

Abstract
Angiosarcomas apparently derive from blood vessel endothelial cells; however, occasionally their histological features suggest mixed origin from blood and lymphatic endothelia. In the absence of specific positive markers for lymphatic endothelia the precise distinction between these components has not been possible. Here we provide evidence by light and electron microscopic immunohistochemistry that podoplanin, a ∼38-kd membrane glycoprotein of podocytes, is specifically expressed in the endothelium of lymphatic capillaries, but not in the blood vasculature. In normal skin and kidney, podoplanin colocalized with vascular endothelial growth factor receptor-3, the only other lymphatic marker presently available. Complementary immunostaining of blood vessels was obtained with established endothelial markers (CD31, CD34, factor VIII-related antigen, and Ulex europaeus I lectin) as well as podocalyxin, another podocytic protein that is also localized in endothelia of blood vessels. Podoplanin specifically immunolabeled endothelia of benign tumorous lesions of undisputed lymphatic origin (lymphangiomas, hygromas) and was detected there as a 38-kd protein by immunoblotting. As paradigms of malignant vascular tumors, poorly differentiated (G3) common angiosarcomas (n = 8), epitheloid angiosarcomas (n = 3), and intestinal Kaposi’s sarcomas (n = 5) were examined for their podoplanin content in relation to conventional endothelial markers. The relative number of tumor cells expressing podoplanin was estimated and, although the number of cases in this preliminary study was limited to 16, an apparent spectrum of podoplanin expression emerged that can be divided into a low-expression group in which 0–10% of tumor cells contained podoplanin, a moderate-expression group with 30–60% and a high-expression group with 70–100%. Ten of eleven angiosarcomas and all Kaposi’s sarcomas showed mixed expression of both lymphatic and blood vascular endothelial phenotypes. By double labeling, most podoplanin-positive tumor cells coexpressed endothelial markers of blood vessels, whereas few tumor cells were positive for individual markers only. From these results we conclude that (1) podoplanin is a selective marker of lymphatic endothelium; (2) G3 angiosarcomas display a quantitative spectrum of podoplanin-expressing tumor cells; (3) in most angiosarcomas, a varying subset of tumor cells coexpresses podoplanin and endothelial markers of blood vessels; and (4) all endothelial cells of Kaposi’s sarcomas expressed the lymphatic marker podoplanin.
It is not unequivocally accepted that all angiosarcomas originate exclusively from endothelia of the blood vasculature; it has been suggested that some also contain components of lymphatic lineage. 1-3 Classification of angiosarcomas is based mainly on morphological criteria, such as rudimentary vasoformation with abortive vessels filled with erythrocytes. Immunohistochemical studies of expression of a large panel of established endothelial markers have not clarified this situation, mainly because these antibodies also showed overlapping immunostaining of lymphatic endothelia. A notable apparent exception was a monoclonal antibody designated PAL-E 4 directed toward an undefined human endothelial antigen, expressed in blood vessels only, that immunostains only unfixed cryostat sections. Thus, any potential subclassification of angiosarcomas has been limited so far primarily by lack of positive endothelial markers that reliably distinguish between the phenotypes of blood and lymphatic vessels.
Recently, we have observed in rat and human kidneys 5 that podoplanin, a ∼38-kd glomerular podocyte membrane mucoprotein, was localized in endothelia of small vessels with topographic and morphological features of lymphatic capillaries. Furthermore, a podoplanin-related rat glycoprotein was found in the endothelium of presumptive lymphatic vessels. 6 Another protein shown to be expressed uniquely on lymphatic endothelia was the receptor for the vascular endothelial growth factor C, designated vascular endothelial growth factor receptor-3 (VEGFR-3) or flt-4. 7-9 Thus we have reasoned that, if antibodies specific for human podoplanin and flt-4 immunolabel the same type of vessel, they could serve as selective immunohistochemical markers in pathologically altered tissues. Here we have confirmed and extended this hypothesis in an immunohistochemical pilot study of normal endothelium, benign vascular tumors, and a small number of angiosarcomas.
Materials and Methods
Human Tissues
Paraffin blocks of 40 cases of vascular tumors and five cases of gastrointestinal Kaposi’s sarcomas from HIV-infected patients (Tables 1 and 2) ▶ ▶ were obtained from the archive of the Institute of Clinical Pathology, University of Vienna. Native renal cortex was dissected from kidneys removed because of hypernephroid carcinoma (n = 10); native lung tissue was dissected from lobectomy specimens removed because of non-small cell carcinoma (n = 5). Cutaneous lymphangiomas (n = 3), normal lymph nodes (n = 3), and normal skin (n = 5) were obtained from the Department of Surgery, University of Vienna. Two fetal hygroma colli were from spontaneous abortions in the 22d and 23rd gestational week.
Table 1.
Podoplanin and Vascular Endothelial Markers in Benign Vascular Tumors
| Histological tumor type | No. of cases | Podoplanin | Vascular markers |
|---|---|---|---|
| Capillary hemangioma | 7 | − | + |
| Cavernous hemangioma | 6 | − | + |
| Mixed lymphatic and hemangioma | 2 | +/−* | −/+* |
| Venous hemangioma | 3 | − | + |
| Lymphangioma | 11 | + | +/− |
*mixed patterns
Table 2.
Podoplanin and Vascular Endothelial Markers in Angiosarcomas
| Patient | Tumor type and localization | % of reactive tumor cells | ||
|---|---|---|---|---|
| Podoplanin | CD31 | Podocalyxin | ||
| S.R. | AS, vena cava sup. | 0 | 80 | 40 |
| K.K. | AS, spine | 5 | 70 | 30 |
| S.A. | epitheloid AS, spine | 5 | 50 | 50 |
| W.I. | AS, arteria pulmonalis | 10 | 20 | 20 |
| H.H. | epitheloid AS, skin | 10 | 70 | 80 |
| A.K. | epitheloid AS skin | 30 | 80 | 80 |
| V.I. | AS, heart | 30 | 70 | 80 |
| H.L. | AS, skin | 30 | 30 | 50 |
| C.L. | AS, heart | 40 | 80 | 70 |
| G.K. | AS, skin | 60 | 80 | 90 |
| D.L. | AS, pleura | 70 | 80 | 40 |
| M.T. | KS, stomach | 100 | 100 | 100 |
| T.D. | KS, stomach | 100 | 100 | 100 |
| J.G. | KS, small intestine | 100 | 100 | 100 |
| G.S. | KS, large intestine | 100 | 100 | 80 |
| K.T. | KS, oral mucosa | 100 | 100 | 70 |
AS, angiosarcoma; KS, Kaposi’s sarcoma.
