

Abstract
PSF (PTB-associated splicing factor) is a multi-functional protein that participates in transcription and RNA processing. While phosphorylation of PSF has been shown to be important for some functions, the sites and the kinases involved are not well understood. Although PSF does not contain a typical RS domain, we report here that PSF is phosphorylated in vivo to generate an epitope(s) that can be recognized by a monoclonal antibody specific for phosphorylated RS motifs within SR proteins. PSF can be phosphorylated by human and yeast SR kinases in vivo and in vitro at two isolated RS motifs within its N terminus. A functional consequence of SR phosphorylation of PSF is to inhibit its binding to the 3’ polypyrimidine tract of pre-mRNA. These results indicate that PSF is a substrate of SR kinases whose phosphorylation regulates its RNA binding capacity and ultimate biological function.
INTRODUCTION
The removal of intervening sequences (introns) from pre-mRNA (splicing) is catalyzed by the spliceosome. The formation of the spliceosome happens in a stepwise manner via the organization of small ribonucleoproteins (snRNPs), U1, U2, U4/U6 and U5 and non-snRNP proteins. Following U1 snRNP association with the 5’ splice site, SF1 and U2AF65 cooperatively bind to the branch point and polypyrimidine tract (Py) to recruit U2snRNP to form the A complex. The incorporation of tri-snRNPs, U4/U6.U5 is followed by spliceosome rearrangement to form the catalytic center, C complex [1, 2]. In higher eukaryotes, the Py of the 3′ splice site is highly conserved and recognized by several proteins, including U2AF65 and PSF [1, 3]. U2AF65, a member of the SR family[4], through recognition of the branch point and Py, can recruit U2 snRNA to base-pair with the branch sequence [3]. PSF is a 100 KDa protein, which was originally co-purified with Py binding protein (PTB) [5]. Subsequently, Patton et al. [6] found that PSF, but not PTB, could interact with the Py independently.
Several lines of evidence have shown that PSF is a multi-functional protein [7]. PSF is generally isolated in a complex [8] with a similar, multifunctional RNP protein, p54nrb/nonO [9, 10]. Both contain the DBHS (Drosophila Behavior; Human Splicing) domain, a region present in several other proteins that function in quite different ways to regulate gene expression [7, 9, 11]. In addition to its Py binding, PSF as a complex with p54nrb/nonO, binds to U5snRNA, indicating a role in spliceosome formation [12]. Its association with U4/U6.U5 tri-snRNP [12, 13] further indicates that PSF participates in the second catalytic step of splicing [14]. Transcription and pre-mRNA splicing are coupled by the C-terminal domain (CTD) of RNA polymerase II [15]. PSF and p54nrb/nonO bind strongly to both the hypo- or hyper-phosphorylated forms of the CTD in vivo and in vitro [16]. Strong transcriptional activators enhance levels of splicing and 3’-end cleavage, and this stimulation requires the CTD [17]. PSF but not p54nrb/nonO preferentially binds to strong activation domains to mediate transcriptional activator- and CTD-dependent stimulation of pre-mRNA processing [18]. As further evidence for a function in transcriptional initiation, PSF forms a ternary complex with the DNA-binding domains of several nuclear hormone receptors and with Sin3A to recruit class I histone deacetylases (HDACs) for repression [19]. Similarly, PSF and p54nrb/nonO interact with the steroidogenic factor (SF-1) to recruit Sin3A and HDACs to a repressive complex on the human CYP17 promoter [20]. In addition to these effects on splicing and transcription, the PSF-p54nrb/nonO complex appears to function as a double-stranded break rejoining factor [21]. PSF and p54nrb/nonO have been identified as components of a nuclear RNA retention complex for regulating late expression of polyoma RNAs [22]. Finally, PSF binding to a cis-acting regulatory element (INS) within Human Immunodeficiency Virus type I (HIV-1) gag/pol and env mRNAs leads to the degradation of INS-containing mRNA, whereas binding of p54nrb/nonO to the INS leads to nuclear export of INS-mRNAs into the cytoplasm [23].
The serine/arginine-rich proteins (SR proteins) are a highly conserved family and play roles in both constitutive and alternative pre-mRNA splicing [4]. Typically SR proteins contain at least one RNA recognition motif (RRM) and an RS-rich domain at the carboxyl-terminus. SR proteins can be extensively phosphorylated by kinases of the SRPK and Clk families [24] as well as by topoisomerase I [25]. RS phosphorylation occurs exclusively on serines within the RS motif. ‘Non-typical’ RS substrates which lack a defined RS domain have not been reported in mammalian cells. However, in yeast, the RGG-containing, RS tract-devoid RNA binding protein, Npl3, has been confirmed as a substrate for the SR kinase, Sky1 in vivo and in vitro [26, 27]. Not only do SR kinases have high specificity of their substrate proteins, but they also show preference toward certain RS context. Wang et al. [28] showed that SRPK2 has a stringent preference for RS dipeptides, and the consensus may be used to predict potential phosphorylation sites in candidate RS domain-containing proteins.
The degree of SR phosphorylation can regulate subcellular localization, protein-protein interactions and the splicing reaction itself [29]. SR proteins are well-known to coordinate protein-protein interactions by their RS domain and recruit other components of splicing [30]. For example, SR proteins interact with exonic splicing enhancer (ESE) sequences [29] and with the intronic branchpoint [31]. SR domains are required for mature spliceosome assembly [31, 32], for recruiting the U4/U6.U5 tri-snRNP into the spliceosome [33, 34], and for the second catalytic step of splicing [35]. In general, SR proteins have been shown to participate throughout the splicing reaction mediating both protein-RNA and direct RNA contacts throughout the splicing process. Simultaneous collaboration of multiple SR proteins may be required to remove a single intron [35, 36].
Both PSF and p54nrb/nonO are phospho-proteins [37-39, 40]. Tyrosine-phosphorylated PSF and p54nrb/nonO have been purified from the nuclear envelope of neuroblastoma cells, but the functional consequences were not investigated [37]. PSF and p54nrb can be phosphorylated in vitro by protein kinase C (PKC) [38, 39]. While this appeared to enhance PSF’s non-specific binding to DNA in vitro, no in vivo confirmation was provided [38]. Hyperphosphorylation of N-terminal serine/thronine residues in PSF was reported to alter its protein-protein interactions, but neither the kinase nor mechanism was identified [40].
Neither PSF nor p54nrb/nonO contains prototypic RS tracts, but both each contains scattered RS dipeptides. The participation of these two proteins in RNA processing and their multi-functional similarities to SR proteins prompted us to address this issue experimentally. Here we show that PSF can be phosphorylated in vivo to induce the formation of an epitope that can be recognized by an anti-SR antibody. PSF, but not p54nrb/nonO, can be phosphorylated in vitro by SR kinases, SRPK1 and DSK1 at an RS site within the N-terminus. SR phosphorylation of PSF inhibits its binding to the polypyrimidine tract of pre-mRNA introns. These results indicate that PSF is an unconventional substrate of SR kinases whose phosphorylation regulates its binding activity and its ultimate function in pre-mRNA splicing.
