

Abstract
Purpose
Botulinum toxin A (Botox) is commonly used for strabismus treatment. Although other muscles atrophy after intramuscular injection with Botox, extraocular muscles (EOMs) do not. A continuous process of myonuclear addition in normal uninjured adult myofibers in rabbit EOMs was studied. In this study, the effect of Botox-induced muscle paralysis on myofiber remodeling in adult EOMs was examined.
Methods
The superior rectus muscles of adult rabbits were each injected with 5 units of Botox. The contralateral muscle received injections of saline only. Bromodeoxyuridine (BrdU) was administered for various periods after Botox treatment, followed by various BrdU-free periods. Myonuclear addition, the number of BrdU-positive satellite cells, and the number of MyoD-positive satellite cells were quantified, as were alterations in expression of immature myosins.
Results
Intramuscular injection of Botox resulted in a significant increase in both the number of BrdU-positive myonuclei and satellite cells. MyoD expression in both satellite cells and myonuclei was significantly increased after Botox injection in EOMs. In Botox-treated EOMs, an increased number of myofibers positive for the neonatal myosin heavy chain (MyHC) isoform was detected in the orbital layer.
Conclusions
Botox-induced EOM paralysis resulted in a significant short-term increase in satellite cell activation and myo-nuclear addition in single myofibers in adult rabbit EOMs compared with control muscles. The appearance of MyoD-positive myonuclei suggests that protein synthesis becomes upregulated after Botox injection, and this, in turn, may help explain the minimal effects on myofiber size in EOMs after Botox injection. Understanding the effect of Botox on satellite cell activation and myonuclear addition in existing myofibers may suggest new ways to maximize the clinical effectiveness of Botox in patients with strabismus.
The direct intramuscular injection of botulinum toxin A (Botox) is a common treatment for numerous muscle contractility disorders, from cervical dystonia and blepharospasm1,2 to spasticity in patients with cerebral palsy.3 Botox, however, was initially developed as a treatment for strabismus and focal dystonias,4–6 and it remains an effective and widely used treatment for these conditions. A single injection of Botox results in a temporary paralysis of the neuromuscular junctions. Previous studies have demonstrated that, in the short term, paralysis with Botox results in terminal sprouting of the motor nerves.7–9 The time course of this nerve sprouting is muscle-dependent. In several botulinum-treated muscles, despite the return of muscle activity, the resultant new terminals are retained for more than a year.7,9 In other muscles, recovery of function at the original neuromuscular junction results in retraction of the sprouts and restoration of normal function to the original nerves.10 The duration of paralysis also seems to vary, lasting from 3 to 12 months,11 although the mean duration in most long-term studies is approximately 3 months.12–14 It appears that intrinsic differences between skeletal muscle types alter their responsiveness to and recovery from the effects of Botox injection.
The extraocular muscles (EOMs) appear to be unique in their response to Botox injection. In one study, reduction of force generation, particularly at low-frequency stimulation, is significant, although not complete, by 18 hours.15 Of note, no changes in muscle contraction velocity characteristics are evident, suggesting a functional effect on all muscle fiber types in the treated EOMs. Furthermore, in contrast to limb skeletal muscles, where muscle atrophy develops after Botox injection,16,17 little evidence of muscle atrophy is seen after Botox injection into the EOMs,18 despite evidence of the muscles’ rapid paralysis after treatment.15 Only the orbital singly innervated fiber type shows a structural change as a result of the Botox injections, and in fact these fibers are hypertrophied.18 A recent report demonstrated a long-term loss of only the EOM-specific myosin heavy chain (MyHC) isoform in rat ex-traocular muscle as a result of Botox injection.19 In addition, the proportion of other MyHC isoforms changes during the time of EOM muscle paralysis, with an initial shift toward slower isoforms and a slow return of faster isoforms. However, it appears that there are few long-lasting changes in the EOMs after Botox treatment.
The EOMs have several unique molecular and physiologic properties compared with other skeletal muscles and, in fact, have been considered a distinct allotype.20 One novel feature is the presence of activated satellite cells within adult, uninjured EOMs of rabbits, mice, monkeys, and humans.21–23 These satellite cells, unlike those of limb skeletal muscles, continuously divide in the normal, uninjured adult EOM and become randomly and stably integrated into existing EOM myofibers.21 It is possible that all skeletal muscles share this trait of continuous remodeling, but the EOMs appear to represent the most active end of this continuum. This process of myonuclear addition is balanced by a continuous process of myonuclear apoptosis, as visualized with TUNEL staining, and segmental cytoplasmic remodeling, as visualized by the presence of activated caspase-3 within defined cytoplasmic domains.24 We hypothesize that the minimal changes seen in EOMs after Botox treatment and the hypertrophy of specific fiber types18,19 can be accounted for by increases in satellite cell activation and myonuclear addition. The purpose of this study was to determine the effect of Botox treatment on myofiber remodeling.
Materials and Methods
Adult New Zealand White rabbits were obtained from Bakkon Rabbitry (Viroqua, WI) and housed with Research Animal Resources at the University of Minnesota. All studies were approved by the Institutional Animal Care and Use Committee, and followed the guidelines of the National Institutes of Health and the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research.
The rabbits were anesthetized with an intramuscular injection of ketamine and xylazine (1:1; 10 mg/kg and 2 mg/kg, respectively). Proparacaine drops were placed in the conjunctival cul-de-sac to reduce blinking. In the first group of rabbits, 5 units of Botox was injected under direct visualization into the belly of one superior rectus muscle. Injections were randomized; half of the Botox-injected muscles were on the right side and half on the left side. The contralateral muscle received an injection of an equal volume of saline only to serve as a control. A minimum of four muscles were examined for each experimental injection protocol.
