

Abstract
The biogenesis and accumulation of the beta amyloid protein (Aβ) is a key event in the cascade of oxidative and inflammatory processes that characterises Alzheimer’s disease. The presenilins and its interacting proteins play a pivotal role in the generation of Aβ from the amyloid precursor protein (APP). In particular, three proteins (nicastrin, aph-1 and pen-2) interact with presenilins to form a large multi-subunit enzymatic complex (γ-secretase) that cleaves APP to generate Aβ. Reconstitution studies in yeast and insect cells have provided strong evidence that these four proteins are the major components of the γ-secretase enzyme. Current research is directed at elucidating the roles that each of these protein play in the function of this enzyme. In addition, a number of presenilin interacting proteins that are not components of γ-secretase play important roles in modulating Aβ production. This review will discuss the components of the γ-secretase complex and the role of presenilin interacting proteins on γ-secretase activity.
Keywords: Alzheimer’s disease, Amyloid precursor protein, Beta amyloid, Presenilin, Interacting proteins, Gamma secretase
Introduction: the generation of beta amyloid by presenilin and its binding proteins
Alzheimer’s disease (AD) is a complex, progressive neurodegenerative disorder that is neuropathologically characterised by extensive neuronal loss and the presence of neurofibrillary tangles and senile plaques. While the majority of AD cases are sporadic, ∼5% of AD cases are familial (FAD) with mutations in three genes, amyloid precursor protein, presenilin 1 (PS1) and presenilin 2 (PS2) accounting for the majority of cases. A major feature of both sporadic and familial forms of AD, is the accumulation and deposition of a small peptide referred to as beta amyloid (Aβ) within brain tissue of AD sufferers. The mechanisms that underlie the disease processes are poorly understood. However, the accumulation of Aβ is thought to play a pivotal role in neuronal loss or dysfunction through a cascade of events that include the generation of free radicals, mitochondrial oxidative damage and inflammatory processes (reviewed in Refs. [1, 2]). The primary event that results in the abnormal accumulation of Aβ is thought to be the dysregulated proteolytic processing of its parent molecule, the amyloid precursor protein (APP).
The APP molecule is a transmembrane glycoprotein that is proteolytically processed by two competing pathways, the non-amyloidogenic and amyloidogenic (Aβ forming) pathways. How these pathways are regulated remains unclear. However, there are many factors including diet, hormonal status, and genetic mutations that influence the processing of APP to generate Aβ (reviewed in Refs. [1, 3]). Three major secretases are postulated to be involved in the proteolytic cleavage of APP. These include α-secretase (of which the metalloproteases ADAM17/TACE and ADAM 10 are likely candidates), beta APP cleaving enzyme (BACE, formally known as β-secretase) and the γ-secretase. The α-secretase cleaves within the Aβ domain of APP thus precluding the formation Aβ and generating non-amyloidogenic fragments and a secreted form of APP (α-APPs). In the amyloidogenic pathway (Fig. 1), BACE cleaves near the N-terminus of the Aβ domain on the APP molecule, liberating another soluble form of APP, β-APPs and a C-terminal fragment (C99) containing the whole Aβ domain. The final step in the amyloidogenic pathway is the intramembranous cleavage of the C99 fragment by γ-secretase, to liberate the Aβ peptide (reviewed in Ref. [1]). Identification of multiple cleavage sites within the APP molecule has suggested the presence of other secretases, namely ε- and ζ-secretase; that generate the APP intracellular domain (AICD) and a longer fragment of Aβ (Aβ46), respectively [4–7]. Although different enzymes may exists, the same γ-secretase complex that generates Aβ may also generate AICD and Aβ46 as these products are inhibited by γ-secretase inhibitors [4–7] indicating that this enzyme has multiple cleavage sites. However, over-expression and mutagenesis studies have shown that the production of Aβ and AICD may be independent catalytic events [8, 9].
Fig 1.
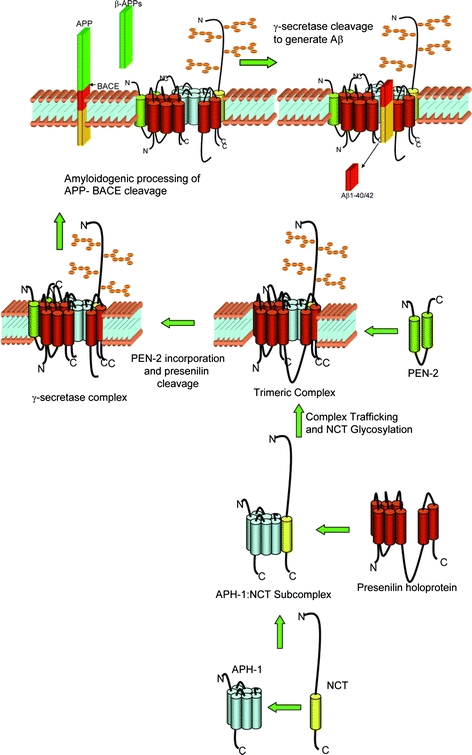
Formation of the γ-secretase complex and Aβ generation. A schematic representation of the proposed formation of the γ-secretase complex and the enzymatic activity resulting in Aβ generation. Evidence has been provided to show that PS, nicastrin (NCT), aph-1, and pen-2 are the only components of the γ-secretase complex. Nicastrin and aph-1 are involved in the early formation of the complex, interacting with each other and forming a dimeric sub-complex. The presenilin holoprotein is incorporated into the aph-1: NCT sub-complex, nicastrin undergoes post-translational modifications and the complex is transported to the cell surface (or other Aβ generating compartments, i.e. TGN) as a trimeric complex. Pen-2 is incorporated into the complex and presenilin is cleaved forming the active γ-secretase complex. Following cleavage of APP by BACE to liberate secreted APP, the APP-C99 fragment left embedded within the membrane undergoes regulated intramembranous processing (RIP) by the γ-secretase complex, liberating Aβ1–40 or Aβ1–42 (Modified from Ref. [165])
The γ-secretase enzyme is thought to be an aspartyl protease that has the unusual ability to regulate intramembrane proteolysis (RIP) for a growing list of type 1 integral membrane proteins which include, APP, APP like proteins (APLPs), E-Cadherin, CD44, lipoprotein receptor related protein (LRP), Notch, sterol regulatory element-binding protein (SREBP), interferon response element (IRE1) and activated transcription factor 6 (ATF-6) (for recent review see [10]). The mechanism of RIP and γ-secretase activity is unknown. However a transient hydrophilic environment for catalysis within the lipid membrane must be created. Furthermore, the enzyme (or an enzyme domain) must have the ability to bend and unwind the α-helical substrates, exposing their amide bonds to hydrolysis. It is therefore conceivable that the γ-secretase enzyme is an integral protein of the lipid bi-layer and contains a number of proteins that may have different functions within an enzyme complex. The exact conformation or molecular architecture of the γ-secretase enzyme remains unknown. However, large molecular mass complexes of ∼250 kDa [11, 12], ∼500 kDa [13–17] and ∼2,000 kDa [13, 18] have been identified.
Over the last few years biochemical and genetic approaches have identified four components of the γ-secretase complex, presenilins, nicastrin, anterior pharynx defective (aph-1) and presenilin enhancer 2 (pen-2). Over-expression and expression knockdown studies have provided strong evidence that these proteins are essential for γ-secretase activity [15, 19–24]. Subsequent reconstitution studies in non-mammalian cells have provided evidence that these are the only components responsible for γ-secretase catalytic activity [25–27]. However the active component and the active catalytic site of this enzymatic complex are yet to be identified.
The roles of presenilin and its binding partners within the γ-secretase complex in Aβ generation
Presenilin 1: the catalytic component of the γ-secretase enzyme
Presenilin 1 (PS1) is thought to be the most critical component of the γ-secretase complex as evidence indicates that it is the catalytic component of this proteolytic enzyme. The finding that mutations in PS1 account for the majority of inherited early onset forms of AD and result in the overproduction of the highly amyloidogenic Aβ42 [28–30] provided the initial evidence that PS1 facilitates γ-secretase activity. Subsequent studies showed that PS1 ablation or mutagenesis of two highly conserved aspartate residues within transmembrane domains 6 and 7 of PS1 results in a reduction in Aβ levels [31–34]. Furthermore, aspartyl protease inhibitors and transition state analogue inhibitors which are designed to target the active site of the protease, all reduce Aβ40 and Aβ42 levels and have been shown to affinity label and bind to PS1 [35–37]. In addition, physical interactions between PS1 and the APP-C99 fragment have been identified [38–40]. The studies outlined above provide substantial evidence to indicate that PS1 is the catalytic subunit of γ-secretase. However, this transmembrane protein is not typical of a conventional aspartyl protease, and lacks the typical D(T/S)G motif for an aspartyl protease active site (recently reviewed in [41]. Aspartyl proteases require the presence of two aspartate residues for the hydrolysis of the peptide bond. Although PS1 lacks the typical D(T/S)G aspartyl protease active site it does contain the two aspartyl residues (D257 and D385) which are either critical for the active site on the γ-secretase complex or constitute the active site [42]. Other proteases that contain atypical aspartyl protease motifs and have weak homology with PS1 include the bacterial membrane embedded aspartyl protease, type 4 prepilin protease and the signal peptidases, which are all transmembrane proteins. These proteins share the unusual aspartyl motif GxGD and perform intramembranous cleavage of type I membrane proteins (reviewed in [43]). The topology and atypical characteristics of these proteins make them an unprecedented class of protease and as is the case for PS1, would require interaction with of other proteins within the complex.
The interaction between the components of the γ-secretase complex is not sufficient to create a macromolecular enzyme complex such as γ-secretase (i.e. total mass of all the components is ∼200 kDa). Thus, dimer or trimer formation must occur within the complex. To date the only member of the complex that is known to undergo this process is PS1. The full length PS1 protein is rapidly endoproteolytically cleaved within its characteristic large hydrophilic loop into amino- and carboxy-terminal fragments (NTF/CTF) of ∼27 and ∼17 kDa, respectively [44, 45]. These fragments are thought to interact with each other to form the catalytic component of the γ-secretase enzyme since in the absence of full length PS1, co-expressed NTF and CTF were able to generate γ-secretase activity in presenilin deficient mammalian cells [46, 47]. The stoichiometry and the nature of the interaction between these fragments remain unclear. It has been shown by many studies that the NTF:CTF form a heterodimer in mammalian cells [12, 48–51] leading to suggestions that this heterodimer is the active γ-secretase [52]. However, Cervantes and colleagues [53] provided evidence that the presenilin fragments can form a tetramer by identifying heterodimers as well as NTF and CTF homodimers in yeast. Evidence for heterodimer and NTF homodimer (but not CTF homodimer) formation has been provided by photo-affinity labelled cross-linking studies [54]. This formation provides a core of aspartyl residues required for aspartyl protease activity. However, it has yet to be established whether the hypothetical “core” is formed between fragments from one PS1 molecule or multiple molecules within the complex. Further, if PS1 is the “catalytic core” of the γ-secretase complex the roles of the other interacting proteins need to be determined.