MATERIALS AND METHODS
Plasmid constructions
The cloning of full-length pET15b-PSF is described elsewhere [6]. PSF truncations (1-18, 28-47, 253-272, 331-350 and 400-419) were cloned into pGEX (Amersham) separately, and their inserts were amplified by the polymerase chain reaction (PCR) using pfu DNA polymerase (Stratagene). PCR fragments were cloned into BamHI and EcoRI sites of the vector separately. The ORF of ASF/SF2 was amplified by PCR using pfu DNA polymerase (Stratagene), and the amplified ASF/SF2 PCR fragment was subcloned into the BamHI and XhoI sites of pTrcHis (Invitrogen). pTrcHis-nonO was cloned by the same strategy. To clone pET29a-Dsk1, Dsk1 was amplified by PCR. The PCR product was digested with BamHI and XhoI and then subcloned into pET29a (Novagen). pRSETb-SRPK1 was a generous gift of Xiang-Dong Fu (University Of California, San Diego). p54nrb was obtained from Dr. Krainer (Cold Spring Harbor Laboratories). Glutathione-S-transferase (GST) plasmid is a commercial product (Amersham). To clone pCR3.1·PSF and pCR3.1-PSF (HA), the ORF of PSF was cloned into pCR3.1 (Invitrogen). Then, an HA tag was created at the 3′ end of the ORF of PSF.
In vitro phosphorylation assay
Protein substrates were purified and separately incubated at 1μM with purified SRPK1, 0.5μM, or Dsk1, 0.5μM, in a total volume of 20 μl containing a kinase buffer (50mM Tris-HCl buffer, pH7.4, 10mM MgCl2, 1mM dithiothreitol) in the presence of 50 μM ATP and 2μCi of [γ-32P]ATP at 23°C for 30 min. The reaction was terminated by boiling in SDS-PAGE sample loading buffer. Samples were fractionated on an 8% or a12.5% SDS-polyacrylamide gel, and protein phosphorylation was detected by autoradiography [41].
Purification of recombinant proteins
SRPK1, Dsk1, and putative substrates (nonO, p54nrb, PSF, and ASF/SF2) were each expressed as N-terminally, 3XHis-tagged proteins. Bacterial strains of interest were grown to an OD600 of 0.5-0.7 at 37°C in LB containing ampicillin, kanamycin or both. Cultures were induced with IPTG. The harvested cells were washed with PBS and resuspened in HKI buffer [42] and sonicated. The lysate was centrifuged at 5000 rpm for 25min in an HS4 rotor (Sovall) to separate supernatant and cell debris. The supernatant was incubated with Ni-NTA beads (Qiagen) for 1.5hr at 4°C. The beads were spun down and washed 3 times with HKI buffer. The proteins were eluted with HKI buffer containing 250 mM imidazole. Purified proteins were dialyzed against buffer D (20 mM HEPES-KCl [pH7.9], 100 mM KCl, 0.2 mM EDTA, 20% glycerol (v/v), protease inhibitors) before being aliquoted and frozen at -70°C. Strains expressing GST-fusion proteins were cultured and induced in the same manner, but purified on glutathione-agarose beads in HKE buffer [42] with 5mM of reduced glutathione.
Bacterial co-transformation was performed as described elsewhere [43]. Briefly, proteins of interest were constructed with 6xHis-tagged vectors (ampicillin resistant), where as the kinase, pET29a-Dsk1 (kanamycin-resistant) or pET29a-SRPK1 lacked the 6xHis-tag. Following drug selection and minidigest-confirmation of co-transformed single colonies, the His-tagged proteins were purified on Ni-NTA beads as described earlier.
Expression of PSF in insect cells
To express PSF in insect sf21 cells, a SalI-XhoI fragment, containing the full-length ORF of PSF was ligated into the SalI-XhoI sites of the transfer vector, pFASTBACHTc (Gibco, BRL). Dsk1 was cloned into the transfer vector, pFASTBAC, using the same strategy. Recombinant viral DNA was generated with the BAC-To-BAC HT Expression System (Gibco, BRL) according to the manufacturer’s instructions. Recombinant viral DNAs were transfected into sf21 cells by using Cellfectin (Gibco, BRL). Supernatants of transfected cultured cells were collected as recombinant virus stocks. Large-scale infections were performed by infecting 100 ml of sf21 cells (∼106/ml) in flasks with the virus stock of either PSF alone or PSF with Dsk1. The cells were harvested after 48-72 hrs of infection. PSF protein was purified by virtue of its His-tag protein as described above.
Antibodies
For anti-PSF antisera, full-length PSF with 6xHis was purified from E. coli and used for immunizing a rabbit. The rabbit blood was collected six weeks following initial injection and two boosts. Derivation of the SR phosphorylation-specific monoclonal antibody, mAb3C5 was described elsewhere [41, 44, 45] and was generously provided as ascites fluid by Dr. Turner (University of Birmingham, Birmingham, U.K.). Anti-HA monoclonal antibody (clone 12CA5) was purchased from Roche Molecular Biochemicals. The strain of mAb104 [46] was obtained from ATCC. The supernatant of mAb104 cell culture was used without dilution.
Transient transfection and immunoprecipitation
COS-7 cells were transfected by lipofectin using FuGene6 (Roche Molecular Biochemicals) according to manufacturer’s recommendations. 100 μl of serum-free DMEM (GibcoBRL) was added into 3 μl of FuGene6 transfectin reagent drop-by-drop and incubated at room temperature for 5 min. The diluted FuGene6 transfectin reagent was then added to 1 μg of cDNA drop-by-drop and incubated at room temperature for 15 min. The resultant FuGene-cDNA mixture was added to one 100 mm plate of COS-7 cells with 2 ml of fresh medium. At 48 h post-transfection, the transfected cells of each well were harvested and lysed with 150 μl of LOIPB buffer (25 mM Tris·HCl [pH7.8], 150 mM NaCl, 2 mM EDTA, 0.5% NP40 (v/v)). The lysates were spun and the supernatants were pre-cleared with protein A beads. 400 μl of the cell extracts was incubated with 4 μl of anti-HA monoclonal antibody for an hour at 4°C. 30 μl of protein A beads were added and incubated at 4°C for 30min. The bead complex was collected, washed 3 times with LOIPB buffer, boiled in SDS sample buffer, and then loaded onto a SDS polyacrylamide gel. Protein samples were transferred to a PVDF membrane, and western blots were performed using mab3C5 and anti-PSF polyclonal antisera. Enhanced chemiluminescence (ECL) Western blotting (Amersham Corp.) was performed to detect the protein samples as the manufacturers’ instructions.
RNA substrate preparation and in vitro RNA-protein gel shift
All RNA oligos were made by Oligo Therapeutics Inc. Synthesized RNA oligos, b/p+Py (NT4) (5′-UGCUGACCCUGUCCCUUUUUUUUCCACAGCUCG), which contains branch sequence (b/p) (UGC UGAC) and polypyrimidine tract (Py) (CCCUUUUUUUU), and scrambled branch sequence (NT3) (AUGCCCAUCGACCUCAAUAAU), were kinased with [γ-32p]ATP for 45min. The samples were mixed with RNA loading buffer (98% formamide, 10 mM EDTA, 0.1% xylene cyanol (v/v), 0.1% bromophenol blue (v/v)), and loaded onto a urea-6% polyacrylamide gel. Purified labeled oligos were eluted and ethanol precipitated. Different concentrations of protein (0.5 to 2 μM) were incubated with radio-labeled RNA oligos (∼1 ng) in binding buffer (20 mM HEPES-KCl [pH7.9], 100 mM KCl, 1 mM DTT, 1u/μl ribonuclease (RNase) inhibitor (Life Technologies), 0.2 mM EDTA,0.5mg/ml tRNA and protease inhibitors (Roche Molecular Biochemicals) for 25min at 30°C. Radiolabeled RNA-protein complexes were analyzed on 0.5X TBE 6% native polyacrylamide gels [47].
