

Abstract
The maspin protein has tumor suppressor activity in breast and prostate cancers. It inhibits cell motility and invasion in vitro and tumor growth and metastasis in nude mice. Maspin is structurally a member of the serpin (serine protease inhibitors) superfamily but deviates somewhat from classical serpins. We find that single-chain tissue plasminogen activator (sctPA) specifically interacts with the maspin reactive site loop peptide and forms a stable complex with recombinant maspin [rMaspin(i)]. Major effects of rMaspin(i) are observed on plasminogen activation by sctPA. First, rMaspin(i) activates free sctPA. Second, it inhibits sctPA preactivated by poly-d-lysine. Third, rMaspin(i) exerts a biphasic effect on the activity of sctPA preactivated by fibrinogen/gelatin, acting as a competitive inhibitor at low concentrations (<0.5 μM) and as a stimulator at higher concentrations. Fourth, 38-kDa C-terminal truncated rMaspin(i) further stimulates fibrinogen/gelatin-associated sctPA. rMaspin(i) acts specifically; it does not inhibit urokinase-type plasminogen activator, plasmin, chymotrypsin, trypsin, or elastase. Our kinetic data are quantitatively consistent with a model in which two segregated domains of maspin interact with the catalytic and activating domains of sctPA. These complex interactions between maspin and sctPA in vitro suggest a mechanism by which maspin regulates plasminogen activation by sctPA bound to the epithelial cell surface.
Since the isolation of the human maspin gene from mammary epithelial cells (1), accumulating evidence supports its function as a tumor suppressor that inhibits motility, invasion, and metastasis (2–4). To explore the diagnostic and therapeutic potential of maspin, we have initiated investigations into the molecular mechanisms underlying its biological activities. Identification of the maspin target was the first fundamental objective.
Maspin is structurally a novel serine protease inhibitor (serpin) that shares sequence homology with other inhibitory serpins, including plasminogen activator inhibitor type-1 and -2 (PAI-1 and PAI-2), as well as noninhibitory proteins like ovalbumin (1). Functional studies demonstrate that recombinant maspin produced by baculovirus-infected insect cells [rMaspin(i)] acts at the cell membrane (3) and that its activity in inhibiting cell migration and invasion across a Matrigel matrix depends on an intact reactive site loop (RSL) (1–4). Based on this information, one may predict that maspin inhibits a cell surface serine protease, which has dual activities in promoting cell motility and invasion. An example of such a protease is urokinase-type plasminogen activator (uPA) in the uPA receptor/uPA system (5). PAI-1 and PAI-2, inhibitors of uPA, decrease cell invasion by inhibiting the proteolytic degradation of extracellular matrix (6, 7). Recent studies suggest that PAI-1 also inhibits cell motility via an integrin-mediated pathway by its interaction with cell surface-bound uPA and vitronectin (8, 9).
The RSL of maspin is shorter than that of other serpins (1), casting doubt on its ability to act as a classical inhibitory serpin. Pemberton et al. (10) reported that purified rMaspin(i) is a substrate instead of an inhibitor to an array of serine proteases, including uPA and tissue-type plasminogen activator (tPA). However, our evidence that the RSL is crucial for the biological activity of maspin would be consistent with maspin acting as an inhibitory serpin.
Here we provide evidence that biologically active rMaspin(i) specifically binds to purified single-chain tissue-type plasminogen activator (sctPA) and has biphasic effects on sctPA, activated to convert plasminogen to plasmin. rMaspin(i) had no effect on several other serine proteases, including uPA, under our assay conditions. Our model based on kinetic analyses provides insight into the dual interactions between rMaspin(i) and sctPA.
MATERIALS AND METHODS
Cell Culture and Materials.
Human mammary carcinoma MDA-MB-435 cells (American Type Culture Collection) were cultured in αMEM (GIBCO/BRL) supplemented with 5% fetal calf serum (HyClone) (11). When the cells were approximately 70% confluent, the medium was switched to DFCI-1 medium (12). Following another 24 hr of continued cell culture, the conditioned medium was collected. The SulfoLink coupling gel was purchased from Pierce. Maspin RSL peptide NH2-CIEVPGARILQHKDEL-COOH was synthesized and was HPLC-purified at the Molecular Biology Core Facility of the Dana–Farber Cancer Institute. rMaspin(i) was produced and purified as described (2). The 38-kDa N-terminal fragment of maspin resulting from limiting digestion by trypsin was a gift from LXR BioTechnology, Inc. (Richmond, CA) (10). sctPA, glutamate-type plasminogen (Glu-plasminogen), high molecular weight urokinase, chromogenic plasmin substrate Spectrozyme PL, chromogenic urokinase substrate Spectrozyme UK, inhibitory monoclonal antibody against sctPA, and defined DESAFIB-X des-AA-fibrinogen were purchased from American Diagnostica (Greenwich, CT). Unless otherwise specified, all other chemicals and reagents were purchased from Sigma.