Nicastrin: the substrate docking site and scaffolding for building the γ-secretase complex
Nicastrin was discovered to be part of the γ-secretase complex when it was found to co-immunoprecipitate with PS1 [55]. Nicastrin is a type I transmembrane protein with a 670 amino acid long hydrophilic N-terminal domain, a transmembrane domain, and a relatively short cytoplasmic C-terminus of twenty amino acid residues [55]. The protein undergoes glycosylation and sialylation within the secretory pathway to yield a mature ∼150 kDa protein, the largest component of the γ-secretase complex [14], [56–58]. Nicastrin has been shown to be a critical component of the γ-secretase complex as inhibition of nicastrin function in vitro and in vivo results in a complete loss of APP and Notch cleavage [14, 55, 59–61]. Within this complex, nicastrin appears to be involved in the early formation of the complex, particularly in the formation of the first sub-complex between itself and aph-1 (Fig. 1). Recent evidence suggests that nicastrin is critical for the correct assembly of the γ-secretase complex within the endoplasmic reticulum and the intracellular trafficking of the complex to the cell surface [62, 63]. Furthermore, nicastrin is essential for the interaction between the complex and APP-C99 and thus may act as a receptor for γ-secretase substrates [63, 64]. Therefore, although nicastrin is a critical component, evidence to date suggests that it is unlikely to be a catalytic component of the γ-secretase complex. Instead, nicastrin acts as a scaffold for the building of the active complex, and a possible substrate docking site.
Anterior pharynx defective 1 (aph-1): scaffolding partner for nicastrin and may have possible catalytic activity
The aph-1 protein is a ∼30 kDa, 308 amino acid long 7-transmembrane protein [65] existing in two homologous forms, located on chromosome 1 and chromosome 15 (aph-1a & aph-1b respectively; [19]). In addition, as the result of alternate splicing, a long and short isoform of aph-1a have also been reported, with the short isoform more abundantly expressed in most tissues [21]. Although the exact function of these isoforms remains unknown, multiple γ-secretase complexes containing aph-1 isoforms are thought to exist [66, 67]. The aph-1a isoform appears to be incorporated into active γ-secretase complexes. Deletion of aph-1a causes a reduction in Aβ generation [66] and is the major isoform present in γ-secretase complexes during embryonic development [67]. The γ-secretase complexes incorporating aph-1b are thought to have a redundant function as aph-1b has been found not to be involved in the regulation of other γ-secretase components or γ-secretase activity [66].
The aph-1 protein forms a stable and intermediate ∼140 kDa complex with Nicastrin, before binding PS1. Aph-1 has been shown to interact with immature and mature forms of presenilins and nicastrin [21]. The GXXXG motif located within transmembrane 4 of aph-1 has been shown to be important in this interaction [68], [69]. The aph-1: nicastrin sub-complex appears to play a role in the stabilisation and assembly of PS1 in the γ-secretase complex (Fig 1; [24, 70, 68]). However, the exact role of aph-1 in the γ-secretase complex remains unknown. Expression studies have shown that removal of aph-1 destabilises the presenilin holoprotein and the addition of aph-1 leads to the accumulation of the holoprotein [11, 19–24, 71]. Thus together with nicastrin, aph-1 is thought to contribute the initial formation and trafficking of the γ-secretase complex and thereby provide a scaffold for the proteolytic processing of PS1 and the formation of the active site.
Aph-1 is critical for the activity of the fully constructed γ-secretase complex. Evidence has been provided that as well as binding to immature components of γ-secretase in early stages of complex formation, aph-1 also interacts with the mature forms of PS1, nicastrin and pen-2 [68, 72]. Furthermore, recent evidence has been provided to suggest that this interaction occurs on the cell surface where it also binds the γ-secretase substrate, Notch and facilitates its cleavage [72]. These studies suggest that apart from the scaffolding function it shares with nicastrin, aph-1 has another function within the fully constructed, active γ-secretase complex. Structural and functional similarities between aph-1 and other proteases that possess the ability for intramembranous cleavage (such as rhomboid, [68, 73] suggests that this transmembrane protein may have an enzymatic function within the complex. This postulated novel function for aph-1 needs to be validated by future studies.
Presenilin Enhancer 2 (pen-2): the elusive presenilinase or “molecular clamp” that holds the complex together?
The pen-2 protein is a ∼12 kDa, 101 amino acid long protein containing two transmembrane domains and its N- and C-terminals facing the lumen of the endoplasmic reticulum [74, 75]. Pen-2 has been shown to interact with Nicastrin, aph-1, and also PS1. Without this interaction pen-2 has been shown to be degraded, possibly by the proteasome [76, 77]. Ablation of pen-2 results in significantly reduced PS1 endoproteolysis and Aβ production, suggesting that pen-2 is essential for both presenilinase and γ-secretase activity [15, 19, 22–24]. Likewise, combinational expression studies have shown that when pen-2 is co-expressed with the other components of the γ-secretase complex in a step-wise manner, endoproteolysis of the PS1 holoprotein was enhanced [11, 22, 24–27, 71]. A trimeric sub-complex between presenilin, nicastrin and aph-1 has been identified in cells lacking pen-2 [24, 70] suggesting that pen-2 may be the final protein incorporated into the γ-secretase complex (Fig. 1). Recent evidence has suggested that pen-2 is incorporated into the γ-secretase complex through an interaction with presenilins. The “DYLSF” domain of pen-2 and a “NF” motif on transmembrane 4 of PS1 have been shown to be critical for the interaction between these proteins [78–80]. The cleavage of PS1 occurs within the hydrophilic loop and is distal from the interaction site on transmembrane 4, suggesting that another domain of pen-2 is responsible for the cleavage of PS1 or pen-2 may present PS1 to the elusive “presenilinase” which may be another member of the γ-secretase complex. The C-terminal end of pen-2 has been shown to be important for γ-secretase activity since altering the length of this region by addition or deletion of residues has been shown to reduce Aβ40 and 42 generation without altering the binding of pen-2 to the complex [78] suggesting that pen-2 may have an alternative function within the complex. One suggestion put forward by Hasegawa and colleagues [78] is that the C-terminus of pen-2 may be a linker/space molecule that maintains spatial interactions between proteins within the complex. However, recent evidence using pen-2 C-terminal loss of function mutations suggest that the C-terminus acts as a “molecular clamp” holding together the presenilin fragments and the whole γ-secretase complex [81]. If this is the case then PEN-2 is an integral part of the catalytic process holding the complex together whilst the γ-secretase products are generated.
The newest member of the complex: TMP21, a modulator of γ-secretase activity
Recently another member of the γ-secretase complex was immuno-purified from wild-type PS1 and PS2 blastocyte derived cells [82]. The type 1 transmembrane protein termed TMP21 is a member of the p24 cargo-family which may have a signalling role in the sorting and transport of proteins from the endoplasmic reticulum to the Golgi [83, 84]. It was shown by Chen and colleagues [82] that TMP21 is a member of the complex as it was isolated in a high molecular weight presenilin complex, interact with all of the known components of the γ-secretase complex, co-localised with the complex components in the ER, Golgi and cell surface and destabilised from the complex in the absence of the presenilins and pen-2. Although the over-expression of TMP21 did not alter γ-secretase activity, its suppression resulted in and increase in Aβ40 and Aβ42. However, suppression of TMP21 did not alter the production of AICD, notch cleavage to generate NICD or cleavage of E-cadherin to generate CICD. These results are consistent with the notion that γ- and ε-secretase cleavage activities are independently regulated and indicate a role for TMP21 in modulating γ-secretase activity to generate Aβ. This role for TMP21 appears to be independent of its role in protein transport since the suppression of both TMP21 and p24a (a member of the p24 cargo family that interacts with TMP21) does not result in additional increases in Aβ production to that observed following the suppression of TMP21 only [82]. This finding led the authors to postulate that there are two pools of TMP21, a major pool that is stabilised by, p24a and has no role in Aβ production and a minor pool that modulates Aβ production, independent of p24a. Although, further investigation is required to determine the precise mechanism of action, it appears that TMP21 may function to regulate intramembrane proteolysis controlling γ-secretase activity and thus preventing the over-production of Aβ. This modulator role for TMP21 is important to consider when elucidating mechanisms on how γ-secretase activity is altered (for example by presenilin mutations) resulting in enhanced Aβ40 and Aβ42 production.
Presenilin 2 (PS2): capable of forming a fully functional γ-secretase complex
The presenilin proteins share an overall homology of 67% with the highest similarities in the predicted transmembrane domains. Fewer similarities are found at the N-terminus and in the central region of the hydrophilic loop [85, 86]. Although there are some structural similarities between PS1 and PS2 several lines of evidence suggest that these proteins may have distinct functions. Mice lacking PS1 die before birth and have severe skeletal and brain deformities, whilst those lacking PS2 only develop a mild pulmonary fibrosis and haemorrhage with age [33, 87–90]. Compared to neuronal cultures isolated from PS1 ablated mice, those isolated from PS2 knockout mice exhibit higher Aβ production and are less sensitive to γ-secretase inhibitors [89, 91]. Lai and colleagues [91] also provided evidence to suggest that distinct PS1 containing and PS2 containing complexes exist [91]. If this is the case, evidence to date suggests that PS2 containing complexes have different functions and have less γ-secretase processing power than PS1 containing complexes. However, recent in vitro evidence has suggested that chimeras of PS1 and PS2 fragments retain γ-secretase activity [92] suggesting that active complexes containing both PS1 and PS2 could occur. However, this is unlikely to occur in vivo as PS2 and PS1 transgenic mice have differential effects on γ-secretase activity [93]. In this study mice lacking PS1 were crossbred with transgenic mice containing wild type or mutant PS2/PS1. Compared to mice harbouring wild-type PS1, those harbouring wild-type PS2 had a ∼4-fold reduction in Aβ40 and ∼2-fold reduction Aβ42 brain levels. In addition, the authors reported that although mutations in PS1 lead to higher brain levels of Aβ40 and Aβ42 than PS2 mutations, those mice harbouring PS2 mutations had higher brain levels of Aβ42, whilst Aβ40 brain levels remain unchanged [93]. Taken together these studies suggest that PS1 containing complexes predominate in generating Aβ40 and Aβ42. However, the finding that PS2 is involved in Aβ42 (but not Aβ40) generation suggests that two Aβ generating γ-secretase complexes (γ-40 secretase and γ-42 secretase) exist, with γ-42 secretase complexes containing both PS1 and PS2.
The roles of presenilin binding proteins not incorporated into the active γ-secretase complex
In addition to Aβ generation, the presenilins have been implicated in a variety of intracellular processes including membrane trafficking, Notch signalling, neuronal plasticity, cell adhesion, regulation of calcium homeostasis, the unfolded protein response, and apoptosis. Therefore it is not surprising that the presenilins have a growing list of binding partners (see Table 1). Though the presenilin binding proteins that are incorporated into the γ-secretase complex are critical for the activity of this enzyme, those that are not integral components have equally important roles in modulating Aβ production. A number of non-complex presenilin binding proteins have been shown to alter γ-secretase activity (see Table 1). However, only a few have had supporting evidence that their direct interaction with the presenilins modulate γ-secretase activity. These are described below and shown in Fig. 2.