Materials
Fluorescein isothiocyanate (FITC)-conjugated rabbit anti-mouse IgG F(ab′)2, FITC- and tetramethylrhodamine-conjugated goat anti-rabbit IgG F(ab′)2 were obtained from Jaxell-Accurate (Westbury, NY). Texas red-conjugated goat anti-mouse IgG, biotinylated horse anti-mouse IgG, goat anti-rabbit IgG, and streptavidin-biotin complex peroxidase system were purchased from Vector Laboratories (Burlingame, CA). Goat anti-rabbit and goat anti-mouse IgG conjugated to 5- or 10-nm gold particles were from Amersham (Auroprobe, Buckinghamshire, UK). Alkaline phosphatase-conjugated rabbit anti-mouse IgG and alkaline phosphatase-conjugated goat anti-rabbit Fc fragment specific IgG were from Promega (Madison, WI). Ulex europaeus I lectin (UEA I), rabbit polyclonal antibodies directed to human factor VIII-related antigen and to UEA I lectin and mouse monoclonal antibody directed to CD31 (clone JC 70A) were from Dako (Glostrup, Denmark). Mouse monoclonal antibody to CD34 (clone QBEnd/10) was from Novocastra Laboratories (Newcastle on Tyne, UK). Mouse monoclonal anti-human endothelial antibody (clone PAL-E) was obtained from Harlan Sera Lab (Crawley Down, UK). Mouse monoclonal antibodies to human VEGFR-3 were prepared as described. 9 WGA-sepharose 4B was purchased from Pharmacia LKB (Uppsala, Sweden). Sodium dodecyl sulfate (SDS), acrylamide, TEMED, ammonium persulfate, high- and low molecular weight standards, Tween 20 and PDG-6 columns were from BioRad Laboratories (Richmond, CA). Lowicryl K4M was obtained from Chemische Werke Lowi (Waldkraiburg, Germany). Diaminobenzidine (DAB) was from Serva (Heidelberg, Germany). Leupeptin, antipain, pepstatin, Triton X-100, dithiothreitol (DTT), ammonium sulfate, ethylendiamine tetraacetic acid (EDTA), heparin, protease type XIV from Streptomyces griseus, phosphotungstic acid, and 4′,6-diamidino-2-phenylindole were from Sigma (Deisenhofen, Germany). Pefablock SCR was from Pentapharm (Basel, Switzerland). Nitro blue tetrazolium and 5′-bromo-4-chloro-3-indolylphosphate (NBT/BCIP) was obtained from Kirkegaard and Perry (Gaithersburg, MD). Nitrocellulose (0.45 μm pore size) was from Schleicher and Schuell (Dassel, Germany). Human umbilical vein endothelial cells (HUVEC) and endothelial cell growth supplement were purchased from Technoclone, Vienna. PenStrep, M199 medium and fetal calf serum were obtained from GIBCO BRL (Gaithersburg, MD).
Antibodies Specific for Human Podoplanin and Podocalyxin
Rabbit anti-human podoplanin IgG was raised against the recombinant human homologue of the rat 43-kd glycoprotein podoplanin (Kowalski H, Soleiman A, Endres D, Breiteneder-Geleff S, Kerjashki D, manuscript in preparation). Rabbit sera were affinity-purified using nitrocellulose strips containing recombinant protein, as described. 10 Mouse monoclonal anti-podocalyxin antibody (clone GB6) was produced using as antigen a wheat germ agglutinin-binding fraction of a Triton X-100 lysate of isolated human glomeruli, as described for rat podocalyxin. 11
Immunofluorescence
Unfixed 4-μm-thick cryostat sections of normal human kidney, placenta, and skin were incubated with rabbit anti-human podoplanin IgG (0.5 μg/ml), monoclonal mouse anti-podocalyxin IgG (10 μg/ml), or monoclonal mouse anti-VEGFR-3 IgG (1μg/ml) followed by FITC-conjugated goat anti-rabbit or rabbit anti-mouse F(ab′)2 fragments. For double immunofluorescence, cryostat or deparaffinized, rehydrated paraffin sections were incubated with 10% egg albumin for 15 minutes, followed by a mix of antibodies specific for human podoplanin, human VEGFR-3, and individual endothelial markers (CD31, podocalyxin (clone GB6), and PAL-E) for 14 hours at 4°C. Sections were then incubated with FITC-conjugated goat anti-rabbit IgG and Texas red-conjugated goat anti-mouse IgG, and examined in an Axiophot microscope equipped for epifluorescence (Zeiss, Oberkochen, Germany). Negative controls included omission of first antibodies or substitution of primary antibodies by irrelevant rabbit or mouse IgG.
Immunoperoxidase on Paraffin Sections
Three-μm-thick deparaffinized serial sections of skin, lymph node, and vascular tumors were directly incubated with anti-CD34 IgG. For immunostaining with other endothelial probes pretreatment of sections was required; protease digestion (Sigma protease XIV, 0.1%) was used for CD31, podocalyxin, factor VIII-related antigen, and UEA I. Microwave pretreatment (600 W for 10 minutes) was performed for anti-human podoplanin IgG. Sections were incubated with primary antibodies for 1 hour at 20°C, followed by biotinylated horse anti-mouse IgG or goat anti-rabbit IgG for 30 minutes and detection by a streptavidin-peroxidase complex, according to the manufacturer’s instructions. Peroxidase reaction product was visualized by DAB and H2O2 .