Purification of Maspin-Binding Proteins with an Affinity Column.
The RSL affinity column was made according to instructions by the manufacturer. Briefly, 5 mg of the maspin RSL peptide in PBS at pH 7.4 was mixed with 1 ml of SulfoLink gel suspension. The gel mixture was incubated at room temperature for 1 hr and packed into a column (1 cm i.d.). The column was washed thoroughly with PBS before loading 50 ml of conditioned DFCI-1 medium from the MDA-MB-435 cell culture. The nonspecific binding proteins were removed by thorough washing with PBS until no protein was detected in the washing (13). The protein specifically bound to the column was eluted with 0.1 M glycine, pH 2.5, into 0.5-ml fractions that were simultaneously neutralized with 0.1 vol of 1.0 M Tris⋅HCl, pH 9.5.
Zymography.
Zymogram gels with incorporated gelatin substrate were prepared as described by Heussen and Dowdle (14). Plasminogen at a final concentration of 10 μg/ml was incorporated into the separating gel for the detection of plasminogen activator activity. Protein samples were loaded at indicated amounts and subjected to electrophoretic separation at 10 mA at 4°C. The gel was then washed twice with 2% Triton X-100 in double distilled H2O for 1 hr. The gel was subsequently incubated at 37°C in 0.1 M glycine, pH 8.3, for 2½ hr at 37°C, stained with 0.25% Coomassie brilliant blue R-250, and destained with a solution of methanol/acetic acid/water (1:1:8, vol/vol).
Formation of sctPA–Maspin Complex.
rMaspin(i) and sctPA were mixed at different molar ratios in 50 mM Tris⋅HCl, pH 7.5, containing 0.1% Triton X-100 in a final volume of 25 μl. Samples were incubated at 37°C for 15 min and denatured at 85°C in SDS/PAGE sample buffer for 2 min. The denatured protein samples were separated on 7.5% SDS/PAGE and subsequently transferred to poly(vinylidene difluoride) membrane. Maspin and complexes containing maspin were detected by Western blotting by using 1,000-fold diluted polyclonal antiserum against maspin, AbS3A, and ECL reagents (Amersham). AbS3A was designed to recognize amino acids 169–182 in the center of the maspin molecule (1).
In a control experiment, sctPA and ovalbumin were mixed at various molar ratios and incubated under the conditions described previously. Following SDS/PAGE and electrotransfer, ovalbumin was detected by Western blotting with the monoclonal antibody OVA-14 at 1:1,000 dilution (Sigma) and ECL.
Coupled Assay for Plasminogen Activation by Free sctPA.
A total of 150 μl of reaction mixture contained 0.2 mM Spectrozyme PL (a chromogenic plasmin substrate) and 0.4 NIH unit of sctPA in 50 mM Tris⋅HCl, pH 7.5/0.1% Triton X-100. No sctPA was added in the blank. Glu-plasminogen at a final concentration of 83 nM and rMaspin(i) were added to start the reaction. The absorbance of the reaction mixtures at 405 nm was monitored at 37°C over the next 2-hr period with a Bio-Rad model 3550 microplate reader. The initial velocity of plasmin production was determined from the plots of A405 vs. t2 based on the following equation, derived from the two step process of plasmin production plus activity:
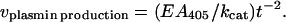 |
1 |
where A405 and E are the absorbance and absorption coefficient at 405 nm of the amidolytic product of Spectrozyme PL, t is time in min, and kcat is the rate constant for overall catalysis.
Coupled Assay for Plasminogen Activation by Immobilized sctPA.
A 96-well ELISA titer plate was coated with sctPA and poly-d-lysine at final concentrations of 2.7 NIH units/ml and 20 μg/ml, respectively, in 0.2 M bicarbonate buffer, pH 9.3, for 1 hr at 37°C. The blank was coated with poly-d-lysine only. The plate was subsequently washed three times with PBS, pH 7.2. A total of 150 μl of reaction mixture containing 0.2 mM Spectrozyme PL, 83 nM Glu-plasminogen, and various concentrations of rMaspin(i) in 50 mM Tris⋅HCl, pH 7.5/0.1% Triton X-100 was added to the coated wells to start the reaction. Initial velocities for the production of plasmin were calculated based on the changes in A405 by using Eq. 1.
sctPA Activity Assay in the Presence of Fibrinogen/Gelatin.