RESULTS
PSF expressed in mammalian cells was recognized by phospho-RS specific antibody
To test whether PSF is phosphorylated by SR kinases, we first expressed the protein in COS-7 monkey kidney cells and asked whether the expressed protein could be recognized by phosphor-RS specific antibodies such as mAB3C5 [41, 44]. The full-length cDNA of human PSF was cloned into a mammalian cell expression vector, pCR3.1, which utilizes a CMV promoter for ectopic over-expression. An HA tag was inserted at the C terminus of the PSF ORF (pCR3.1·PSF-HA) for immunoprecipitation purposes. Either PCR3.1, pCR3.1·PSF or pCR3.1·PSF-HA was transiently transfected into Cos-7 cells. After 48 hr, whole cell lysates were prepared and subjected to anti-HA immunoprecipitation followed by western blotting. In the lysates of pCR3.1·PSF-HA -transfected cells, PSF-HA is recognized by both anti-PSF (Fig 1A, lane 3) and mAb3C5 (Fig 1B, lane 2), whereas no protein is recognized by either antibody in the lysates of mock-transfected cells (Fig 1A, lane 2; Fig 1B, lane 1). To confirm that the immunoprecipitation was specific for the HA-tag, we included the lysates of untagged pCR3.1·PSF-transfected cells in the western blotting against anti-PSF. No band was detected in this sample (Fig 1A, lane 1). These results indicate that an epitope is created on PSF expressed in mammalian cells that is recognized by a monoclonal antibody that is specific for phosphorylated RS motifs.
Figure 1.
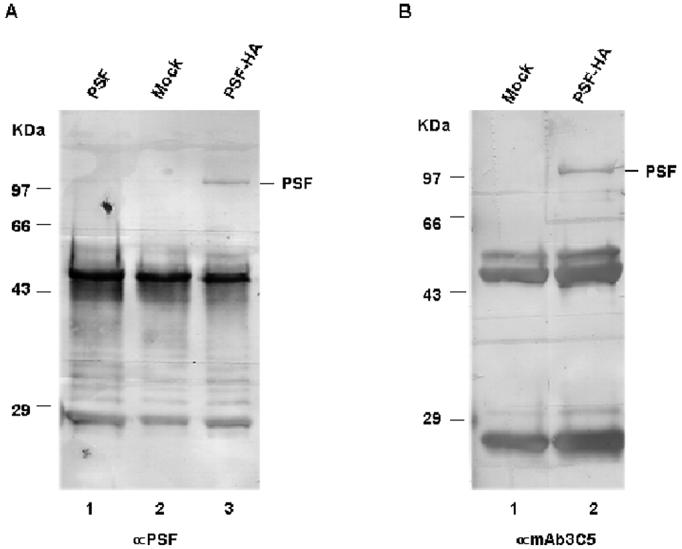
PSF can be recognized by SR phosphorylation-specific mAb 3C5 in mammalian cells. HA-tagged PSF (pCR3.1·PSF-HA), untagged PSF (pCR3.1·PSF) or the empty expression vector (mock) was transiently transfected into Cos-7 cells. After 48 hrs, supernatants from whole cell lysates were collected and immunoprecipitated with an anti-HA monoclonal antibody. Samples were resolved on 10% SDS-PAGE, transferred to a PVDF membrane, and then western blotting was performed with anti-PSF (A) or mAb 3C5 (B). Bands at ∼50 and 25 kD in all samples result from reaction of the anti-mouse secondary antibody against heavy and light immunoglobulin chains.
SR Kinases are sufficient to phosphorylate PSF in E. coli
Since PSF was recognized by phospho-RS specific antibody in mammalian cells, we decided to test in bacteria whether a prototypic SR protein kinase is necessary and sufficient (in the absence of other eukaryotic-specific co-factors) to phosphorylate PSF. Full-length His-tagged versions of PSF, ASF/SF2, and nonO were separately transformed and expressed in E. coli either alone or together with Dsk1 or its human orthologue, SRPK1 (PSFD or PSFS, respectively, in Figs. 2A and B). The proteins were purified and then tested by western blotting with mAb3C5. The prototypic RS protein, ASF/SF2, when co-expressed with Dsk1, was strongly recognized by mAb3C5 (Fig. 2A, lane 7), indicating that no other eukaryotic components were required. Likewise, PSF, co-expressed with either Dsk1 (PSFD, Fig. 2 A and B. lanes 5) or with SRPK1 (PSFS, Fig. 2B, Lane 6) was bound by mAb3C5, whereas PSF expressed alone was not (Fig. 2A and B, lanes 4). On the other hand, p54nrband nonO did not react with the SR phosphorylation-specific mAb3C5 when co-expressed with the kinase (Fig 2A, lane 3 and data not shown).
Figure 2.
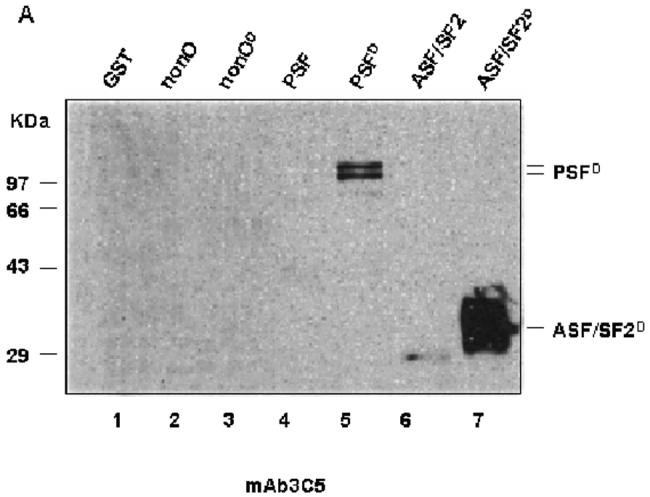
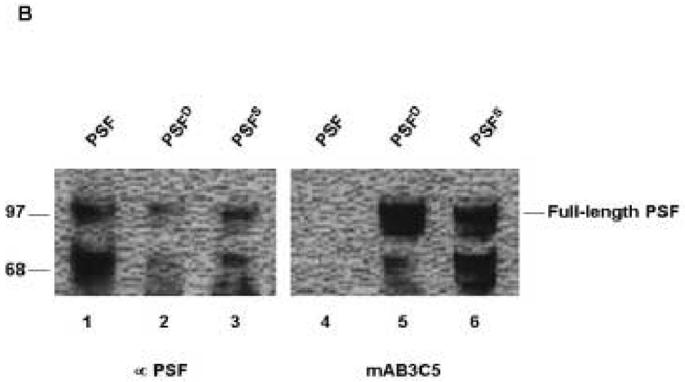
PSF co-expressed with SR kinases in bacteria can be recognized by SR phosphorylation-specific mAb3C5. (A). 2 μM of GST, or His-tagged proteins (nonO, PSF or ASF/SF2) or the same proteins co-expressed with Dsk1 (nonOD, PSFD or ASF/SF2D) were resolved by 10% SDS-PAGE, transferred to PVDF membranes, and then western blotted with mAb3C5. (B). 2 μM of purified PSF alone or PSF co-expressed with Dsk1 (PSFD) or SRPK1 (PSFS) were separated as above and filters were blotted with anti-PSF (left panel) or mAb3C5 (right panel). The reactive species at 68 kD is a proteolytic fragment of PSF previously observed at high abundance [14].
Our results indicate that in E coli, SRPK1 or Dsk1 is sufficient to create a phospho-epitope(s) for PSF that can be recognized by mAb3C5. However, we observed significantly reduced intensity for PSF relative to ASF/SF2 (Fig. 2A) under conditions of equal protein loading (data not shown). This indicates that the interaction between PSFD and this monoclonal antibody is weaker than that of ASF/SF2. This result might reflect the scarcity of reactive RS dipeptides within PSF or that the fraction of PSF phosphorylated is less. The faster migrating band of PSF of 68 kD, which was routinely recognized by both anti-PSF and mAb3C5 (Fig. 2B and re-addressed below), is very likely a N-terminal-containing, proteolytically-truncated form of the same size that was previously reported in mammalian cells [14].
