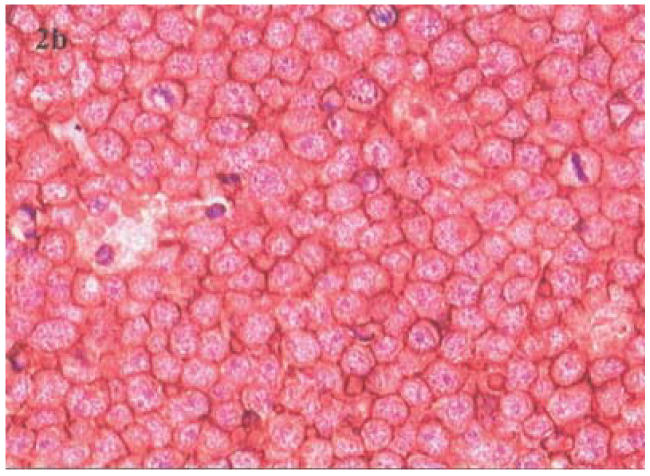
An immunoperoxidase stained paraffin section of the lymphoma in showing characteristic membrane localization for the B-cell CD 20 antigen.
All lymphoma cases were negative for HHV-8 as revealed by immunostaining for LANA, whereas, control BCBL-1 cells and Kaposi's sarcoma sections showed presence of HHV-8 LANA by immunofluorescence or immunoperoxidase (Figure 3 a–b).
Figure 3 (a).
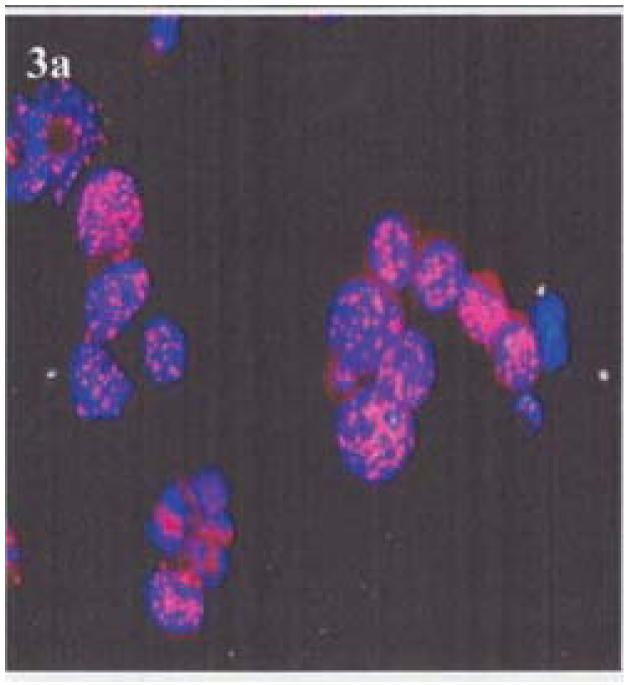
LANA Immunofluorescence in the nuclei of control BCBL-1 cells
Figure 3 (b).
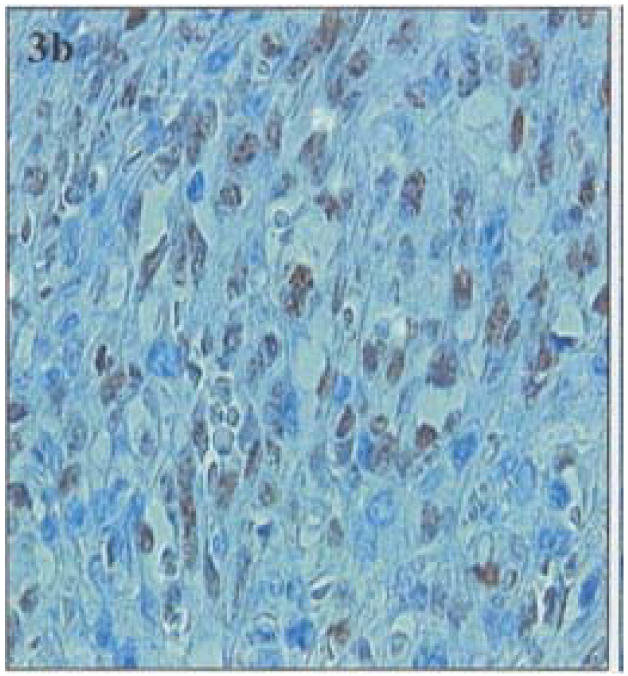
LANA positive cells in nodular KS control demonstrated by immunoperoxidase
In situ hybridization: All four tested Burkitt's lymphomas (Table 1&2) and five out of the eight tested DLB-CL (62.5%) (Table 1&2) had nuclear EBER reactivity in the tumor cells. Four (4/4) tested HD (100%) showed EBER hybridization within Reed-Stenberg (RSC) and Hodgkin's cells (HS) (Figure 4). Co-existence of HHV-8 (PCR) and EBER was seen in seven tumors (41%) (Table 2). Whereas most (13/17) tested lymphomas consistently expressed EBER-EBV (72%), only 45% (10/22) expressed HHV-8 by PCR (Table 2).
