

Abstract
This study focuses on the mechanism of action of N-alkylthio β-lactams, a new family of antibacterial compounds that show promising activity against Staphylococcus and Bacillus microbes. Previous investigations have determined that these compounds are highly selective towards which bacteria they target, and possess completely unprecedented structure-activity profiles for a β-lactam antibiotic. Unlike penicillin, which inhibits cell wall crosslinking proteins and affords a broad spectrum of bacteriocidal activity, these N-thiolated lactams are bacteriostatic in their behavior and act through a different mechanistic mode. Our current findings indicate that the compounds react rapidly within the bacterial cell with co-enzyme A (CoA) through in vivo transfer of the N-thio group to produce an alkyl-CoA mixed disulfide species, which then interferes with fatty acid biosynthesis. Our studies on coenzyme A disulfide reductase show that the CoA thiol redox buffer is not perturbed by these compounds; however, the lactams appear to act as prodrugs. The experimental evidence that these β-lactams inhibit fatty acid biosynthesis in bacteria, and the elucidation of coenzyme A as a primary cellular target, offers opportunities for the discovery of other small organic compounds that can be developed as therapeutics for MRSA and anthrax infections.
Keywords: N-thiolated β-lactams, MRSA, Bacillus anthracis, mechanism of action, glutathione, coenzyme A
Introduction
N-Alkylthio β-lactams represent a recently discovered family of antibacterial compounds that selectively inhibit the growth of methicillin-resistant Staphylococcus aureus (MRSA)1–6 and Bacillus anthracis7 (Figure 1). Despite their apparent structural similarities to the penicillins and other members of the β-lactam family of antibiotics, these N-thiolated β-lactam compounds behave in many ways differently to all other known antibacterial β-lactams. First, these N-thiolated compounds only target a few genera of bacteria such as Staphylococcus and Bacillus, yet show anticancer properties8–12, and are non-lethal to healthy human cells (fibroblasts).2 Secondly, the bioactivity of these N-thiolated lactams islargely insensitive to changes in structure of the ring substituents at the C3 and C4 centers of the lactam ring, strongly suggesting that the compounds exert their antibacterial effects on a completely different cellular target and through a different mode of action than previously studied β-lactam antibacterials. Unfortunately, neither the target nor the mode of action of these new antibacterial agents is presently known.
Figure 1.

Structures of N-alkylthio β-lactams and penicillins, two different families of β-lactam antibiotics.
Results and Discussion
Structure-Activity Studies
Ongoing experiments in our laboratory on the N-thiolated systems have explored the effect of the substituents around the β-lactam ring on antibacterial activity.1–7 Our results so far indicate that the structure-bioactivity patterns for these N-thiolated lactams are completely different to those of other β-lactam antibacterial drugs. In fact, these are the first β-lactam compounds which possess antibiotic properties despite the absence of any ionizable ring functionality (typically a carboxylic acid) within its core structure. A wide variety of substituents at the C3 and C4 positions can be tolerated without significant effects on biological activity, and stereochemistry at these centers is also irrelevant. However, the N-alkylthio moiety is absolutely essential.6 Finding that thiolation of the nitrogen is a prerequisite for activity, we then further investigated the effect of various organothio moieties. While arylthio substituents are tolerated on the nitrogen, small alkylthio substituents (four carbons or less) provide for the strongest bioactivity. Further, alkylthio groups bearing a single branched site (i.e., isopropyl and sec-butyl) produce better activities against S. aureus than do the straight chain n-alkylthio analogs, while a high degree of branching (i.e., t-butylthio) results in a significant decrease in activity. The oxidation state of the sulfur also plays a substantial role in the activity of the compounds: N-sulfenyl and N-sulfinyl groups provide for much more elevated anti-MRSA activities than N-sulfonyl or N-sulfonic acid functionalities.6
The clear dependency of substituents at the lactam nitrogen, as compared to the C3 and C4 ring centers, suggests that this organothio moiety is central to the compound’s biological mechanism. By contrast, the broad tolerance of substituents at the other two positions suggests that these groups may not play a direct role in the binding or reactivity of the drug with a biological target, but more likely, may influence bacterial cell uptake and membrane permeability. Further, it was recently shown that structurally related N-alkylthio-2-oxazolidinones (Figure 2) closely mirror these same trends, demonstrating that the four-membered β-lactam ring itself is not even essential for bioactivity.13
Figure 2.

Structure of N-alkylthio 2-oxazolidinone antibiotics.
Genera-Selective Antibacterials
Antimicrobial screening of the various lactam derivatives outlined above included a fairly wide range of Gram-positive and Gram-negative bacteria. Approximately 65 microbes of different genera or species were examined.14 Although there were fluctuations in the levels of bioactivity seen among the different lactam variants against any particular bacterium, the data was consistent in terms of which microbes were most susceptible. This indicates that these N-thiolated lactams function as narrow-spectrum antibiotics for Staphylococcus, Micrococcus, and Bacillus bacteria, although they also are somewhat less powerful inhibitors of Bacteroides, Streptococcus, Neisserria gonorrhoeae, Salmonella typhimurium, Vibrio cholerae, and Mycobacterium tuberculosis. On the other hand, these compounds were found to be consistently ineffective against Enterobactor cloacae, Escherichia coli, Klebsiella pneumoniae, Listeria monocytogenes, and Proteus mirabilis. Thus, activity does not appear to be related to whether the microbe is Gram-positive or Gram-negative, unlike that seen for many other antibacterial drugs. Peculiarly, the most sensitive microbes were those from four distinct taxonomic orders defined by their genetic, morphological and metabolic traits. Although nine genera in total were found to be susceptible to the lactams, few taxonomic relationships could be used to ascertain the spectrum of activity. The class of bacteria most sensitive to the N-thiolated β-lactams was the "Bacilli", which consists of two orders: Bacillales and Lactobacillales. Susceptible members of this taxonomic class included species of Bacillus, Staphylococcus and Streptococcus. Enterococcus, Lactococcus and Listeria which are also affiliated with the "Bacilli" were mostly insensitive to the drugs. This sporadic but narrow-spectrum of activity supports the conclusions of the above-described structure-activity data, that the compounds are acting in a different manner from traditional broad spectrum β-lactam antibiotics.
Effects of the lactam on cell growth were studied by measuring cell survival for methicillin-susceptible S. aureus (MSSA) and MRSA, respectively, when cultured in the absence or presence of lactam 1 over a 2 hour time frame. In the absence of lactam 1, MSSA and MRSA grew logarithmically; in the presence of 1, bacterial growth was immediately halted. While reproduction ceased at the MIC level of the lactam as well as at 10x MIC, the number of viable cells remained constant throughout the duration of the experiment. Consequently, bacteria growth was clearly inhibited by lactam 1, but little to no decrease in cell population was observed. These data provide substantial evidence that even at high drug concentrations, N-methylthio β-lactams are bacteriostatic agents toward staphylococci, and do not reduce cell counts that would be expected if the effects were bacteriocidal.14
Thus, the finding that N-thiolated β-lactams are bacteriostatic agents possessing a narrow range of activity strongly implies that the biological target is internalized within the cell. Preliminary information on the cellular target of a new antibiotic can be extracted from the intrinsic nature of other known antibacterial drugs: those antibiotics that inhibit intracellular processes such as protein synthesis (chloramphenicol) or secondary metabolites (sulfa drugs) are generally bacteriostatic agents whereas those that disrupt the outer periphery of cells such as the bacterial membrane (polymyxins, bacitracin, cationic lipoproteins), the cell wall (penicillin, vancomycin) or directly damage DNA (metronidazole) are bactericiodal. Thus, our first efforts to determine mechanism of action were directed at identifying the chemical pathway by which the lactams exert their effects, and which cellular process or processes are most greatly affected by the lactams.
Considerations Regarding the Compound’s Mode of Action
To account for the antibiotic properties of these compounds, at least three types of reactions between N-methylthio β-lactams and a biological nucleophile (Nu−) can be envisioned: acylation (ring opening), alkylation, and sulfenylation (Scheme 1). For the β-lactam class of molecules, it is naturally assumed that nucleophilic addition occurs on the 3 carbonyl center of the reactive four-membered ring. As in the case of penicillins and cephalosporins, the four-membered lactam ring is susceptible to this type of acylation mechanism through recognition by and reaction with an active site serine of cell wall transpeptidases.15,16 It is through this mechanism, the covalent interaction with the penicillin-binding proteins (PBP’s), that β-lactam antibiotics are known to eradicate bacteria. During this process, a serine residue in the active site of the PBP’s adds to the carbonyl center of penicillin to create a stable ester linkage. The addition reaction occurring on the hydroxyl of serine is enhanced by ring strain and a carboxylic acid moiety located in proximity to the β-lactam carbonyl. Repositioning or eliminating the carboxylate negates the ability of the antibiotics to irreversibly bind to the PBPs. Therefore, the lack of an acidic functionality in the N-methylthio β-lactams suggests that binding within the PBP active site, and subsequent nucleophilic attack on the carbonyl carbon, would be extremely unlikely. Moreover, chemical studies verified the ring's stability of N-thiolated β-lactams. Compound 1 is stable to cleavage in water when exposed to equimolar amounts of potassium hydroxide and serine. In addition, 1 is completely unreactive to commercial β-lactamases even after several days in pH 7.4 buffer. Moreover, lactam 1 does not inhibit penicillinase in catalyzing the hydrolysis of penicillin G.
Scheme 1.