Taken with the results of Figs. 1, PSF appears to be a substrate for the SR kinases, Dsk1 and SRPK1, and the phosphorylation of PSF generates a conformational epitope similar to that found in conventional SR proteins.
PSF is phosphorylated by SRPK1 and Dsk1 in vitro
SR proteins are phosphorylated by human SRPK1 and fission yeast Dsk1 in a highly specific manner [4, 41]. To determine whether PSF is a substrate of these two SR kinases, we performed in vitro phosphorylation assays. SRPK1 or Dsk1 was incubated with PSF, ASF/SF2, nonO or GST proteins separately. As with the conventional substrate ASF/SF2 (Fig. 3A and B, lanes 7), we found that PSF can be phosphorylated by either Dsk1 (Fig. 3A, lane 5) or SRPK1 (Fig. 3B, lane 5), albeit to a lesser extent. NonO and its human orthologue p54nrb share high sequence identity with PSF, including an RSR tripeptide motif immediately N-terminal to the RRMs. However, neither nonO (Fig. 3A and B, lanes 3) nor p54nrb (data not shown) were phosphorylated by SRPK1 or Dsk1.
Figure 3.
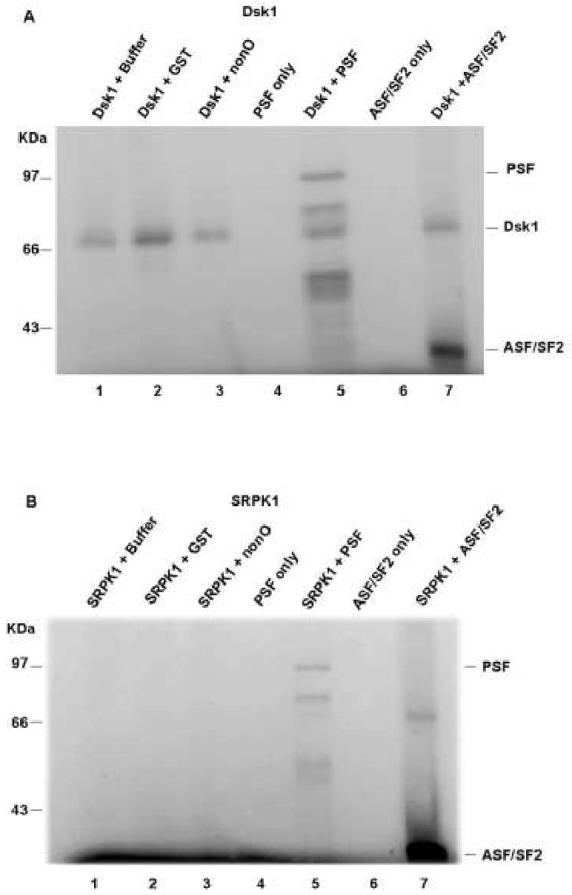
In vitro phosphorylation of PSF by SRPK1 or Dsk1. 1μM of the indicated bacterially expressed and purified recombinant proteins were incubated with0.5 μM Dsk1 (A) or with the same concentration of SRPK1 (B) in the presence of [γ-32P]ATP at 23°C for 30 min. Samples were resolved by 10% SDS-PAGE and visualized by phosphoimaging. Positions of phosphorylated PSF and autophosphorylated kinase are indicated.
It was previously shown that autophosphorylation of Dsk1 is constitutive and occurs prior to substrate protein phosphorylation [41]. Autophosphorylation of SRPK1, on the other hand, is concentration-dependent [24]. Thus we suspect that the ∼70kD band in lanes containing Dsk1 is autophosphorylated Dsk1. We suspect that the ∼66kD band in Lane 7 of Fig.3B, as well as the very faint equivalently-sized band in other lanes that contain SRPK1, is either a bacterial contaminant, which accompanies SRPK1 purification, or a degradation product of SRPK1 that is particularly stable in the presence of the substrate ASF. Regardless, the results strongly indicate that PSF, but not its highly similar partner, p54nrb/nonO, is an in vitro substrate of both SR kinases.
An isolated RS dipeptide near the N-terminus of PSF is the target of SR phosphorylation
PSF does not contain a conventional RS domain [4], but it contains 5 interspersed RS dipeptides. To test whether one or more of these is a genuine RS dipeptide responsible for the phosphorylation by SRPK1, we constructed GST-fusions of regions spanning each RS dipeptide, then expressed and purified each of them from E Coli (Fig. 4A). Only the N-terminal fragment (residues 1-18) was phosphorylated above background levels (Fig. 4B). Mutation of the RS dipeptide (residues 7 and 8 to LG) within the N-terminal region eliminated SRPK1 phosphorylation (Fig. 4C). That 7-RS-8 is the only site of SR phosphorylation was confirmed by the lack of reactivity of mAbC35 against truncation mutants (26-707 and 313-707) that lack this region but contain all other RS dipeptide candidates (data not shown). Thus, this isolated motif can convey SR kinase substrate specificity and anti-SR antibody immunological specificity to PSF.
Figure 4.
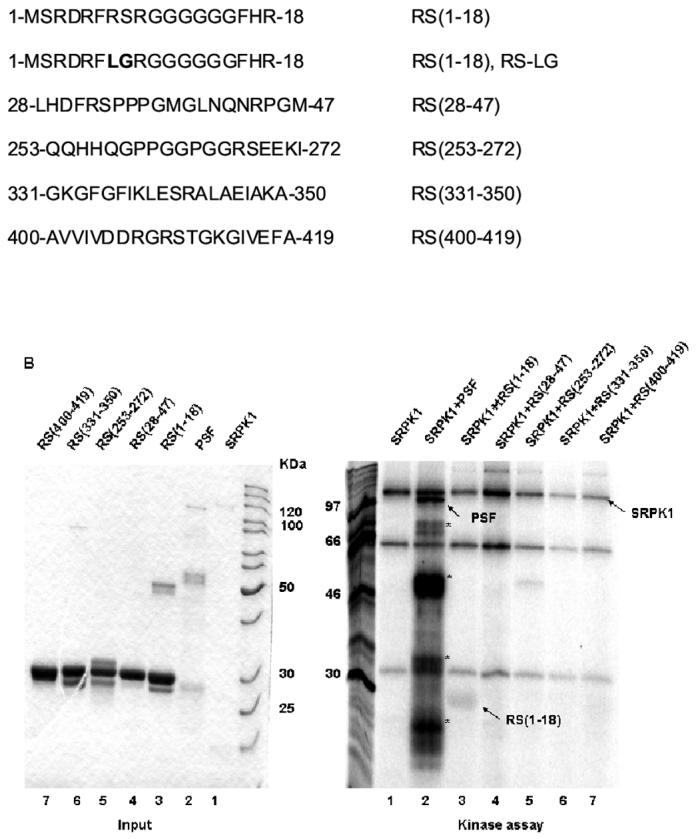
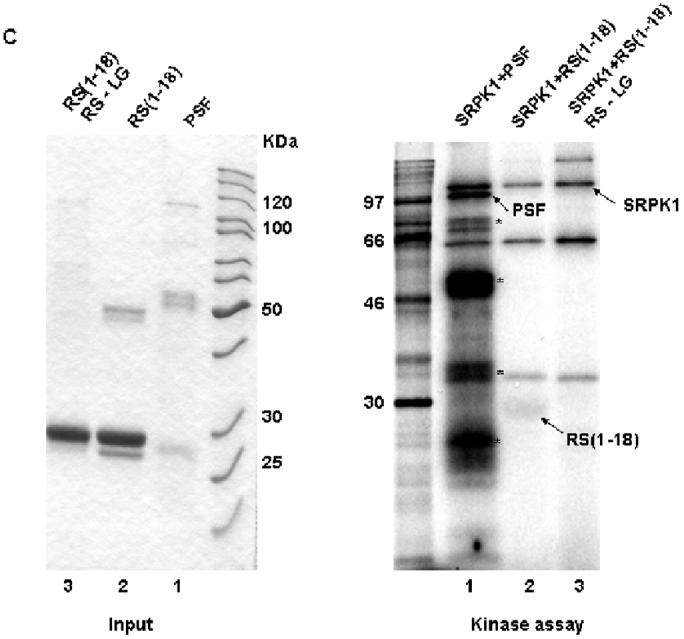
An N-terminal RS dipeptide within PSF is phosphorylated by SRPK1. (A). Amino acid sequences spanning the five RS dipeptides, the two mutations of 7-RS-8 within the N-terminal (1-18) region, and their relative positions within PSF. All peptides were expressed and purified as GST fusion proteins. (B) The indicated bacterially expressed GST fusion proteins were incubated with SRPK1 in the presence of [γ-32P]ATP at 23°C for 30 min (right panel). Integrity and equal inputs for each fusion protein was confirmed by Commassie staining (left panel). (C) Bacterially expressed RS(1-18) wildtype and mutated peptides (left panel) were incubated with SRPK1 in the presence of [γ-32P]ATP at 23°C for 30 min (right panel). The samples were resolved on 10% SDS-PAGE and visualized by phosphoimaging. Asterisks denote degradation products of full-length PSF. The apparent increase in signal of SRPK1-mediated phosphorylation in this experiment relative to that in Fig. 3B results from the use of a higher specific activity 32P-ATP substrate
We carried the experiment out on both the 7-RS-8 mutant and on No reactivity was found with the mAb. This further confirmed our conclusion that 7-RS-8 was the only site of SR phosphorylation in PSF. We have inserted this text as data not shown on p XX of the Results section.