Spectrozyme PL at a final concentration of 0.2 mM was added to a preincubation mixture of 0.4 NIH unit of sctPA, fibrinogen (0.4 μM), and gelatin (133 μg/ml) in 50 mM Tris⋅HCl, pH 7.5/0.1% Triton X-100 (1 hr at 37°C). No sctPA was added in the blank. Glu-plasminogen at a final concentration of 83 nM and rMaspin(i) or trypsin-cleaved rMaspin(i) when indicated were added to start the assay. The final volume of each reaction was 150 μl. The absorbance of the reaction mixtures at 405 nm was monitored at 37°C over the next 2-hr period. To determine the dissociation constant of the sctPA/rMaspin(i) intermediate, the concentrations of plasminogen and rMaspin(i) were varied under the optimized sctPA assay conditions.
Activity Assays of Other Serine Proteases.
The amidolytic activities of uPA, sctPA, plasmin, elastase, chymotrypsin, and trypsin were assayed by using specific synthetic chromogenic substrates Spectrozyme UK (0.2 mM in 50 mM Tris⋅HCl, pH 8.0), Spectrozyme PL (0.2 mM in 50 mM Tris⋅HCl/0.1% Triton X-100, pH 7.5), N-methoxysuccinyl-Ala-Ala-Pro-Val p-nitroanilide (0.2 mM in 0.1 M Hepes/0.5 M NaCl, pH 7.5), N-succinyl-Ala-Ala-Pro-Phe p-nitroanilide (0.2 mM in 0.2 M phosphate buffer, pH 6.2), and N-succinyl-Ala-Ala-Pro-Phe p-nitroanilide (0.2 mM in 50 mM Tris⋅HCl, pH 7.5), respectively. The enzyme substrate and rMaspin(i) at a final concentration of 0.5 μM, when indicated, were added to start the assays. The activities of these enzymes were also determined with the coupled plasminogen activation assay used for sctPA.
RESULTS
Maspin RSL Peptide Specifically Binds sctPA.
MDA-MB-435 cells, which are inhibited by rMaspin(i) in both motility and invasion assays (3), were chosen as the starting material to find a maspin-binding protein by affinity purification. The DFCI-1 medium conditioned by these cells was put onto a maspin RSL peptide column. After a thorough wash with PBS, the column was eluted with 0.1 M glycine, pH 2.5, resulting in a single-peak elution profile. The amount of protein eluted accounts for approximately 0.25% of the starting material, and fractions 5 and 6 had the highest protein concentrations. SDS/PAGE followed by silver staining revealed a marked selective increase of a 70-kDa protein in fractions 5 and 6 as compared with the starting material and all the other elution fractions. Fractions 5 and 6 also had several minor components ranging from 14 to 90 kDa, the identities of which are yet to be determined (data not shown).
We tested the elution fractions for proteolytic activities by substrate-incorporated zymographic gel electrophoresis. On the plasminogen-dependent zymogram gel (Fig. 1A), five clear zones resulted from localized degradation of gelatin. Fraction 5 (and fraction 6, data not shown) gave rise to a clear zone of 70 kDa, consistent with the SDS/PAGE result and similar to the molecular mass of sctPA. As compared with the commercial sctPA, the proteolytic activity of fractions 5 and 6 was approximately 1 NIH unit/ml as estimated by reflection densitometry of the zymogram. The amount of tPA protein was estimated to be 15 ng/ml by a tPA-specific ELISA kit (American Diagnostica). All of the other eluted fractions were negative in these assays (data not shown). In addition, all of the elution fractions tested negative for plasminogen-independent protease and metalloprotease activities by the corresponding substrate-incorporated zymographic analyses (data not shown). Purified commercial human high molecular weight uPA, human sctPA, and porcine pancreatic elastase were used as positive controls and size references.
Figure 1.

(A) Purification of sctPA from MDA-MB-435-conditioned medium. Zymographic analysis of the plasminogen-dependent activity is shown. Lanes 1–5 are 15 μg of protein sample from MDA-MB-435-conditioned medium, 15 μg of protein of elution fraction 5, 0.45 NIH unit of high molecular weight uPA standard, 0.43 NIH unit of commercial sctPA standard, and 6.15 NIH units of elastase. (B) Dose-dependent inhibition of plasminogen activator activity by tPA-blocking monoclonal antibody. A coupled assay for plasminogen activation by free enzyme was performed. The rates of plasminogen activation by 0.4 NIH unit of pure sctPA (solid line) and by 15 ml of fraction 5 (dashed line) are normalized and presented as percentages of the corresponding positive controls in the absence of antibody. The error bars represent standard deviations from two parallel experiments.