Bromodeoxyuridine (BrdU) immunolabeling was used to visualize DNA synthesis and mitosis in adult EOMs. To compare this labeling with that in our previously published work,21 three postinjection BrdU-labeling protocols were used (Table 1, referred to in the text figure legends as, e.g., 3,8,10).
Table 1.
BrdU-Labeling Protocols
| Days Since Botox Injection | Days of BrdU | Days BrdU-Free | Total Days Since Botox Injection |
|---|---|---|---|
| 3 | 8 | 10 | 21 |
| 7 | 7 | 7 | 21 |
| 7 | 7 | 1 | 15 |
BrdU, in sterile isotonic saline, was injected intraperitoneally at a dose of 50 mg/kg body weight at the postinjection times and days indicated in Table 1. The rabbits were killed after the specified BrdU-free period, with an overdose of barbiturate anesthesia. The superior, lateral, medial, and inferior rectus muscles were removed, frozen in methylbutane chilled to a slurry in liquid nitrogen, and serially sectioned at 12 μm. The even-numbered sections were processed for expression of BrdU (1:100; Chemicon, Temecula, CA) and dystrophin (1:20; Novocastra-Vector Laboratories, Burlingame, CA) to ascertain whether the labeled nuclei were within the muscle sarcolemma and therefore were myonuclei. The odd-numbered sections were immunostained for BrdU and laminin (1:20; Sigma-Aldrich, St. Louis, MO) to ascertain whether the BrdU-labeled nuclei were within the muscle basal lamina and were satellite cells. For double-labeling experiments, sections were quenched with hydrogen peroxide, incubated with blocking serum, followed by biotin-avidin blocking reagent (Vector Laboratories), and every other section was incubated in primary antibody to either dystrophin or laminin. After a rinse in phosphate-buffered saline (PBS), the sections were reacted with a peroxidase avidin-biotin complex (ABC; Vectastain; Novacastra-Vector Laboratories). The peroxidase was developed with diaminobenzidine, and the sections were rinsed in PBS. For BrdU localization, the sections were incubated in 2 N HCl for 1 hour at 37°C, followed by neutralization in borate buffer and a PBS rinse. The sections were incubated in the primary antibody to BrdU. The sections were rinsed in PBS and incubated in reagents from the alkaline phosphatase ABC kit (Vector Laboratories) and reacted with the alkaline phosphatase black substrate kit. All BrdU myonuclear counts were performed on BrdU-dystrophin–labeled sections. The dystrophin or laminin immunostained brown and the BrdU-positive nuclei were black.
BrdU labeling and the number of myofibers were quantified in a masked fashion with the aid of a morphometry system (Bioquant Systems, Nashville, TN), by methods described previously.21,24 Briefly, the overall percentage of BrdU-positive myonuclei or cells in the satellite cell position that expressed BrdU, as identified by location between the dystrophin-positive sarcolemma and the laminin-positive basal lamina in serial sections, was determined as a percentage of all myofibers in cross-section. Cross-sections through control and treated EOMs from four rabbits were analyzed, to determine the labeling index for each of these antigens. These were calculated as percentages of the total number of labeled myonuclei or total number of labeled nuclei in the satellite cell position divided by the total number of myofibers counted. Counts were made in a minimum of four random fields in the orbital and global layers of four injected and four control muscles from each rabbit. Between 500 and 1500 myofibers were included in the total number of myofibers in cross-section, and four sections were counted for each muscle. Sections were chosen at random distances from the insertional tendon to the midbelly region of the analyzed muscles. The percentage positive was compared between the orbital and global layers and analyzed for statistical significance using either an unpaired two-tailed t-test or an analysis of variance (ANOVA) and the Dunn multiple comparison tests (Prism and Statmate software; Graph-Pad, San Diego, CA). An F-test was used to verify that the variances were not significantly different. Data were considered significantly different if P ≤ 0.05. Counts are expressed as the mean ± SEM.
Activated satellite cells were identified with an antibody to MyoD (1:25; Dako, Carpinteria, CA), according to standard methods of immunohistochemical localization. The number of MyoD-positive satellite cells was determined by counting four random fields in three cross sections of each muscle in both the orbital and global layers of Botox-treated and control muscles. Muscles from four rabbits were analyzed for each post-Botox injection interval. Data were analyzed as described for the EOM sections.
Immunohistochemical localization of neonatal and developmental myosin heavy chain isoforms (neonatal and developmental MyHC: 1:20; Novocastra-Vector Laboratories) was performed on randomly selected sections through the middle of the muscle and the tendon ends, using standard methods. Four fields in both the orbital and global layers were counted in sections prepared from four rabbits at each post-Botox interval. The data were analyzed as described for the EOM sections.
Results
Several BrdU-labeling periods were used to examine the relatively short-term changes in satellite cell activation and myonuclear addition that might occur after Botox injection into the EOM. In the first group of animals, daily intraperitoneal injections of BrdU were begun 1 week after a single injection of Botox into the superior rectus muscle, followed by euthanasia on day 15 (7,7,1). In these animals, there was a 2.5-fold increase in new myonuclei and a 4.5-fold increase in BrdU-positive satellite cells in both the orbital and global layers of Botox-treated EOM myofibers compared with saline-injected control specimens (Figs. 1, 2).