Probable reaction pathways of N-thiolated β-lactams with nucleophiles.
Further evidence of the stability of N-thiolated lactams to cleavage by bacterial penicillinases is shown in Figure 3. Two experiments are shown,: on the left (figure a), 20 ug of lactam 1 was added to one well (left side well) bored into the agar, and 20 μg of penicillin G was added to the other well (right side), and the plate was inoculated with a penicillin-susceptible form of S. aureus. After 24 hours of incubation, large growth inhibition zones were observed for both antibiotics. This experiment was then repeated (figure b), but this time 20 μg of penicillinase enzyme was mixed into the agar prior to incubation with MRSA. The consequences of this are clear: penicillin G (right well) loses all of its antibiotic activity while lactam 1 retains all of its anti-MRSA activity.
Figure 3.

Comparison of bioactivities of lactam 1 and penicillin G toward MRSA in the absence (a) versus presence (b) of bacterial penicillinase. In (a), when no penicillinase is present, lactam 1 and penicillin each produces large zones of growth inhibition. However, when penicillinase is present in the growth media (b), penicillin G is completely devoid of activity while lactam 1 retains its full activity.
However, to prove unequivocally that the N-thiolated β-lactam was not inhibiting the PBP’s, light microscopy and electron microscopy were used to probe for damage or thinning of the cell wall in S. aureus treated with high concentrations of lactam 1.

Bacteria exposed to antibiotics that disrupt the cell wall (ie, β-lactams) or cytoplasmic membrane (ie, polymyxins) can be observed by scanning electron microscopy (SEM) for physiological damage elicited by the drugs. Cultures of S. aureus inoculated with either lactam 1 or penicillin G were inspected by SEM for changes in cell size and appearance in comparison to a culture with no antibiotic (Figure 4).14 The SEM samples were prepared from Kirby-Bauer agar diffusion plates by incision of the agar along the outermost regions of cleared zones (see Figure 3a) where sub-lethal doses of the drugs are present and bacterial growth is only partially inhibited.17 The first image in Figure 4a portrays the appearance of S. aureus grown with no antibiotic present. In its natural state, staphylococci grow to about 1 μm in diameter as grape-like clusters of spherical-shaped cells. Cultures with lactam 1 (Figure 4b) also show the presence of surviving colonies of bacteria which resemble S. aureus in its natural state, appearing spherical and uniform with no apparent deformities. This provided the first substantial evidence that N-thiolated β-lactams do not inhibit bacterial transpetidases nor cause rupturing of the cytoplasmic membrane in staphylococcus in the manner of penicillin, which produce cocci that are wrinkled, concaved, and sheared due to deterioration of the bacterial cell wall.
Figure 4.

Scanning electron microscope images of S. aureus cultured on agar in the presence of (a) no antibiotic, (b) lactam 1, and (c) penicillin G. These images show that cells which survive explosure to N-thiolated lactam 1 (b) look normal in morphology and clustering behavior, while those treated with penicillin (c) suffer severe damage indicative of inhibition of cell wall biosynthesis.
The inhibition of cell wall biosynthesis in Gram-positive bacteria can also be detected by light microscopy upon Gram-staining of the cells. This allows bacteria to be readily distinguished by the thickness and structural integrity of their cell walls. Figure 5 shows the results of these experiments, and the appearances of S. aureus cells exposed to (a) no antibiotic, (b) lactam 1, and (c) penicillin G after staining.14 Untreated S. aureus possesses a well-developed, thick cell wall which retains the crystal-violet stain upon decolorization (Figure 5a). Cells treated with lactam 1 likewise stain uniformly purple, indicating a fully intact and mature cell wall (Figure 5b). However, bacteria treated with penicillin G stain mostly pink, indicative of a weakened or sheared cell wall (Figure 5c). This confirms the results of the SEM experiments, which indicate that the N-thiolated lactams inhibit S. aureus through a pathway other than interfering with cell well biosynthesis.
Figure 5.

Light microscopy of S. aureus exposed to (a) no antibiotic, (b) lactam 1, and (c) penicillin G. Staphylococci which appear purple after Gram-staining (a, b) have intact cell walls; those which stain pink (c) have deformed cell walls.
Next, we considered the possibility that the compounds could serve as alkylating agents towards cellular nucleophiles, as depicted hypothetically in Scheme 2. Initial structure-activity studies, however, did not support this. It could be predicted that nucleophilic attack on the carbon of the alkylthio moiety could be deterred by placement of bulkier, branched alkyl groups on the sulfur center. In fact, what we observed was completely opposite to this prediction, that the N-ethylthio, N-isopropylthio, and N-sec-butylthio lactams were increasingly more active, not less active, than the N-methylthio analogue 1.6 But we decided to look into this further by studying the ability of lactam 1 to serve as a DNA alkylating agent in vitro. Most known nucleic acid alkylating drugs react with nucleotides to inhibit DNA replication or compromise the integrity of the super helices. Examples of DNA alkylating agents include leinamycin19 and mitomycin20, which exhibit bactericidal behavior. The N-methylthio lactam compounds, however, are bacteriostatic and thus seem unlikely to function as alkylating agents. Nevertheless, experiments were conducted to confirm this.
Scheme 2.

Potential nucleophilic attack on the alkyl residue of the N-alkylthio side chain.
First, studies were run to determine if N-thiolated β-lactams could directly cause strand breakage of supercoiled DNA.14 Plasmid pBR322 was incubated in the presence of varying amounts of lactam 1 (5–100 μM) at 37°C in sodium phosphate buffer (50 mM; pH 7.4) for 24 hours, and double strand (fragmentation) and single strand (linearization) breakage was then analyzed by agarose gel electrophoresis containing 1% ethidium bromide (Figure 6).
Figure 6.

Supercoiled DNA treated with lactam 1 at 5–100 μM. Plasmid pBR322 (0.5 μg) was incubated with lactam 1 at 37°C in sodium phosphate buffer (50 mM, pH 7.4) for 24 hrs and analyzed by agarose gel electrophoresis (ethidium bromide staining). Lane 1: marker. Lane 2: pBR322. Lane 3: pBR322 + DMSO. Lane 4. pBR322 + 5 μM 1. Lane 5: pBR322 + 10 μM 1. Lane 6: pBR322 + 25 μM 1. Lane 7: pBR322 + 50 μM 1. Lane 8: pBR322 + 100 μM 1. Lane 9: marker.
The results show that in lanes 2 and 3, two bands were observed for the plasmid in the absence of lactam 1. With the supercoiled pBR322 giving rise to the denser band, a weak band of relaxed or "nicked" DNA was formed by the electrical current and passage through the gel. Lanes 3–8 contained plasmid samples treated with various concentrations of lactam 1. It was evident that the compound did not cause fragmentation or relaxation of the superhelix. The bands were equivalent in location and illumination to those observed for the untreated plasmids (lanes 2 and 3) thus indicating that chemical modification of the DNA did not occur in the presence of 5–100 μM of lactam 1.
Recognizing that DNA alkylators are often in a prodrug form, requiring chemical activation before lethal effects on the nucleotide can occur, we carried out some additional experiments. Leinamycin, for example, is transformed into a potent alkylating antibiotic by a thiol-mediated reaction with cysteine or glutathione.21 Cells susceptible to leinamycin possess thiol-rich intracellular environments which we considered may also be needed for N-thiolated β-lactams to be able to alkylate their cellular target. A second study was therefore performed to examine the stability of plasmid pBR322 (5 μM) to treatment with a large amount of lactam 1 (100 μM) in the presence of various thiols (100 μM) (Figure 7). In lane 1, the plasmid was treated with the lactam for 24 hours.
Figure 7.