Table 1.
Presenilin interacting proteins
| Interacting protein | Proposed function | Interaction with presenilins shown to modulate Aβ production | Reference |
|---|---|---|---|
| Complex components | |||
| APH-1 | Component of the γ-secretase complex; role for initial formation of the complex, or a proteolytic role | Yes | [19, 65, 67, 123]. |
| Nicastrin | Component of γ-secretase complex; possible role as a receptor for APP-C99 | Yes | [14, 55–58]. |
| PEN-2 | Component of the γ-secretase complex, role in proteolytic cleavage of the presenilins | Yes | [19, 24, 70, 76, 77]. |
| TMP21 | Recently identified component of the γ-secretase complex, role in modulating Aβ production | Yes | [82] |
| γ-secretase substrates | |||
| APP-C99 | Substrate for the γ-secretase complex: precursor to Aβ generation | Yes | [38–40] |
| E/N cadherin | γ-secretase substrate; role in Cell-Cell Adhesion | No | [124] |
| IRE1 | γ-secretase substrate; role in unfolded protein response | No | [125, 126] |
| LRP | γ-secretase substrate; Lipid metabolism | Yes | [119] |
| Notch1 | γ-secretase substrate; precursor to NICD generation | Yes | [112] |
| Interacting binding proteins | |||
| ABP-280, Filamin homolog1 | Actin binding protein | No | [127] |
| Bcl-2 | Anti-apoptotic molecule | No | [128] |
| Bcl-XL | Anti-apoptotic molecule | No | [129] |
| Calmyrin | Possible calcium-myrstol switch | No | [130] |
| CALP- calsenilin like protein | Novel member of the calsenilin/KChIp protein family | No | [131] |
| Calsenilin | Calcium binding protein | Yes | [94, 96–99, 101, 102] |
| CLIP-170 | Linking membrane organelles to microtubules | No | [132] |
| DRAL | LIM-domain containing protein | No | [133] |
| FHL2 | Role in PI3K/Akt activation | No | [134] |
| FKBP38 | Role in mitochondria mediated apoptosis | No | [135] |
| Go | Signalling molecule | No | [136] |
| GSK3β | Wnt signalling, serine threonine protein kinase | No | [137] |
| HC5/ZETA | Subunits of the catalytic 20S proteasome | Yes | [138] |
| Met1 | Putative methyltransferase | No | [139] |
| Modifier of cell adhesion protein (MOCA) | Regulates proteasomal activity on APP | Yes | [140] |
| Omi/HtrA2 | Serine protease, proapoptotic | No | [141] |
| PAG | Neuronal proliferation protein | No | [142] |
| PAMP and PARL | Metalloproteases | No | [143] |
| PKA | Serine/Theronine protein kinase; β-catenin phosphorylation | No | [144] |
| Plakoglobin | Role in b-catenin/Tcf-4 activity | No | [145] |
| PLD1 | Phospholipid-modifying enzyme | Yes | [146] |
| PSAP | PDZ like protein | No | [147] |
| QM/Jif1 | Negative regulator of c-Jun | No | [148] |
| Rab proteins | Vesicle mediated protein trafficking | Yes | [149, 150] |
| RyR2 | Cardiac ryanodine receptor | No | [151] |
| SEL-10 | Ubiquitination of proteins | Yes | [152] |
| Sorcin | Calcium binding regulator of ryanodine receptor | No | [153] |
| Syntaxin 1A | Synaptic plasma-membrane protein | No | [154] |
| Syntaxin 5 | ER-Golgi vesicular transport | Yes | [155, 156] |
| Tau | Microtubule binding protein | No | [137] |
| Telencephalin | Neuron specific adhesion molecule | No | [40] |
| TPIP | Tetratricopeptide repeat-containing protein | No | [157] |
| Ubiquilin | Ubiquitin domain-containing protein | No | [158–160] |
| X11 family of proteins | Cytoplasmic adaptor proteins | Yes | [110, 111] |
| β-catenin/δ-catenin (NPARP) | Wnt signalling. Cell adhesion | No | [161–163] |
| μ-Calpain | Calcium-dependent thiol protease | No | [164] |
Fig. 2.
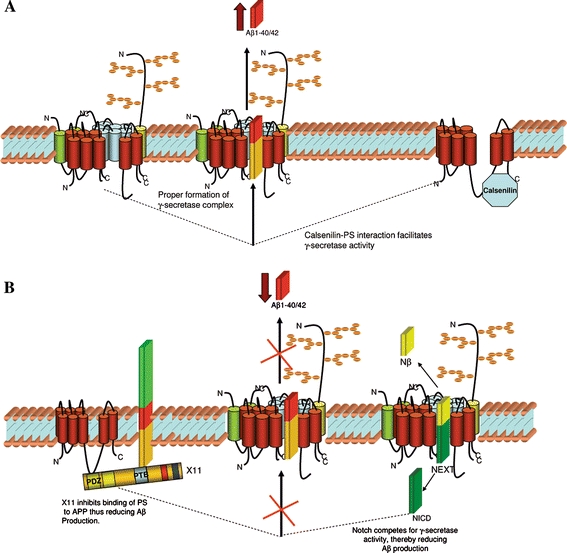
The postulated roles for presenilin interacting proteins not incorporated into the γ-secretase complex in Aβ generation. A schematic representation of the postulated roles of presenilin interacting proteins in (A) promoting or (B) attenuating γ-secretase activity and Aβ generation. (A) Together with the formation of the γ-secretase complex, calsenilin have been shown to interact with the presenilins and promote Aβ generation when over-expressed in mammalian cells. The role of calsenilin in Aβ generation remains unclear but maybe involved in the presentation of the substrate to the γ-secretase complex. (B) The presenilin interacting protein, X11 and other γ-secretase substrates that compete for enzyme activity such as Notch fragments (i.e. NEXT) have all been shown to reduce Aβ generation when over-expressed in mammalian cells. X11 may reduce Aβ production by binding to PS and APP preventing PS mediated cleavage of APP or its presentation to the γ-secretase complex
Calsenilin
Calsenilin (also called DREAM or kChIP3) is a calcium binding protein, that was first identified through yeast two hybrid screening using the PS2 C-terminus as the bait protein [94]. The interaction between calsenilin and the presenilins appear to have a role in APP processing. It was initially demonstrated that calsenilin can form a complex with PS1 and PS2 and regulate presenilin fragment formation in mammalian cells [95]. This study also showed that calsenilin may facilitate presenilin mediated apoptosis, since calsenilin was shown to preferentially interact with the C-terminal fragment of PS2 that results from caspase cleavage of the presenilin holoprotein [96]. Expression studies in vivo (Xenopus) and in vitro (mammalian cells) have shown that the interaction between presenilin and calsenilin may be involved in ER calcium release and apoptosis [97, 98]. Over-expressing calsenilin in neuronal cells resulted in cell death which was enhanced by the co-expression of wild-type or PS1 mutations [99]. These studies have suggested that the interaction between calsenilin and presenilins may function in apoptotic pathways. However, several lines of evidence suggest that calsenilin may also be implicated in APP processing and Aβ generation. Initial in vitro studies showed that over-expression of calsenilin in mouse neuroblastoma cells resulted in a gene dose dependent accumulation of Aβ42 [100]. Using a γ-secretase luciferase based assay and a cell free system established by Li and colleagues [18] to characterise the γ-secretase complex in purified cell membranes, Jo and colleagues [101] recently reported that the over-expression of calsenilin resulted in an increase in γ-secretase activity. This calsenilin/presenilin, mediated activity was markedly reduced using a well-characterised γ-secretase inhibitor (L-685, 458). Furthermore, compared to wild-type mice, brain Aβ40 and Aβ42 levels were reduced in calsenilin ablated mice [102]. Together these studies would suggest that calsenilin has a second function, to facilitate γ-secretase activity.
X11/Mint proteins
A family of cytoplasmic adaptor proteins called X11 (also known as munc-18-interacting proteins—mints) have also shown to have a direct role in APP processing. In particular, several studies have reported that X11α and X11β stabilise APP and inhibit the secretion of Aβ and APP from neuronal cells [103–106]. The molecular mechanisms by which the X11s influence APP processing and Aβ production are not clear. However, this effect appears to be modulated by a direct interaction between X11 and APP. The interaction between the phosphotyrosine binding (PTB) domain within X-11 and the YENPTY motif within APP appears to modulate Aβ generation, since introduction of point mutations within these domains enhanced Aβ generation [103, 106–109]. The binding of X11 appears to stabilise APP, leading to its intracellular accumulation [103, 106–109]. It is conceivable that the X11–APP interaction may inhibit secretase accessibility to APP resulting in impaired Aβ secretion. The X11 proteins have multiple binding domains and have been shown to interact with the PS1 C-terminal fragment through their PDZ domain [110]. The authors also showed an interaction between PS1 and APP in the presence of X11 that was otherwise ablated in the absence of X11. This suggests that an interaction between APP and PS1 is mediated through X11. A subsequent study has reported that X11β and alcadein (and X11β-binding protein) expression prevents the binding of PS1 to APP [111]. The significance of the results from these interaction studies remains unclear. However, they do suggest that X11 proteins may act as inhibitory molecules preventing PS1 mediated proteolytic processing of APP and thus attenuating Aβ production.
Competing substrates
Some presenilin binding proteins reduce the likelihood of Aβ generation by competing for the catalytic activity offered by γ-secretase. One example is Notch1 which undergoes cleavage by γ-secretase to liberate an intracellular domain (NICD) that translocates to the nucleus where it co-activates transcription of genes involved in several developmental pathways [112–115]. The first evidence of substrate competition came from one study that treated neurons with the Noch1 ligand, delta, and found a dose dependent reduction in Aβ levels [116]. In contrast, the over-expression of APP in these neurons resulted in a reduction in Notch signalling [116]. Further evidence for substrate competition was provided by Kimberley and colleagues [117] where APP and Notch based substrates directly competed for γ-secretase activity in a cell free assay. Similar results were achieved in a subsequent study where cells expressing Notch showed a reduction in Aβ production [118]. Additional evidence for substrate competition has been provided recently for another substrate of γ-secretase, the cytoplasmic tail of the low-density lipoprotein receptor—related protein (LRP), which has also been shown to interact with PS1 [119]. The co- expression of the LRP C-terminal fragment and APP in CHO cells has been shown to result in a reduction in secreted levels of Aβ40 and Aβ42 [119].
Taken together, these studies not only suggest substrate competition but also the presence of a single γ-secretase. However, the presence of multiple γ-secretase complexes cannot be ruled out. As discussed above there is evidence in the literature for the formation of γ-secretase complexes with distinct functions. All of the studies described above are undertaken in conditions where there is a large amount of substrate available which could down-regulate the activity of one γ-secretase for another. Whether substrate competition occurs under physiological conditions and if there are certain conditions that promote a higher expression of one substrate over another remains to be determined.