Figure 4.
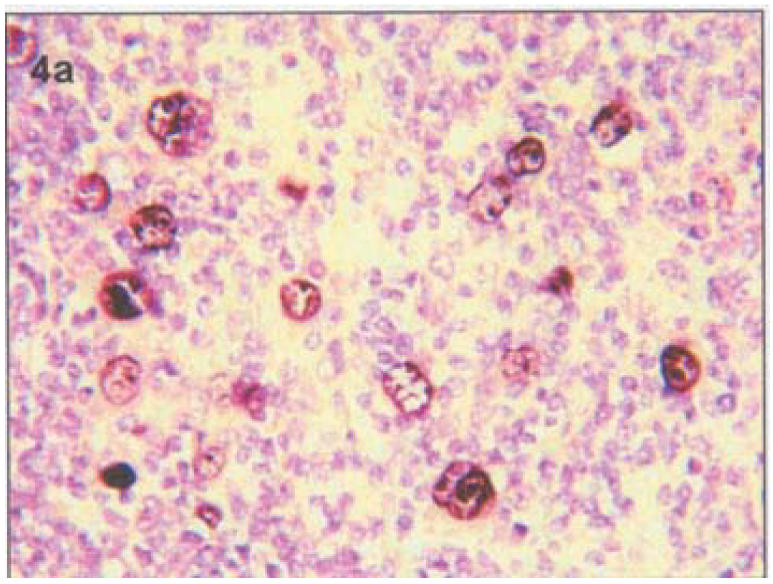
In in-situ hybridization (ISH) showing EBER localization at the inner nuclear membrane and nucleoli in HD
Polymerase chain reaction (PCR): HHV-8 DNA (Table 2) was detected in ten (10) of the twenty two (40.9%) malignant lymphomas all LANA negative by immunostaining. The positive cases were sub-classified into: BL 2/6 (33%), DLB-CL 4/10 (40%), B-LBL 1/1 and HD 3/5 (60%).
Consistent HHV-8 positivity was demonstrable with DNA from frozen biopsies whereas mostly negative results were seen with DNA extracted from biopsies after formalin fixation and paraffin embedding (Table 1). Comparison of PCR products from ML and KS showed a much stronger band in KS biopsies (Figure 5), which may suggest higher viral DNA content in KS compared to ML except in BCBL (not shown). All non-lymphoma or non- KS tissues from normal individuals were negative (not shown).
Figure 5.

PCR detection of KSHV/HHV-8 sequence in malignant lymphoma and Kaposi's sarcoma. The test band in each lane correspond to 233 bp, lane 1 (ladder), lane 2 DLC-BL, lane 3 BL, lanes 4, 5 and 6 DLB-CL, Lane 7 HD, lanes 8 and 9 EKS, lanes 10 and 11 AKS. Lane 12 is water. Note stronger bands in KS compared to lymphomas.
PCR for EBV DNA was concordant with the results (15/19 tested) of EBER in situ hybridization (Table 1). Seven (7/14) tumors had also demonstrable HHV-6 DNA by PCR. No CMV DNA was demonstrable by PCR in these seven tested tumors. The presence of HHV-8, EBV or HHV-6 in the tested material was not clearly related to age, sex, HIV serostatus or type of lymphoma. Three cases showed co-expression of all three herpes viruses (Table 1).