Gel electrophoresis analysis of super-coiled DNA treated with N-thiolated lactam 1 in the absence or presence of thiols. Plasmid pBR322 (0.5μg) was incubated with lactam 1 at 37°C in sodium phosphate buffer (50 mM, pH 7.2) for 24 hrs and analyzed by agarose gel electrophoresis (ethidium bromide staining). Restriction digests conducted with EcoR1 were performed at 37°C for 1 hr. Lane 1: pBR322 + 100 μM 1. Lane 2: pBR322 + 100 μM glutathione. Lane 3: pBR322 + 100 μM glutathione + 100 μM 1. Lane 4: pBR322 + 100 μM DTT. Lane 5: pBR322 + 100 μM DTT + 100 μM 1. Lane 6: pBR322 + 100 μM 2-mercaptoethanol. Lane 7: pBR322 + 100 μM 2-mercaptoethanol + 100 μM 1. Lane 8: linearized pBR322 (EcoR1digest). Lane 9: pBR322 + EcoR1 + 100 μM 1. Lane 10: pBR322 + DMSO. Lane 11: pBR322 + EcoR1 + DMSO.
As expected, the supercoil remained completely intact with only the antibiotic present. DNA samples loaded in lanes 2–7 were incubated with glutathione (lanes 2, 3), dithiothreitol (DTT; lanes 4, 5), or β-mercaptoethanol (lanes 6, 7). Samples which also contained the lactam are located in lanes 3, 5, and 7. To verify the cleavability of the plasmid, pBR322 was also linearized with the endonuclease, EcoR1. Samples containing the digested DNA are located in lanes 8 and 11. A loading comprised of lactam 1 and EcoR1 was applied to lane 9 to establish if the compound could function as an endonuclease inhibitor. After 24 hrs, the superhelix of the DNA appeared completely unaltered by the lactam either in the presence, or absence, of glutathione, DTT, or β-mercaptoethanol. The bands of lanes 1–7 were comparable to that of the antibiotic-free plasmid loading (lane 10), indicating no DNA cleavage by the N-thiolated lactam.
Disruption of the super-helix by chemical modifications of nucleotide bases can cause the arrest of DNA replication. To further substantiate that N-thiolated β-lactams are not alkylating native nucleic acids or affecting nucleotide assimilation into double-stranded DNA, pulse-labeling experiments with 3H-thymidine were next performed to monitor DNA replication in living bacteria, in the presence versus in the absence of lactam 1 and controls.14 The results of the radio- uptake experiments are shown in Figure 8. Thymidine incorporation was measured as a percent of the control for the three different antibiotics. As expected, ciprofloxacin was the most potent, inhibiting 65% of thymidine utilization after 30 minutes. Conversely, with lactam 1 and penicillin G present, ≥90% of the nucleotide was converted into DNA. Lactam 1 appeared not to affect thymidine incorporation for at least the first 30 minutes. After that, however, the rate of thymidine incorporation leveled off.
Figure 8.

The rate of radiolabeled thymidine incorporation into growing S. aureus cells (relative to an untreated control) for ciprofloxacin (CIP) and lactam 1.
Gene expression is a biological process targeted by many families of antibiotics including tetracyclines, macrolides, aminoglycosides, streptogramins, and oxazolidinones. In most cases, drugs that affect transcription or translation are bacteriostatic, suggesting that one of these processes may be inhibited by N-thiolated β-lactams. To test this hypothesis, incorporation of 3H-uridine and 3H-isoleucine into RNA and proteins, respectively, was measured in a broth culture of S. aureus treated with lactam 1. RNA labeling studies14 with 3H-uridine were conducted to assess the influence of lactam 1 on transcription in S. aureus. Cultures were grown to early logarithmic phase and inoculated with 3H-uridine and lactam 1, rifampicin (an inhibitor of RNA synthesis), or DMSO (Figure 9). As expected, rifampicin immediately blocked the transcription of RNA, with nucleotide uptake being less than 20% of that for the DMSO control after 20 minutes. Lactam 1 was also found to modulate uridine incorporation, though certainly not to the extent or rapidity of rifampicin. After 20 minutes, the nucleotide uptake in the presence of 1 remained nearly 80% of the untreated (DMSO) control, suggesting that transcription can not be the primary process targeted by the lactam.
Figure 9.

The rate of radiolabeled uridine incorporation into growing S. aureus cells (relative to an untreated control) in the presence of rifamicin and lactam 1.
To determine if the primary role of growth inhibition by N-thiolated β-lactams could be as a transcription inhibitor, the rate of protein synthesis was monitored by the uptake of 3H-isoleucine.14 An overnight culture of S. aureus was grown to early logarithmic phase and inoculated with 3H-isoleucine (5 μCi/ml). The cultures were then treated with either lactam 1 or chloramphenicol (a translation inhibitor), or DMSO as a control (Figure 10).
Figure 10.

The rate of radiolabeled isoleucine incorporation into growing S. aureus cells (relative to an untreated control) in the presence of chloramphenicol (CHL), lactam 1, and DMSO.
For the initial 10 minutes, a linear progression of isoleucine incorporation was observed for the culture containing lactam 1, but beyond that, effects on protein synthesis became much more pronounced.22 While these studies indicate that the lactams have a delayed effect on protein synthesis in bacteria, we decided to then investigate the effect of the lactams on fatty acid biosynthesis in S. aureus.23–26 In this case, these experiments demonstrated that treatment of S. aureus cells with lactam 1 either at the MIC (10 μg/mL) or twice the MIC (20 μg/mL) had a striking effect on the rate of uptake of radiolabeled acetate compared to the controls, DMSO and penicillin G (Figure 11). This indicates that a primary mode of action for the compounds in S. aureus is inhibition of fatty acid biosynthesis.
Figure 11.
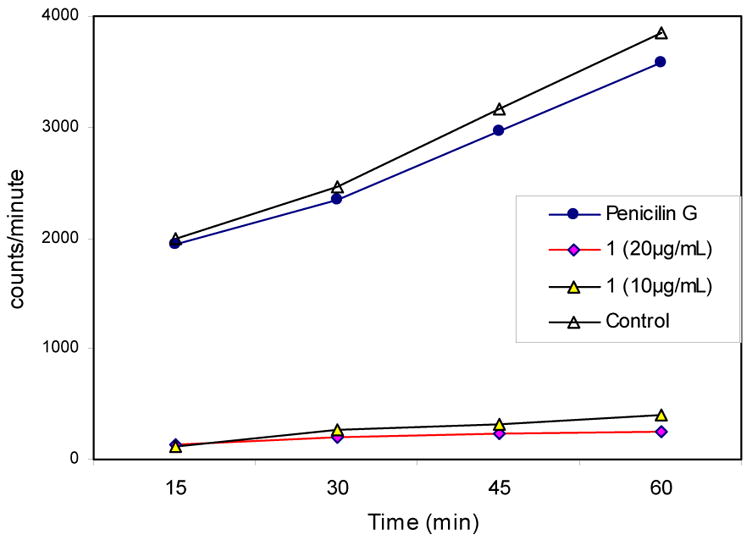
Effect of N-alkylthio β-lactam 1 on fatty acid synthesis. Measurement of uptake of radiolabeled acetate. Incidence of radiation versus time for S. aureus treated with: penicillin G (2 μg/mL, 2xMIC), lactam 1 (20 μg/mL, 2xMIC), (c) compound 1 (10 μg/mL, 1xMIC), (d) DMSO control in the presence of 3H acetate.
N-Thiolated β-Lactams as Thiolating Agents
N-Thiolated β-lactams have similar sulfenylating properties to the sulfur transfer reagents used for their preparation (Figure 12). Previous chemical studies of N-methylthio lactams revealed that the NS bond is resistant to cleavage by most types of nucleophiles, but is susceptible to thiophiles.27,28
Figure 12.

Electrophilic sulfur-transfer reagents.
Further analysis of the cultures of S. aureus treated with sub-MIC amounts of N-alkylthio β-lactam 1 under steady state growth conditions showed the formation of the dethiolated β-lactam in the lysate, which could easily be isolated from the growth media by extraction with ethyl acetate. The 1H NMR spectrum of the crude isolate revealed the clean formation of the N-protio β-lactam as represented by the broad singlet positioned at 6.37 ppm (Figure 13).14 The loss of the N-methylthio substituent was also confirmed by the absence of the methylthio protons signal at 2.41 ppm. The evidence from this and prior experiments suggests that the methylthio moiety is cleaved by a protein or metabolite following entry of the lactam into the cytoplasm. This provides clear evidence that a primary metabolic pathway for the N-thiolated lactams proceeds through a thiol transfer process (Scheme 3). In light of these findings, we turned to consider the likelihood that an intracellular thiol was the recipient of this transfer to produce a mixed disulfide within the cell.9
Figure 13.

1H NMR spectra of (a) lactam 1 prior to inoculation of S. aureus and (b) lactam 1 following incubation with S. aureus for 24 hours. The N-H signal noted in (b) is indicative of a thio-transfer reaction.
Scheme 3.

Thio-transfer from the lactam 1 to a nucleophilic reactant present in the lysate of cultured S. aureus.
We initially considered the likelihood that N-thiolated β-lactams may inhibit bacterial growth by transfer of the alkylthio moiety onto a sulfhydryl-containing target such as a cysteine residue in a protein. A survey conducted in our laboratory using the 20 essential L-amino acids showed that cysteine was the only one that reacted directly with lactam 1 under physiological conditions (pH 7.4), affording the N-dethiolated β-lactam. Similar results were obtained with glutathione, a tripeptide that is found in the cytoplasm of most bacteria and in human cells (Figure 14).
Figure 14.
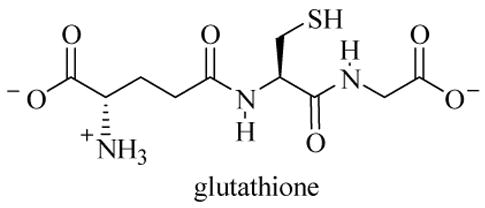
Structure of glutathione.
To evaluate whether glutathione could affect the anti-staphylococcal properties of N-thiolated β-lactams, a Kirby-Bauer diffusion experiment was run.22 Equimolar amounts of glutathione and lactam 1 were added to the same well while only the lactam was added to another well. Following overnight incubation, the plate inoculated with S. aureus was examined for growth inhibition. As expected, a large zone of inhibition was found around the well containing the lactam 1, but no zone was found around the well containing lactam 1 that had been co-mixed with glutathione. It was subsequently observed by varying the ratio of lactam-to-glutathione that the antimicrobial activity of N-methylthio lactam 1 against S. aureus can be partially or completely reduced, depending on the quantity of glutathione present in the media.
To further substantiate the neutralizing effect of glutathione on the lactams, an agar plate was prepared with a 1 mg of glutathione (in pH 7.4 buffer) added into a well located at the center of the plate, and 20 μg of the N-methylthio β-lactams 1–3 were each loaded into three surrounding wells (Figure 15). The ability of glutathione to protect the bacteria from the anti-proliferation effects of these antibiotics was again clearly evident by the presence of concaved zones between the wells containing glutathione (in the center) and the three lactams (on the outside).
Figure 15.