Concluding comments
It is well established that together with presenilin, its binding proteins, nicastrin, aph-1 and pen-2 are the only proteins that constitute the γ-secretase complex responsible for generating Aβ from APP. The function(s) of each γ-secretase component within the complex is gradually becoming apparent. However, there are many facets of γ-secretase activity that are yet to be identified such as the domain responsible for actual catalytic activity; the mechanisms of enzymatic activity within the lipid bi-layer; and the existence of single or multiple γ-secretase complexes. Further investigation using appropriate protein expression models and reconstitution studies such as those used to validate the protein components of γ-secretase complex is required to provide some insight into the processes that underlie the catalytic activity of this enzyme. Indeed, reconstitution of the γ-secretase complex in insect cells and purification of this complex from the cells have provided researchers with the first 3D images of the structure of this intricate enzyme [120, 121]. Identification of the specific catalytic domain(s) within the complex would provide a site that could be a potential target to develop agents that specifically modulate Aβ levels without altering the other known (and unknown) activities of the γ-secretase enzyme. Recent insight into this notion was provided by the finding that certain compounds selectively block the cleavage of APP but not Notch [122]. However, it is important to note that other presenilin interacting proteins not incorporated into the complex and the pathways they are involved in play important roles in APP processing and Aβ generation. These interactions assist in many facets of γ-secretase activity, including modulating substrate/enzyme availability, and thus are important to consider when developing appropriate therapeutic strategies.
Abbreviations
- APP
amyloid precursor protein
- Aβ
beta amyloid
- AICD
APP intracellular domain
- aph-1
anterior pharynx defective 1
- pen-2
presenilin enhancer 2
- PS1
presenilin 1
- PS2
presenilin 2
- TMP21
transmembrane protein 21
Footnotes
Special issue dedicated to John P. Blass.
References
- 1.Verdile G, Fuller S, Atwood CS, Laws SM, Gandy SE, Martins RN (2004) The role of beta amyloid in Alzheimer’s disease: still a cause of everything or the only one who got caught? Pharmacol Res 50:397–409 [DOI] [PubMed]
- 2.Reddy PH (2006) Amyloid precursor protein-mediated free radicals and oxidative damage: implications for the development and progression of Alzheimer’s disease. J Neurochem 96:1–13 [DOI] [PubMed]
- 3.Gandy S, Petanceska S (2001) Regulation of alzheimer beta-amyloid precursor trafficking and metabolism. Adv Exp Med Biol 487:85–100 [DOI] [PubMed]
- 4.Yu C, Kim SH, Ikeuchi T, Xu H, Gasparini L, Wang R, Sisodia SS (2001) Characterization of a presenilin-mediated amyloid precursor protein carboxyl-terminal fragment gamma. Evidence for distinct mechanisms involved in gamma-secretase processing of the APP and Notch1 transmembrane domains. J Biol Chem 276:43756–43760 Epub 42001 Oct 43752 [DOI] [PubMed]
- 5.Weidemann A, Eggert S, Reinhard FB, Vogel M, Paliga K, Baier G, Masters CL, Beyreuther K, Evin G (2002) A novel epsilon-cleavage within the transmembrane domain of the Alzheimer amyloid precursor protein demonstrates homology with Notch processing. Biochemistry 41:2825–2835 [DOI] [PubMed]
- 6.Zhao G, Mao G, Tan J, Dong Y, Cui MZ, Kim SH, Xu X (2004) Identification of a new presenilin-dependent zeta-cleavage site within the transmembrane domain of amyloid precursor protein. J Biol Chem 279:50647–50650 [DOI] [PubMed]
- 7.Zhao G, Cui MZ, Mao G, Dong Y, Tan J, Sun L, Xu X (2005) gamma-Cleavage is dependent on zeta-cleavage during the proteolytic processing of amyloid precursor protein within its transmembrane domain. J Biol Chem 280:37689–37697 [DOI] [PubMed]
- 8.Bergman A, Religa D, Karlstrom H, Laudon H, Winblad B, Lannfelt L, Lundkvist J, Naslund J (2003) APP intracellular domain formation and unaltered signaling in the presence of familial Alzheimer’s disease mutations. Exp Cell Res 287:1–9 [DOI] [PubMed]
- 9.Kim HS, Kim EM, Lee JP, Park CH, Kim S, Seo JH, Chang KA, Yu E, Jeong SJ, Chong YH, Suh YH (2003) C-terminal fragments of amyloid precursor protein exert neurotoxicity by inducing glycogen synthase kinase-3beta expression. Faseb J 17:1951–1953 Epub 2003 Aug 1915 [DOI] [PubMed]
- 10.Wolfe MS, Kopan R (2004) Intramembrane proteolysis: theme and variations. Science 305:1119–1123 [DOI] [PubMed]
- 11.Kimberly WT, LaVoie MJ, Ostaszewski BL, Ye W, Wolfe MS, Selkoe DJ (2003) Gamma-secretase is a membrane protein complex comprised of presenilin, nicastrin, Aph-1, and Pen-2. Proc Natl Acad Sci USA 100:6382–6387 [DOI] [PMC free article] [PubMed]
- 12.Yu G, Chen F, Levesque G, Nishimura M, Zhang DM, Levesque L, Rogaeva E, Xu D, Liang Y, Duthie M, St George-Hyslop PH, Fraser PE (1998) The presenilin 1 protein is a component of a high molecular weight intracellular complex that contains beta-catenin. J Biol Chem 273:16470–16475 [DOI] [PubMed]
- 13.Farmery MR, Tjernberg LO, Pursglove SE, Bergman A, Winblad B, Naslund J (2003) Partial purification and characterization of gamma-secretase from post-mortem human brain. J Biol Chem 278:24277–24284 [DOI] [PubMed]
- 14.Edbauer D, Winkler E, Haass C, Steiner H (2002) Presenilin and nicastrin regulate each other and determine amyloid beta-peptide production via complex formation. Proc Natl Acad Sci USA 99:8666–8671 [DOI] [PMC free article] [PubMed]
- 15.Steiner H, Winkler E, Edbauer D, Prokop S, Basset G, Yamasaki A, Kostka M, Haass C (2002) PEN-2 is an integral component of the gamma-secretase complex required for coordinated expression of presenilin and nicastrin. J Biol Chem 277:39062–39065 [DOI] [PubMed]
- 16.Li T, Ma G, Cai H, Price DL, Wong PC (2003) Nicastrin is required for assembly of presenilin/gamma-secretase complexes to mediate Notch signaling and for processing and trafficking of beta-amyloid precursor protein in mammals. J Neurosci 23:3272–3277 [DOI] [PMC free article] [PubMed]
- 17.Nyabi O, Bentahir M, Horre K, Herreman A, Gottardi-Littell N, Van Broeckhoven C, Merchiers P, Spittaels K, Annaert W, De Strooper B (2003) Presenilins mutated at Asp-257 or Asp-385 restore Pen-2 expression and Nicastrin glycosylation but remain catalytically inactive in the absence of wild type Presenilin. J Biol Chem 278:43430–43436 [DOI] [PubMed]
- 18.Li YM, Lai MT, Xu M, Huang Q, DiMuzio-Mower J, Sardana MK, Shi XP, Yin KC, Shafer JA, Gardell SJ (2000) Presenilin 1 is linked with gamma-secretase activity in the detergent solubilized state. Proc Natl Acad Sci USA 97:6138–6143 [DOI] [PMC free article] [PubMed]
- 19.Francis R, McGrath G, Zhang J, Ruddy DA, Sym M, Apfeld J, Nicoll M, Maxwell M, Hai B, Ellis MC, Parks AL, Xu W, Li J, Gurney M, Myers RL, Himes CS, Hiebsch R, Ruble C, Nye JS, Curtis D (2002) Aph-1 and pen-2 are required for Notch pathway signaling, gamma-secretase cleavage of beta APP, and presenilin protein accumulation. Dev Cell 3:85–97 [DOI] [PubMed]
- 20.Lee SF, Shah S, Li H, Yu C, Han W, Yu G (2002) Mammalian APH-1 interacts with presenilin and nicastrin and is required for intramembrane proteolysis of amyloid-beta precursor protein and Notch. J Biol Chem 277:45013–45019 [DOI] [PubMed]
- 21.Gu Y, Chen F, Sanjo N, Kawarai T, Hasegawa H, Duthie M, Li W, Ruan X, Luthra A, Mount HT, Tandon A, Fraser PE, St George-Hyslop P (2003) APH-1 interacts with mature and immature forms of presenilins and nicastrin and may play a role in maturation of presenilin–nicastrin complexes. J Biol Chem 278:7374–7380 [DOI] [PubMed]
- 22.Hu Y, Fortini ME (2003) Different cofactor activities in gamma-secretase assembly: evidence for a nicastrin-Aph-1 subcomplex. J Cell Biol 161:685–690 [DOI] [PMC free article] [PubMed]
- 23.Luo WJ, Wang H, Li H, Kim BS, Shah S, Lee HJ, Thinakaran G, Kim TW, Yu G, Xu H (2003) PEN-2 and APH-1 coordinately regulate proteolytic processing of presenilin 1. J Biol Chem 278:7850–7854 [DOI] [PubMed]
- 24.Takasugi N, Tomita T, Hayashi I, Tsuruoka M, Niimura M, Takahashi Y, Thinakaran G, Iwatsubo T (2003) The role of presenilin cofactors in the gamma-secretase complex. Nature 422:438–441 [DOI] [PubMed]
- 25.Edbauer D, Winkler E, Regula JT, Pesold B, Steiner H, Haass C (2003) Reconstitution of gamma-secretase activity. Nat Cell Biol 5:486–488 [DOI] [PubMed]
- 26.Hayashi I, Urano Y, Fukuda R, Isoo N, Kodama T, Hamakubo T, Tomita T, Iwatsubo T (2004) Selective reconstitution and recovery of functional gamma-secretase complex on budded baculovirus particles. J Biol Chem 279:38040–38046 [DOI] [PubMed]