Discussion
In this study, the overall increase in registered malignant lymphoma (ML) from 1992 to 1994 appears to be an increase of AIDS-related lymphoma (ARL) since 1993. The apparent decline of ML/ARL after 1995 could not be statistically confirmed due to the relatively small number of registered cases and may be due to improper documentation of cases. The prevalence of ARL in Tanzania is difficult to ascertain since there is no routine screening of all lymphoma patients for HIV. However, the association between NHL and HIV seems, overall, weaker in Africa than in the more developed countries4. Different explanations have been put forward for this such as early acquisition of Epstein-Barr virus (EBV) in childhood in Africa with lower lymphoma risk if the virus is reactivated by immunosuppression21. A lower susceptibility to malignant lymphoma among African populations than among white populations has been hypothesized, since incidence rates were lower in black than white people of USA1. Under reporting of NHL and competitive mortality from other AIDS-associated illnesses may also cause this apparent lack of ARL since patients with severe immunodeficiency in Africa are likely to die from infectious diseases before manifesting ML22.
The recently described human herpes virus (KSHV/HHV-8) is closely related to herpes virus Saimiri (HVS) and also to Epstein-Barr virus (EBV)5 and correspondingly also lymphomagenic. Thus although first described in AIDS-related as well as in endemic and classical/sporadic Kaposi's sarcoma5, 6, 23, HHV-8/KSV was also shown to be associated with peripheral blood mononuclear cells (PBMC) of B-cell phenotype in AIDS patients with and without KS24, 25, and in lymphoid cells of other conditions such as body cavity based lymphoma (BCBL) a subset of ARL8, 11, 27–31. Given the historical endemic occurrence of KS and therefore HHV-8 infection in the Tanzania population it seemed appropriate to study the possible association of HHV-8 also with Tanzanian malignant lymphomas. Our present observations indicate a frequent (45%) presence of HHV-8 (by PCR) in various types of African malignant lymphomas including Hodgkin's disease, in HIV as well as non-HIV infected patients. However, HHV-8 cell association in these tumors was not proved by LANA immunostaining. These lymphomas were often also clonally infected with EBV (EBER+) but not in a strictly concordant way indicating the specificity of the PCR assays7.
Our findings clearly show that HHV-8 in ML patients is not related exclusively to HIV infection and does not segregate to any single type of B cell lymphoma but is also seen in HD cases. Outside Africa, HHV-8 has been shown to associate with multicentric Castleman's disease (MCD)29, angioimmunoblastic lymphadenopathy (AILD) and benign lymphadenopathy with follicular hyperplasia28, 30, 32. However, in another study of Spanish lymphoma HHV-8 was associated exclusively with BCBL3.
The present findings therefore could support the notion that HHV-8 is a lymphotropic virus6, 24, 31 but similar to EBV34 may also infect non-lymphoid cell types23, 35 and become activated not only due to HIV infection but also under conditions leading to EKS, suggesting a possible role for other non-HIV related co-factors in EKS evolution. However, quantitative differences in co-factors common to AKS and EKS cannot at present be excluded. Furthermore, our finding by PCR of much less HHV-8 DNA in ML than in KS indicates a lower viral load in ML possibly suggesting that HHV-8 infection may not be causally associated with ML since to be so, the virus should infect all or most cells of the tumor clone. The pathogenic role of HHV-8 in KS and the various types of lymphomas such as BCBL is still unclear but several “pirated” viral genes, homologous to cellular genes may be functionally expressed and thereby influence KS development15, 36, 37. However, the restricted HHV-8 association with various ML other than BCBL and MCD23 does not support a causal role for HHV-8 in ML in general and could indicate its presence as a passenger virus in tissues from individuals in high HHV-8 prevalence regions. This was clearly confirmed by failure to demonstrate HHV 8 in all lymphomas as demonstrated by negative anti-LANA immunostaining. Nevertheless, a pathognomonic role of HHV-8 in KS and BCBL tumorigenesis is well documented as also indicated by findings that anti-herpes viral treatment might induce long-term remission of KS38.