Figure 1.

Photomicrographs of a superior rectus muscle labeled with daily BrdU for 7 days 1 week after a single injection of Botox. BrdU is immunostained black and dystrophin, which demarcates the sarco-lemma, is immunostained brown. Horizontal arrow: a BrdU-positive myonucleus; vertical arrow: BrdU-positive cell in the satellite cell position. Bar, 50 μm.
Figure 2.
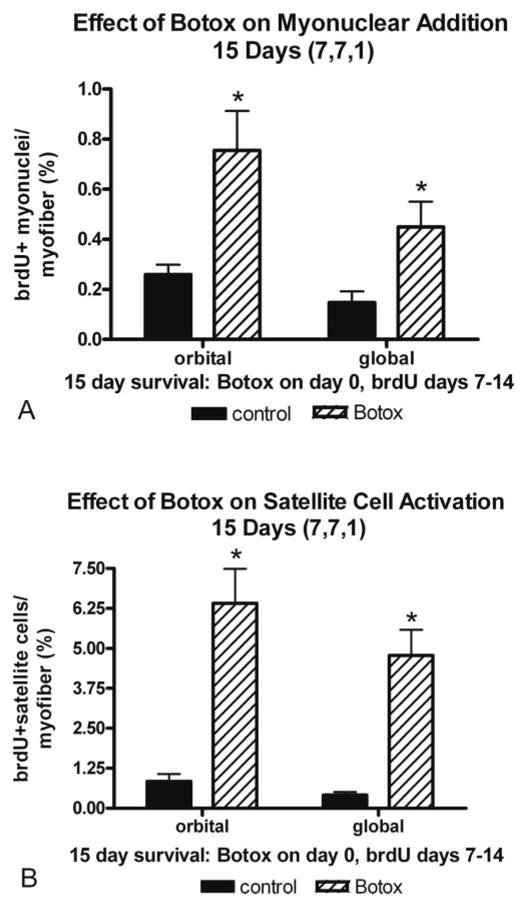
Quantification of (A) myonuclear addition (B) and satellite cell activation, as measured by BrdU incorporation. Botox was administered on day 0, BrdU was administered daily from days 7 to 14 after the Botox injection, and the animal was euthanatized on day 15 (7,7,1) *Significant difference from the saline-injected control.
To visualize the activation of satellite cells starting at an earlier time point after the Botox injection, BrdU was injected every day for 8 days starting 3 days after a Botox injection, followed by a 10-day BrdU-free period (3,8,10; Fig. 3). In these muscles, the number of BrdU-positive myonuclei in the orbital layer was unchanged compared with the noninjected control. However, in the global layer the number of BrdU-labeled myonuclei increased compared with the contralateral control. When BrdU-incorporation began 3 days after Botox injection, satellite cell division was elevated in both the orbital and global layers compared with the contralateral control muscles (Fig. 3).
Figure 3.
Quantification of (A) myonuclear addition (B) and satellite cell activation, as measured by BrdU incorporation. Botox was administered on day 0, BrdU was administered daily from days 3 to 8 after the Botox injection, and the animal was euthanatized on day 21 (3,8,10) *Significant difference from the saline-injected control.
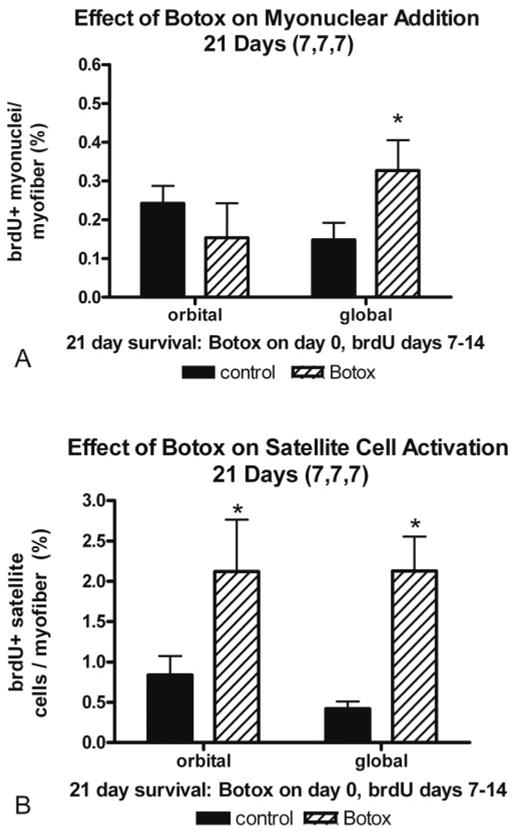
To give BrdU-labeled satellite cells a longer opportunity for potential fusion into existing myofibers, BrdU was injected every day for 7 days beginning 1 week after Botox injection, followed by a BrdU-free period of 7 days (7,7,7; Fig. 4). Satellite cells were significantly upregulated compared to the saline-injected control muscle. Of note, only the global layer showed a significant increase in myonuclear addition.
Figure 4.