- 27.Zhang L, Lee J, Song L, Sun X, Shen J, Terracina G, Parker EM (2005) Characterization of the reconstituted gamma-secretase complex from Sf9 cells co-expressing presenilin 1, nicastrin [correction of nacastrin], aph-1a, and pen-2. Biochemistry 44:4450–4457 [DOI] [PubMed]
- 28.Borchelt DR, Thinakaran G, Eckman CB, Lee MK, Davenport F, Ratovitsky T, Prada CM, Kim G, Seekins S, Yager D, Slunt HH, Wang R, Seeger M, Levey AI, Gandy SE, Copeland NG, Jenkins NA, Price DL, Younkin SG, Sisodia SS (1996) Familial Alzheimer’s disease-linked presenilin 1 variants elevate Abeta1–42/1–40 ratio in vitro and in vivo. Neuron 17:1005–1013 [DOI] [PubMed]
- 29.Lemere CA, Lopera F, Kosik KS, Lendon CL, Ossa J, Saido TC, Yamaguchi H, Ruiz A, Martinez A, Madrigal L, Hincapie L, Arango JC, Anthony DC, Koo EH, Goate AM, Selkoe DJ (1996) The E280A presenilin 1 Alzheimer mutation produces increased A beta 42 deposition and severe cerebellar pathology. Nat Med 2:1146–1150 [DOI] [PubMed]
- 30.Xia W, Zhang J, Kholodenko D, Citron M, Podlisny MB, Teplow DB, Haass C, Seubert P, Koo EH, Selkoe DJ (1997) Enhanced production and oligomerization of the 42-residue amyloid beta-protein by Chinese hamster ovary cells stably expressing mutant presenilins. J Biol Chem 272:7977–7982 [DOI] [PubMed]
- 31.De Strooper B, Saftig P, Craessaerts K, Vanderstichele H, Guhde G, Annaert W, Von Figura K, Van Leuven F (1998) Deficiency of presenilin-1 inhibits the normal cleavage of amyloid precursor protein. Nature 391:387–390 [DOI] [PubMed]
- 32.Wolfe MS, Xia W, Ostaszewski BL, Diehl TS, Kimberly WT, Selkoe DJ (1999) Two transmembrane aspartates in presenilin-1 required for presenilin endoproteolysis and gamma-secretase activity. Nature 398:513–517 [DOI] [PubMed]
- 33.Steiner H, Duff K, Capell A, Romig H, Grim MG, Lincoln S, Hardy J, Yu X, Picciano M, Fechteler K, Citron M, Kopan R, Pesold B, Keck S, Baader M, Tomita T, Iwatsubo T, Baumeister R, Haass C (1999) A loss of function mutation of presenilin-2 interferes with amyloid beta-peptide production and notch signaling. J Biol Chem 274:28669–28673 [DOI] [PubMed]
- 34.Kimberly WT, Xia W, Rahmati T, Wolfe MS, Selkoe DJ (2000) The transmembrane aspartates in presenilin 1 and 2 are obligatory for gamma-secretase activity and amyloid beta-protein generation. J Biol Chem 275:3173–3178 [DOI] [PubMed]
- 35.Li YM, Xu M, Lai MT, Huang Q, Castro JL, DiMuzio-Mower J, Harrison T, Lellis C, Nadin A, Neduvelil JG, Register RB, Sardana MK, Shearman MS, Smith AL, Shi XP, Yin KC, Shafer JA, Gardell SJ (2000) Photoactivated gamma-secretase inhibitors directed to the active site covalently label presenilin 1. Nature 405:689–694 [DOI] [PubMed]
- 36.Esler WP, Kimberly WT, Ostaszewski BL, Diehl TS, Moore CL, Tsai JY, Rahmati T, Xia W, Selkoe DJ, Wolfe MS (2000) Transition-state analogue inhibitors of gamma-secretase bind directly to presenilin-1. Nat Cell Biol 2:428–434 [DOI] [PubMed]
- 37.Seiffert D, Bradley JD, Rominger CM, Rominger DH, Yang F, Meredith JE Jr, Wang Q, Roach AH, Thompson LA, Spitz SM, Higaki JN, Prakash SR, Combs AP, Copeland RA, Arneric SP, Hartig PR, Robertson DW, Cordell B, Stern AM, Olson RE, Zaczek R (2000) Presenilin-1 and -2 are molecular targets for gamma-secretase inhibitors. J Biol Chem 275:34086–34091 [DOI] [PubMed]
- 38.Verdile G, Martins RN, Duthie M, Holmes E, St. George-Hyslop PH, Fraser PE (2000) Inhibiting amyloid precursor protein C-terminal cleavage promotes an interaction with presenilin 1. J Biol Chem 275:20794–20798 [DOI] [PubMed]
- 39.Xia W, Ray WJ, Ostaszewski BL, Rahmati T, Kimberly WT, Wolfe MS, Zhang J, Goate AM, Selkoe DJ (2000) Presenilin complexes with the C-terminal fragments of amyloid precursor protein at the sites of amyloid beta-protein generation. Proc Natl Acad Sci USA 97:9299–9304 [DOI] [PMC free article] [PubMed]
- 40.Annaert WG, Esselens C, Baert V, Boeve C, Snellings G, Cupers P, Craessaerts K, De Strooper B (2001) Interaction with telencephalin and the amyloid precursor protein predicts a ring structure for presenilins. Neuron 32:579–589 [DOI] [PubMed]
- 41.Liz MA, Sousa MM (2005) Deciphering cryptic proteases. Cell Mol Life Sci 62:989–1002 [DOI] [PMC free article] [PubMed]
- 42.Wolfe MS (2003) The secretases of Alzheimer’s disease. Curr Top Dev Biol 54:233–261 [DOI] [PubMed]
- 43.Xia W, Wolfe MS (2003) Intramembrane proteolysis by presenilin and presenilin-like proteases. J Cell Sci 116:2839–2844 [DOI] [PubMed]
- 44.Thinakaran G, Borchelt DR, Lee MK, Slunt HH, Spitzer L, Kim G, Ratovitsky T, Davenport F, Nordstedt C, Seeger M, Hardy J, Levey AI, Gandy SE, Jenkins NA, Copeland NG, Price DL, Sisodia SS (1996) Endoproteolysis of presenilin 1 and accumulation of processed derivatives in vivo. Neuron 17:181–190 [DOI] [PubMed]
- 45.Podlisny MB, Citron M, Amarante P, Sherrington R, Xia W, Zhang J, Diehl T, Levesque G, Fraser P, Haass C, Koo EH, Seubert P, St George-Hyslop P, Teplow DB, Selkoe DJ (1997) Presenilin proteins undergo heterogeneous endoproteolysis between Thr291 and Ala299 and occur as stable N- and C-terminal fragments in normal and Alzheimer brain tissue. Neurobiol Dis 3:325–337 [DOI] [PubMed]
- 46.Levitan D, Lee J, Song L, Manning R, Wong G, Parker E, Zhang L (2001) PS1 N- and C-terminal fragments form a complex that functions in APP processing and Notch signaling. Proc Natl Acad Sci USA 98:12186–12190 [DOI] [PMC free article] [PubMed]
- 47.Laudon H, Mathews PM, Karlstrom H, Bergman A, Farmery MR, Nixon RA, Winblad B, Gandy SE, Lendahl U, Lundkvist J, Naslund J (2004) Co-expressed presenilin 1 NTF and CTF form functional gamma-secretase complexes in cells devoid of full-length protein. J Neurochem 89:44–53 [DOI] [PubMed]
- 48.Capell A, Grunberg J, Pesold B, Diehlmann A, Citron M, Nixon R, Beyreuther K, Selkoe DJ, Haass C (1998) The proteolytic fragments of the Alzheimer’s disease-associated presenilin-1 form heterodimers and occur as a 100–150-kDa molecular mass complex. J Biol Chem 273:3205–3211 [DOI] [PubMed]
- 49.Seeger M, Nordstedt C, Petanceska S, Kovacs DM, Gouras GK, Hahne S, Fraser P, Levesque L, Czernik AJ, George-Hyslop PS, Sisodia SS, Thinakaran G, Tanzi RE, Greengard P, Gandy S (1997) Evidence for phosphorylation and oligomeric assembly of presenilin 1. Proc Natl Acad Sci USA 94:5090–5094 [DOI] [PMC free article] [PubMed]
- 50.Steiner H, Capell A, Pesold B, Citron M, Kloetzel PM, Selkoe DJ, Romig H, Mendla K, Haass C (1998) Expression of Alzheimer’s disease-associated presenilin-1 is controlled by proteolytic degradation and complex formation. J Biol Chem 273:32322–32331 [DOI] [PubMed]
- 51.Beher D, Elle C, Underwood J, Davis JB, Ward R, Karran E, Masters CL, Beyreuther K, Multhaup G (1999) Proteolytic fragments of Alzheimer’s disease-associated presenilin 1 are present in synaptic organelles and growth cone membranes of rat brain. J Neurochem 72:1564–1573 [DOI] [PubMed]
- 52.Yu G, Chen F, Nishimura M, Steiner H, Tandon A, Kawarai T, Arawaka S, Supala A, Song YQ, Rogaeva E, Holmes E, Zhang DM, Milman P, Fraser P, Haass C, St. George-Hyslop P (2000) Mutation of conserved aspartates affect maturation of presenilin 1 and presenilin 2 complexes. Acta Neurol Scand Suppl 176:6–11 [DOI] [PubMed]
- 53.Cervantes S, Gonzalez-Duarte R, Marfany G (2001) Homodimerization of presenilin N-terminal fragments is affected by mutations linked to Alzheimer’s disease. FEBS Lett 505:81–86 [DOI] [PubMed]
- 54.Schroeter EH, Ilagan MX, Brunkan AL, Hecimovic S, Li YM, Xu M, Lewis HD, Saxena MT, De Strooper B, Coonrod A, Tomita T, Iwatsubo T, Moore CL, Goate A, Wolfe MS, Shearman M, Kopan R (2003) A presenilin dimer at the core of the gamma-secretase enzyme: insights from parallel analysis of Notch 1 and APP proteolysis. Proc Natl Acad Sci USA 100:13075–13080 [DOI] [PMC free article] [PubMed]
- 55.Yu G, Nishimura M, Arawaka S, Levitan D, Zhang L, Tandon A, Song YQ, Rogaeva E, Chen F, Kawarai T, Supala A, Levesque L, Yu H, Yang DS, Holmes E, Milman P, Liang Y, Zhang DM, Xu DH, Sato C, Rogaev E, Smith M, Janus C, Zhang Y, Aebersold R, Farrer LS, Sorbi S, Bruni A, Fraser P, St. George-Hyslop P (2000) Nicastrin modulates presenilin-mediated notch/glp-1 signal transduction and betaAPP processing. Nature 407:48–54 [DOI] [PubMed]
- 56.Leem JY, Vijayan S, Han P, Cai D, Machura M, Lopes KO, Veselits ML, Xu H, Thinakaran G (2002) Presenilin 1 is required for maturation and cell surface accumulation of nicastrin. J Biol Chem 277:19236–19240 [DOI] [PubMed]
- 57.Tomita T, Katayama R, Takikawa R, Iwatsubo T (2002) Complex N-glycosylated form of nicastrin is stabilized and selectively bound to presenilin fragments. FEBS Lett 520:117–121 [DOI] [PubMed]
- 58.Yang DS, Tandon A, Chen F, Yu G, Yu H, Arawaka S, Hasegawa H, Duthie M, Schmidt SD, Ramabhadran TV, Nixon RA, Mathews PM, Gandy SE, Mount HT, St. George-Hyslop P, Fraser PE (2002) Mature glycosylation and trafficking of nicastrin modulate its binding to presenilins. J Biol Chem 277:28135–28142 [DOI] [PubMed]