SR Phosphorylation reduces the binding of PSF to the polypyrimidine tract
To test whether SR phosphorylation affects the ability of PSF to bind to its pre-mRNA target, we performed an in vitro gel mobility shift assay. In Fig 5A, bacterially produced non-phosphorylated PSF was incubated at increasing concentrations with a radiolabeled RNA oligonucleotide containing both the branch point and the Py (b/p+Py) from the intron of the Adenovirus major late (Adml) transcript [47]. With purified bacterially expressed protein, the slower migrating, more intense species corresponds to full-length PSF, while the lower species is likely the 68 kD breakdown product of Figs. 2B and 3 which was previously shown [14] to bind to RNA. Both species show specific competition with cold bp+Py but not with a scrambled sequence competitor, NT3.
Figure 5.
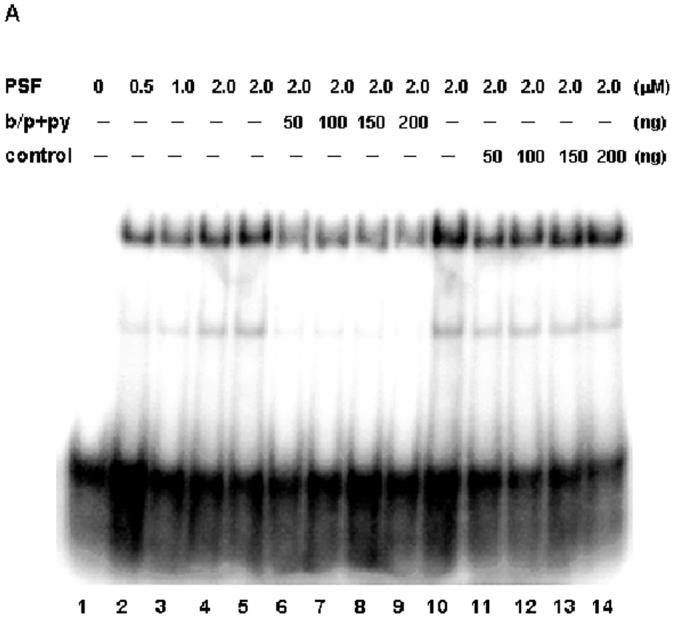
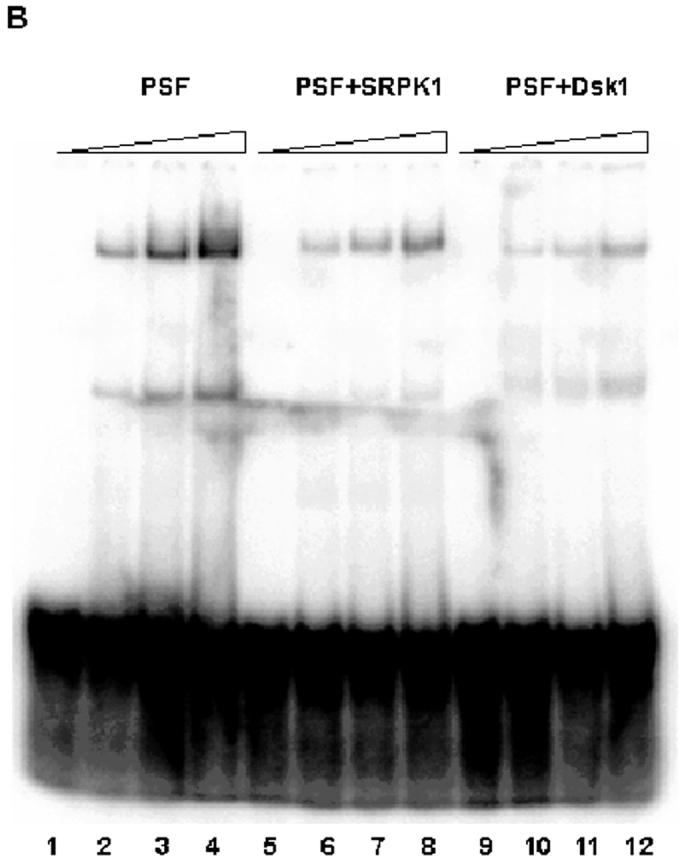
SR phosphorylation regulates the in vitro binding of PSF to the polypyrimidine tract of the 3’ splice site. (A) Specificity of PSF-pyrimidine (py) tract binding. PSF expressed alone in and purified from E coli was incubated with a labeled RNA oligo (∼1ng) containing the branch point (bp) and py (bp+py) at 30°C for 25 min (lane, 1-4). Specific (lanes 5-9) or non-specific (lanes 10-14) unlabeled competitors were added at varying concentrations to the binding reactions prior to resolution on a 6% native polyacrylamide gel and visualization by phosphoimaging. (B) SR phosphorylation reduces binding. PSF (lanes 1-4) or PSF co-expressed with Dsk1 (PSFD) (lanes 5-8) or SRPK1 (PSFS) (lanes 9-12) were incubated separately with the labeled RNA bp+py-containing oligo (∼1ng) at 30°C for 25 min. Samples were resolved and visualized as in(A).
Having established specificity, we then tested equivalent concentrations (as judged by Commassie staining of inputs; data not shown) of non-phosphorylated and phosphorylated PSF (produced in and purified from E. coli) in the same assay (Fig. 5B). We found that PSF-RNA interaction was significantly reduced by SR-phosphorylation of PSF by either Dsk1 or SRPK1.