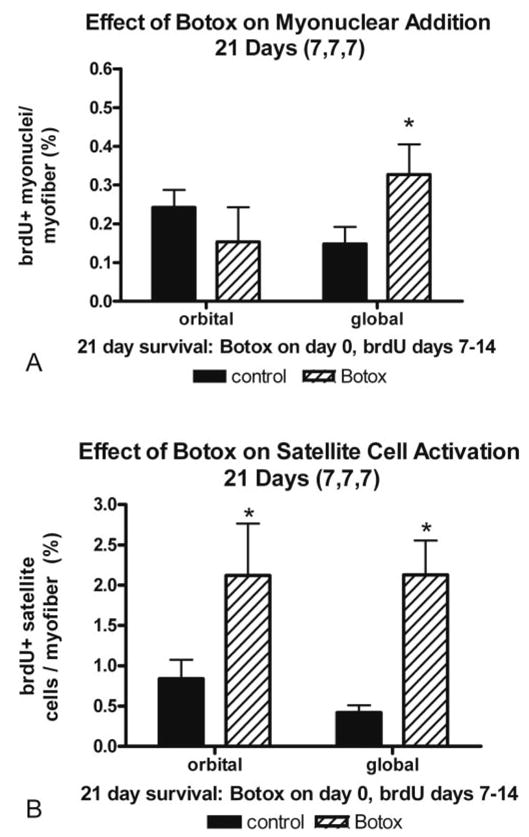
Quantification of (A) myonuclear addition and (B) satellite cell activation, as measured by BrdU incorporation. Botox was administered on day 0, BrdU was administered daily from days 7 to 14 after the Botox injection, and the animal was euthanatized on day 21 (7,7,7) *Significant difference from the saline-injected control.
Botox treatment of EOMs resulted in a significant increase in the percentage of activated satellite cells, as evidenced by expression of the myogenic regulatory factor MyoD. When compared with normal adult EOMs, there was a 9.3% and 7.3% increase in MyoD-positive cells in the orbital and global layers respectively (Figs. 5, 6). These cells tended to be randomly distributed throughout the treated muscle in much the same random distribution as in the normal control rectus muscles. In contrast to normal EOMs, MyoD-positive myonuclei were present (Figs. 5, 6).
Figure 5.
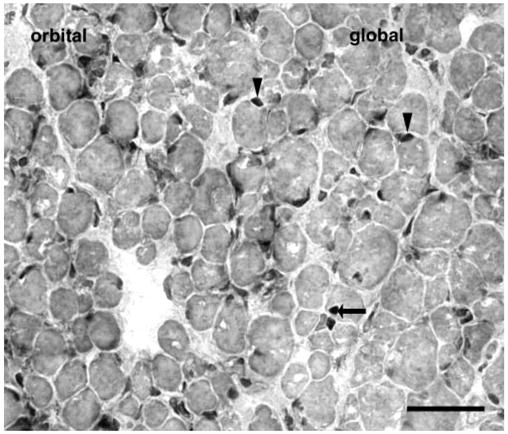
Injection of normal adult superior rectus with Botox resulted in a significant increase in the number of MyoD-positive satellite cells (arrowheads) compared with the normal EOM. Normally, the myonuclei in adult skeletal muscle are negative for MyoD expression. However, Botox treatment resulted in the appearance of MyoD-positive myonuclei, which were randomly located throughout the treated muscle (arrows).
Figure 6.
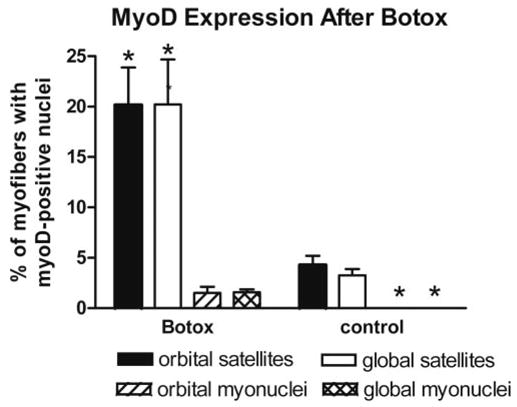
The number of MyoD-positive satellite cells and myonuclei was quantified in the rectus muscles. *Significant increase compared with the noninjected controls. **Essentially no MyoD-positive myonuclei in control superior rectus muscle.
Alterations in expression of the two immature MyHC isoforms, developmental and neonatal, were examined in the Botox-treated EOMs. The only significant alteration of MyHC isoform expression in the EOMs was an increase in neonatal MyHC expression in the orbital layer in the middle region of the Botox-treated rectus muscles, where the percentage of positive went from 8.4% ± 2.4% in control muscles to 45.34% ± 9.15% after Botox treatment. No significant difference was seen in expression of developmental MyHC.
Discussion
A single injection of Botox resulted in a significant short-term increase in satellite cell activation and myonuclear addition in adult rabbit EOMs. Concomitant with this, myonuclear expression of MyoD occurred. Few changes were seen in expression of neonatal and developmental MyHC 14 or 21 days after a single Botox injection in the EOMs; the orbital layer only showed an increase in neonatal myosin expression.