In conclusion the present study shows a slightly increased frequency of ML in Tanzania during the HIV epidemic and an association of HHV-8 by PCR with some ML cases of various types. However, no HHV-8 clonal association by immunohistochemistry for LANA was seen suggesting a passenger relationship with the lymphoma tissue. Thus although there is clear evidence for a pathogenic role of HHV-8 in AKS and EKS and primary effusion lymphoma (PEL, BCBL), association of HHV-8 with various ML in Tanzania only reflect a local tissue homing and activation of carrier cells in a high HHV-8 prevalent area.
This study, which to our knowledge is the first in Tanzania to show this type of relationship, clearly demonstrates that HHV-8 has no pathogenic role in the studied ML. This implies that measures for controlling HHV-8 such as vaccination will not eradicate ML. Control of EBV may however, be considered in special categories of ML. The findings calls for more studies to clearly elucidate the factors responsible for the low frequency of ML in Tanzania and Africa in general despite the ongoing HIV/AIDS epidemic.
Acknowledgments
This study was supported by SIDA through its Department for Research Cooperation (SAREC) (Ephata Kaaya) and Swedish Cancer Society (E. Castaños-Velez). The technical assistance of Joseph Lawrence, Vera Nelson and Angelika Magogo is highly appreciated.
References
- 1.Beral V, Peterman R, Jaffe H. AIDS-associated and non-Hodgkin's lymphoma. Lancet. 1991;937:805–809. doi: 10.1016/0140-6736(91)92513-2. [DOI] [PubMed] [Google Scholar]
- 2.Centers for Diseases Control, Revision of the case definition of acquired immunodeficiency syndrome for national reporting-USA. MMWR. 1985;34:373–375. [PubMed] [Google Scholar]
- 3.Lim ST, Levine AM. Recent advances in AIDS-related lymphoma. C A Cancer J Clin. 2005;55:229–264. doi: 10.3322/canjclin.55.4.229. [DOI] [PubMed] [Google Scholar]
- 4.Dal Maso L, Franceschi S. Epidemiology of non-Hodgkin's lymphomas and other haemolymphopoietic neoplasms in people with AIDS. Lancet Oncol. 2003;4:110–119. doi: 10.1016/s1470-2045(03)00983-5. [DOI] [PubMed] [Google Scholar]
- 5.Chang Y, Cesarman E, Pessin MS, et al. Identification of Herpes-virus-like DNA sequences in AIDS-associated Kaposi's sarcoma. Science. 1994;266:1865–1869. doi: 10.1126/science.7997879. [DOI] [PubMed] [Google Scholar]
- 6.Viejo-Borbolla A, Ottinger M, Schulz TF. HHV-8 biology and role in pathogenesis of KS and other AIDS-related malignancies. Curr HIV/AIDS Rep. 2004;1:5–11. doi: 10.1007/s11904-004-0001-3. [DOI] [PubMed] [Google Scholar]
- 7.Schalling M, Ekman M, Kaaya EE, Linde A, Biberfeld P. A role of a new herpes virus (KSHV) in different forms of Kaposi's sarcoma. Nature Medicine. 1995;1:707–702. doi: 10.1038/nm0795-707. [DOI] [PubMed] [Google Scholar]
- 8.Ansari MO, Dawson DB, Nador R, et al. Primary body cavity based AIDS-related lymphomas. Am J Clin Pathol. 1996;105:221–229. doi: 10.1093/ajcp/105.2.221. [DOI] [PubMed] [Google Scholar]
- 9.Cesarman E, Chang Y, Moore PS, Said JW, Knowles DM. Kaposi's sarcoma-associated herpes virus-like DNA sequences in AIDS-related body-cavity-based lymphomas. NEJM. 1995;332:1186–1191. doi: 10.1056/NEJM199505043321802. [DOI] [PubMed] [Google Scholar]