While there is considerable degradation of full length PSF, we have repeated this experiment several times with independent preparations of phosphorylated and nonphosphorylated PSF and obtained the same results (data not shown). If a degraded version of PSF was giving rise to the minor (faster mobility) gel shifted band as suspected [14], this species shows the same binding and competition behavior as the slower mobility gel-shifted (presumably) full-length PSF-containing RNA-complex. In bacteria, SR phosphorylation (as judged by the data of Fig.2) and other experiments (eg, protein yields following Ni-resin purification; data not shown) appear to stabilize PSF relative to the nonphosphorylated form. Phosphorylated preparations of PSF undoubtedly contain nonphosphorylated substrate. Therefore, we suspect that the inhibitory effects of SR-phosphorylation of PSF observed in Fig.5B are an underestimate. We conclude that SR-phosphorylation of PSF regulates its ability to bind to the polypyrimidine tract of pre-mRNA introns.
DISCUSSION
When over-expressed in mammalian cells, PSF appears to be phosphorylated by an unidentified SR kinase, likely one of the SRPK family, such that an SR phosphorylation-specific monoclonal antibody (mAb3C5) could recognize it. These results were confirmed with an additional anti-SR monoclonal antibody, mAb104, for PSF expressed by baculoviral infection of insect cells (data not shown). We found that recombinant SRPK1 or Dsk1, SR kinases shown to have high specificity for their substrates [24, 41], phosphorylate PSF in vitro. Furthermore, PSF is a substrate for both SR kinases when co-expressed with them in bacteria, as evidenced by their recognition with mAb3C5. On the other hand, neither nonO nor its human homologue, p54nrb, which share high sequence identity/similarity with PSF, is phosphorylated under the same conditions. These results provided strong evidence that PSF is a non-classical substrate of SR kinases and that the phosphorylation is specific for RS dipeptides [24, 41]. That conclusion was confirmed by mutagenesis experiments that demonstrated SR phosphorylation near the N-terminus at position 8.
PSF is an atypical SR substrate in that it contains no obvious RS domain. While unusual, this is not unprecedented. In budding yeast, Npl3p was reported to be phosphorylated by mammalian SRPK1 and by its yeast homolog Sky1 [26]. As with Np13p, in which only 1 of 8 RS dipeptides was phosphorylated [27], only 1 of 5 RS dipeptides was phosphorylated in PSF by SRPK1. According to Wang et al. [28], a serine (position 0) with arginines on both sides (positions -1 and +1), and with an arginine in position -3, is the preferred substrate for SR kinases [28]. In PSF, 7-RS-8 fits the consensus precisely (Fig. 4A), and this motif explains why only RS (1-18), out of 5 candidates was phosphorylated by SRPK1. On the other hand, p54nrb/nonO contains an RS dipeptide, RSR, just N-terminal to its RNA recognition motif [9, 10]. Perhaps it cannot be phosphorylated by SRPK1 because it lacks either an arginine at position -3 or the other criterion of Wang et al. [28], a basic path surrounding the targeted serine. We noted that the level of phosphorylation of the N-terminal, RS-containing fragment was weaker than that observed for full-length PSF. We suspect that this short fusion fragment might not be folded as natively as the full-length protein. It is also possible that other SRPK1 contacts presented in full-length PSF were lost in the deleted version.
PSF is an hnRNP protein that associates with RNA and snRNP complexes at multiple stages. For example, PSF binds to both U5snRNA [12] and the U4/U6.U5 tri-snRNP [13]. Since PSF also accumulates in the spliceosome complex B [6], it likely participates in bridging this complex with tri-snRNP. PSF interacts with the 5’ splice site[48], but in addition, appears to accumulate in complex C where it could participate in the second step of catalysis [14]. Therefore, we felt it would be informative to determine the effects of SR phosphorylation of PSF on RNA binding. To address this, we chose to determine the functional consequence of SR phosphorylation on the binding of PSF to the intronic polypyrimidine track (Py). We observed a significant and specific reduction of Py binding by phosphorylated PSF. This is similar to what was observed for the non-conventional SR protein Np13, in which phosphorylation decreased its association with poly (A)+ RNA [35]. Our binding experiment of Fig. 5B indicates that the extent of phosphorylation correlates with the degree of inhibition. However, we cannot distinguish whether phosphorylation eliminates binding completely or whether it reduces binding affinity, because the preparations of phosphoryated PSF most probably contain unphosphorylated substrate. The same caveat holds for the conclusions drawn on Np13[35]. In addition it was shown that modulation of Sky1-mediated SR phosphorylation of Np13 is required for its proper nuclear localization [27].
SR phosphorylation of conventional substrates, such as ASF/SF2, can alter their interactions with RNA or with other proteins to affect pre-mRNA processing and mRNA export [49-51]. Indeed, a phosphorylation-dephosphorylation cycle is necessary for splicing [52-54]. PSF has been shown in two-hybrid assays to interact with protein phosphatase type 1 (PP1) [55]. Microcystin-LR inhibits PP1 and PP2A by irreversibly binding to their catalytic subunits, thereby blocking the catalytic function of the spliceosome and the assembly of complexes B and C [55]. Both these complexes have been shown to contain PSF [6, 14]. The finding that SRPK1, co-localizes in speckles with the splicing factors SC35 and B1C8 [56], indicates that an SR kinase, PSF and a putative phosphatase capable of reversing the reaction are present in the same place at the same time. SR proteins have been reported in Drosophila for regulating alternative splicing and sex determination, and phosphorylation by Doa, a LAMMER kinase, plays an important role in this regulation [57].
While PSF can serve as a substrate for SRPK1, our data do not address whether PSF is phosphorylated exclusively by SRPK1 in mammalian cells. PSF was reported to be hyperphosphorylated on serine and threonine residues during apoptosis—an event that dissociates PSF from PTB, leading to binding interactions with new partners [58]. It remains to be determined whether differential phosphorylation of PSF plays a role in these events. Further investigation is necessary to determine additional functional consequences of unconventional SR phosphorylation of PSF.