Effect of glutathione (GSH) on the antimicrobial properties of N-thiolated β-lactams 1–3 against S. aureus. Each 6-mm well contains 20 μg of the indicated β-lactam, and the center well contains 1 mg of glutathione. The neutralizing effect of glutathione on the lactams is apparent from the indented regions of growth inhibition after 24 hours of incubation.
We then followed this experiment up with an additional assay to evaluate the stability of other N-alkylthio lactam derivatives to glutathione. Figure 16 compares the relative antibacterial activities of three N-alkylthio-substituted lactams, N-methylthio lactam 1, N-ethylthio lactam 4, and N-sec-butylthio lactam 5 against S. aureus varying in the presence of different amounts of glutathione. For the plate shown in Figure 16a, 20 μg glutathione was added to the wells containing 20 μg of the three lactams, and for the plate shown for Figure 16b, 50 μg of glutathione was added to the three wells. As observed in Figure 16a, N-ethylthio lactam 4 and N-sec-butylthio lactam 5 both possess complete activity against S. aureus in the presence of 20 μg of glutathione, while N-methylthio lactam 1 is completely inactivated. At the higher amount of glutathione (50 μg) shown in Figure 16b, the N-ethylthio lactam also loses all of its activity, while the N-sec-butylthio compound is still strongly active. Thus, these studies show that the differences in biological activity of the different N-alkylthio lactam derivatives seem to be due to the different reactivities of those lactams towards thiophilic nucleophiles.
Figure 16.

Comparison of bioactivities of N-methylthio lactam 1, N-ethylthio lactam 4, and N-sec-butylthio lactam 5 against S. aureus in the presence of glutathione. In (a), each well contains 20 μg of the lactam and 20 μg of glutathione. In b, 50 μg of glutathione is added to each well containing 20 μg of the lactam. Growth inhibition was checked after 24 hours of incubation.
From these experiments, we surmised that the protective role of glutathione likely explains the selectivity that is observed among the different bacterial genera, such that those microbes having the highest levels of glutathione in their cytoplasm are more resistant to the antibacterial effects of these lactams. Closer inspection of the list of bacteria that we tested indicates that the microbes having the greatest susceptibilities to the lactams also are those that reportedly contain low cytoplasmic levels of glutathione.29 In addition, E. coli and P. aeruginosa, which show no susceptibility to these drugs, have high levels of glutathione. The correlations between reported thiol levels and the observed bioactivities of the N-alkylthio β-lactams are shown in Table 1.
Table 1.
Correlation between reported30 percentages of intracellular glutathione and CoA and the observed bacterial susceptibility to N-alkylthio β-lactams (by agar diffusion).
| Species | % GSHa | % CoA | Susceptibility |
|---|---|---|---|
| Staphylococcus aureus | <1 | 67 | highb |
| Bacillus subtilis | <2 | 21 | high |
| Micrococcus luteus | <0.2 | 17 | high |
| Mycobacterium tuberculosis | <0.2 | 14 | mediumd |
| Pseudomonas aeruginosa | 70 | 15 | none |
| Escherichia coli | 80 | 11 | none |
Percentages of GSH and CoA (relative to total thiol content) derived from data.30
high: average zones of inhibition >20 mm.
M. tuberculosis showed no activity in the Kirby-Bauer assays, but showed activity in MIC assays.
The role of glutathione in protecting bacteria from the inhibitory effects of the N-thiolated lactams can now be understood, but what remains unclear is the reason for why bacteria expressing little or no glutathione are so sensitive to the inhibitory effects of the lactams, and what the identity of the cellular target in those bacteria is. It was therefore important to investigate further the cellular target in S. aureus. We tried several different strategies to do this, but the most informative experiment was obtained by the lysate from a culture of S. aureus with resin-bound β-lactam 6 (Scheme 4). This resin was stirred with the N-H β-lactam in the presence of Hunig’s base to afford the resin-bound N-thiolated lactam 6. The identity of the resin-bound lactam (and its phthalimide precursor) was confirmed by infrared spectroscopy. Resin 6 was stirred for 24 hours with the homogenized S. aureus lysate, washed with hot water and ethanol to remove non-bound components, and subsequently reacted with diisobutylaluminum hydride (DIBAL) in anhydrous methylene chloride to cleave off any covalently-bound constituents. Purification of the resulting cleavage product and analysis by HPLC and NMR revealed the presence of coenzyme A (CoA).
Scheme 4.

Identification of primary cellular target in S. aureus using resin-bound lactam 6.
These results are consistent with a previous report by Fahey (Table 1) that CoA is the predominant small thiol (and thus the major thiophilic nucleophile) in S. aureus29, and our own observation that bacteria with the highest reported levels of CoA (Staphylococcus, Bacillus, and Micrococcus) are most susceptible to the N-alkylthio β-lactams.
The role of coenzyme A disulfide reductase in bacterial cells
The unusually high concentrations of CoA in some bacteria, particularly Staphylococcus and Bacillus, stems from the role of CoA within the thiol redox buffer (Scheme 5). Thiol redox buffer systems are present in all cells, and consist of free cytosolic thiols which are present in high (often millimolar) concentrations. An NAD(P)H-dependent disulfide reductase30–32 regulates the balance of this equilibrium in bacterial cells by converting the disulfide to the reductively active thiol form. Eukaryotes and many prokaryotes utilize glutathione in their thiol-redox buffers. Indeed, glutathione was long thought to be ubiquitous.29 However, recent findings have shown that some prokaryotes, including those in the Staphylococcus and Bacillus genera, do not produce glutathione, and instead rely upon CoA or mycothiol in their thiol redox buffer. This buffer consists of millimolar levels of free CoA and a CoA disulfide reductase, an NADPH-dependent enzyme in the flavoprotein disulfide reductase family. Originally isolated from S. aureus by delCardayre and coworkers in the late 1990’s, CoA disulfide reductase has been thoroughly characterized.30,31 A member of the flavoprotein disulfide reductase family, CoA disulfide reductase contains a single cysteine residue and an integral but noncovalently bound flavin molecule.32 Reduction of CoA disulfide occurs through two steps: thiol-disulfide exchange with the active site cysteine, followed by flavin-mediated hydride transfer from NADPH to reduce the cysteine-CoA disulfide bond and regenerate the active site (Scheme 6).
Scheme 5.

A generalized thiol redox buffer. The thiol-disulfide equilibrium is responsible for absorbing reactive oxygen species (ROS) generated in the cell by oxidative stress, and is closely maintained by an enzyme, disulfide reductase.
Scheme 6.

Mode of action of CoA disulfide reductase.
The correlation between the antibacterial activity of the N-alkylthio β-lactams and the CoA-based thiol-redox buffer system led us to consider whether CoA disulfide reductase might be a key target in the mode of action of these compounds. Despite a coarse resemblance, glutathione disulfide reductase and coenzyme A disulfide reductase are substantially different in mechanism and substrate selectivity, and so it was postulated that the activity of the N-alkylthio β-lactams against CoA-rich bacteria might be a function of these differences. In particular, we hypothesized that the mixed CoA-alkyl disulfide produced by the alkylthio transfer from the β-lactam might inhibit the CoA-dependent pathway in the manner shown in Scheme 7. Although this might take place through a non-covalent inhibition, the asymmetric CoA disulfide might also transfer the alkylthio portion to the active site cysteine for covalent modification.
Scheme 7.

A possible result of transferring the alkylthio residue from the lactam to CoA, leading to the disruption of the CoA disulfide reductase.
To test these hypotheses, asymmetric CoA disulfides 8a-c were prepared33 from free CoA (trilithium salt) and the corresponding alkyl methanethiolsulfonates (Scheme 8).
Scheme 8.