- 59.Chung HM, Struhl G (2001) Nicastrin is required for Presenilin-mediated transmembrane cleavage in Drosophila. Nat Cell Biol 3:1129–1132 [DOI] [PubMed]
- 60.Hu Y, Ye Y, Fortini ME (2002) Nicastrin is required for gamma-secretase cleavage of the Drosophila Notch receptor. Dev Cell 2:69–78 [DOI] [PubMed]
- 61.Lopez-Schier H, St Johnston D (2002) Drosophila nicastrin is essential for the intramembranous cleavage of notch. Dev Cell 2:79–89 [DOI] [PubMed]
- 62.Zhang T, Tran S, Clouser C, Pignoni F, Zhang YW, Luo WJ, Wang H, Lin P, Vetrivel KS, Liao F, Li F, Wong PC, Farquhar MG, Thinakaran G, and Xu H (2005) Nicastrin controls aspects of photoreceptor neuron specification and differentiation in the Drosophila eye Nicastrin is critical for stability and trafficking but not association of other presenilin/gamma-secretase components. Dev Dyn 234:590–601 [DOI] [PubMed]
- 63.Shah S, Lee SF, Tabuchi K, Hao YH, Yu C, LaPlant Q, Ball H, Dann CE III, Sudhof T, Yu G (2005) Nicastrin functions as a gamma-secretase-substrate receptor. Cell 122:435–447 [DOI] [PubMed]
- 64.Berezovska O, Ramdya P, Skoch J, Wolfe MS, Bacskai BJ, Hyman BT (2003) Amyloid precursor protein associates with a nicastrin-dependent docking site on the presenilin 1-gamma-secretase complex in cells demonstrated by fluorescence lifetime imaging. J Neurosci 23:4560–4566 [DOI] [PMC free article] [PubMed]
- 65.Goutte C, Tsunozaki M, Hale VA, Priess JR (2002) APH-1 is a multipass membrane protein essential for the Notch signaling pathway in Caenorhabditis elegans embryos. Proc Natl Acad Sci USA 99:775–779 [DOI] [PMC free article] [PubMed]
- 66.Shirotani K, Edbauer D, Prokop S, Haass C, Steiner H (2004) Identification of distinct gamma-secretase complexes with different APH-1 variants. J Biol Chem 279:41340–41345 [DOI] [PubMed]
- 67.Ma G, Li T, Price DL, Wong PC (2005) APH-1a is the principal mammalian APH-1 isoform present in gamma-secretase complexes during embryonic development. J Neurosci 25:192–198 [DOI] [PMC free article] [PubMed]
- 68.Lee SF, Shah S, Yu C, Wigley WC, Li H, Lim M, Pedersen K, Han W, Thomas P, Lundkvist J, Hao YH, Yu G (2004) A conserved GXXXG motif in APH-1 is critical for assembly and activity of the gamma-secretase complex. J Biol Chem 279:4144–4152 [DOI] [PubMed]
- 69.Niimura M, Isoo N, Takasugi N, Tsuruoka M, Ui-Tei K, Saigo K, Morohashi Y, Tomita T, Iwatsubo T (2005) Aph-1 contributes to the stabilization and trafficking of the gamma-secretase complex through mechanisms involving intermolecular and intramolecular interactions. J Biol Chem 280:12967–12975 [DOI] [PubMed]
- 70.LaVoie MJ, Fraering PC, Ostaszewski BL, Ye W, Kimberly WT, Wolfe MS, Selkoe DJ (2003) Assembly of the gamma-secretase complex involves early formation of an intermediate subcomplex of Aph-1 and nicastrin. J Biol Chem 278:37213–37222 [DOI] [PubMed]
- 71.Kim SH, Ikeuchi T, Yu C, Sisodia SS (2003) Regulated hyperaccumulation of presenilin-1 and the “gamma-secretase” complex. Evidence for differential intramembranous processing of transmembrane subatrates. J Biol Chem 278:33992–34002 [DOI] [PubMed]
- 72.Hansson EM, Stromberg K, Bergstedt S, Yu G, Naslund J, Lundkvist J, Lendahl U (2005) Aph-1 interacts at the cell surface with proteins in the active gamma-secretase complex and membrane-tethered Notch. J Neurochem 92:1010–1020 [DOI] [PubMed]
- 73.Urban S, Lee JR, Freeman M (2001) Drosophila rhomboid-1 defines a family of putative intramembrane serine proteases. Cell 107:173–182 [DOI] [PubMed]
- 74.De Strooper B (2003) Aph-1, Pen-2, and Nicastrin with Presenilin generate an active gamma-secretase complex. Neuron 38:9–12 [DOI] [PubMed]
- 75.Morais VA, Crystal AS, Pijak DS, Carlin D, Costa J, Lee VM, Doms RW (2003) The transmembrane domain region of nicastrin mediates direct interactions with APH-1 and the gamma-secretase complex. J Biol Chem 278:43284–43291 [DOI] [PubMed]
- 76.Bergman A, Hansson EM, Pursglove SE, Farmery MR, Lannfelt L, Lendahl U, Lundkvist J, Naslund J (2004) Pen-2 is sequestered in the endoplasmic reticulum and subjected to ubiquitylation and proteasome-mediated degradation in the absence of presenilin. J Biol Chem 279:16744–16753 [DOI] [PubMed]
- 77.Crystal AS, Morais VA, Fortna RR, Carlin D, Pierson TC, Wilson CA, Lee VM, Doms RW (2004) Presenilin modulates Pen-2 levels posttranslationally by protecting it from proteasomal degradation. Biochemistry 43:3555–3563 [DOI] [PubMed]
- 78.Hasegawa H, Sanjo N, Chen F, Gu YJ, Shier C, Petit A, Kawarai T, Katayama T, Schmidt SD, Mathews PM, Schmitt-Ulms G, Fraser PE, St. George-Hyslop P (2004) Both the sequence and length of the C terminus of PEN-2 are critical for intermolecular interactions and function of presenilin complexes. J Biol Chem 279:46455–46463 [DOI] [PubMed]
- 79.Watanabe N, Tomita T, Sato C, Kitamura T, Morohashi Y, Iwatsubo T (2005) Pen-2 is incorporated into the gamma-secretase complex through binding to transmembrane domain 4 of presenilin 1. J Biol Chem 280:41967–41975 [DOI] [PubMed]
- 80.Kim SH, Sisodia SS (2005) Evidence that the “NF” motif in transmembrane domain 4 of presenilin 1 is critical for binding with PEN-2. J Biol Chem 280:41953–41966 [DOI] [PubMed]
- 81.Prokop S, Haass C, Steiner H (2005) Length and overall sequence of the PEN-2 C-terminal domain determines its function in the stabilization of presenilin fragments. J Neurochem 94:57–62 [DOI] [PubMed]
- 82.Chen F, Hasegawa H, Schmitt-Ulms G, Kawarai T, Bohm C, Katayama T, Gu Y, Sanjo N, Glista M, Rogaeva E, Wakutani Y, Pardossi-Piquard R, Ruan X, Tandon A, Checler F, Marambaud P, Hansen K, Westaway D, St. George-Hyslop P, Fraser P (2006) TMP21 is a presenilin complex component that modulates gamma-secretase but not epsilon-secretase activity. Nature 440:1208–1212 [DOI] [PubMed]
- 83.Blum R, Feick P, Puype M, Vandekerckhove J, Klengel R, Nastainczyk W, Schulz I (1996) Tmp21 and p24A, two type I proteins enriched in pancreatic microsomal membranes, are members of a protein family involved in vesicular trafficking. J Biol Chem 271:17183–17189 [DOI] [PubMed]
- 84.Barr FA, Preisinger C, Kopajtich R, Korner R (2001) Golgi matrix proteins interact with p24 cargo receptors and aid their efficient retention in the Golgi apparatus. J Cell Biol 155:885–891 [DOI] [PMC free article] [PubMed]
- 85.Levy-Lahad E, Wasco W, Poorkaj P, Romano DM, Oshima J, Pettingell WH, Yu CE, Jondro PD, Schmidt SD, Wang K, et al (1995) Candidate gene for the chromosome 1 familial Alzheimer’s disease locus. Science 269:973–977 [DOI] [PubMed]
- 86.Rogaev EI, Sherrington R, Rogaeva EA, Levesque G, Ikeda M, Liang Y, Chi H, Lin C, Holman K, Tsuda T, et al (1995) Familial Alzheimer’s disease in kindreds with missense mutations in a gene on chromosome 1 related to the Alzheimer’s disease type 3 gene. Nature 376:775–778 [DOI] [PubMed]
- 87.Shen J, Bronson RT, Chen DF, Xia W, Selkoe DJ, Tonegawa S (1997) Skeletal and CNS defects in Presenilin-1-deficient mice. Cell 89:629–639 [DOI] [PubMed]
- 88.Wong PC, Zheng H, Chen H, Becher MW, Sirinathsinghji DJ, Trumbauer ME, Chen HY, Price DL, Van der Ploeg LH, Sisodia SS (1997) Presenilin 1 is required for Notch1 and DII1 expression in the paraxial mesoderm. Nature 387:288–292 [DOI] [PubMed]
- 89.Herreman A, Hartmann D, Annaert W, Saftig P, Craessaerts K, Serneels L, Umans L, Schrijvers V, Checler F, Vanderstichele H, Baekelandt V, Dressel R, Cupers P, Huylebroeck D, Zwijsen A, Van Leuven F, De Strooper B (1999) Presenilin 2 deficiency causes a mild pulmonary phenotype and no changes in amyloid precursor protein processing but enhances the embryonic lethal phenotype of presenilin 1 deficiency. Proc Natl Acad Sci USA 96:11872–11877 [DOI] [PMC free article] [PubMed]
- 90.Rozmahel R, Mount HT, Chen F, Nguyen V, Huang J, Erdebil S, Liauw J, Yu G, Hasegawa H, Gu Y, Song YQ, Schmidt SD, Nixon RA, Mathews PM, Bergeron C, Fraser P, Westaway D, St. George-Hyslop P (2002) Alleles at the Nicastrin locus modify presenilin 1-deficiency phenotype. Proc Natl Acad Sci USA 99:14452–14457 [DOI] [PMC free article] [PubMed]
- 91.Lai MT, Chen E, Crouthamel MC, DiMuzio-Mower J, Xu M, Huang Q, Price E, Register RB, Shi XP, Donoviel DB, Bernstein A, Hazuda D, Gardell SJ, Li YM (2003) Presenilin-1 and presenilin-2 exhibit distinct yet overlapping gamma-secretase activities. J Biol Chem 278:22475–22481 [DOI] [PubMed]
- 92.Stromberg K, Hansson EM, Laudon H, Bergstedt S, Naslund J, Lundkvist J, Lendahl U (2005) gamma-secretase complexes containing N- and C-terminal fragments of different presenilin origin retain normal gamma-secretase activity. J Neurochem 95:880–890 [DOI] [PubMed]