To address whether the purification of sctPA from the maspin RSL peptide affinity column was because of an artificial interaction of sctPA with the supporting matrix or with other nonspecific proteins, a parallel purification experiment was performed in which 50 ml of MDA-MB-435-conditioned DFCI-1 medium was loaded to SulfoLink gel coupled with pure recombinant glutathione S-transferase. The resulting elution profile had one peak as shown by protein concentration assays. However, all fractions tested negative in either plasminogen-dependent or -independent zymograms (data not shown).
Coupled plasminogen activation assays further confirmed the presence of sctPA in fractions 5 and 6. As shown in Fig. 1B, fraction 5 (and fraction 6, data not shown) and the commercial sctPA standard lost their activities in a dose-dependent manner with an increasing concentration of the anti-tPA antibody. In contrast, the uPA standard was not inhibited by this antibody (data not shown). Furthermore, fresh DFCI-1 culture medium with 1% FCS had no detectable plasminogen activator activity, whereas the conditioned DFCI-1 medium from MDA-MB-435 cell culture exhibited a low activity, attenuated about 10-fold relative to fractions 5 and 6 (data not shown). Thus, our data indicate that the sctPA in fractions 5 and 6 did not originate from serum but rather was secreted from MDA-MB-435 cells.
rMaspin(i) and sctPA Form a Detergent-Resistant Complex.
Physical interaction between maspin and sctPA proteins is suggested by the purification of sctPA from the maspin RSL peptide affinity column. Quantitative ELISA also demonstrated a dose-dependent binding of pure sctPA to immobilized rMaspin(i) (data not shown). To address the question of whether maspin acts as a typical inhibitory serpin to form a stable complex with sctPA (15), mixtures of pure rMaspin(i) and sctPA in various molar ratios were denatured with SDS and analyzed by Western blotting with the maspin antibody AbS3A. As shown in Fig. 2A, when the molar ratio of rMaspin(i)/sctPA was between 1 and 5, a band was detected with the size of the combined molecular masses of sctPA and rMaspin(i), approximately 110 kDa. This band was not observed with sctPA alone (lane 6) or polymerized rMaspin(i) (lane 7). rMaspin(i) in these reaction mixtures was not significantly degraded as shown by a constant 42-kDa band. The detection of the 110-kDa band in Fig. 2A indicates the formation of a detergent-resistant sctPA–rMaspin(i) complex. Thus, it is likely that rMaspin(i) interacts with sctPA as a typical inhibitory serpin such as PAI-1 (16).
Figure 2.

Formation of a detergent-resistant complex between rMaspin(i) and sctPA. (A) Western blotting of sctPA–rMaspin(i) complex. The monomer form of rMaspin(i), dimer form of rMaspin(i), and commercial sctPA–rMaspin(i) complex had molecular masses of 42, 84, and 110 kDa, respectively. Lanes 1–6 are the mixtures of pure sctPA and rMaspin(i) at indicated molar ratios. Lane 7 is polymerized rMaspin(i) standard. (B) Western blotting of the sctPA–ovalbumin complex. Lanes 1–5 are the mixtures of sctPA and ovalbumin at indicated molar ratios.
Under the same conditions, no new band was seen with the mixture of pure sctPA and ovalbumin, a bona fide example of a noninhibitory serpin (Fig. 2B). As the molar ratio between sctPA and ovalbumin approached 1, ovalbumin was significantly degraded, unlike rMaspin(i), as shown by the disappearance of the intact 45-kDa ovalbumin and the concomitant appearance of a lower band of approximately 42–43 kDa.
rMaspin(i) Stimulates Free sctPA Activity but Inhibits Immobilized sctPA.
sctPA converts plasminogen to plasmin. To examine the effect of rMaspin(i) on this activity, we first employed the coupled plasminogen activation assay in the absence of sctPA cofactors (16). Surprisingly, rMaspin(i) had only a stimulatory effect on sctPA activity (Fig. 3A). sctPA activity was steadily elevated as the concentration of rMaspin(i) was increased until it approached a saturation level that was dependent on sctPA concentration (data not shown). This stimulatory effect of rMaspin(i) on sctPA was specific because ovalbumin and BSA had no effect under the same conditions (data not shown).
Figure 3.