Formation of mixed CoA disulfides 8a-c.
Rates of reduction of the CoA mixed disulfides versus the native CoA disulfide by CoA disulfide reductase were determined spectrophotometrically and the values of Vmax, Km, and kcat were found through best fit analysis35 to the Michaelis-Menton equation (Table 2). In each case, the rates of catalysis of the mixed disulfides were found to be within an order of magnitude of those of the native substrate (CoA disulfide). The most drastic difference in rate of reduction was observed for the sec-butyl CoA mixed disulfide (8c), whose Vmax and kcat were approximately 20% of CoA disulfide. This steady drop in both the Vmax and Km upon incrementally increasing the bulk of the disulfide alkyl residue from methyl to ethyl to sec-butyl reflects diminishing reducibility and increasing stability of the disulfides by the reductase. Despite the retardation of reductase activity, the mixed CoA disulfides do not substantially inhibit CoA disulfide reductase.
Table 2.
Kinetic data for the reduction of CoA disulfides by CoA disulfide reductase. 36
| Compound | Vmax (μM/min) | Km(μM) | kcat(min−1) |
|---|---|---|---|
| CoA-disulfide | 1.9 | 6 | 190 |
| CoA-methyl disulfide (8a) | 2.6 | 8 | 260 |
| CoA-ethyl disulfide (8b) | 1.6 | 5 | 160 |
| CoA-sec-butyl disulfide (8c) | 0.57 | 4 | 57 |
Kinetic data was determined by spectrophotometric measurement (340 nm, 25°C) of the rate of NADPH oxidation in the presence of CoA disulfide reductase (10 nM). Concentrations of disulfides were varied from 2–200 μM.
Although these studies conclude that CoA disulfide reductase is not the primary enzymatic target of the N-alkylthio β-lactams, the capping of the active cysteine in CoA disulfide reductase through formation of an unreactive disulfide remained an intriguing possibility. If mixed CoA disulfides interact with CoA disulfide reductase in such a way that the alkyl thiol is preferentially expelled instead of transferred, then the cysteine-CoA disulfide bond is selectively formed over the cysteine-alkyl disulfide bond. This postulate is supported by the report of delCardayre et al, who noted that in the reaction of CoA-glutathione mixed disulfide with CoA disulfide reductase, the CoA selectively formed the disulfide bond with the enzyme, while free glutathione was released.31 To explore this latter possibility, the rate of catalysis of CoA disulfide reductase in the presence of methylthio methanesulfonate was measured (Scheme 9). This thio-transfer compound was indeed found to be a viable substrate for CoA disulfide reductase, having a Vmax of 2.3 μM/min and a Km of 172 μM. The fact that the Vmax is nearly identical to that of the other CoA disulfides (Table 2) indicates that there is no difference in the rate of cleavage between a cysteine-CoA disulfide and a cysteine-methyl disulfide.
Scheme 9.

Disulfide capping of the active site cysteine in CoA disulfide reductase.
Since the thiol-redox buffer is not itself affected by the N-alkylthio β-lactams, we are now looking into the possibility that the mixed CoA-alkyl disulfides might directly inhibit one or more CoA-dependent enzymes, such as those involved in fatty acid biosynthesis. This coincided with our findings that lactam 1 inhibits the uptake of acetate, and thus fatty acid biosynthesis, in S. aureus.
The demonstrated ability of CoA-mixed disulfides to inhibit bacterial fatty acid synthesis represents a significant advance in the quest for novel antibacterials. However, due to the inability of CoA mixed disulfides to traverse the cell membrane, these CoA disulfides are not directly useful as therapeutics. This research shows that N-alkylthio β-lactams can serve as a prodrug to produce the CoA mixed disulfides within the bacterial target, and that it is through the formation of these mixed disulfides that inhibition of bacterial fatty acid synthesis can be effected.
Finally, it might be informative to compare the properties of these N-thiolated lactams to those of the natural substance allicin, from freshly crushed garlic cloves (Figure 18).36–39 In addition to exhibiting activity against a wide range of bacteria40–43, allicin also shows antifungal44–45, antiviral47,49, antiparisitic49,50, and anticancer properties. Like the N-thiolated lactams, allicin shows the strongest activity against those bacteria that express low levels of glutathione, and blocks fatty acid biosynthesis.51–54 Additionally, both compounds partially inhibit protein and nucleic acid synthesis.55 Allicin is reported to be a specific inhibitor of acetyl-CoA synthetases through reversible and non-covalent binding.56 On the other hand, allicin reacts directly with sulfhydryl residues of various proteins such as alcohol dehydrogenases, thioredoxin reductase, and RNA polymerase, and it is through this covalent modification that it is believed may also inhibit RNA polymerase.57 The similarities in the reactivities and biological properties of allicin (and related compounds such as ajoene58–59, thiolactomycin, and cerulenin) with those of N-thiolated lactams do strongly suggest common biochemical targets and mechanisms of action.
Figure 18.