The effect of Botox on EOM activity is initiated very rapidly.15,25,26 Just 18 hours after an intramuscular injection of Botox into EOM, muscle force generation decreases to 93% compared with contralateral controls.15 Clinical doses of Botox result in paralysis of all myofiber types in the EOMs equally.15,27 Presynaptic axonal sprouting is initiated soon after Botox exposure and neuromuscular junction paralysis. One study demonstrated that axonal sprouting begins within 24 hours of muscle inactivity.28 By 8 days after Botox injection, evidence of renewed muscle activity due to neurite sprouting from the affected nerve is present.29,30 Thus, although muscle activity is reduced significantly after Botox administration, some small level of muscle activity appears to be retained almost continuously in the postinjection period. The rapid initiation of myonuclear addition and satellite cell activation in the present study correlates well with the rapid reaction of muscle to Botox. The initiation of sprouting within 24 hours of Botox administration may play a role in activating the satellite cells, as they are preferentially associated with myoneural junctions.31
Denervation results in upregulation of many cellular processes, presumably due to increased expression and release of various myogenic growth factors including hepatocyte growth factor (HGF), fibroblast growth factor (FGF), insulin-like growth factor (IGF), and ciliary neurotrophic factor (CNTF).32–34 In fact, experimentally induced upregulation of IGF-I accelerates muscle and motor neuron regeneration.33 There are a large number of growth factors that are normally upregulated in adult EOMs compared with adult limb muscle. These myogenic growth factors may play a role in the short-term satellite cell activation that has been documented after Botox treatment.24,35 Previous work has demonstrated that IGF activates satellite cells as the mechanism for muscle growth.36 A possible explanation for the lack of atrophy in the EOMs after Botox administration may be that growth factor release, known to cause muscle growth, would upregulate satellite cell activation and allow fiber size to be maintained. Studies are ongoing to examine the effect of total nerve removal on these processes. It may well be that this response will be similar with interventions, such as axotomy. It is possible that these activated satellite cells may enter the cell cycle but ultimately apoptose. Apoptosis is known to occur in a number of degenerative central nervous system disorders.37 It is also possible that some of the activated satellite cells return to quiescence.38
As identified by MyoD immunoreactivity, control EOMs contain approximately five activated satellite cells per 100 myofibers in cross section.21,23,24 Paralysis by Botox significantly increases the population of activated satellite cells within the 2 weeks after injection so that there are 20 to 30 activated satellite cells per 100 myofibers in cross section, an increase of five- to sixfold over control. From these numbers, it is clear that there is more than a sufficient number of activated satellite cells to account for the BrdU-positive myonuclei seen as a result of Botox treatment. Because Botox treatment is known to result in rapid initiation of neurite sprouting from the paralyzed nerves,39 it may be that the stimuli for both these processes are related. Surgical denervation of limb muscle also results in the upregulation of MyoD expression,40 and this increase is irrespective of myofiber type.41 However, the alteration in MyoD is activity independent. The presence of the denervated nerve in a spinal cord isolation model of injury is sufficient to retain MyoD expression in satellite cells in the limb muscles examined.40 Both MyoD and myogenin mRNA are significantly increased 7 days after Botox treatment of adult rat triceps muscle, but protein expression was not examined in this study.42 Botox treatment results in a significant increase in MyoD expression in the functionally denervated EOM, and this study showed that a population of these activated satellite cells become integrated into existing myofibers. These newly added myonuclei become stably integrated into the existing myofibers, and these labeled myonuclei are present 60 days after BrdU labeling.21
Botox treatment of the EOMs resulted in the appearance of MyoD-positive myonuclei, which are not present in normal EOMs. The upregulation of MyoD-expression in myonuclei is seen after hypertrophy induced by both muscle stretch and functional overload.43–45 In a model of stretch-induced muscle hypertrophy, the appearance of myonuclear MyoD correlates with increased protein synthesis in the stretched muscles.43 This upregulation of myonuclear MyoD in stretch-activated hypertrophy is independent of satellite cell activation and division.46 When growing limb muscle is treated with Botox, there are increased numbers of MyoD-positive nuclei, some of which appear to be myonuclei based on their published photomicrographs.47 This upregulation of MyoD-positive nuclei occurs even though significant myofiber atrophy occurred in these Botox-treated immature muscles. However, these Botox-treated muscles in growing rats are unchanged in terms of myonuclei per millimeter myofiber length, suggesting that myonuclear number and myofiber size may be under different control mechanisms.
Intramuscular Botox injection results in myofiber atrophy in almost all the muscles thus far examined.17,29,48 The only known exception appears to be the extraocular muscles, and the short-term absence of significant Botox-induced myofiber atrophy in the EOM has been clearly documented.18,19 In fact, in the first 35 days after injection of Botox into monkey medial rectus, there is an increase in the mean muscle fiber cross-sectional area specifically in the orbital singly innervated fiber type.18 In another study, the mean cross-sectional diameters are unchanged in the global layer at any time after Botox injection.19 This implies that the EOMs retain some mechanism that allows them to maintain myofiber cross-sectional area when functionally denervated. We hypothesize that the up-regulation of satellite cell activation, increased myonuclear addition, and upregulation of MyoD expression in myofiber nuclei may be responsible for maintenance of myofiber size in these muscles. As previously noted, MyoD-positive myonuclei appear in the Botox-treated muscles. If, as has been shown in other studies, upregulation of MyoD expression in adult myonuclei is associated with increased protein synthesis,43,46 this, in turn, would help maintain myofiber cross-sectional area in the extraocular muscles.
Few studies have examined alterations in MyHC isoform expression after Botox treatment in the EOMs. In one study, the EOM responded to Botox injection by short-term alterations in MyHC isoform expression, increasing the number of fibers positive for the slower isoforms.19 By 8 months after Botox injection, the proportion of myofibers expressing fast MyHCs increases, although they do not return to normal. The only long-term difference in the Botox-treated EOMs is the complete absence of expression of EOM-specific MyHC.19 In laryngeal muscles, which are very similar to EOMs in MyHC expression patterns, Botox injection results in an increased number of myofibers positive for the fast MyHC isoforms.49 In a study of gracilis muscle that is surgically denervated, increased expression of developmental MyHC and MyoD expression in satellite cells is seen; however, these changes were not quantified.50 No clear pattern of MyHC isoform changes has emerged from the studies thus far. In the present study, expression of only the developmental and neonatal MyHC isoforms was examined. The process of satellite cell activation and myonuclear addition suggested that there are modifications to other properties in the EOMs that are considered to be more “developmental” in nature. It is interesting that little significant change in the expression of these molecules was observed at the time points examined. As we have seen after EOM stretching, however, increased expression of the immature MyHC isoforms does not correlate with increased myonuclear addition in the single myofibers examined (Christiansen S, et al. IOVS 2004;45:ARVO E-Abstract 1585). It may be that signals controlling myonuclear addition are distinct from those controlling MyHC isoform composition, and the changes may be a result of denervation alone.