The dose-dependent effect of rMaspin(i) on plasminogen activation by free sctPA activity (A) and by poly-d-lysine-coimmobilized sctPA (B). The initial velocities are presented as the rate of plasmin production in units of pM/min (⧫). The error bars represent the standard deviations of two parallel experiments. The dashed line represents calculated initial velocities by using Eq. 2 with α = 0.01 pM/min, β = 8.3 × 10−7/min, ξ = 0.425, ψ = 1.7 × 10−7 pM−1, and ζ = 1.0 × 10−16 pM−2 for A and α = 0.435 pM/min, ξ = 0.1, ψ = 4.0 × 10−7 pM−1, and ζ = 1.0 × 10−16 pM−2 for B.
It is well known that binding of proteins such as fibrin or fibrinogen greatly activates sctPA under physiological conditions (16). To examine the effect of rMaspin(i) on activated sctPA, we first screened several activating components by the coupled plasminogen activation assay. sctPA coimmobilized with poly-d-lysine at a final concentration of 20 μg/ml most effectively activated sctPA by approximately 20-fold (Fig. 3B). rMaspin(i) added to this preactivated sctPA inhibited the conversion of plasminogen to plasmin in a nearly linear dose-dependent manner, and at 1.0 μM, blocked sctPA activity by nearly 90%.
rMaspin(i) Exerts a Biphasic Effect on Fibrinogen/Gelatin-Associated sctPA.
Fibrin(ogen) is a physiological activator of sctPA in fibrinolysis reactions (17). To further characterize the biochemical properties of rMaspin(i) under a condition closer to in vivo, the effect of rMaspin(i) on the plasminogen activation by sctPA was examined in the presence of DESAFIB-X des-AA-fibrinogen, a defined fragment of fibrinogen that gives rise to linear stimulation of sctPA, used in combination with gelatin for a greater stimulation (data not shown). rMaspin(i) added along with plasminogen exhibited a biphasic inverse bell-shaped dose-response curve with a nadir at 0.5 μM rMaspin(i) (Fig. 4A). Under the same conditions, recombinant PAI-1 was a potent inhibitor and completely blocked sctPA activity at a final concentration of 0.025 μM (data not shown).
Figure 4.

(A) The dose-dependent effect of rMaspin(i) (○) and trypsin-cleaved maspin (•) on the plasminogen activation by fibrinogen/gelatin-activated sctPA. Dashed lines represent calculated initial velocities by using Eq. 2 with α = 0.21, β = 1.45 × 10−7/min, ξ = 0.1, ψ = 6.0 × 10−8 pM−1, and ζ = 1.0 × 10−16 pM−2 for rMaspin(i), and α = 1.01, β = 1.727 × 10−6/min, ξ = 0.425, ψ = 1.7 × 10−7 pM−1, and ζ = 1.0 × 10−16 pM−2 for trypsin-cleaved maspin. (B) Double-reciprocal plot of 1/v vs. 1/s for the plasminogen activation by fibrinogen/gelatin-activated sctPA at low concentrations of rMaspin(i). The substrate, S, is plasminogen. (C) Replot of the slopes of the double-reciprocal plot vs. the concentration of rMaspin(i). The intercept on the horizontal axis indicates the Ki value as 0.13 μM. The initial velocities are presented as rate of the production of plasmin in units of pM/min. The error bars represent the standard deviations of two parallel experiments.
The inhibitory effect of rMaspin(i) at low concentrations (<0.5 μM) is primarily because of its competition with plasminogen to bind sctPA, as shown by the double-reciprocal plots of 1/vinitial vs. 1/[plasminogen], which intercept close to the 1/v axis (Fig. 4B). The inhibitory effect of rMaspin(i) was inversely dependent on the concentration of plasminogen. At 0.5 μM, rMaspin(i) strongly blocked this plasminogen activation by sctPA by approximately 75%. The replot of the slopes of the double-reciprocal plots vs. rMaspin(i) concentration was linear. The Ki value deduced from this replot was 0.13 μM (Fig. 4C).
38-kDa C-Terminal Truncated rMaspin(i) Further Stimulates sctPA.