Structure of allicin, the active constituent from crushed garlic.
Conclusions
Over the past several years, substantial progress has been made toward understanding the structural and mechanistic basis of the observed antibacterial properties of N-alkylthio β-lactams. As described, the metabolism of these compounds proceeds through a thiol-transfer pathway to produce alkyl-CoA mixed disulfides within the cell. Further, it was demonstrated that the bacteria most susceptible (Staphylococcus and Bacillus) are those which utilize CoA in their thiol-redox buffers. Although the high levels of coenzyme A required for the activity of these compounds arises from the role of CoA within the thiol-redox buffer, kinetics studies on the CoA disulfide reductase enzyme provided strong evidence that the CoA thiol-redox buffer itself is not appreciably affected by the N-alkylthio β-lactams. The finding that bacterial fatty acid synthesis is strongly inhibited by the N-alkylthio β-lactams has led to the hypothesis that CoA-dependent enzymes within this pathway may be inhibited by alkyl-CoA mixed disulfides. In vitro experiments in MRSA show that alkyl-CoA mixed disulfides, the ones thought to form from the interaction of CoA and the lactams inside the cell, are in fact biologically inactive against these microbes in culture. This is presumably due to the inability of the disulfides to cross the cell membrane into the cytoplasm. This finding highlights the utility of the N-alkylthio β-lactams as prodrugs to produce alkyl CoA mixed disulfides within the bacterial cells. Furthermore, it was demonstrated that covalent attachment of an alkylthio group to the active site cysteine of CoA disulfide reductase is reversible, due to an NADPH-mediated reduction of the disulfide bond to regenerate the active sulfhydryl form of the enzyme.
While there is strong likelihood that these thiolating compounds produce multiple inhibitory effects on thiol-dependent enzymatic pathways, the finding that coenzyme A is a primary target of the N-thiolated lactams brings further attention to CoA and, specifically, CoA-dependent processes, as a viable target for the development of antibacterial drugs. Within this context, these studies may prove informative, particular in light of the increasing incidences of bacterial drug resistance and the growing need for new antibiotics with alternative mechanisms of action. Although this current study has ruled out CoA disulfide reductase as the primary enzyme affected, other CoA-dependent enzymes which utilize a thiol in its active site may be involved. Our laboratory is now examining this in more detail.
Experimental
General Procedures
Commercially-available reagents and solvents were used without further purification. Lactams 1–5 were prepared and purified as previously reported.1,6
Antimicrobial Susceptibility Testing of Lactams 1–5: Culture Preparation
From a freezer stock in tryptic soy broth (Difco Laboratories, Detroit, MI) and 20% glycerol, a culture of each microorganism was grown on tryptic soy agar (TSA) plates (Becton-Dickinson Laboratories, Cockeysville, MD) at 37°C for 24 hours. A 108 suspension was then made in sterile phosphate buffered saline (pH 7.4) and swabbed across fresh TSA plates. Kirby-Bauer Agar Diffusion Assays: A 108-standardized cell count suspension was made in sterile phosphate buffered saline (pH 7.4) and swabbed across fresh TSA plates. Circular wells (6-mm diameter) were cut into the inoculated plates and 20 μL of a 1 mg/mL stock solution of the test lactam in dimethylsulfoxide (DMSO) was pipeted into the wells. The plates were incubated for 24 hours at 37°C and the antimicrobial susceptibilities were determined by measuring the zones of growth inhibition around each well.
Minimum Inhibitory Concentration (MIC) Assays. Media Preparation
The minimum inhibitory concentrations were determined by agar dilution. The test media were prepared in 24-well plates (Costar 3524, Cambridge, MA) by adding a known concentration of the test drug in DMSO together with a solution of Mueller-Hinton II agar (Becton-Dickinson Laboratories, Cockeysville, MD) for a total volume of 1 mL in each well. Calculations of the overall concentration of antibiotic in the wells were standardized by measuring from a 1 mg/mL stock solution of the test drug. At this concentration, the microliter volume is equivalent to the micrograms in solution. The amount of agar solution added to the wells was determined by subtracting 1000 μL from the quantity of test drug in each well to give a combined volume of 1 mL. Following preparation of the well plates, the media were allowed to solidify at room temperature for 24 hours before inoculation. Inoculation: From an 24-hour culture of each organism on TSA plates (Becton-Dickinson Laboratories, Cockeysville, MD), the staphylococcal strains were grown overnight in 5 mL of tryptic soy broth (Difco Laboratories, Detroit, MI) at 37°C. One microliter of each culture was then applied to the appropriate well of agar and incubated at 37°C overnight. After 24 hr, the MICs were determined by examining the wells for growth. Growth Studies. Overnight cultures of the test strains were grown to logarithmic phase in Mueller-Hinton Broth (MHB). An inoculum of 106 cfu/mL was added to freshly-prepared MHB and grown for 1 hr at 35°C while shaking. The test compounds diluted to 1, 5, or 10 times the MICs in DMSO was applied to each tube. Viable cell counts were determined by plating adequate dilutions of each culture. The plates were incubated and colony counts were taken after 24 hrs. Turbidity measurements were determined by transferring 0.2 mL aliquots of culture to a 48-well plate (Costar 3524, Cambridge, MA) and optical density readings taken at 630 nm with a Bio-Tek EL800 plate reader. Metabolism studies. To a fresh 106 cfu/ml suspension of S. aureus (ATCC 25923) in 9 mL of sterile saline was added 1 mL of a 400 μM solution of lactam 1 in DMSO. After 1 hr, 10 mL of deionized H2O was added and the solution was extracted 3 times with 5 mL of ethyl acetate. The organic layers were combined, dried with magnesium sulfate, and the solvent was removed under reduced pressure. The residue was dissolved in 500 μL of CDCl3 and the chemical structure was elucidated by 1H NMR.
Studies on the Inhibition of CoA Disulfide Reductase by Disulfides and Thiosulfonates
A sample of CoA disulfide reductase was provided by Professor Al Claiborne (Wake Forest University). Coenzyme A (hexahydrate, trilithium salt), coenzyme A disulfide, and methylthio methanesulfonate were obtained from Sigma-Aldrich Co. The remaining thiosulfonate reagents and disulfides were synthesized as described below.
sec-Butylthio Methanethiolsulfonate
To a mixture of sodium methanethiolsulfonate (88 mg, 0.86 mmol, 3.2 eq.) and sec-butyl disulfide (50 μL, 0.27 mmol, 1 eq.) in dry CH2Cl2 (0.5 mL) was added I2 (137 mg, 0.54 mmol, 2 eq.) The mixture was allowed to stir at rt under an inert atmosphere. After 10 hr, TLC indicated no disulfide remaining. The reaction mixture was diluted to 25 mL with CH2Cl2, then 10 mL of Na2S2O3 (1M) was added with stirring until the reddish color disappeared (about 10 minutes). The CH2Cl2 layer was separated, washed with water (2x20 mL), dried over MgSO4, then concentrated in vacuo to give 63 mg (70%) of a pale yellow oil. 1H NMR (250 MHz, CDCl3) ™ 3.50–3.42 (m, 1H); 3.27 (s, 3H); 1.75-1.64 (m, 2H); 1.43 (d, J=7.0 Hz, 3H); 0.98 (t, J=7.4 Hz, 3H). 13C NMR (63 MHz, CDCl3) ™ 51.4, 49.9, 29.9, 21.8, 11.2.
Ethylthio Methanesulfonate
Prepared according to the procedure described above, except the reaction time was 16 h. Obtained 45 mg (60%) as a pale yellow oil. 1H NMR (250 MHz, CDCl3) ™ 3.267 (s, 3H); 3.14 (q, J=7.6 Hz, 2H); 1.39 (t, J=7.4 Hz, 3H). 13C NMR (63 MHz, CDCl3) ™ 50.8, 30.9, 15.0.
Methyl-CoA Disulfide (8a)
To a solution of coenzyme A (5 mg of the trilithium salt, 6.37 μmol) in methanol (300 μL) was added methylthio methanesulfonate (106 μL of a 10 mg/mL methanolic solution, 1.1 mg, 6.4 μmol, 1 eq.). The resulting solution was stirred at rt under N2 for 30 min. The solvent was evaporated under a stream of nitrogen, and the residual solid was taken up in water (1 mL), analyzed by HPLC, and lyophilized. Compound 8a (1.6 mg, 30%) was isolated as a flocculent white solid. 1H NMR: (400 MHz, DMSO-d6): ™ 8.36 (s, 1H); 8.16 (d, J=5.4 Hz, 1H); 8.12 (s, 1H); 7.69 (t, J=6.0 Hz, 1H); 7.23 (broad s, 2H); 5.87 (d, J=6.4 Hz, 2H); 4.64 (t, J=5.2 Hz, 1H); 4.21 (broad s, 1H); 3.94 (broad s, 2H); 3.78 (m, 4H); 2.74 (t, J=7.0 Hz, 2H); 2.38 (s, 2H); 2.37 (s, 3H); 2.56-2.23 (m, 2H); 0.92 (s, 3H); 0.63 (s, 3H). MS (MALDI): m/z 814.550 (M+, all sites protonated).
Ethyl-CoA Disulfide (8b)
Prepared as described for 8a. 4.0 mg (80%) was a obtained as a flocculent white solid. 1H NMR (400 MHz, DMSO-d6): ™ 8.35 (s, 1H); 8.15 (t, J=5.6 Hz, 1H); 8.11 (s, 1H); 7.69 (t, J=5.8 Hz, 1H); 7.22 (broad s, 2H); 5.87 (d, J=6.4 Hz, 1H); 4.77 (broad s, 1H); 4.62 (t, J=5.6 Hz, 1H); 4.19 (broad s, 1H); 3.93 (broad s, 2H); 3.77 (broad s, 4H); 2.72-2.64 (m, 4H); 2.35 (s, 2H); 2.23 (t, J=7.2 Hz, 2H); 1.20 (t, J=7.2 Hz, 3H); 0.92 (s, 3H); 0.62 (s, 3H). MS (MALDI): m/z 828.619 (M+, all sites protonated).
sec-Butyl-CoA Disulfide (8c)