- 10.Carbone A, Gloghini A, Vaccher F, et al. KSHV/HHV-8 positive solid lymphomas: a tissue based variant of pleural effusion lymphoma. J Mol Diagn. 2005;7:17–27. doi: 10.1016/S1525-1578(10)60004-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Cesarman E, Moore PS, Rao PH, Inghirami G, Knowles DM, Chang Y. In vitro establishment and characterization of two acquired immunodeficiency syndrome-related lymphoma lines (BC-1 and BC-2) containing Kaposi's sarcoma-associated DNA sequences. Blood. 1995;86:2808–2714. [PubMed] [Google Scholar]
- 12.Gao S-JL, Kingsley M, Li W, et al. KSHV antibodies among Americans, Italians and Ugandans with and without Kaposi's sarcoma. Nature Medicine. 1996;2:925–928. doi: 10.1038/nm0896-925. [DOI] [PubMed] [Google Scholar]
- 13.Enbom M, Urassa W, Massambu C, Thorstensson R, Mhalu F, Linde A. Detection of human herpesvirus 8 DNA in serum from blood donors with HHV-8 antibodies indicates possible blood borne virus transmission. J Med Virol. 2002;68:264–267. doi: 10.1002/jmv.10183. [DOI] [PubMed] [Google Scholar]
- 14.Kedes DHE, Oberskalski E, Busch M, Kohn R, Flood J, Ganem D. The seroepidemiology of human herpesvirus 8 (Kaposi's sarcoma associated herpesvirus): distribution of infection in KS-risk groups and evidence for sexual transmission. Nature Medicine. 1996;2:918–924. doi: 10.1038/nm0896-918. [DOI] [PubMed] [Google Scholar]
- 15.Biberfeld P, Ensoli B, Stürzl M, Schulz T. KSHV/HHV-8, cytokines, growth factors and HIV in pathogenesis of Kaposi's sarcoma. Curt Opin Infect Dis. 1998;11:97–105. doi: 10.1097/00001432-199804000-00002. (1998) [DOI] [PubMed] [Google Scholar]
- 16.Harris NL, Jaffe ES, Stein H, et al. A revised European-American classification of lymphoid neoplasms: A proposal from the International lymphoma study group. Blood. 1994;84:1361–1392. [PubMed] [Google Scholar]
- 17.Kaaya EE, Parravicini C, Ordonez C, et al. Heterogeneity of the spindle cells in Kaposi's sarcoma: Comparison of cells in lesions and culture. JAIDS. 1995;10:295–305. [PubMed] [Google Scholar]
- 18.Shi S-R, Key ME, Kalra KL. Antigen retrieval in Formalin-fixed, paraffin-embedded tissues: An enhancement method for immunohistochemical staining based on microwave oven heating of tissue sections. J Histochem Cytochem. 1991;39:741–748. doi: 10.1177/39.6.1709656. [DOI] [PubMed] [Google Scholar]
- 19.Cinque P, Vago L, Dahl H, et al. Polymerase chain reaction on cerebrospinal fluid for diagnosis of virus-associated opportunistic diseases of the central nervous system in HIV-infected patients. AIDS. 1996;10:951–958. doi: 10.1097/00002030-199610090-00004. [DOI] [PubMed] [Google Scholar]
- 20.Rezikyan S, Kaaya EE, Ekman M, et al. B-cell lymphomagenesis in SIV-immunosuppressed cynomolgus monkeys. Intl J Cancer. 1995;61:574–579. doi: 10.1002/ijc.2910610423. [DOI] [PubMed] [Google Scholar]
- 21.Sitas F, Bezwoda WR, Levin V, et al. Association between human immunodeficiency virus type 1 infection and cancer in the black population of Johannesburg and Soweto, South Africa. Br J Cancer. 1997;75:1704–1707. doi: 10.1038/bjc.1997.290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Morgan D, Mahe C, Mayanja B, et al. HIV-1 infection in rural Africa: is there a difference in median time to AIDS and survival compared with that in industrialized countries? AIDS. 2002;16:597–603. doi: 10.1097/00002030-200203080-00011. [DOI] [PubMed] [Google Scholar]
- 23.Chang Y, Ziegler J, Wabinga H, et al. Kaposi's sarcoma-associated herpesvirus and Kaposi's sarcoma in Africa. Arch Intern Med. 1996;156:202–204. [PubMed] [Google Scholar]
- 24.Ambroziak JA, Blackbourn DJ, Herndier BG, et al. Herpes-like sequences in HIV-infected and uninfected Kaposi's sarcoma patients. Science. 1995;268:582–583. doi: 10.1126/science.7725108. [DOI] [PubMed] [Google Scholar]