- 93.Mastrangelo P, Mathews PM, Chishti MA, Schmidt SD, Gu Y, Yang J, Mazzella MJ, Coomaraswamy J, Horne P, Strome B, Pelly H, Levesque G, Ebeling C, Jiang Y, Nixon RA, Rozmahel R, Fraser PE, St. George-Hyslop P, Carlson GA, Westaway D (2005) Dissociated phenotypes in presenilin transgenic mice define functionally distinct gamma-secretases. Proc Natl Acad Sci USA 102:8972–8977 [DOI] [PMC free article] [PubMed]
- 94.Buxbaum JD, Choi EK, Luo Y, Lilliehook C, Crowley AC, Merriam DE, Wasco W (1998) Calsenilin: a calcium-binding protein that interacts with the presenilins and regulates the levels of a presenilin fragment. Nat Med 4:1177–1181 [DOI] [PubMed]
- 95.Buxbaum JD (2004) A role for calsenilin and related proteins in multiple aspects of neuronal function. Biochem Biophys Res Commun 322:1140–1144 [DOI] [PubMed]
- 96.Choi EK, Zaidi NF, Miller JS, Crowley AC, Merriam DE, Lilliehook C, Buxbaum JD, Wasco W (2001) Calsenilin is a substrate for caspase-3 that preferentially interacts with the familial Alzheimer’s disease-associated C-terminal fragment of presenilin 2. J Biol Chem 276:19197–19204 [DOI] [PubMed]
- 97.Leissring MA, Yamasaki TR, Wasco W, Buxbaum JD, Parker I, LaFerla FM (2000) Calsenilin reverses presenilin-mediated enhancement of calcium signaling. Proc Natl Acad Sci USA 97:8590–8593 [DOI] [PMC free article] [PubMed]
- 98.Lilliehook C, Chan S, Choi EK, Zaidi NF, Wasco W, Mattson MP, Buxbaum JD (2002) Calsenilin enhances apoptosis by altering endoplasmic reticulum calcium signaling. Mol Cell Neurosci 19:552–559 [DOI] [PubMed]
- 99.Jo DG, Chang JW, Hong HS, Mook-Jung I, Jung YK (2003) Contribution of presenilin/gamma-secretase to calsenilin-mediated apoptosis. Biochem Biophys Res Commun 305:62–66 [DOI] [PubMed]
- 100.Jo DG, Kim MJ, Choi YH, Kim IK, Song YH, Woo HN, Chung CW, Jung YK (2001) Pro-apoptotic function of calsenilin/DREAM/KChIP3. Faseb J 15:589–591 [DOI] [PubMed]
- 101.Jo DG, Jang J, Kim BJ, Lundkvist J, Jung YK (2005) Overexpression of calsenilin enhances gamma-secretase activity. Neurosci Lett 378:59–64 [DOI] [PubMed]
- 102.Lilliehook C, Bozdagi O, Yao J, Gomez-Ramirez M, Zaidi NF, Wasco W, Gandy S, Santucci AC, Haroutunian V, Huntley GW, Buxbaum JD (2003) Altered Abeta formation and long-term potentiation in a calsenilin knock-out. J Neurosci 23:9097–9106 [DOI] [PMC free article] [PubMed]
- 103.Borg JP, Yang Y, De Taddeo-Borg M, Margolis B, Turner RS (1998) The X11alpha protein slows cellular amyloid precursor protein processing and reduces Abeta40 and Abeta42 secretion. J Biol Chem 273:14761–14766 [DOI] [PubMed]
- 104.Sastre M, Turner RS, Levy E (1998) X11 interaction with beta-amyloid precursor protein modulates its cellular stabilization and reduces amyloid beta-protein secretion. J Biol Chem 273:22351–22357 [DOI] [PubMed]
- 105.McLoughlin DM, Irving NG, Brownlees J, Brion JP, Leroy K, Miller CC (1999) Mint2/X11-like colocalizes with the Alzheimer’s disease amyloid precursor protein and is associated with neuritic plaques in Alzheimer’s disease. Eur J Neurosci 11:1988–1994 [DOI] [PubMed]
- 106.Tomita S, Ozaki T, Taru H, Oguchi S, Takeda S, Yagi Y, Sakiyama S, Kirino Y, Suzuki T (1999) Interaction of a neuron-specific protein containing PDZ domains with Alzheimer’s amyloid precursor protein. J Biol Chem 274:2243–2254 [DOI] [PubMed]
- 107.McLoughlin DM, Miller CC (1996) The intracellular cytoplasmic domain of the Alzheimer’s disease amyloid precursor protein interacts with phosphotyrosine-binding domain proteins in the yeast two-hybrid system. FEBS Lett 397:197–200 [DOI] [PubMed]
- 108.Zhang Z, Lee CH, Mandiyan V, Borg JP, Margolis B, Schlessinger J, Kuriyan J (1997) Sequence-specific recognition of the internalization motif of the Alzheimer’s amyloid precursor protein by the X11 PTB domain. Embo J 16:6141–6150 [DOI] [PMC free article] [PubMed]
- 109.Tanahashi H, Tabira T (1999) X11L2, a new member of the X11 protein family, interacts with Alzheimer’s beta-amyloid precursor protein. Biochem Biophys Res Commun 255:663–667 [DOI] [PubMed]
- 110.Lau KF, McLoughlin DM, Standen C, Miller CC (2000) X11 alpha and X11 beta interact with presenilin-1 via their PDZ domains. Mol Cell Neurosci 16:557–565 [DOI] [PubMed]
- 111.Araki Y, Tomita S, Yamaguchi H, Miyagi N, Sumioka A, Kirino Y, Suzuki T (2003) Novel cadherin-related membrane proteins, Alcadeins, enhance the X11-like protein-mediated stabilization of amyloid beta-protein precursor metabolism. J Biol Chem 278:49448–49458 [DOI] [PubMed]
- 112.Ray WJ, Yao M, Mumm J, Schroeter EH, Saftig P, Wolfe M, Selkoe DJ, Kopan R, Goate AM (1999) Cell surface presenilin-1 participates in the gamma-secretase-like proteolysis of Notch. J Biol Chem 274:36801–36807 [DOI] [PubMed]
- 113.Sastre M, Steiner H, Fuchs K, Capell A, Multhaup G, Condron MM, Teplow DB, Haass C (2001) Presenilin-dependent gamma-secretase processing of beta-amyloid precursor protein at a site corresponding to the S3 cleavage of Notch. EMBO Rep 2:835–841 [DOI] [PMC free article] [PubMed]
- 114.Roncarati R, Sestan N, Scheinfeld MH, Berechid BE, Lopez PA, Meucci O, McGlade JC, Rakic P, D’Adamio L (2002) The gamma-secretase-generated intracellular domain of beta-amyloid precursor protein binds Numb and inhibits Notch signaling. Proc Natl Acad Sci USA 99:7102–7107 [DOI] [PMC free article] [PubMed]
- 115.Okochi M, Steiner H, Fukumori A, Tanii H, Tomita T, Tanaka T, Iwatsubo T, Kudo T, Takeda M, Haass C (2002) Presenilins mediate a dual intramembranous gamma-secretase cleavage of Notch-1. Embo J 21:5408–5416 [DOI] [PMC free article] [PubMed]
- 116.Berezovska O, Jack C, Deng A, Gastineau N, Rebeck GW, Hyman BT (2001) Notch1 and amyloid precursor protein are competitive substrates for presenilin1-dependent gamma-secretase cleavage. J Biol Chem 276:30018–30023 [DOI] [PubMed]
- 117.Kimberly WT, Esler WP, Ye W, Ostaszewski BL, Gao J, Diehl T, Selkoe DJ, Wolfe MS (2003) Notch and the amyloid precursor protein are cleaved by similar gamma-secretase(s). Biochemistry 42:137–144 [DOI] [PubMed]
- 118.Lleo A, Berezovska O, Ramdya P, Fukumoto H, Raju S, Shah T, Hyman BT (2003) Notch1 competes with the amyloid precursor protein for gamma-secretase and down-regulates presenilin-1 gene expression. J Biol Chem 278:47370–47375 [DOI] [PubMed]
- 119.Lleo A, Waldron E, von Arnim CA, Herl L, Tangredi MM, Peltan ID, Strickland DK, Koo EH, Hyman BT, Pietrzik CU, Berezovska O (2005) Low density lipoprotein receptor-related protein (LRP) interacts with presenilin 1 and is a competitive substrate of the amyloid precursor protein (APP) for gamma-secretase. J Biol Chem 280:27303–27309 [DOI] [PubMed]
- 120.Ogura T, Mio K, Hayashi I, Miyashita H, Fukuda R, Kopan R, Kodama T, Hamakubo T, Iwastubo T, Tomita T, Sato C (2006) Three-dimensional structure of the gamma-secretase complex. Biochem Biophys Res Commun 343:525–534 [DOI] [PubMed]
- 121.Lazarov VK, Fraering PC, Ye W, Wolfe MS, Selkoe DJ, Li H (2006) Electron microscopic structure of purified, active gamma-secretase reveals an aqueous intramembrane chamber and two pores. Proc Natl Acad Sci USA 103:6889–6894 [DOI] [PMC free article] [PubMed]
- 122.Fraering PC, Ye W, LaVoie MJ, Ostaszewski BL, Selkoe DJ, Wolfe MS (2005) Gamma-secretase substrate selectivity can be modulated directly via interaction with a nucleotide-binding site. J Biol Chem 280:41987–41996 [DOI] [PMC free article] [PubMed]
- 123.Shirotani K, Edbauer D, Kostka M, Steiner H, Haass C (2004) Immature nicastrin stabilizes APH-1 independent of PEN-2 and presenilin: identification of nicastrin mutants that selectively interact with APH-1. J Neurochem 89:1520–1527 [DOI] [PubMed]
- 124.Georgakopoulos A, Marambaud P, Friedrich VL Jr, Shioi J, Efthimiopoulos S, Robakis NK (2000) Presenilin-1: a component of synaptic and endothelial adherens junctions. Ann NY Acad Sci 920:209–214 [DOI] [PubMed]
- 125.Katayama T, Imaizumi K, Sato N, Miyoshi K, Kudo T, Hitomi J, Morihara T, Yoneda T, Gomi F, Mori Y, Nakano Y, Takeda J, Tsuda T, Itoyama Y, Murayama O, Takashima A, St. George-Hyslop P, Takeda M, Tohyama M (1999) Presenilin-1 mutations downregulate the signalling pathway of the unfolded-protein response. Nat Cell Biol 1:479–485 [DOI] [PubMed]
- 126.Niwa M, Sidrauski C, Kaufman RJ, Walter P (1999) A role for presenilin-1 in nuclear accumulation of Ire1 fragments and induction of the mammalian unfolded protein response. Cell 99:691–702 [DOI] [PubMed]
- 127.Zhang W, Han SW, McKeel DW, Goate A, Wu JY (1998) Interaction of presenilins with the filamin family of actin-binding proteins. J Neurosci 18:914–922 [DOI] [PMC free article] [PubMed]
- 128.Alberici A, Moratto D, Benussi L, Gasparini L, Ghidoni R, Gatta LB, Finazzi D, Frisoni GB, Trabucchi M, Growdon JH, Nitsch RM, Binetti G (1999) Presenilin 1 protein directly interacts with Bcl-2. J Biol Chem 274:30764–30769 [DOI] [PubMed]
- 129.Passer BJ, Pellegrini L, Vito P, Ganjei JK, D’Adamio L (1999) Interaction of Alzheimer’s presenilin-1 and presenilin-2 with Bcl-X(L). A potential role in modulating the threshold of cell death. J Biol Chem 274:24007–24013 [DOI] [PubMed]