The approximate relative number of podoplanin-positive tumor cells was estimated in paraffin sections of immunoperoxidase-labeled sections. The number of reactive cells was counted in 10 medium-power fields per tumor (magnification, ×40) of nonnecrotic areas, and the identity of cells was subsequently confirmed by high magnification (×460). The number of tumor cells expressing CD31 and podocalyxin were similarly determined. Relative numbers of immunoreactive cells were expressed as percentages of the total number of tumor cells.
Immunoelectron Microscopy
Normal skin was fixed in 4% paraformaldehyde in 20 mmol/L phosphate-buffered saline (PBS) for 6 hours at 20°C and embedded in Lowicryl K4M. 12 Ultrathin sections were incubated in PBS/1% egg albumin, followed by rabbit anti-podoplanin IgG (5 μg/ml) and goat anti-rabbit IgG conjugated to 10-nm gold particles. Other sections were double-labeled with rabbit anti-podoplanin IgG and monoclonal mouse anti-podocalyxin IgG, followed by goat anti-rabbit IgG conjugated to 5-nm gold particles and goat anti-mouse IgG conjugated to 10-nm gold particles.
Cell Culture
HUVECs were cultured in M199 medium, supplemented with 20% fetal calf serum, 1% PenStrep, 0.1% heparin, and 50 μg/ml endothelial cell growth supplement, and used up to passage 4.
Immunoblotting
Membrane protein fractions were prepared from tissue samples of hygroma colli (n = 2), lymphangioma (n = 3), human placenta (n = 5), human lung (n = 5), and isolated human glomeruli, and subconfluent HUVECs as described. 13 Briefly, tissues were frozen in liquid N2, powdered, and suspended in PBS containing protease inhibitors (2 mg/L pepstatin, antipain, leupeptin, and 10 mg/L Pefablock). Cultured cells were rinsed in PBS and scraped from the dishes. Tissues and cells were incubated in Na2CO3, pH 11.5, and the resulting membrane vesicle fraction was processed for immunoblotting as described, 5 using polyclonal rabbit anti-human podoplanin antibody (0.5μg/ml), or monoclonal anti-podocalyxin antibody GB6 (1μg/ml) as primary antibodies.
Results
Podoplanin Is Selectively Expressed in Lymphatic Endothelia of Human Tissues
The specificity of affinity purified rabbit anti-human podoplanin IgG used for immunohistochemistry was confirmed by immunoblotting of membrane protein fractions of human renal glomeruli, lung and placentary membrane protein fractions. In these samples, a 38-kd protein characteristic for human podoplanin was specifically labeled (Figure 1 ▶ , lanes D-F).
Figure 1.

Podoplanin is identified as a 38-kd protein in human tissues and cell lines by immunoblotting with an affinity-purified specific antibody raised in rabbit against a recombinant fusion protein containing the ectodomain. Tissues and cells were extracted with Na2CO3 (pH 11.5) to yield membrane protein fractions that were then used for immunoblotting. A-C: Membrane protein fractions stained with Coomassie Blue of lymphangioma (A), placenta (B), and lung (C). D-F: Immunoblots on these fractions revealed the 38-kd band specific for human podoplanin in lung (D), placenta (E), isolated glomeruli (F), and fetal lymphangioma (hygroma colli) (G). H: Podoplanin was not detected in membrane protein fractions of HUVECs.
Podoplanin was first localized in normal skin and kidney cortex in unfixed cryostat sections. Anti-podoplanin IgG specifically labeled capillaries distinct from those recognized by PAL-E (Figure 2, A and B) ▶ . Double labeling with anti-podoplanin IgG and antibodies to VEGFR-3, at present the only established marker for lymphatic endothelium, resulted in perfect overlap of the fluorescence signals (Figure 2, C ▶ -E).
Figure 2.

Localization of podoplanin in lymphatic capillaries in cryostat sections of skin and kidney by indirect double immunofluorescence with affinity purified rabbit anti-podoplanin IgG. A and B: Normal human skin and kidney cortex labeled with rabbit anti-podoplanin IgG (detected by FITC-anti-rabbit IgG) and blood vessel endothelium-specific mouse monoclonal antibody PAL-E (detected by Texas Red-anti-mouse IgG). Lymphatic endothelia are labeled in green, indicating expression of podoplanin. Endothelia of blood capillaries appear red, indicating the endothelium-specific antigen recognized by PAL-E. C-E: Section of human skin double-labeled with mouse monoclonal antibody directed toward the vascular endothelial growth factor receptor-3 (VEGFR-3) labeled here in green (C), and with anti-podoplanin IgG, indicated by a red fluorescence signal (D). When the two signals are merged, almost perfect overlap is observed, as indicate by a yellow signal (E). Epi, epidermis; L, lymphatic capillary; C, blood capillary; T, tubule. Magnifications, ×400 (A and B); ×450 (C-E).
Anti-podoplanin IgG specifically and selectively labeled dermal lymphatics also in paraffin sections. Expression of podoplanin correlated with the caliber of lymphatic vessels and was prominent in capillaries composed of a single layer of endothelial cells, but not in larger lymphatic vessels with smooth muscle cells. In lymph nodes anti-podoplanin IgG labeled small hilar lymphatic vessels and sinusoidal lining cells, but not large lymphatic vessels and the high endothelial venules (HEV) (Figure 3A) ▶ . By contrast, anti-podocalyxin IgG and CD31 immunolabeled HEVs (Figure 3B) ▶ , while sinus endothelial cells were only weakly stained.
Figure 3.

Localization of podoplanin, podocalyxin, and CD31 in consecutive paraffin sections of a normal human lymph node by immunoperoxidase. A: Podoplanin is located in marginal sinusoidal lining cells (SIN), but not in endothelial cells of HEV. B: The endothelial marker podocalyxin immunostains sinus lining cells and HEV endothelia. C: Endothelia of HEV are prominently stained with CD31. Magnification, ×450.