ACKNOWLEDGEMENTS
We thank Dr. B. Turner for providing mab3C5 anti-serum, Dr. X.-D. Fu for the SRPK1 construct, Dr. A. Krainer for the p54nrb construct. We thank P. Dudek for helpful discussions and Ms. Maya Ghosh for technical assistance. The work was supported by NIH grants GM40639 to RJL and CA92318 to PWT.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
REFERENCES
- [1].Kramer A. The structure and function of proteins involved in mammalian pre-mRNA splicing. Annu. Rev. Biochem. 1996;65:367–409. doi: 10.1146/annurev.bi.65.070196.002055. [DOI] [PubMed] [Google Scholar]
- [2].Staley JP, Guthrie C. Mechanical devices of the spliceosome: motors, clocks, springs, and things. Cell. 1998;92:315–326. doi: 10.1016/s0092-8674(00)80925-3. [DOI] [PubMed] [Google Scholar]
- [3].Valcarcel J, Gaur RK, Singh R, Green MR. Interaction of U2AF65 RS region with pre-mRNA branch point and promotion of base pairing with U2 snRNA. Science. 1996;273:1706–1709. doi: 10.1126/science.273.5282.1706. [DOI] [PubMed] [Google Scholar]
- [4].Fu XD. The superfamily of arginine/serine-rich splicing factors. RNA. 1995;1:663–680. [PMC free article] [PubMed] [Google Scholar]
- [5].Patton JG, Mayer SA, Tempst P, Nadal-Ginard B. Characterization and molecular cloning of polypyrimidine tract-binding protein: a component of a complex necessary for pre-mRNA splicing. Genes Dev. 1991;5:1237–1251. doi: 10.1101/gad.5.7.1237. [DOI] [PubMed] [Google Scholar]
- [6].Patton JG, Porro EB, Galceran J, Tempst P, Nadal-Ginard B. Cloning and characterization of PSF, a novel pre-mRNA splicing factor. Genes Dev. 1993;7:393–406. doi: 10.1101/gad.7.3.393. [DOI] [PubMed] [Google Scholar]
- [7].Shav-Tal Y, Zipori D. PSF and p54(nrb)/NonO--multi-functional nuclear proteins. FEBS Lett. 2002;531:109–114. doi: 10.1016/s0014-5793(02)03447-6. [DOI] [PubMed] [Google Scholar]
- [8].Zhang WW, Zhang LX, Busch RK, Farres J, Busch H. Purification and characterization of a DNA-binding heterodimer of 52 and 100 kDa from HeLa cells. Biochem. J. 1993;290:267–272. doi: 10.1042/bj2900267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Dong B, Horowitz DS, Kobayashi R, Krainer AR. Purification and cDNA cloning of HeLa cell p54nrb, a nuclear protein with two RNA recognition motifs and extensive homology to human splicing factor PSF and Drosophila NONA/BJ6. Nucleic Acids Res. 1993;21:4085–4092. doi: 10.1093/nar/21.17.4085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].Yang YS, Hanke JH, Carayannopoulos L, Craft CM, Capra JD, Tucker PW. NonO, a non-POU-domain-containing, octamer-binding protein, is the mammalian homolog of Drosophila nonAdiss. Mol. Cell. Biol. 1993;13:5593–5603. doi: 10.1128/mcb.13.9.5593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Kiesler E, Miralles F, Ostlund Farrants AK, Visa N. The Hrp65 self interaction is mediated by an evolutionarily conserved domain and is required for nuclear import of Hrp65 isoforms that lack a nuclear localization signal. J. Cell Sci. 2003;116:3949–3956. doi: 10.1242/jcs.00690. [DOI] [PubMed] [Google Scholar]
- [12].Peng R, Dye BT, Perez I, Bernard DC, Thompson AB, Patton JG. PSF and p54nrb bind a conserved stem in U5snRNA. RNA. 2002;8:1334–1347. doi: 10.1017/s1355838202022070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].Teigelkamp S, Mundt C, Achsel T, Will CL, Luhrmann R. The human U5 snRNP-specific 100-kD protein is an RS domain containing, putative RNA helicase with significant homology to the yeast splicing factor Prp28. RNA. 1997;3:1313–1326. [PMC free article] [PubMed] [Google Scholar]
- [14].Gozani O, Patton JG, Reed R. A novel set of spliceosome-associated proteins and the essential splicing factor PSF bind stably to pre-mRNA prior to catalytic step II of the splicing reaction. EMBO J. 1994;13:3356–3367. doi: 10.1002/j.1460-2075.1994.tb06638.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Maniatis T, Reed R. An extensive network of coupling among gene expression machines. Nature. 2002;416:499–506. doi: 10.1038/416499a. [DOI] [PubMed] [Google Scholar]
- [16].Emili A, Shales M, McCracken S, Xie W, Tucker PW, Kobayashi R, Blencowe BJ, Ingles CJ. Splicing and transcription-associated proteins PSF and p54nrb/nonO bind to the RNA polymerase II CTD. RNA. 2002;8:1102–1111. doi: 10.1017/s1355838202025037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Rosonina E, Bakowski MA, McCracken S, Blencowe BJ. Transcriptional activators control splicing and 3′-end cleavage levels. J. Biol. Chem. 2003;278:43034–43040. doi: 10.1074/jbc.M307289200. [DOI] [PubMed] [Google Scholar]
- [18].Rosonina E, Ip JY, Calarco JA, Bakowski MA, Emili A, McCracken S, Tucker PW, Ingles CJ, Blencowe BJ. Role for PSF in mediating transcriptional activator-dependent stimulation of pre-mRNA processing in vivo. Mol. Cell. Biol. 2005;15:6734–6746. doi: 10.1128/MCB.25.15.6734-6746.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Mathur M, Tucker PW, Samuels HH. PSF is a novel corepressor that mediates its effect through Sin3A and the DNA binding domain of nuclear hormone receptors. Mol. Cell. Biol. 2001;21:2298–2311. doi: 10.1128/MCB.21.7.2298-2311.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [20].Sewer MB, Nguyen VQ, Huang CJ, Tucker PW, Kagawa N, Waterman MR. Transcriptional activation of human CYP17 in H295R adrenocortical cells depends on complex formation among p54(nrb)/NonO, protein-associated splicing factor, and SF-1, a complex that also participates in repression of transcription. Endocrinology. 2002;143:1280–1290. doi: 10.1210/endo.143.4.8748. [DOI] [PubMed] [Google Scholar]
- [21].Bladen CL, Udayakumar D, Takeda Y, Dynan WS. Identification of the polypyrimidine tract binding protein-associated splicing factor.p54(nrb) complex as a candidate DNA double-strand break rejoining factor. J. Biol. Chem. 2005;280:5205–5210. doi: 10.1074/jbc.M412758200. [DOI] [PubMed] [Google Scholar]
- [22].Zhang Z, Carmichael GG. The fate of dsRNA in the nucleus: a p54(nrb)-containing complex mediates the nuclear retention of promiscuously A-to-I edited RNAs. Cell. 2001;106:465–475. doi: 10.1016/s0092-8674(01)00466-4. [DOI] [PubMed] [Google Scholar]
- [23].Zolotukhin AS, Michalowski D, Bear J, Smulevitch SV, Traish AM, Peng R, Patton J, Shatsky IN, Felber BK. PSF acts through the human immunodeficiency virus type 1 mRNA instability elements to regulate virus expression. Mol. Cell. Biol. 2003;23:6618–6630. doi: 10.1128/MCB.23.18.6618-6630.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [24].Colwill K, Feng LL, Yeakley JM, Gish GD, Caceres JF, Pawson T, Fu XD. SRPK1 and Clk/Sty protein kinases show distinct substrate specificities for serine/arginine-rich splicing factors. J. Biol. Chem. 1996;271:24569–24575. doi: 10.1074/jbc.271.40.24569. [DOI] [PubMed] [Google Scholar]
- [25].Rossi F, Labourier E, Forne T, Divita G, Derancourt J, Riou JF, Antoine E, Cathala G, Brunel C, Tazi J. Specific phosphorylation of SR proteins by mammalian DNA topoisomerase I. Nature. 1996;381:80–82. doi: 10.1038/381080a0. [DOI] [PubMed] [Google Scholar]