As plasminogen activation proceeds, rMaspin(i) cleaved by sctPA and by plasmin (10) may accumulate and further modify the activity of sctPA. To address this question, purified 38-kDa C-terminal truncated rMaspin(i), cleaved by trypsin and containing the entire sequence upstream of the P1 residue, was investigated for its effect on fibrinogen/gelatin activated sctPA. This polypeptide did not inhibit the preactivated sctPA but further stimulated it, suggesting that the N-terminal domain of cleaved (relaxed) maspin may have a higher affinity than fibrinogen/gelatin for the activation site of sctPA (Fig. 4A) and is responsible for the stimulatory interaction with sctPA. Thus, the cleaved product of rMaspin(i) is likely to exert a positive feedback on physiological plasminogen activation. On the other hand, maspin RSL peptide interacts specifically with sctPA on an affinity column and inhibits sctPA activated by fibrinogen/gelatin in our coupled assays (data not shown), suggesting a critical role of maspin C-terminal domain in the inhibitory interaction with the catalytic domain of sctPA. These data demonstrate that the opposing effects of rMaspin(i) on sctPA activity originate from two segregated functional domains.
rMaspin(i) Specifically Inhibits sctPA.
Inhibitory serpins with arginine at the P1 site usually crossinhibit a spectrum of trypsin-like serine proteases (18). The possible crossreactivity of rMaspin(i), which has arginine at its P1 position, with five commercial trypsin-like serine proteases (sctPA, high molecular weight uPA, plasmin, trypsin, and chymotrypsin) and elastase was examined both by the coupled plasminogen activation assay with fibrinogen/gelatin and by the optimal amidolytic activity assays with the corresponding synthetic substrate and no fibrinogen/gelatin. As summarized in Table 1, rMaspin(i) inhibited only the fibrinogen/gelatin-associated sctPA. The high specificity of rMaspin(i) toward sctPA is surprising and further demonstrates that maspin is a novel inhibitory serpin.
Table 1.
Effect of rMaspin(i) on proteolytic activities
| Protease | rMaspin(i)* | rMaspin(i)/ fibrinogen/gelatin† |
|---|---|---|
| sctPA | — | Ki = 0.13 μM |
| uPA | — | — |
| Plasmin | — | — |
| Chymotrypsin | — | — |
| Trypsin | — | — |
| Elastase | — | — |
A — indicates that rMaspin(i) had no detectable effect on proteolysis.
Assayed under the optimized condition for each corresponding enzyme in the absence of fibrinogen/gelatin.
Assayed under the optimized condition for sctPA in the presence of fibrinogen/gelatin.
DISCUSSION
The mammary tumor suppressor gene maspin encodes a specific inhibitor of sctPA as shown by (i) the specific affinity of sctPA for the immobilized maspin RSL peptide, (ii) the formation of a detergent-resistant sctPA–rMaspin(i) complex, and (iii) specific novel effects of rMaspin(i) on plasminogen activation by sctPA, acting as a stimulating factor when sctPA is free and as an inhibitor when sctPA is tightly associated with an activating factor. The biphasic effects of rMaspin(i) on sctPA derive from two segregated domains of the maspin molecule. The interplay between rMaspin(i) and other factors in the plasminogen activation by sctPA is complex and suggests a mechanism by which maspin modulates the cellular responses to physiological or pathological signals.
A simplified kinetic model and an equation derived from it (Fig. 5) may explain the novel effects of rMaspin(i) on the initial velocity of plasminogen activation by free and preactivated sctPA. Here we propose that rMaspin(i) is a kinetically bifunctional protein. The stimulatory effect of both rMaspin(i) on free sctPA (Fig. 3A) and of 38-kDa C-terminal truncated rMaspin(i) on preactivated sctPA (Fig. 4A) results from the predominant interaction of the N terminus of rMaspin(i) with the activation domain of sctPA. In contrast, when the activation domain of sctPA is tightly bound to another activating component (i.e., poly-d-lysine), rMaspin(i) only can use its RSL region to compete with plasminogen for the catalytic site of sctPA, in which case β in Eq. 2 (shown in Fig. 5) is 0. The effect of rMaspin(i) on sctPA preactivated by fibrinogen/gelatin (Fig. 4A) represents a case when rMaspin(i) can compete for both the catalytic and the activating domains of sctPA. rMaspin(i) at low concentrations (<0.5 μM) primarily competes with plasminogen for the catalytic site of sctPA, acting as an inhibitor (Fig. 4 B and C). As rMaspin(i) concentration is further increased, its stimulatory effect eventually overrides the inhibitory effect and becomes dominant. In this case, all the kinetic parameters in Eq. 2 appear. As shown in Figs. 3 and 4A, all the calculated initial velocities using Eq. 2 with different kinetic parameter settings match our experimental results. Thus this kinetic model is quantitatively consistent with our data.
Figure 5.

Models for interactions between rMaspin(i) and sctPA. Positions left and right of sctPA represent bindings to regulatory and catalytic sites, respectively. Ks are equilibrium constants and ks are rate constants. v and M in Eq. 2 represent the initial velocity of plasmin production in pM/min and the concentration of rMaspin(i) (or 38-kDa C-terminal truncated rMaspin(i)) in μM, respectively. α, β, χ, ξ, ψ, and ζ are kinetic constants.