By electron microscopy, dermal lymphatic capillaries were identified by conventional morphological criteria, such as absence of a continuous basal lamina and pericytes, and lack of erythrocytes in their lumen. Podoplanin was localized by immunogold procedures predominantly at the luminal surface of lymphatic endothelia, while a small fraction (∼10%) was also found on the abluminal surface and the intercellular regions (Figure 4A) ▶ . Endothelia of blood vessels were consistently devoid of gold particles (Figure 4B) ▶ .
Figure 4.

Localization of podoplanin in dermal lymphatic and blood capillaries, using indirect immunogold labeling with affinity purified rabbit anti-podoplanin IgG on ultrathin Lowicryl sections. A: This vessel was identified as lymphatic capillary with overlapping, interdigitating endothelial junctions (J) with desmoses (D), and lack of erythrocytes in the lumen. Podoplanin is distributed on the endothelial surface, with strong preference of the luminal surface. Few gold particles are found on the basolateral surface (arrowheads). B: Dermal blood capillary with an erythrocyte in its lumen, immunolabeled with anti-podoplanin IgG as in A. Gold particles were not found on endothelial cells nor on pericytes and erythrocytes. C: Double-labeling of a dermal lymphatic capillary with anti-podocalyxin IgG (10-nm gold particles, arrowheads), and anti-podoplanin IgG (5-nm gold particles, arrows). Both markers were found on the endothelial surface. MV, microvillus; J, junction; D, desmosome-like structure; Ery, erythrocyte. Magnification, ×40,000.
Collectively, these findings indicate that podoplanin and VEGFR-3 colocalize and are specifically and selectively expressed on endothelial cells of small lymphatic capillaries but not in blood vessels of any size, and that rabbit anti-podoplanin IgG is a valid immunohistochemical marker of lymphatic capillaries.
Podoplanin Is Expressed in Endothelial Cells of Benign Lymphatic Tumors
Podoplanin was localized in endothelia of 11 lymphangiomas of skin, oral mucosa, and mesenterium, whereas adjacent blood vessels were not labeled (Figure 5D) ▶ . Anti-podoplanin IgG specifically labeled a ∼38-kd protein by immunoblotting on membrane protein fractions of two lymphangiomas, indicating the presence of authentic podoplanin (Figure 1G) ▶ . Controls included omission of the primary antibody, or replacement by other endothelium-specific proteins such as podocalyxin. Anti-podocalyxin IgG was calibrated by immunoblotting of human glomerular lysates, a major source of this protein, and in HUVECs (Figure 6 ▶ , lane D), and specifically labeled a protein doublet with an apparent molecular mass of 170 kd (Figure 6 ▶ , lane B).
Figure 5.
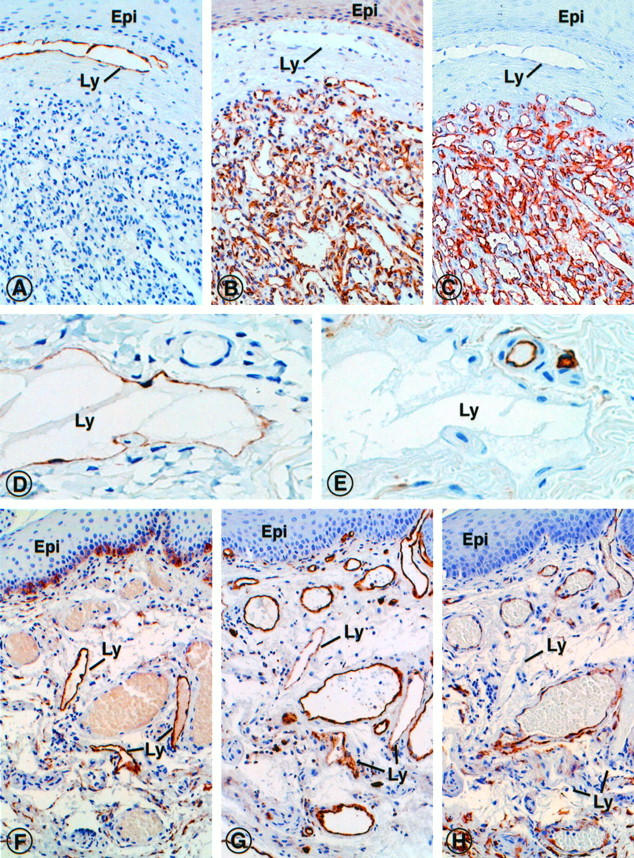
Localization of the lymphatic phenotypic marker podoplanin and indicators of endothelia of blood vessels (podocalyxin, CD31 and CD34) in consecutive paraffin sections of benign vascular tumors. A-C: Dermal capillary hemangioma immunostained for podoplanin (A), podocalyxin (B) and CD34 (C). Although podoplanin was not found within the tumor, the markers of blood vessels were strongly expressed. By contrast, a dermal lymphatic vessel (Ly) beneath the epidermis (Epi) was strongly labeled with antibodies to podoplanin, but not with those to podocalyxin or CD34. D and E: Dermal lymphangioma immunostained for podoplanin (D), and CD34 (E). Podoplanin is expressed by the endothelial lining cells of the cavernous lymphangioma space (Ly), normal blood vessels in the adjacent connective tissue were not marked (D). An inverse labeling pattern was obtained with the endothelial marker CD34 (E). F-H: Mixed angioma of rectal mucosa, immunostained with antibodies specific for podoplanin (F), CD31 (G), and CD34 (H). Although podoplanin was detected exclusively in lymphatic vessels (Ly), CD31 and CD34 were localized primarily in blood vessels, and to a variable extent also in the lymphatics. Basal epidermal cells were spuriously labeled with all antibodies used, which could represent unspecific background staining. Epi, epithelium; Ly, lymphatic capillary. Magnifications, ×450 (A-C), ×750 (D-E), ×450 (F-H).
Figure 6.

Podocalyxin, a sialoglycoprotein of glomerular podocytes, was used as immunohistochemical indicator of human endothelia of blood vessels, using a specific monoclonal antibody designated GB6. A and C: Membrane protein fractions of isolated human glomeruli (A) and of cultured HUVECs (C) stained by Coomassie Blue. Monoclonal mouse antibody GB6 specifically immunoblots podocalyxin as a 170-kd glycoprotein in glomeruli (B) and HUVECs (D).