In summary, paralysis of extraocular muscle, as a result of Botox treatment, resulted in a significant short-term increase in satellite cell activation and myonuclear addition and expression of MyoD in myofiber nuclei within single myofibers in adult rabbit EOM compared with normal noninjected eye muscles. No concomitant changes in expression of immature MyHC isoforms were seen except in the orbital layer, suggesting that the mechanisms that control these two processes are distinct. Because functional denervation results in significantly increased myofiber remodeling compared with control EOMs, the upregulation of these processes may explain the minimal histologic and morphometric changes that occur after Botox administration in the extraocular muscles compared with limb muscles. Future studies will examine other possible factors, including release of trophic factors from nerves.
Footnotes
Supported by Grants EY13979 and EY11375 from the National Eye Institute, the Neuro-ophthalmology Research Fund, the Minnesota Medical Foundation (LKM), the Minnesota Lions and Lionesses, and an unrestricted grant to the Department of Ophthalmology from Research to Prevent Blindness, Inc.
Disclosure: I. Ugalde, None; S.P. Christiansen, None; L.K. McLoon, None
References
- 1.Scott AB, Kennedy RA, Stubbs HA. Botulinum toxin injection as a treatment for blepharospasm. Arch Ophthalmol. 1985;103:347–350. doi: 10.1001/archopht.1985.01050030043017. [DOI] [PubMed] [Google Scholar]
- 2.Brin MF, Fahn S, Moskowitz D, et al. Localized injections of Botulinum toxin for treatment of focal dystonia and hemifacial spasm. Mov Disord. 1987;2:237–254. doi: 10.1002/mds.870020402. [DOI] [PubMed] [Google Scholar]
- 3.Koman LA, Mooney JF, Smith B, Goodman A, Mulvaney T. Management of cerebral palsy with botulinum-A toxin: preliminary investigation. J Pediatr Orthop. 1993;13:489–495. doi: 10.1097/01241398-199307000-00013. [DOI] [PubMed] [Google Scholar]
- 4.Scott AB. Botulinum toxin injection into extraocular muscles as an alternative to strabismus surgery. Ophthalmology. 1980;87:1044–1049. doi: 10.1016/s0161-6420(80)35127-0. [DOI] [PubMed] [Google Scholar]
- 5.Scott AB, Rosenbaum A, Collins CC. Pharmacologic weakening of extraocular muscles. Invest Ophthalmol. 1973;12:924–927. [PubMed] [Google Scholar]
- 6.Elston JS, Lee JP, Powell CM, Hogg C, Clark P. Treatment of strabismus in adults with botulinum toxin A. Br J Ophthalmol. 1985;69:718–724. doi: 10.1136/bjo.69.10.718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Holland RL, Brown MC. Nerve growth in botulinum toxin poisoned muscles. Neuroscience. 1981;6:1167–1179. doi: 10.1016/0306-4522(81)90081-6. [DOI] [PubMed] [Google Scholar]
- 8.Angaut-Petit D, Molgo J, Comella JX, Faille L, Tabti N. Terminal sprouting in mouse neuromuscular junctions poisoned with botulinum type A toxin: morphological and electrophysiological features. Neuroscience. 1990;37:799–808. doi: 10.1016/0306-4522(90)90109-h. [DOI] [PubMed] [Google Scholar]
- 9.Holds JB, Alderson K, Fogg SG, Anderson RL. Motor nerve sprouting in human orbicularis muscle after botulinum A injection. Invest Ophthalmol Vis Sci. 1990;31:964–967. [PubMed] [Google Scholar]
- 10.De Paiva A, Meunier FA, Molgo J, Aoki KR, Dolly JO. Functional repair of motor endplates after botulinum neurotoxin type A poisoning: biphasic switch of synaptic activity between nerve sprouts and their parent terminals. Proc Natl Acad Sci USA. 1999;96:3200–3205. doi: 10.1073/pnas.96.6.3200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Tan EK, Jankovic J. Botulinum toxin A in patients with oromandibular dystonia: long-term follow-up. Neurology. 1999;53:2102–2107. doi: 10.1212/wnl.53.9.2102. [DOI] [PubMed] [Google Scholar]
- 12.Brin MF. Dosing, administration, and a treatment algorithm for use of botulinum toxin A for adult-onset spasticity. Spasticity Study Group. Muscle Nerve Suppl. 1997;6:S208–220. doi: 10.1002/(sici)1097-4598(1997)6+<208::aid-mus15>3.0.co;2-1. [DOI] [PubMed] [Google Scholar]
- 13.Defazio G, Abbruzzese G, Girlanda P, et al. Botulinum toxin A treatment for primary hemifacial spasm: a 10 year multicenter study. Arch Neurol. 2002;59:418–420. doi: 10.1001/archneur.59.3.418. [DOI] [PubMed] [Google Scholar]
- 14.Calace P, Coretese G, Piscopo R, et al. Treatment of blepharospasm with botulinum neurotoxin type A: long-term results. Eur J Ophthalmol. 2003;13:331–336. doi: 10.1177/112067210301300401. [DOI] [PubMed] [Google Scholar]