The specific inhibitory effect of rMaspin(i) on sctPA but not on uPA or the other four serine proteases (Table 1) may underline the significance of the second kringle domain of sctPA in regulating plasminogen activation. sctPA is secreted as an active enzyme and is further strongly activated by specific interactions, primarily through its second kringle domain, with a wide variety of extracellular matrix components including fibrin(ogen) (19, 20), laminin (21), fibronectin (22), and lysine-rich molecules as shown here. Because sctPA activated by poly-d-lysine or fibrinogen/gelatin is inhibited by rMaspin(i), it is likely that the second kringle domain of sctPA plays a critical role in the fine modulation of its activity by both stimulators and inhibitors. It is worth noting that the biochemical homologue of sctPA, urokinase (uPA), has a single kringle domain, the function of which is not clear.
The plasminogen activation by sctPA also can be inhibited by PAI-1 (Ki = 0.033 μM (16)) and PAI-2 (Ki = 0.55 μM (23)). rMaspin(i), with a Ki of 0.13 μM, is comparable with PAI-2 and less effective than PAI-1 in inhibiting this reaction in vitro. However, based on our kinetic models for the interaction between rMaspin(i) and sctPA, these Ki values have little use in predicting the relative inhibitory potencies of these inhibitors in vivo unless other factors involved in the corresponding in vivo sctPA complex are identified and characterized. It is known that the interaction with extracellular matrix components may change the activities of both sctPA and PAIs (16, 24). Our data suggest that maspin may become a stronger inhibitor of sctPA if the activation domain of sctPA or the N-terminal domain of maspin is blocked.
The antiproteolytic activity of rMaspin(i) correlates well with its biological activities in inhibiting tumor invasion and metastasis. However, despite a few reports suggesting that tPA is involved in cell growth (25), cell motility, and invasion (26), the function of tPA in epithelial cell biology and tumorigenesis is not well understood. It is important to note that the biphasic effect of rMaspin(i) on fibrinogen/gelatin-activated sctPA coincides with its bell-shaped dose-dependent inhibition of cell invasion (2). Our preliminary data also show that sctPA stimulates the motility of mammary tumor cells, and this stimulatory effect is inhibited by rMaspin(i) (unpublished data). Extensive biological studies are under way in our laboratory to address whether the proteolytic inhibition of sctPA by maspin represents a general regulatory mechanism in cell invasion and motility.
In summary, we provide evidence here that the mammary tumor suppressor gene maspin specifically inhibits sctPA. Moreover, maspin represents a kinetically bifunctional serpin, the partition of which between the substrate form and the inhibitor form depends not only on its own structural features but also on the microenvironment of the target enzyme. The requirement of other sctPA-stimulating factors (e.g., fibrinogen or polylysine) in the inhibitory interaction between rMaspin(i) and sctPA may reflect a novel mechanism by which maspin regulates sctPA activity on the cell surface.
Acknowledgments
We thank Drs. Liliana Ossowski, Daniel Rifkin, Laszlo Lorand and David W. Christianson and Dana J. Witten for helpful discussions and critical reviews of the manuscript. This work was supported by National Institutes of Health Grant CA 61253 (to R.S.) and National Research Service Award Fellowship 5-T32-CA 09361-14 (to S.S.).
ABBREVIATIONS
- PAI-1 and PAI-2
plasminogen activator inhibitor type-1 and -2
- RSL
reactive site loop
- uPA
urokinase-type plasminogen activator
- tPA
tissue-type plasminogen activator
- rMaspin(i)
recombinant maspin produced by baculovirus-infected insect cells
- sctPA
single-chain tissue-type plasminogen activator
- Glu-plasminogen
glutamate-type plasminogen
Footnotes
Contribution of this paper was initiated by Ruth Sager and, after her death (March 1997), completed by Arthur B. Pardee.