In capillary, cavernous, and venous hemangiomas (n = 16), podoplanin was not found, whereas lymphatic vessels outside the tumors were intensely labeled (Figure 5A) ▶ . Immunostaining of consecutive sections with vascular endothelial markers (CD31, CD34, factor VIII-related antigen, UEA I, podocalyxin) resulted not only in complementary labeling of tumor blood vessels, but also in variable labeling of lymphatic endothelium (Figure 5, B and C) ▶ .
Collectively, these results indicated that podoplanin was specifically expressed by endothelia of benign lymphatic tumorous lesions, similar to nonneoplastic lymphatic endothelium.
Angiosarcomas Coexpress Podoplanin and Markers of Blood Vessel Endothelium
Eight common angiosarcomas (cutaneous, soft tissue, and visceral type) and three epitheloid cutaneous angiosarcomas (Table 2) ▶ were immunostained with anti-podoplanin IgG. All were high-grade tumors (G3) and contained nonvasoformative or poorly vasoformative areas (Figure 7A) ▶ . Podoplanin was variably expressed in 10 of 11 tumors in 5–70% of tumor cells (Table 2) ▶ . Podoplanin-expressing tumor cells were arranged in clusters of 5 to 30 cells that were confluent in tumors with high levels of expression, and frequently formed abortive lumina with occasional erythrocytes. Isolated, single podoplanin-containing tumor cells were also occasionally encountered, some of which showed atypical mitotic figures (Figure 7B ▶ , inset). Invariably, podoplanin was detected in highest concentrations at tumor cells’ membranes (Figure 7, B and C) ▶ .
Figure 7.
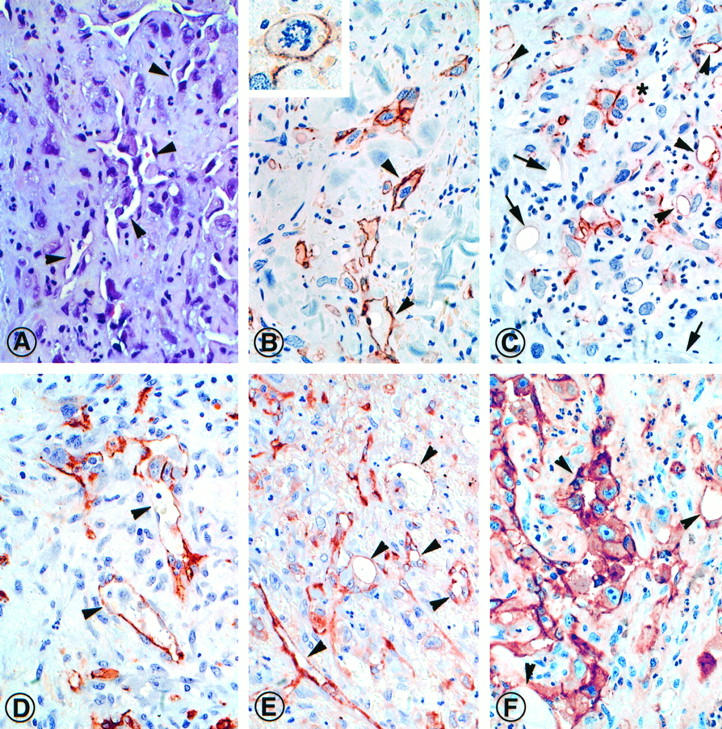
Expression of podoplanin and endothelial markers (CD31, CD34 and factor VIII related antigen) exemplified in a dermal high grade (G3) epitheloid angiosarcoma (Patient A. K., Table 2 ▶ ). Consecutive paraffin sections were stained with hematoxylin and eosin (A) and immunolabeled for localization of podoplanin (B and C), CD34 (D), factor VIII-related antigen (E), and CD31 (F). In approximately 30% of tumor cells podoplanin was detected. Most podoplanin-expressing cells formed small groups and clusters that were frequently associated with vascular clefts (arrowheads). Individual tumor cells were also found outside these groups (B, inset). Occasionally vasoformative structures contain both podoplanin-positive and -negative lining cells (C, asterisk). Small vascular clefts without podoplanin-containing cells are prominent in some areas of the tumor (C, arrows). Although the distribution of CD34 was similar to podoplanin (D), factor VIII-related antigen (E) and CD31 (F) are more widely distributed in this tumor. Magnification, ×450.
A quantitative estimate of the relative numbers of podoplanin-expressing tumor cells in individual angiosarcomas was obtained by direct microscopic examination of immunostained slides and defined as a percentage of the entire tumor cell population. Although the number of cases analyzed was obviously small and suited only for a pilot study, a spectrum became apparent. We divided the cases into a low expression group (n = 5) with only 0–10% of podoplanin-positive tumor cells, a moderate expression group (n = 5) with 30–60% positive cells, and a high expression group (n = 6) with 70–100% positive cells. Two tumors of the first group originated in large blood vessels and two others were derived from the soft tissue of the spine. Tumors with high expression of podoplanin originated predominantly from skin and heart (Table 2) ▶ . There was no correlation of podoplanin expression and histological type of angiosarcoma. Localization of vascular endothelial markers in consecutive paraffin sections showed some topographic, but no quantitative, association with podoplanin (Figure 7, D ▶ -F).
To determine precisely whether podoplanin-producing tumor cells expressed endothelial antigens of blood vessels simultaneously, double immunofluorescence with IgGs specific for podoplanin and podocalyxin was performed. Both markers labeled at least 30% of tumor cells and were concentrated in clusters (Figure 8) ▶ , in agreement with the results obtained by immunoperoxidase labeling. Most tumor cells (>70%) coexpressed podoplanin and podocalyxin (Figure 8C) ▶ , whereas only a small fraction expressed podoplanin only, and none podocalyxin alone. Podoplanin was localized also with this technique in highest concentration on cell surfaces, while podocalyxin and CD31 were predominately found in the cytoplasm of tumor cells (Figure 8, A and B) ▶ . Tumor cells located in the black background in these preparations failed to stain with either marker.