- 25.Whitby D, Howard MR, Tenant-Flowers M, et al. Detection of Kaposi's sarcoma associated herpesvirus in peripheral blood of HIV-infected individuals and progression to Kaposi's sarcoma. Lancet. 1995;346:799–802. doi: 10.1016/s0140-6736(95)91619-9. [DOI] [PubMed] [Google Scholar]
- 26.Moore PS, Gao SJ, Dominguez G, et al. Primary Characterization of a herpes virus agent associated with Kaposi's sarcoma. J Virol. 1996;70:549–558. doi: 10.1128/jvi.70.1.549-558.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hermine O, Michel M, Buzyn-Veil A, Gessain A. Body-cavity-based lymphoma in an HIV-seronegative patient without Kaposi's sarcoma-associated herpes virus-like DNA sequences. NEJM. 1996;334:272–273. doi: 10.1056/NEJM199601253340417. [DOI] [PubMed] [Google Scholar]
- 28.Memar O M, Rady P L, Tyring S K. Human herpes virus-8: detection of novel herpes virus-like DNA sequences in Kaposi's sarcoma and other lesions. J Mol Med. 1995;73:603–609. doi: 10.1007/BF00196354. [DOI] [PubMed] [Google Scholar]
- 29.Soulier J, Grollet L, Oksenhendlar E, Cacoub P, Cazals-Hatem D, Sigaux F. Kaposi's sarcoma-associated DNA sequences in multicentric Castleman's disease. Blood. 1995;86:1276–1280. [PubMed] [Google Scholar]
- 30.Luppi M, Barozzi P, Maiorana A, et al. Human herpes-virus-8 DNA sequences in human immunodeficiency virus-negative angioimmunoblastic lymphadenopathy and benign lymphadenopathy with germinal centre hyperplasia and increased vascularity. Blood. 1996;87:3903–3909. [PubMed] [Google Scholar]
- 31.Carbone A, Gloghini A. AIDS-related lymphoma: from pathogenesis to pathology. Br J Haematol. 2005;130:662–670. doi: 10.1111/j.1365-2141.2005.05613.x. [DOI] [PubMed] [Google Scholar]
- 32.Malnati MS, Dagna L, Ponzoni M, Lusso P. Human herpesvirus 8 (HHV-8/KSHV) and hematologic malignancies. Rev Clin Exp Hematol. 2003;7:375–405. [PubMed] [Google Scholar]
- 33.Pastore C, Gloghini A, Volpe G, et al. Distribution of Kaposi's sarcoma herpesvirus sequences among lymphoid malignancies in Italy and Spain. B J Dermatol. 1995;91:918–920. doi: 10.1111/j.1365-2141.1995.tb05410.x. [DOI] [PubMed] [Google Scholar]
- 34.Greenspan JS, Greenspan D, Lennette, et al. Replication of Epstein-Barr virus within the epithelial cells of oral “hairy” leukoplakia, an AIDS-associated lesion. NEJM. 1985;313:1564–1571. doi: 10.1056/NEJM198512193132502. [DOI] [PubMed] [Google Scholar]
- 35.Boshoff C, Schulz TF, Kennedy MM, et al. Kaposi's sarcoma-associated herpes virus (KSHV) infects endothelial and spindle cells. Nature Medicine. 1995;1:1274–1278. doi: 10.1038/nm1295-1274. [DOI] [PubMed] [Google Scholar]
- 36.Nicholas J, Zong JC, Alcendor DJ, et al. Novel organizational features, captured cellular genes and strain variability within the genome of KSHV/HHV-8. J Natl Cancer Inst. 1998;23:79–88. doi: 10.1093/oxfordjournals.jncimonographs.a024179. [DOI] [PubMed] [Google Scholar]
- 37.Zhong J-C, Metroka C, Reitz MS, Nicholas J, Hayward G. Strain variability among Kaposi's sarcoma-Associated herpes virus (Human Herpesvirus 8) Genomes: Evidence that a large cohort of United States AIDS patients may have been infected by a single common isolate. J of Virology. 1997;71:2505–2511. doi: 10.1128/jvi.71.3.2505-2511.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Morfeldt L, Torssander J. Long-term remission of Kaposi's sarcoma following foscarnet treatment in HIV-infected patients. Scand J Infect Dis. 1994;26:749–752. doi: 10.3109/00365549409008645. [DOI] [PubMed] [Google Scholar]