- [26].Yun CY, Fu XD. Conserved SR protein kinase functions in nuclear import and its action is counteracted by arginine methylation in Saccharomyces cerevisiae. J. Cell Biol. 2000;150:707–718. doi: 10.1083/jcb.150.4.707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Gilbert W, Siebel CW, Guthrie C. Phosphorylation by Sky1p promotes Npl3p shuttling and mRNA dissociation. RNA. 2001;7:302–313. doi: 10.1017/s1355838201002369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Wang HY, Lin W, Dyck JA, Yeakley JM, Songyang Z, Cantley LC, Fu XD. SRPK2: a differentially expressed SR protein-specific kinase involved in mediating the interaction and localization of pre-mRNA splicing factors in mammalian cells. J. Cell Biol. 1998;140:737–750. doi: 10.1083/jcb.140.4.737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Graveley BR. Sorting out the complexity of SR protein functions. RNA. 2000;6:1197–1211. doi: 10.1017/s1355838200000960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Hertel KJ, Graveley BR. RS domains contact the pre-mRNA throughout spliceosome assembly. Trends Biochem. Sci. 2005;30:115–118. doi: 10.1016/j.tibs.2005.01.002. [DOI] [PubMed] [Google Scholar]
- [31].Shen H, Green MR. A pathway of sequential arginine-serine-rich domain splicing signal interactions during mammalian spliceosome assembly. Mol. Cell. 2004;16:363–373. doi: 10.1016/j.molcel.2004.10.021. [DOI] [PubMed] [Google Scholar]
- [32].Shen H, Kan JL, Green MR. Arginine-serine-rich domains bound at splicing enhancers contact the branchpoint to promote prespliceosome assembly. Mol. Cell. 2004;13:367–376. doi: 10.1016/s1097-2765(04)00025-5. [DOI] [PubMed] [Google Scholar]
- [33].Fetzer S, Lauber J, Will CL, Luhrmann R. The [U4/U6.U5] tri-snRNP-specific 27K protein is a novel SR protein that can be phosphorylated by the snRNP-associated protein kinase. RNA. 1997;3:344–355. [PMC free article] [PubMed] [Google Scholar]
- [34].Roscigno RF, Garcia-Blanco MA. SR proteins escort the U4/U6.U5 tri-snRNP to the spliceosome. RNA. 1995;1:692–706. [PMC free article] [PubMed] [Google Scholar]
- [35].Cazalla D, Newton K, Caceres JF. A novel SR-related protein is required for the second step of Pre-mRNA splicing. Mol. Cell. Biol. 2005;25:2969–2980. doi: 10.1128/MCB.25.8.2969-2980.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [36].Huang Y, Steitz JA. SRprises along a messenger′s journey. Mol. Cell. 2005;17:613–615. doi: 10.1016/j.molcel.2005.02.020. [DOI] [PubMed] [Google Scholar]
- [37].Otto H, Dreger M, Bengtsson L, Hucho F. Identification of tyrosine phosphorylated proteins associated with the nuclear envelope. Euro. J. Biochem. 2001;268:420–428. doi: 10.1046/j.1432-1033.2001.01901.x. [DOI] [PubMed] [Google Scholar]
- [38].Akhmedov AT, Lopez BS. Human 100-kDa homologous DNA-pairing protein is the splicing factor PSF and promotes DNA strand invasion. Nucleic Acids Res. 2000;28:3022–3030. doi: 10.1093/nar/28.16.3022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Deloulme JC, Prichard L, Delattre O, Storm DR. The prooncoprotein EWS binds calmodulin and is phosphorylated by protein kinase C through an IQ domain. J. Biol. Chem. 1997;272:27369–27377. doi: 10.1074/jbc.272.43.27369. [DOI] [PubMed] [Google Scholar]
- [40].Shav-Tal Y, Cohen M, Lapter S, Dye B, Patton JG, Vandekerckhove J, Zipori D. Nuclear relocalization of the pre-mRNA splicing factor PSF during apoptosis involves hyperphosphorylation, masking of antigenic epitopes, and changes in protein interactions. Mol. Biol. Cell. 2001;12:2328–2340. doi: 10.1091/mbc.12.8.2328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Tang Z, Yanagida M, Lin RJ. Fission Yeast Mitotic Regulator Dsk1 is an SR Protein-Specific Kinase. J. Biol. Chem. 1998;273:5963–5969. doi: 10.1074/jbc.273.10.5963. [DOI] [PubMed] [Google Scholar]
- [42].Li C, Schwabe JR, Banayo E, Evans RM. Coexpression of nuclear receptor partners increases their solubility and biological activities. Proc. Natl. Acad. Sci. 1997;94:2278–2283. doi: 10.1073/pnas.94.6.2278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [43].Yue BG, Ajuh P, Akusjärvi G, Lamond AI, Kreivi JP. Functional coexpression of serine protein kinase SRPK1 and its substrate ASF/SF2 in Escherichia Coli. Nucleic Acids Res. 2000;28:e14. doi: 10.1093/nar/28.5.e14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].Bridge E, Xia DX, Carmo-Fonseca M, Cardinali B, Lamond AI, Pettersson U. Dynamic organization of splicing factors in adenovirus-infected cells. J. Virol. 1995;69:281–290. doi: 10.1128/jvi.69.1.281-290.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [45].Turner BM, Davies S, Whitfield WG. Characterization of a family of nuclear and chromosomal proteins identified by a monoclonal antibody. Euro.Cell Biology. 1985;38:344–352. [PubMed] [Google Scholar]
- [46].Zahler AM, Lane WS, Stolk JA, Roth MB. SR proteins: a conserved family of pre-mRNA splicing factors. Genes Dev. 1992;6:837–847. doi: 10.1101/gad.6.5.837. [DOI] [PubMed] [Google Scholar]
- [47].Berglund JA, Abovich N, Rosbash M. A cooperative interaction between U2AF65 and mBBP/SF1 facilitates branchpoint region recognition. Genes Dev. 1998;12:858–867. doi: 10.1101/gad.12.6.858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48].Kameoka S, Duque P, Konarska MM. p54(nrb) associates with the 5′ splice site within large transcription/splicing complexes. EMBO J. 2004;23:1782–1791. doi: 10.1038/sj.emboj.7600187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [49].Tacke R, Chen Y, Manley JL. Sequence-specific RNA binding by an SR protein requires RS domain phosphorylation: creation of an SRp40-specific splicing enhancer. Proc. Natl. Acad. Sci. 1997;94:1148–1153. doi: 10.1073/pnas.94.4.1148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].Xiao SH, Manley JL. Phosphorylation of the ASF/SF2 RS domain affects both protein-protein and protein-RNA interactions and is necessary for splicing. Genes Dev. 1997;11:334–344. doi: 10.1101/gad.11.3.334. [DOI] [PubMed] [Google Scholar]
- [51].Huang Y, Yario TA, Steitz JA. A molecular link between SR protein dephosphorylation and mRNA export. Proc. Natl. Acad. Sci. 2004;101:9666–9670. doi: 10.1073/pnas.0403533101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [52].Cao W, Jamison SF, Garcia-Blanco MA. Both phosphorylation and dephosphorylation of ASF/SF2 are required for pre-mRNA splicing in vitro. RNA. 1997;3:1456–1467. [PMC free article] [PubMed] [Google Scholar]
- [53].Prasad J, Colwill K, Pawson T, Manley JL. The protein kinase Clk/Sty directly modulates SR protein activity: Both hyper-and hypophosphorylation inhibit splicing. Mol. Cell. Biol. 1999;19:6991–7000. doi: 10.1128/mcb.19.10.6991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [54].Xiao S, Manley JL. Phosphorylation-dephosphorylation differentially affects activities of splicing factor ASF/SF2. EMBO J. 1998;17:6359–6367. doi: 10.1093/emboj/17.21.6359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [55].Hirano K, Edodi F, Patton JG, Hartshorne DJ. Interaction of protein phosphatase type 1 with a splicing factor. FEBS Letters. 1996;389:191–194. doi: 10.1016/0014-5793(96)00577-7. [DOI] [PubMed] [Google Scholar]
- [56].Wang HY, Lin W, Dyck JA, Yeakley JM, Songyang Z, Cantley LC, Fu XD. SRPK2: a differentially expressed SR protein-specific kinase involved in mediating the interaction and localization of pre-mRNA splicing factors in mammalian cells. J. Cell Biol. 1998;140:737–750. doi: 10.1083/jcb.140.4.737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [57].Du C, McGuffin ME, Dauwalder B, Rabinow L, Mattox W. Protein phosphorylation plays an essential role in the regulation of alternative splicing and sex determination in Drosophila. Mol. Cell. 1998;2:741–750. doi: 10.1016/s1097-2765(00)80289-0. [DOI] [PubMed] [Google Scholar]
- [58].Shav-Tal Y, Cohen M, Lapter S, Dye B, Patton JG, Vandekerckhove J, Zipori D. Nuclear relocalization of the pre-mRNA splicing factor PSF during apoptosis involves hyperphosphorylation, masking of antigenic epitopes, and changes in protein interactions. Mol. Biol. Cell. 2001;12:2328–2340. doi: 10.1091/mbc.12.8.2328. [DOI] [PMC free article] [PubMed] [Google Scholar]