Figure 8.

Colocalization of podoplanin and podocalyxin in a cardiac common angiosarcoma (Patient C. L. (Table 2) ▶ , classified as containing 40% podoplanin-positive tumor cells) using indirect double immunofluorescence on a single paraffin section. A cluster of tumor cells is selected to show podoplanin on the cell surfaces (A). The same tumor cells also express podocalyxin on both surface membranes and cytoplasm (B). When the two signals are merged, podocalyxin and podoplanin colocalize on the cell surface of all tumor cells, as indicated by the yellow fluorescence signal (C). Magnification, ×1000.
All Kaposi’s sarcomas investigated in this study (5 originating in the alimentary canal from HIV-infected individuals) were in the early patch-forming stage, with conspicuous jagged vascular spaces dissecting either the gastrointestinal (Figure 9A) ▶ or the oral mucosa. In all cases the atypical endothelial cells lining these spaces expressed podoplanin on their cell surface (100% of podoplanin-positive tumor cells, Figure 9B ▶ ). In consecutive sections the Kaposi’s sarcoma spindle cells revealed moderate to strong reactivity for the endothelial markers CD31 and CD34 (Figure 9, C and D) ▶ .
Figure 9.

Expression of podoplanin and endothelial markers of blood vessels (CD31, CD34) in Kaposi’s sarcoma of the gastric mucosa (Patient M. T., Table 2 ▶ ). Consecutive paraffin sections were stained with hematoxylin and eosin (A), and immunolabeled for the localization of podoplanin (B), CD31 (C), and CD34 (D). Podoplanin was detected in all tumor cells lining the irregular vascular clefts. A similar reactivity was observed for CD34, a weaker labeling with antibodies to CD31. Arrowhead: gastric gland. Magnification, ×450.
Collectively, these data indicate that tumor cell surfaces of 10 of 11 angiosarcomas expressed variable amounts of the lymphatic marker podoplanin, all tumor cells of early stage Kaposi’s sarcomas reacted with anti-podoplanin IgG, and the majority of tumor cells also coexpressed markers found primarily in blood vessel endothelium (CD31 and podocalyxin).
Discussion
Characterization of cellular phenotypes by immunohistochemical markers has provided a solid basis for current histopathological classifications of neoplastic lesions. Particularly in soft tissue tumors of low differentiation, the conserved expression of several endothelial proteins is frequently the major diagnostic clue leading to identification of these tumors as angiosarcomas. However, as there are “no reliable means to distinguishing blood vascular from lymphatic endothelial differentiation (or origin),” 14 any subclassification of G3 angiosarcomas has so far been impossible. Only recently the receptor for the vascular endothelial growth factor C, VEGFR-3, was discovered and shown to be a specific marker for lymphatic endothelia. In studies on the distribution of renal podoplanin, a novel glomerular membrane mucoprotein, we observed intense immunolabeling of interstitial vessels that showed characteristics of lymphatics. We have confirmed and extended this observation to study the phenotypic makeup of benign and malignant vascular tumors, and we provide suggestive evidence for different classes of angiosarcomas based on their podoplanin content.
Podoplanin was originally found on the surface of rat glomerular epithelial cells (podocytes) and linked to flattening of foot processes (hence the name “podoplanin”) that occurs in glomerular diseases and is related to proteinuria. Database comparison of the DNA-deduced amino acid sequence of rat podoplanin has revealed its identity with a glycoprotein in rat bone (designated E11) that was also observed in vessels of the connective tissue that showed features of lymphatic capillaries. 6 Additional related proteins were found in rat type I alveolar cells, in mouse osteoblasts, osteocytes, and type I epithelial cells of thymus, and in canine MDCK cells, where they serve as receptors for influenza C viruses. 15-18 Podoplanin and its relatives share a high level of identity in their transmembrane domains, but little conserved sequence in their ectodomains apart from several conserved O-glycosylation sites. Thus, podoplanin shows features of a membrane mucoprotein of currently unknown biological function. As heavily O-glycosylated mucoproteins were recently identified as counterreceptors for selectins that mediate adhesion of inflammatory cells, 19 it is possible that podoplanin plays a similar role in lymphatic endothelia.
Evidence that podoplanin is specifically expressed by lymphatic endothelia was based on the following findings: (1) Affinity-purified rabbit anti-podoplanin IgG specifically and exclusively immunolabeled endothelial cells of vessels that were clearly distinct from PAL-E-labeled blood vessels. (2) Podoplanin-positive vessels were also selectively stained with anti-VEGFR-3 IgG in double-labeling experiments. (3) Dermal lymphatic capillaries were identified by ultrastructural morphological criteria and found by immunoelectron microscopy to be selectively labeled by anti-podoplanin antibodies. (4) In benign lymphatic tumorous lesions (lymphangioma, hygroma), podoplanin was localized in endothelial cells. (5) Immunoblotting of lymphangiomas with anti-podoplanin IgG revealed a ∼38-kd protein that was identical with podoplanin in human lung and isolated glomeruli. Collectively these data indicate that podoplanin is expressed in lymphatic endothelia and thus provides a valid marker for the study of the phenotypic characteristics of neoplastic endothelial cells.