Prepared as described for 8a. 4.9 mg (95%) was isolated as a flocculent white solid. 1H NMR (400 MHz, DMSO-d6): ™ 8.36 (s, 1H); 8.14 (t, J=5.6 Hz, 1H); 8.12 (s, 1H); 7.68 (t, J=5.8 Hz, 1H); 7.23 (broad s, 2H); 5.87 (d, J=6.4 Hz, 1H); 4.79 (broad s, 1H); 4.64 (t, J=5.4 Hz, 1H); 4.21 (broad s, 1H); 3.95 (broad s, 2H); 3.78 (broad s, 4H); 2.90-2.74 (m, 1H); 2.69 (t, J=7.0 Hz, 2H); 2.37 (s, 2H); 2.29-2.23 (m, 2H); 1.60-1.53 (m, 1H); 1.48-1.40 (m, 1H); 1.19 (d, J=6.8 Hz, 3H); 0.92 (s, 3H); 0.88 (t, J=7.4 Hz, 3H); 0.63 (s, 3H). MS (MALDI): m/z 856.592 (M+, all sites protonated).
Co-A Disulfide Reductase Inhibition Assays
Spectrophotometric assays were done using a Perkins-Elmer spectrophotometer equipped with a 6-cuvette changer. Measurements were made at 340 nm and 25oC. Absorbances were measured every 5–10 s for 3 minutes versus a blank consisting of the buffer solution. Using Tris-HCl (50 mM, pH 7.80), stock solutions of CoA disulfide reductase (500 nM), NADPH (100 μM) CoA disulfide (1.0 mM), and the mixed disulfides (1.0 mM) were prepared. CoA disulfide reductase was kept cold during the experiments; all other solutions were allowed to warm to rt.
Determination of CoA Disulfide Reductase Inhibition for Mixed Disulfides 8a-c and CoA Disulfide
To each 1-mL cuvette was added NADPH, CoA disulfide reductase, and pH 7.4 phosphate buffer solution. Addition of the disulfide initiated the kinetic run. Amounts were added volumetrically from stock solutions (described above) such that the following final concentrations were reached: buffer: 50 mM; CoA disulfide reductase: 10 nM; NADPH: 50 μM. Disulfide concentrations were varied as follows:
[CoA disulfide] = 20, 40, 80, 100 μM (Trial 1). 5, 10, 20, 40, 60, 80, 100 μM (Trial 2)
[8a] = 5, 10, 20, 40, 60, 100, 150 μM
[8b] = 2, 10, 20, 40, 60, 100, 150, 200 μM
[8c] = 20, 40, 60, 100, 150 μM
Kinetic parameters were determined by finding the best hyperbolic fit to the Michaelis-Menton equation using Kaleidagraph® software, version 3.5.1.34
Determination of Kinetic Data for Small Disulfides
Run as described above, except that the stock solutions of CoA disulfide reductase were diluted to 100 nM, to minimize pipetting errors and improve reproducibility. The following disulfide concentrations were used:
[8a] = 25, 50, 75, 100 μM
[8b] = 50, 75, 100 μM
[8c] = 13, 25, 38, 50, 63, 75, 100 μM
Measurement of Inhibition Effects for β-Lactams 1 and 2
To each 1 mL-cuvette was added NADPH, CoAD, and lactams 1 or 2 to produce the following final concentrations: buffer: 50 mM. NADPH: 50 μM; CoA disulfide: 20 μM; CoA disulfide reductase: 20 nM. Concentrations of N-alkylthio β-lactam were varied as follows:
[1] = 0, 5, 10, 15, 20, 25, 50, 70 μM
[2] = 0, 10, 15, 20, 25, 50, 70 μM
Addition of the enzyme was done last, to mark the start of the run.
Determination of Inhibition of CoA Disulfide Reductase by Methylthio Methanesulfonate
To each 1 mL cuvette was added NADPH, CoA disulfide reductase, and pH 7.4 phosphate buffer solution. Addition of the methylthio methanesulfonate initiated the kinetic run. Amounts were added volumetrically from stock solutions (described above) such that the following final concentrations were used: buffer: 50 μM; CoA disulfide reductase: 10 nM; NADPH: 50 μM; Methylthio methanesulfonate concentrations were varied as follows: 20, 40, 60, 200, 400, 600, 1000, 2000, 3000 μM.
Scanning Electron Microscopy (SEM) Experiments
Samples were prepared from sections of agar taken from the disk-diffusion experiment. Sterile 6-mm diameter paper discs (Becton-Dickinson Laboratories, Cockeysville, MD) were impregnated with 20 μg of penicillin G potassium salt (Sigma Chemical Co, St. Louis, MO) and lactam 1 from their 1 mg/mL stock solutions in DMSO. From a 24-hour culture of Staphylococcus aureus (ATCC 25923) grown on TSA plates (Becton-Dickinson Laboratories, Cockeysville, MD), a 108 suspension was made in sterile phosphate buffered saline (pH 7.4) and swabbed across two separate plates. The discs were placed on the inoculated plates and incubated for 24 hours at 37° C. After 24 hours, sections of agar in areas containing the division between the zones of inhibition and lawn of bacteria were cut out and placed into Petri dishes for the SEM preparation. The agar sections were flooded with 10 mL of a pre-made glutaraldehyde-osmium fixative. After 1 hour, the sections were removed and washed 3 times with 0.1 M sodium cacodylate. The samples were then sequentially submerged in 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, and 100% ethanol for periods of 5 minute. Following alcohol treatment, the sections were placed into hexamethyldisilazane (HMDS) for chemical drying. The samples were next mounted onto stubs and coated with gold/palladium by a Pelco Model 3 Sputter Coater. The morphology of the cells was examined with a Novascan 30 scanning electron microscope.
Light Microscopy Experiments
Sterile 6-mm paper discs (Becton-Dickinson Laboratories, Cockeysville, MD) containing 20 μg of lactam 1 and penicillin G potassium salt (Sigma Chemical Co, St. Louis, MO) were placed on separate TSA plates inoculated by S. aureus (ATCC 25923) from an overnight culture. The plates were incubated at 37° C for 4–5 hours or until the zones of inhibition were visible. Glass 24 x 60-mm coverslips (Corning Glass works, Corning, NY) were then gently pressed across the zones to adhere the bacteria. Following brief heat fixing over a Bunsen burner flame, the coverslips were flooded by Gram crystal violet stain (Becton Dickinson Laboratories, Cockeysville, MD) for one minute. The coverslips were rinsed with water and flooded by Gram iodine (Difco Laboratories, Detroit, MI) for one minute then decolorized by adding 95% ethanol dropwise until the crystal violet no longer flowed off the coverslips. The coverslips were rinsed again with water and counterstained with Gram safranin (Difco Laboratories, Detroit, MI) for one minute. They were then thoroughly rinsed with water, blotted dry, and mounted on glass microscope slides (Fisher Scientific, Pittsburgh, PA). The slides were viewed with a Nikon bright-field light microscope.
DNA Cleavage Experiments
To 17 μL of sodium phosphate buffer (50 mM, pH 7.4) was added 0.5 μg of pBR322 (ICN Biomedicals Inc, Aurora, OH) in 200 μL microfuge tubes. 2 μL of lactam 1 at 5, 10, 25, 50, and 100 μM concentrations in DMSO were added and the samples were vortexed then incubated at 37°C. After 24 hrs, 2 μL of Blue/Orange 6X Loading Dye (Promega Corp., Madison, WI) was added to 8 μL aliquots of the DNA mixtures. The samples were loaded on a 1.2% agarose gel (1.8 g medium EEO agarose; 150 mL TAE; 1 μg/mL ethidium bromide) and horizontal electrophoresis was performed at 80 V/10 cm for 2.5 hrs in TAE (40 mM Tris-acetate, pH 7.8, 1 mM EDTA). The agarose gel was visualized and photographed under UV transillumination. The same procedure was used in experiments conducted with glutathione, dithiothreitol, and 2-mercaptoethanol. For the enzyme digest samples, 20 μL of sterile deionized H2O, 2 μL of 100 μM 1, 2μL of restriction enzyme buffer H (90 mM Tris-HCl, 10 mM MgCl2, 50 mM NaCl, pH 7.5), 1 μg of pBR322, and 2 μL of EcoR1 were added sequentially to a 1.5 mL microfuge tube and incubated in a 37°C water bath for 1 hr.
Radioisotope-uptake experiments
The effects of lactam 1 on DNA, RNA, and protein synthesis in S. aureus ATCC 25923 was determined by measuring the respective incorporations of [methyl-3H]thymidine, [5-3H]uridine, or L-[4,5-3H]isoleucine (Amersham Life Science). Ciprofloxacin, rifampcin, and chloramphenicol were used as controls for the inhibition of DNA, RNA, and protein synthesis, respectively. Radioactive precursors were added to early logarithmic-phase S. aureus (3 μCi of [methyl-3H]thymidine, 3 μCi of [5-3H]uridine, or 5 μCi of L-[4,5-3H]isoleucine) in Luria Broth in the presence or absence of an antibiotic at twice the MIC value. To assess the effect on DNA, RNA, and protein synthesis, 50 μL samples were removed from each reaction tube at designated time intervals (5, 10, 20, 30, 45, and 60 min) and precipitated in 1 mL of ice cold 10% trichloroacetic acid. After 1 hr, the samples were filtered through glass fiber filters (GF/A; Whatman), washed with 2 mL of ice-cold 5% trichloroacetic acid and 2 mL of ice-cold 95% ethanol, and dried at room temperature overnight. The dried filters were placed in 10 mL vials containing 7 mL of counting fluid (Cytoscint, ICN International). Radioactivity was measured by liquid scintillation (Beckman Instruments).
3H Acetate Uptake Studies
A culture of S. aureus (strain RN4220 in 1% Luria Broth) was used as 1% inoculum in freshly prepared broth (20 mL) and cells were grown at 37oC until the A600 reached 0.11 (1.3 x 107 cfu/mL). The culture was then divided into 4 test tubes (5mL each) and kept in the water bath at 37oC to let the bacteria grow for 5 min, and then sufficient drug was added such that the final concentration of DMSO in each sample was 2% by volume, and each sample contained one of the following: a) penicillin G, (2 ug/mL, 2xMIC); b) lactam 1 (10 μg/mL, 1xMIC); c) lactam 1 (20 μg/mL, 2xMIC); d) no drug added. Each tube was incubated for 5 min. at 37oC, then 2 μL of 3H-acetate (10 μCi/uL in ethanol solution, sodium salt) were added to each tube (4 μCi/mL radioactivity per tube). At intervals of 15, 30, 45, and 60 min, 0.8 mL aliquots were taken from each tube, homogenized, and diluted with chloroform (1 mL) and methanol (2 mL). An additional 1 mL of chloroform and 1 mL of distilled water were added, and the solution was mixed. The organic solution was then washed with distilled water (1 mL), then 2M KCl (3 x 1 mL), then 0.1 M sodium acetate (3 x 1 mL). The radioactivity in the organic phase was then analyzed by scintillation counting.