References
- 1.Zou Z, Anisowics A, Hendrix M J C, Thor A, Neveu M, Sheng S, Rafidi K, Seftor E A, Sager R. Science. 1994;263:526–529. doi: 10.1126/science.8290962. [DOI] [PubMed] [Google Scholar]
- 2.Sheng S, Pemberton P A, Sager R. J Biol Chem. 1994;269:30988–30993. [PubMed] [Google Scholar]
- 3.Sheng S, Carey J, Seftor E A, Dias L, Sager R. Proc Natl Acad Sci USA. 1996;93:11669–11674. doi: 10.1073/pnas.93.21.11669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Sager R, Sheng S, Pemberton P, Hendrix M J C. Curr Top Microbiol Immunol. 1996;213:51–64. doi: 10.1007/978-3-642-61107-0_4. [DOI] [PubMed] [Google Scholar]
- 5.Ossowski L, Glunie G, Masucci M-T, Blasi F. J Cell Biol. 1991;115:1107–1112. doi: 10.1083/jcb.115.4.1107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Fazoli F, Blasi F. Trends Pharmacol Sci. 1994;15:25–29. doi: 10.1016/0165-6147(94)90130-9. [DOI] [PubMed] [Google Scholar]
- 7.Laugn W E, Cao X R, Yu Y B, Shimada H, Kruithof E K O. Cancer Res. 1993;53:6051–6057. [PubMed] [Google Scholar]
- 8.Stefansson S, Lawrence D A. Nature (London) 1996;383:441–443. doi: 10.1038/383441a0. [DOI] [PubMed] [Google Scholar]
- 9.Deng C, Curriden S A, Wang S, Rosenburg S, Loskutoff D J. J Cell Biol. 1996;134:1563–1571. doi: 10.1083/jcb.134.6.1563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Pemberton P A, Wong D T, Gibson H L, Kiefer M C, Fitzpatrick P A, Sager R, Barr P J. J Biol Chem. 1995;270:15832–15837. doi: 10.1074/jbc.270.26.15832. [DOI] [PubMed] [Google Scholar]
- 11.Band V, Zajchowski D, Swisshelm K, Trask D, Kulesa C, Cohen C, Connolly J, Sager R. Cancer Res. 1990;50:7351–7357. [PubMed] [Google Scholar]
- 12.Band V, Sager R. Proc Natl Acad Sci USA. 1989;86:1249–1253. doi: 10.1073/pnas.86.4.1249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bradford M M. Anal Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- 14.Heussen C, Dowdle E B. Anal Biochem. 1980;102:196–202. doi: 10.1016/0003-2697(80)90338-3. [DOI] [PubMed] [Google Scholar]
- 15.Bjork I, Nordling K, Larsson I, Olson S T. J Biol Chem. 1992;267:19047–19050. [PubMed] [Google Scholar]
- 16.Kruithof E K O, Vassalli J-D, Schleuning W-D, Mattaliano R J, Bachmann F. J Biol Chem. 1986;261:11207–11213. [PubMed] [Google Scholar]
- 17.Mullertz, S. (1956) Acta Physiol. Scand. 38, Suppl. 130, 1–66. [PubMed]
- 18.Lomas D A, Evans D L, Upton C, McFadden G, Carrell R W. J Biol Chem. 1993;268:516–521. [PubMed] [Google Scholar]
- 19.Grailhe P, Nieuwenhuizen W, Angles-Cano E. Eur J Biochem. 1994;219:961–967. doi: 10.1111/j.1432-1033.1994.tb18578.x. [DOI] [PubMed] [Google Scholar]
- 20.Downing A K, Driscoll P C, Harvey T S, Dudgeon T J, Smith B O, Baron M, Campbell I D. J Mol Biol. 1992;225:821–833. doi: 10.1016/0022-2836(92)90403-7. [DOI] [PubMed] [Google Scholar]
- 21.Moser T L, Enghild J J, Pizzo S V, Stack M S. J Biol Chem. 1993;268:18917–18923. [PubMed] [Google Scholar]
- 22.Stack M S, Pizzo S V. J Biol Chem. 1993;268:18924–18928. [PubMed] [Google Scholar]
- 23.Astedt B, Bladh B, Christensen U, Lecander I. Scan J Clin Lab Invest. 1985;45:429–435. doi: 10.1080/00365518509155239. [DOI] [PubMed] [Google Scholar]
- 24.Pennica D, Holmes W E, Kohr W J, Harkins R N, Vehar G A, Ward C A, Bennett W F, Velverton E, Seebury P H, Heyneker H L, Goeddel D V, Collen D. Nature (London) 1983;301:214–221. doi: 10.1038/301214a0. [DOI] [PubMed] [Google Scholar]
- 25.Petro G D, Copeta A, Barlati S. Exp Cell Res. 1994;213:286–294. doi: 10.1006/excr.1994.1200. [DOI] [PubMed] [Google Scholar]
- 26.Pittman R N, DiBenedetto A J. J Neurochem. 1995;64:566–575. doi: 10.1046/j.1471-4159.1995.64020566.x. [DOI] [PubMed] [Google Scholar]