Immunocytochemical localization of podoplanin in benign lymphatic tumorous lesions (lymphangiomas, hygromas) resulted in intense continuous labeling of endothelial cells lining the ectatic vascular spaces. Podoplanin was also detected by immunoblotting as a 38-kd protein in membrane protein fractions of lymphangiomas. Conversely, endothelial cells of adjacent nonneoplastic blood vessels were strongly labeled with established endothelial markers, such as CD31, and also with anti-podocalyxin antibodies, but not with anti-podoplanin IgG. Thus, it appears that immunostaining of lymphatic and blood endothelial markers were, in essence, mutually exclusive on lymphangioma cells. This contrasts with Kaposi’s sarcoma, in which cells lining the proliferative vascular channels reacted with both lineage markers. The observation that in the early stages of tumor growth virtually all of the malignant endothelial cells expressed podoplanin provides further evidence that Kaposi’s sarcoma spindle cells are related to lymphatic endothelium. 9,20
Malignant tumors derived from endothelial cells are known to express endothelial antigens in varying quantities. This appears to be related to the ratio of vasoformative to poorly vasoformative to solid areas within a given tumor and also to the sites of tumor origin. 2,3,21-25 Generally, in vascular tumors the degree of differentiation is inversely related to the lesion’s expression of markers specific for the assumed lineage. As a diagnostic aid to distinguish angiosarcoma from other types of sarcomas, a large panel of antibodies specific for vascular endothelia such as factor VIII-related antigen, CD31, and CD34 were used. 26,27 However, this approach fails to disclose mixed phenotypic lineages of hemangiosarcoma or lymphangiosarcoma components in individual high-grade sarcomas, 1-3 and in particular in the rare tumors arising from areas rich in lymphatic vessels and/or persistent chronic lymphedema (lymphedema-associated angiosarcoma). 14,28,29
Here we have used podoplanin as a specific marker of lymphatic endothelium to unravel the relative fractions of lymphatic and vascular differentiated tumor cells in common and epitheloid G3 angiosarcomas and in early Kaposi’s sarcomas. Angiosarcoma cells were identified by cellular atypia, atypical mitotic figures and loss of cellular cohesion, and shedding of the tumor cells into the vascular clefts. These criteria clearly excluded the possibility that reactive nontumorous lymphatic vessels were mistaken for tumor cells. Because the sample of cases is very small (n = 16) in this preliminary trial, it is premature to speculate about the expression of podoplanin (ie, contribution of the lymphatic phenotype) as a criterion for classifying angiosarcomas. Intriguingly, the large majority of podoplanin-expressing tumor cells also coexpressed vascular endothelial markers podocalyxin and CD31. These findings indicate that a variable fraction of angiosarcoma cells within any given tumor displays the characteristics of both endothelial phenotypes. It is not clear whether this indicates different sites of origin or dysregulated phenotypic expression due to malignant transformation. An alternative explanation could be that the tumor cells derive from immature endothelial precursor cells.
Collectively, the results of this study establish that podoplanin is a valid marker for lymphatic endothelia. Using specific antibodies as immunohistochemical tools, we have found that podoplanin expression is preserved in endothelia of all benign lymphatic tumorous lesions and all Kaposi’s sarcomas examined. By contrast, 10 of 11 G3 angiosarcomas contained only variable fractions of podoplanin-expressing tumor cells that also coexpressed markers of blood vessel phenotype. It remains to be determined in a larger number of cases whether lymphatic differentiation and podoplanin expression could serve as reliable criteria for recognizing subgroups of G3 angiosarcomas of clinical and biological relevance. The expression of podoplanin in a subset of angiosarcoma tumor cells could suggest their derivation from a common precursor of lymphatic and blood vascular endothelial cells.
Note Added in Proof
In a recent manuscript, it was reported that podoplanin was also detected in endothelia of HIV-associated cutaneous Kaposi’s sarcomas. Weninger W, et al, “Expression of VEGF-3 and podoplanin suggests a lymphatic endothelial origin of Kaposi’s sarcoma tumor cells.” Lab Invest 1999 (in press).
Acknowledgments
We thank Anny Hovorka, Helga Poczewski, and Sabine Rych for excellent technical assistance and Anton Jäger for assistance with the graphical work.
Footnotes
Address reprint requests to Dr. D. Kerjaschki, Department of Clinical Pathology, University of Vienna, AKH, Währinger Gürtel 18–20, A-1090 Vienna, Austria. E-mail: dontscho.kerjaschki@akh-wien.ac.at.
Supported by SFB 005, Projects 7 and 11, from the Austrian Fonds zur Förderung der Wissenschaftlichen Forschung (to D. K. and E. T.). H. K. was supported by a fellowship from the Leukemia Society of America (Grant # 5015–94).
References
- 1.Holden CA, Spaull J, Das AK, McKee PH, Jones EW: The histogenesis of angiosarcoma of the face and scalp: an immunohistochemical and ultrastructural study. Histopathology 1987, 11:37-51 [DOI] [PubMed] [Google Scholar]
- 2.Oshawa M, Naka N, Tomita Y, Kawamori D, Kanno H, Aozasa K: Use of immunohistochemical procedures in diagnosing angiosarcoma: evaluation of 98 cases. Cancer 1995, 75:2867-2874 [DOI] [PubMed] [Google Scholar]
- 3.Orchard GE, Zelger B, Jones EW, Jones RR: An immunocytochemical assessment of 19 cases of cutaneous angiosarcoma. Histopathology 1995, 28:235-240 [DOI] [PubMed] [Google Scholar]
- 4.Schlingenmann RO, Dingjan GM, Emeis JJ, Blok J, Warnaar SO, Ruiter DJ: Monoclonal antibody PAL-E specific for endothelium. Lab Invest 1985, 52:71-76 [PubMed] [Google Scholar]