- 130.Stabler SM, Ostrowski LL, Janicki SM, Monteiro MJ (1999) A myristoylated calcium-binding protein that preferentially interacts with the Alzheimer’s disease presenilin 2 protein. J Cell Biol 145:1277–1292 [DOI] [PMC free article] [PubMed]
- 131.Morohashi Y, Hatano N, Ohya S, Takikawa R, Watabiki T, Takasugi N, Imaizumi Y, Tomita T, Iwatsubo T (2002) Molecular cloning and characterization of CALP/KChIP4, a novel EF-hand protein interacting with presenilin 2 and voltage-gated potassium channel subunit Kv4. J Biol Chem 277:14965–14975 [DOI] [PubMed]
- 132.Johnsingh AA, Johnston JM, Merz G, Xu J, Kotula L, Jacobsen JS, Tezapsidis N (2000) Altered binding of mutated presenilin with cytoskeleton-interacting proteins. FEBS Lett 465:53–58 [DOI] [PubMed]
- 133.Tanahashi H, Tabira T (2000) Alzheimer’s disease-associated presenilin 2 interacts with DRAL, an LIM-domain protein. Hum Mol Genet 9:2281–2289 [DOI] [PubMed]
- 134.Kang DE, Yoon IS, Repetto E, Busse T, Yermian N, Ie L, Koo EH (2005) Presenilins mediate phosphatidylinositol 3-kinase/AKT and ERK activation via select signaling receptors. Selectivity of PS2 in platelet-derived growth factor signaling. J Biol Chem 280:31537–31547 [DOI] [PubMed]
- 135.Wang HQ, Nakaya Y, Du Z, Yamane T, Shirane M, Kudo T, Takeda M, Takebayashi K, Noda Y, Nakayama KI, Nishimura M (2005) Interaction of presenilins with FKBP38 promotes apoptosis by reducing mitochondrial Bcl-2. Hum Mol Genet 14:1889–1902 [DOI] [PubMed]
- 136.Smine A, Xu X, Nishiyama K, Katada T, Gambetti P, Yadav SP, Wu X, Shi YC, Yasuhara S, Homburger V, Okamoto T (1998) Regulation of brain G-protein go by Alzheimer’s disease gene presenilin-1. J Biol Chem 273:16281–16288 [DOI] [PubMed]
- 137.Takashima A, Murayama M, Murayama O, Kohno T, Honda T, Yasutake K, Nihonmatsu N, Mercken M, Yamaguchi H, Sugihara S, Wolozin B (1998) Presenilin 1 associates with glycogen synthase kinase-3beta and its substrate tau. Proc Natl Acad Sci USA 95:9637–9641 [DOI] [PMC free article] [PubMed]
- 138.Van Gassen G, C De Jonghe, Pype S, Van Criekinge W, Julliams A, Vanderhoeven I, Woodrow S, Beyaert R, Huylebroeck D, Van Broeckhoven C (1999) Alzheimer’s disease associated presenilin 1 interacts with HC5 and ZETA, subunits of the catalytic 20S proteasome. Neurobiol Dis 6:376–391 [DOI] [PubMed]
- 139.Zhang SX, Guo Y, Boulianne GL (2001) Identification of a novel family of putative methyltransferases that interact with human and Drosophila presenilins. Gene 280:135–144 [DOI] [PubMed]
- 140.Chen Q, Kimura H, Schubert D (2002) A novel mechanism for the regulation of amyloid precursor protein metabolism. J Cell Biol 158:79–89 [DOI] [PMC free article] [PubMed]
- 141.Gupta S, Singh R, Datta P, Zhang Z, Orr C, Lu Z, Dubois G, Zervos AS, Meisler MH, Srinivasula SM, Fernandes-Alnemri T, Alnemri ES (2004) The C-terminal tail of presenilin regulates Omi/HtrA2 protease activity. J Biol Chem 279:45844–45854 [DOI] [PubMed]
- 142.Zhou Y, Zhang W, Easton R, Ray JW, Lampe P, Jiang Z, Brunkan AL, Goate A, Johnson EM, Wu JY (2002) Presenilin-1 protects against neuronal apoptosis caused by its interacting protein PAG. Neurobiol Dis 9:126–138 [DOI] [PubMed]
- 143.Pellegrini L, Passer BJ, Canelles M, Lefterov I, Ganjei JK, Fowlkes BJ, Koonin EV, D’Adamio L (2001) PAMP and PARL, two novel putative metalloproteases interacting with the COOH-terminus of Presenilin-1 and -2. J Alzheimers Dis 3:181–190 [DOI] [PubMed]
- 144.Kang DE, Soriano S, Xia X, Eberhart CG, De Strooper B, Zheng H, Koo EH (2002) Presenilin couples the paired phosphorylation of beta-catenin independent of axin: implications for beta-catenin activation in tumorigenesis. Cell 110:751–762 [DOI] [PubMed]
- 145.Raurell I, Castano J, Franci C, Garcia de Herreros A, Dunach M (2006) Presenilin-1 interacts with plakoglobin and enhances plakoglobin-Tcf-4 association. Implications for the regulation of beta-catenin/Tcf-4-dependent transcription. J Biol Chem 281:1401–1411 [DOI] [PubMed]
- 146.Cai D, Netzer WJ, Zhong M, Lin Y, Du G, Frohman M, Foster DA, Sisodia SS, Xu H, Gorelick FS, Greengard P (2006) Presenilin-1 uses phospholipase D1 as a negative regulator of beta-amyloid formation. Proc Natl Acad Sci USA 103:1941–1946 [DOI] [PMC free article] [PubMed]
- 147.Xu X, Shi Y, Wu X, Gambetti P, Sui D, Cui MZ (1999) Identification of a novel PSD-95/Dlg/ZO-1 (PDZ)-like protein interacting with the C terminus of presenilin-1. J Biol Chem 274:32543–32546 [DOI] [PubMed]
- 148.Imafuku I, Masaki T, Waragai M, Takeuchi S, Kawabata M, Hirai S, Ohno S, Nee LE, Lippa CF, Kanazawa I, Imagawa M, Okazawa H (1999) Presenilin 1 suppresses the function of c-Jun homodimers via interaction with QM/Jif-1. J Cell Biol 147:121–134 [DOI] [PMC free article] [PubMed]
- 149.Dumanchin C, Czech C, Campion D, Cuif MH, Poyot T, Martin C, Charbonnier F, Goud B, Pradier L, Frebourg T (1999) Presenilins interact with Rab11, a small GTPase involved in the regulation of vesicular transport. Hum Mol Genet 8:1263–1269 [DOI] [PubMed]
- 150.Scheper W, Zwart R, Sluijs P, Annaert W, Gool WA, Baas F (2000) Alzheimer’s presenilin 1 is a putative membrane receptor for rab GDP dissociation inhibitor. Hum Mol Genet 9:303–310 [DOI] [PubMed]
- 151.Takeda T, Asahi M, Yamaguchi O, Hikoso S, Nakayama H, Kusakari Y, Kawai M, Hongo K, Higuchi Y, Kashiwase K, Watanabe T, Taniike M, Nakai A, Nishida K, Kurihara S, Donoviel DB, Bernstein A, Tomita T, Iwatsubo T, Hori M, Otsu K (2005) Presenilin 2 regulates the systolic function of heart by modulating Ca2+ signaling. Faseb J 19:2069–2071 [DOI] [PubMed]
- 152.Li J, Pauley AM, Myers RL, Shuang R, Brashler JR, Yan R, Buhl AE, Ruble C, Gurney ME (2002) SEL-10 interacts with presenilin 1, facilitates its ubiquitination, and alters A-beta peptide production. J Neurochem 82:1540–1548 [DOI] [PubMed]
- 153.Pack-Chung E, Meyers MB, Pettingell WP, Moir RD, Brownawell AM, Cheng I, Tanzi RE, Kim TW (2000) Presenilin 2 interacts with sorcin, a modulator of the ryanodine receptor. J Biol Chem 275:14440–14445 [DOI] [PubMed]
- 154.Smith SK, Anderson HA, Yu G, Robertson AG, Allen SJ, Tyler SJ, Naylor RL, Mason G, Wilcock GW, Roche PA, Fraser PE, Dawbarn D (2000) Identification of syntaxin 1A as a novel binding protein for presenilin-1. Brain Res Mol Brain Res 78:100–107 [DOI] [PubMed]
- 155.Suga K, Saito A, Tomiyama T, Mori H, Akagawa K (2005) Syntaxin 5 interacts specifically with presenilin holoproteins and affects processing of betaAPP in neuronal cells. J Neurochem 94:425–439 [DOI] [PubMed]
- 156.Suga K, Tomiyama T, Mori H, Akagawa K (2004) Syntaxin 5 interacts with presenilin holoproteins, but not with their N- or C-terminal fragments, and affects beta-amyloid peptide production. Biochem J 381:619–628 [DOI] [PMC free article] [PubMed]
- 157.Prihar G, Gonzalez de Chavez F, Baker M, Crook R, McGowan E, Grover A, Hardy J, Hutton M (1999) A novel candidate presenilin-1 interacting protein containing tetratricopeptide repeats. Neuroreport 10:1409–1415 [DOI] [PubMed]
- 158.Mah AL, Perry G, Smith MA, Monteiro MJ (2000) Identification of ubiquilin, a novel presenilin interactor that increases presenilin protein accumulation. J Cell Biol 151:847–862 [DOI] [PMC free article] [PubMed]
- 159.Massey LK, Mah AL, Ford DL, Miller J, Liang J, Doong H, Monteiro MJ (2004) Overexpression of ubiquilin decreases ubiquitination and degradation of presenilin proteins. J Alzheimers Dis 6:79–92 [DOI] [PubMed]
- 160.Massey LK, Mah AL, Monteiro MJ (2005) Ubiquilin regulates presenilin endoproteolysis and modulates gamma-secretase components, Pen-2 and nicastrin. Biochem J 391:513–525 [DOI] [PMC free article] [PubMed]
- 161.Zhou J, Liyanage U, Medina M, Ho C, Simmons AD, Lovett M, Kosik KS (1997) Presenilin 1 interaction in the brain with a novel member of the Armadillo family. Neuroreport 8:2085–2090 [DOI] [PubMed]
- 162.Levesque G, Yu G, Nishimura M, Zhang DM, Levesque L, Yu H, Xu D, Liang Y, Rogaeva E, Ikeda M, Duthie M, Murgolo N, Wang L, VanderVere P, Bayne ML, Strader CD, Rommens JM, Fraser PE, P St. George-Hyslop (1999) Presenilins interact with armadillo proteins including neural-specific plakophilin-related protein and beta-catenin. J Neurochem 72:999–1008 [DOI] [PubMed]
- 163.Stahl B, Diehlmann A, Sudhof TC (1999) Direct interaction of Alzheimer’s disease-related presenilin 1 with armadillo protein p0071. J Biol Chem 274:9141–9148 [DOI] [PubMed]
- 164.Shinozaki K, Maruyama K, Kume H, Tomita T, Saido TC, Iwatsubo T, Obata K (1998) The presenilin 2 loop domain interacts with the mu-calpain C-terminal region. Int J Mol Med 1:797–799 [DOI] [PubMed]
- 165.Periz G, Fortini ME (2004) Functional reconstitution of gamma-secretase through coordinated expression of presenilin, nicastrin, Aph-1, and Pen-2. J Neurosci Res 77:309–322 [DOI] [PubMed]