- 15.Dimitrova DM, Shall MS, Goldberg SJ. Short-term effects of botulinum toxin on the lateral rectus muscle of the cat. Exp Brain Res. 2002;147:449–455. doi: 10.1007/s00221-002-1265-8. [DOI] [PubMed] [Google Scholar]
- 16.Duchen LW. Changes in motor innervation and cholinesterase localization induced by botulinum toxin in skeletal muscle of the mouse: differences between fast and slow muscles. J Neurol Neurosurg Psychiat. 1970;33:40–54. doi: 10.1136/jnnp.33.1.40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hassan SM, Jennekens FGI, Veldman H. Botulinum toxin-induced myopathy in the rat. Brain. 1995;118:533–545. doi: 10.1093/brain/118.2.533. [DOI] [PubMed] [Google Scholar]
- 18.Spencer RF, McNeer KW. Botulinum toxin paralysis of adult monkey extraocular muscle: structural alterations in orbital, singly innervated muscle fibers. Arch Ophthalmol. 1987;105:1703–1711. doi: 10.1001/archopht.1987.01060120101035. [DOI] [PubMed] [Google Scholar]
- 19.Krancj BS, Sketelj J, D’Albis A, Erzen I. Long-term changes in myosin heavy chain composition after botulinum toxin A injection into rat medial rectus muscle. Invest Ophthalmol Vis Sci. 2001;42:3158–3164. [PubMed] [Google Scholar]
- 20.Porter JD, Baker RS. Muscles of a different “color”: the unusual properties of the extraocular muscles may predispose or protect them in neurogenic and myogenic disease. Neurology. 1996;46:30–37. doi: 10.1212/wnl.46.1.30. [DOI] [PubMed] [Google Scholar]
- 21.McLoon LK, Wirtschafter JD. Continuous myonuclear addition to single extraocular myofibers of uninjured adult rabbits. Muscle Nerve. 2002;25:348–358. doi: 10.1002/mus.10056. [DOI] [PubMed] [Google Scholar]
- 22.McLoon LK, Wirtschafter JD. Activated satellite cells are present in uninjured extraocular muscles of mature mice. Trans Am Ophthalmol Soc. 2002;100:119–123. [PMC free article] [PubMed] [Google Scholar]
- 23.McLoon LK, Wirtschafter JD. Activated satellite cells in extraocular muscles of normal adult monkeys and humans. Invest Ophthalmol Vis Sci. 2003;44:1927–1932. doi: 10.1167/iovs.02-0673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.McLoon LK, Rowe J, Wirtschafter JD, McCormick KM. Continuous myofiber remodeling in uninjured extraocular myofibers: myonuclear turnover and evidence for apoptosis. Muscle Nerve. 2004;29:707–715. doi: 10.1002/mus.20012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Moreno-Lopez B, Pastor AM, De La Cruz RR, Delgado-Garcia JM. Dose-dependent, central effects of botulinum neurotoxin type A: a pilot study in the alert behaving cat. Neurology. 1997;48:456–464. doi: 10.1212/wnl.48.2.456. [DOI] [PubMed] [Google Scholar]
- 26.Dengis CA, Steinbach MJ, Kraft SP. Registered eye position: short-and long-term effects of botulinum toxin injected into eye muscle. Exp Brain Res. 1998;119:475–482. doi: 10.1007/s002210050363. [DOI] [PubMed] [Google Scholar]
- 27.Stahl JS, Averbuch-Heller L, Remler BF, Leigh RJ. Clinical evidence of extraocular muscle fiber-type specificity of botulinum toxin. Neurology. 1998;51:1093–1099. doi: 10.1212/wnl.51.4.1093. [DOI] [PubMed] [Google Scholar]
- 28.Pamphlett R. Early terminal and nodal sprouting of motor axons after botulinum toxin. J Neurol Sci. 1989;92:181–192. doi: 10.1016/0022-510x(89)90135-4. [DOI] [PubMed] [Google Scholar]
- 29.Pinter MJ, Van den Noven S, Muccio D, Wallace N. Axotomy-like changes in cat motoneuron electrical properties elicited by botulinum toxin depend on the complete elimination of neuromuscular transmission. J Neurosci. 1991;11:657–666. doi: 10.1523/JNEUROSCI.11-03-00657.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Van Putten MJAM, Padberg M, Tavy DLJ. In vivo analysis of end plate noise of human extensor digitorum brevis muscle after intramuscularly injected botulinum toxin type A. Muscle Nerve. 2002;26:784–790. doi: 10.1002/mus.10274. [DOI] [PubMed] [Google Scholar]
- 31.Hawke TJ, Garry DJ. Myogenic satellite cells: physiology to molecular biology. J Appl Physiol. 2001;91:534–551. doi: 10.1152/jappl.2001.91.2.534. [DOI] [PubMed] [Google Scholar]
- 32.Siegel SG, Patton B, English AW. Ciliary neurotrophic factor is required for motoneuron sprouting. Exp Neurol. 2000;166:205–212. doi: 10.1006/exnr.2000.7528. [DOI] [PubMed] [Google Scholar]