- 5.Breiteneder-Geleff S, Matsui K, Soleiman A, Meraner P, Poczewski H, Kalt R, Schaffner G, Kerjaschki D: Podoplanin, a novel 43-kd membrane protein of glomerular epithelial cells, is down-regulated in puromycin nephrosis. Am J Pathol 1997, 151:1141-1152 [PMC free article] [PubMed] [Google Scholar]
- 6.Wetterwald A, Hofstetter W, Cecchini MG, Lanske B, Wagner C, Fleisch H, Atkinson M: Characterization and cloning of the E11 antigen, a marker expressed by rat osteoblasts and osteocytes. Bone 1996, 18:125-132 [DOI] [PubMed] [Google Scholar]
- 7.Kaipainen A, Korhonen J, Mustonen T, van Hinsbergh VWM, Fang GH, Dumont D, Breitman M, Alitalo K: Expression of the fms-like tyrosine kinase 4 gene becomes restricted to lymphatic endothelium during development. Proc Natl Acad Sci USA 1995, 92:3566-3570 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Jeltsch M, Kaipainen A, Joukov V, Meng X, Lakso M, Rauvala H, Swartz M, Fukumura D, Jain RK, Alitalo K: Hyperplasia of lymphatic vessels in VEGF-C transgenic mice. Science 1997, 276:1423-1425 [DOI] [PubMed] [Google Scholar]
- 9.Jussila L, Valtola R, Partanen TA, Salven P, Heikkilä P, Matikainen M-T, Renkonen R, Kaipainen A, Detmar M, Tschachler E, Alitalo R, Alitalo K: Lymphatic endothelium and Kaposi’s sarcoma spindle cells detected by antibodies against the vascular endothelial growth factor receptor-3. Cancer Res 1998, 58:1599-1604 [PubMed] [Google Scholar]
- 10.Horvat R, Hovorka A, Dekan G, Poczewski K, Kerjaschki D: Endothelial cell membranes contain podocalyxin, the major sialoprotein of visceral glomerular epithelial cells. J Cell Biol 1986, 102:484-491 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kerjaschki D, Sharkey DJ, Farquhar MG: Identification and characterization of podocalyxin, the major sialoprotein of the renal glomerular epithelial cell. J Cell Biol 1984, 98:1591-1596 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Carlemalm E, Garavito M, Villiger W: Resin development for electron microscopy and an analysis of embedding at low temperature. J Microsc 1982, 126:123. [DOI] [PubMed] [Google Scholar]
- 13.Kerjaschki D, Ojjha PP, Susani M, Horvat R, Binder S, Hovorka A, Hillemans P, Pytela R: A β1-integrin receptor for fibronectin in human kidney glomeruli. Am J Pathol 1989, 134:481-489 [PMC free article] [PubMed] [Google Scholar]
- 14.Calonje E, Fletcher CDM: Tumors of blood vessels and lymphatics. Fletcher CDM eds. Diagnostic Histopathology of Tumors. 1995, :pp 43-77 Churchill Livingstone, New York [Google Scholar]
- 15.Rishi AK, Joyce-Brady M, Fisher J, Dobbs LG, Floros J, VanderSpek J, Brody JS, Williams MC: Cloning, characterization, and developmental expression of a rat lung alveolar type I cell gene in embryonic endodermal, and neural derivatives. Dev Biol 1995, 167:249-230 [DOI] [PubMed] [Google Scholar]
- 16.Nose K, Saito H, Kuroki T: Isolation of a gene sequence induced later by tumor-promoting 12-O-tetradecanoylphorbol-13-acetate in mouse osteoblastic cells (MC3T3–E1) and expressed constitutively in ras-transformed cells. Cell Growth Differ 1990, 1:511-518 [PubMed] [Google Scholar]
- 17.Farr A, Nelson A, Hosier S: Characterization of an antigenic determinant preferentially expressed by type I epithelial cells in the murine thymus. J Histochem Cytochem 1992, 40:651-664 [DOI] [PubMed] [Google Scholar]
- 18.Zimmer G, Klenk HD, Herrler G: Identification of a 40-kd cell surface sialoglycoprotein with the characteristics of a major influenza C virus receptor in a Madin-Darby canine kidney cell line. J Biol Chem 1995, 270:17815-17822 [DOI] [PubMed] [Google Scholar]
- 19.Sasseti C, Tangemann K, Singer MS, Kershaw DB, Rosen SD: Identification of podocalyxin-like protein as a high endothelial venule ligand for L-selectin: parallels to CD34. J Exp Med 1998, 187:1965-1975 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Beckstead JH, Wood GS, Fletcher V: Evidence for the origin of Kaposi’s sarcoma from lymphatic endothelium. Am J Pathol 1985, 119:294-300 [PMC free article] [PubMed] [Google Scholar]
- 21.Traweek ST, Kandalaft PL, Mehta P, Battifora H: The human hematopoietic progenitor cell antigen (CD 34) in vascular neoplasia. Am J Clin Pathol 1991, 96:25-31 [DOI] [PubMed] [Google Scholar]
- 22.Orchard GE, Jones EW, Jones RR: Immunohistochemistry in the diagnosis of Kaposi’s sarcoma and angiosarcoma. Br J Biomedical Sci 1995, 52:35-49 [PubMed] [Google Scholar]
- 23.De Young BR, Swanson PE, Argenyi ZB, Ritter JH, Fitsgibbon JF, Stahl DJ, Hoover W, Wick MR: CD31 immunoreactivity in mesenchymal neoplasms of the skin, and subcutis: report of 145 cases and review of putative immunohistologic markers of endothelial differentiation. J Cutan Pathol 1995, 22:215-222 [DOI] [PubMed] [Google Scholar]
- 24.Appleton MAC, Attanoos RL, Jasani B: Thrombomodulin as a marker of vascular and lymphatic tumours. Histopathology 1996, 29:153-157 [DOI] [PubMed] [Google Scholar]
- 25.Pammer J, Weninger W, Mazal PR, Horvat R, Tschachler E: Expression of the CD40 antigen on normal endothelial cells and in benign and malignant tumours of vascular origin. Histopathology 1996, 29:517-524 [DOI] [PubMed] [Google Scholar]
- 26.Albelda SM, Muller WA, Buck CA, Newman PJ: Molecular and cellular properties of PECAM-1 (endoCAM/CD31): a novel vascular cell-cell adhesion molecule. J Cell Biol 1991, 114:1059-1068 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Beschorner WE, Civin CI, Strauss LC: Localization of hematopoietic progenitor cells in tissue with the anti-My-10 monoclonal antibody. Am J Pathol 1985, 119:1-4 [PMC free article] [PubMed] [Google Scholar]
- 28.Sordillo PP, Chapman R, Hajdu SI, Magill GB, Golbey RB: Lymphangiosarcoma. Cancer 1981, 48:1674-1679 [DOI] [PubMed] [Google Scholar]
- 29.Kindblom LG, Stenman G, Angervall L: Morphological and cytogenetic studies of angiosarcoma in Stewart-Treves syndrome. Virchows Archiv A Pathol Anat 1991, 419:439-445 [DOI] [PubMed] [Google Scholar]