- 33.Rabinovsky ED, Gelir E, Gelir S, et al. Targeted expression of IGF-1 transgene to skeletal muscle accelerates muscle and motor neuron regeneration. FASEB J. 2003;17:53–55. doi: 10.1096/fj.02-0183fje. [DOI] [PubMed] [Google Scholar]
- 34.Yamaguchi A, Ishii H, Morita I, Oota I, Takeda H. mRNA expression of fibroblast growth factors and hepatocyte growth factor in rat plantaris muscle following denervation and compensatory overload. Pflugers Arch. 2004;448:539–546. doi: 10.1007/s00424-004-1282-5. [DOI] [PubMed] [Google Scholar]
- 35.Fischer MD, Gorospe JR, Felder E, et al. Expression profiling reveals metabolic and structural components of extraocular muscles. Physiol Genomics. 2002;9:71–84. doi: 10.1152/physiolgenomics.00115.2001. [DOI] [PubMed] [Google Scholar]
- 36.Paul AC, Rosenthal N. Different modes of hypertrophy in skeletal muscle fibers. J Cell Biol. 2002;156:751–760. doi: 10.1083/jcb.200105147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Zhu X, McShea A, Harris PL, et al. Elevated expression of a regulator of the G2/M phase of the cell cycle, neuronal CIP-1-associated regulator of cyclin B, in Alzheimer’s disease. J Neurosci Res. 2004;75:698–703. doi: 10.1002/jnr.20028. [DOI] [PubMed] [Google Scholar]
- 38.Olguin HC, Olwin BB. Pax-7 up-regulation inhibits myogenesis and cell cycle progression in satellite cells: a potential mechanism for self-renewal. Dev Biol. 2004;275:375–388. doi: 10.1016/j.ydbio.2004.08.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Bonner PH, Friedli AF, Baker RS. Botulinum A toxin stimulates neurite branching in nerve-muscle cocultures. Brain Res Dev Brain Res. 1994;79:39–46. doi: 10.1016/0165-3806(94)90047-7. [DOI] [PubMed] [Google Scholar]
- 40.Hyatt JP, Roy RR, Baldwin KM, Edgerton VR. Nerve activity-independent regulation of skeletal muscle atrophy: role of MyoD and myogenin in satellite cells and myonuclei. Am J Physiol. 2003;285:C161–C173. doi: 10.1152/ajpcell.00128.2003. [DOI] [PubMed] [Google Scholar]
- 41.Ishido M, Kami K, Masuhara M. In vivo expression patterns of MyoD, p21 and Rb proteins in myonuclei and satellite cells of denervated rat skeletal muscle. Am J Physiol. 2004;287:C484–C493. doi: 10.1152/ajpcell.00080.2004. [DOI] [PubMed] [Google Scholar]
- 42.Witzemann V, Sakmann B. Differential regulation of MyoD and myogenin mRNA levels by nerve induced muscle activity. FEBS Lett. 1991;282:259–264. doi: 10.1016/0014-5793(91)80490-t. [DOI] [PubMed] [Google Scholar]
- 43.Lowe DA, Lund T, Alway SE. Hypertrophy-stimulated myogenic regulatory factor mRNA increases are attenuated in fast muscle of aged quails. Am J Physiol. 1998;275:C155–C162. doi: 10.1152/ajpcell.1998.275.1.C155. [DOI] [PubMed] [Google Scholar]
- 44.Gallegly JC, Turesky NA, Strotman BA, Gurley CM, Peterson CA, Dupont-Versteegden EE. Satellite cell regulation of muscle mass is altered at old age. J Appl Physiol. 2004;97:1082–1090. doi: 10.1152/japplphysiol.00006.2004. [DOI] [PubMed] [Google Scholar]
- 45.Ishido M, Kami K, Masuhara M. Localization of MyoD, myogenin and cell regulatory factors in hypertrophying rat skeletal muscles. Acta Physiol Scand. 2004;180:281–289. doi: 10.1046/j.0001-6772.2003.01238.x. [DOI] [PubMed] [Google Scholar]
- 46.Lowe DA, Alway SE. Stretch-induced myogenin, MyoD, and MRF4 expression and acute hypertrophy in quail slow-tonic muscle are not dependent upon satellite cell proliferation. Cell Tissue Res. 1999;296:531–539. doi: 10.1007/s004410051314. [DOI] [PubMed] [Google Scholar]
- 47.Chen CM, Stott NS, Smith HK. Effects of botulinum toxin A injection and exercise on the growth of juvenile rat gastrocnemius muscle. J Appl Physiol. 2002;93:1437–1447. doi: 10.1152/japplphysiol.00189.2002. [DOI] [PubMed] [Google Scholar]
- 48.Porter JD, Strebeck S, Capra NF. Botulinum-induced changes in monkey eyelid muscle. Comparison with changes seen in extraocular muscle. Arch Ophthalmol. 1991;109:396–404. doi: 10.1001/archopht.1991.01080030098047. [DOI] [PubMed] [Google Scholar]
- 49.Inagi K, Connor NP, Schultz E, Ford CN, Cook CH, Heisey DM. Muscle fiber-type changes induced by botulinum toxin injection in the rat larynx. Otolaryngol Head Neck Surg. 1999;120:876 –883. doi: 10.1016/S0194-5998(99)70330-X. [DOI] [PubMed] [Google Scholar]
- 50.Yoshimura K, Harii K. A regenerative change during muscle adaptation to denervation in rats. J Surg Res. 1999;81:139–146. doi: 10.1006/jsre.1998.5504. [DOI] [PubMed] [Google Scholar]