Procedure for the Isolation of CoA from S. aureus Lysate using Lactam-bound Resin 6
A quantity of 1 liter of S. aureus (ATCC 25923) in broth was cultured at 37 °C for 24 hours. The culture was then centrifuged and washed. The resultant pellet was washed with phosphate buffer and then resuspended in 5 mL of phosphate buffer. The cells were then sonicated in an ice bath for 30 minutes, stopping every 5 minutes to avoid overheating. The lysed cells were then centrifuged and the lysate was centrifuged again, and the resulting lysate was separated off from the solids. Filtering of the lysate yielded a slightly opaque yellow solution. 0.252 g of resin 6 was swelled in 0.5 mL of DMSO and added to the lysate solution. This mixture was then centrifuged at 200 rpm for 24 hours. Next, the lysate was filtered and the solid was repeatedly washed with boiling water and boiling ethanol. The solid was dried and 196 mg of material was collected. The material was dissolved in 5 mL of dry CH2Cl2, and 1 mL of diisobutylaluminum hydride (1.0 M in hexanes) was added at 0oC. The reaction was worked up with 0.1 M HCl followed by freeze-drying. The crude extract was analyzed by high-performance liquid chromatography using a Shimadzu LC-8A HPLC on an analytical reverse phase column. A 9:1 mixture of acetonitrile:water at a 2 mL/min flow rate was used, and detection was done using a Shimadzu SPD-10A UV-VIS detector. Identification of CoA was achieved by comparison of the retention time to a commercial sample.
Acknowledgments
We are grateful for the expert assistance of Ms. Betty Loraam with the electron microscopy experiments, and Ms. Sonja Dickey with culture preparations and gel electrophoresis. We sincerely thank Professor Al Claiborne at Wake Forest Medical School for a sample of co-enzyme A disulfide reductase and also for helpful discussion. We thank the NIH for supporting these studies through grant R01 AI51351.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Turos E, Konaklieva MI, Ren RXF, Shi H, Gonzalez J, Dickey S, Lim D. Tetrahedron. 2000;56:8193. 2000. [Google Scholar]
- 2.Turos E, Long TE, Konaklieva MI, Coates C, Shim JY, Dickey S, Lim DV, Cannons A. Bioorg Med Chem Lett. 2002;12:2229. doi: 10.1016/s0960-894x(02)00343-8. [DOI] [PubMed] [Google Scholar]
- 3.Coates C, Long TE, Turos E, Dickey S, Lim DV. Bioorg Med Chem Lett. 2003;11:193. doi: 10.1016/s0968-0896(02)00383-8. [DOI] [PubMed] [Google Scholar]
- 4.Long TE, Turos E, Konaklieva MI, Blum AL, Amry A, Baker EA, Suwandi LS, McCain MD, Rahman MF, Dickey S, Lim DV. Bioorg Med Chem. 2003;11:1859. doi: 10.1016/s0968-0896(03)00037-3. [DOI] [PubMed] [Google Scholar]
- 5.Turos E, Coates C, Shim JY, Wang Y, Roettgers JM, Long TE, Reddy GSK, Ortiz A, Culbreath M, Dickey S, Lim DV, Alonso E, Gonzalez J. Bioorg Med Chem. 2005;13:6289. doi: 10.1016/j.bmc.2005.08.011. [DOI] [PubMed] [Google Scholar]
- 6.Heldreth B, Long TE, Jang S, Guntireddygari SKR, Turos E, Dickey S, Lim DV. Bioorg Med Chem. 2006;14:3775. doi: 10.1016/j.bmc.2006.01.029. [DOI] [PubMed] [Google Scholar]
- 7.Turos E, Long TE, Heldreth B, Leslie JM, Guntireddygari SKR, Wang Y, Coates C, Konaklieva M, Dickey S, Lim DV, Alonso E, Gonzalez J. Bioorg Med Chem Lett. 2006;16:2084. doi: 10.1016/j.bmcl.2006.01.070. [DOI] [PubMed] [Google Scholar]
- 8.Smith DM, Kazi A, Smith L, Long TE, Heldreth B, Turos E, Dou QP. Mol Pharm. 2002;61:1348. doi: 10.1124/mol.61.6.1348. [DOI] [PubMed] [Google Scholar]
- 9.Kazi A, Hill R, Long TE, Kuhn DJ, Turos E, Dou QP. Biochem Pharm. 2004;67:365. doi: 10.1016/j.bcp.2003.09.017. [DOI] [PubMed] [Google Scholar]
- 10.Kuhn D, Coates C, Daniel K, Chen D, Bhuiyan M, Turos E, Dou QP. Frontiers in Bioscience. 2004;9:2605. doi: 10.2741/1420. [DOI] [PubMed] [Google Scholar]
- 11.Kuhn D, Wang Y, Minic V, Coates C, Reddy GSK, Daniel KG, Shim JY, Chen D, Landis-Piwowar KR, Miller FR, Turos E, Dou QP. Frontiers in Bioscience. 2005;10:1183. doi: 10.2741/1611. [DOI] [PubMed] [Google Scholar]
- 12.Ping Dou Q. Private communication. Wayne State University; [Google Scholar]
- 13.Mishra RK, Revell KD, Coates CM, Dickey S, Lim DV. Bioorg Med Chem Lett. 2006;16:2081. doi: 10.1016/j.bmcl.2006.01.058. [DOI] [PubMed] [Google Scholar]
- 14.Long Timothy. Ph.D. dissertation. University of South Florida; 2003. [Google Scholar]
- 15.Tipper DJ, Strominger JL. Proc Natl Acad Soc USA. 1965;54:1133. doi: 10.1073/pnas.54.4.1133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Waxman DJ, Strominger JL. In: Chemistry and Biology of β-Lactam Antibiotics. Morin RB, Gorman M, editors. Vol. 3. Academic Press; New York: 1982. p. 209. [Google Scholar]
- 17.The advantage of preparing the samples from agar cultures as opposed to broth is that a concentration gradient is produced as the antibiotics diffuse through the solid media, allowing microscopic examination among a range of exposure amounts.
- 18.For a discussion, seeBlaszczak LC, Brown RF, Cook GK, Hornback WJ, Hoying RC, Indelicato JM, Jordon CL, Katner AS, Kinnick MD, McDonald JH, III, Morin JM, Jr, Munroe JE, Pasini CE. J Med Chem. 1990;33:1656. doi: 10.1021/jm00168a019.
- 19.Hara M, Takahashi I, Yoshida M, Asano K, Kawamoto I, Morimoto M, Nakano H. J Antibiot. 1989;42:333. doi: 10.7164/antibiotics.42.333. [DOI] [PubMed] [Google Scholar]
- 20.Hata T, Hoshi T, Kanamori K, Matsumae A, Sano Y, Shima T, Sugawara R. J Antibiot. 1956;9:141. [PubMed] [Google Scholar]
- 21.Hara M, Saitoh Y, Nakano H. Biochemistry. 1990;29:5676. doi: 10.1021/bi00476a005. [DOI] [PubMed] [Google Scholar]
- 22.In addition to potential inhibitory effects on protein and nucleic acid biosynthesis, we also considered the possibility that the lactams may alter bacterial respiration by inhibiting the cellular uptake and phosphorylation of glucose. Thus, we examined the propensity of lactam 1 to induce metabolic starvation in S. aureus by measuring uptake of radiolabeled-glucose in the presence versus the absence of lactam 1. It was found that sugar uptake was not inhibited to any detectible extent at the MIC of lactam 1. Thus, metabolic respiration does not seem to be perturbed by these N-thiolated lactams. For more details, see Bart Heldreth, Ph.D. dissertation, University of South Florida, 2004.
- 23.Leonardi R, Chohnan S, Zhang YM, Virga KG, Lee RL, Rock CO, Jackowski S. J Biol Chem. 2005;280:3314. doi: 10.1074/jbc.M411608200. [DOI] [PubMed] [Google Scholar]
- 24.Slater-Radosti C, Van Aller G, Greenwood R, Nicholar R, Keller PM, DeWolf WE, Jr, Fan F, Payne DJ, Jaworski DD. J Antimicrob Chemother. 2001;48:1. doi: 10.1093/jac/48.1.1. [DOI] [PubMed] [Google Scholar]
- 25.Higgins DL, Chang R, Debabov DV, Leung J, Wu T, Krause KM, Sandvik E, Hubard JM, Kaniga K, Schmidt DE, Jr, Gao Q, Cass RT, Karr DE, Benton BM, Humphrey PP. Antimicrob Agents Chemother. 2005;49:1127. doi: 10.1128/AAC.49.3.1127-1134.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Bligh EG, Dyer WJ. Can J Biochem Phys. 1959;37:911. doi: 10.1139/o59-099. [DOI] [PubMed] [Google Scholar]
- 27.Shah NV, Cama LD. Heterocycles. 1987;25:221. [Google Scholar]
- 28.Woulfe SR, Iwagami H, Miller MJ. Tetrahedron Lett. 1985;26:3891. [Google Scholar]
- 29.Newton GL, Arnold K, Price MS, Sherrill C, delCardayre SB, Aharonowitz Y, Cohen G, Daview J, Fahey RC, Davis C. J Bacteriol. 1996;178:1990. doi: 10.1128/jb.178.7.1990-1995.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.delCardayre SB, Stock KP, Newton GL, Fahey RC, Davies JE. J Biol Chem. 1998;273:5744. doi: 10.1074/jbc.273.10.5744. [DOI] [PubMed] [Google Scholar]
- 31.delCardayre SB, Davies JE. J Biol Chem. 1998;273:5752. doi: 10.1074/jbc.273.10.5752. [DOI] [PubMed] [Google Scholar]
- 32.Argyrou A, Blanchard JS. Progress in Nucleic Acid Research and Molecular Biology. 2004;78:89. doi: 10.1016/S0079-6603(04)78003-4. [DOI] [PubMed] [Google Scholar]
- 33.Fujiki K, Tanifuji N, Sasaki Y, Yokoyama T. Synthesis. 2002;3:343. [Google Scholar]
- 34.Analysis done using Kaleidagraph software, version 3.5.1, available from Synergy Software.
- 35.Purified CoADR was provided by Professor Al Claiborne (Wake Forest Medical School). For more details, see Kevin Revell, Ph.D. dissertation, University of South Florida, 2006.
- 36.Cavallito C, Bailey JH. J Am Chem Soc. 1944;66:1944. [Google Scholar]
- 37.Ankri S, Mirelman D. Microbes and Infection. 1999;2:125. doi: 10.1016/s1286-4579(99)80003-3. [DOI] [PubMed] [Google Scholar]
- 38.Block E. Scient Am. 1985;252:94. doi: 10.1038/scientificamerican0385-114. [DOI] [PubMed] [Google Scholar]
- 39.Koch HP, Lawson LD. Garlic: The Science and Therapeutic Application of Allium sativum L. Vol. 1. Williams and Wilkins; Baltimore: 1996. p. 233. [Google Scholar]
- 40.Uchida Y, Takahashi T, Sato N. Jpn J Antibiotics. 1975;28:638. [PubMed] [Google Scholar]
- 41.Cellini L, Campli E, Masulli M, Di Bartolomeo S, Allocati N. FEMS Immunol Med Microbiol. 1996;13:273. doi: 10.1111/j.1574-695X.1996.tb00251.x. [DOI] [PubMed] [Google Scholar]
- 42.Gonzales-Fandos E, Garcia-Lopez ML, Sierrra ML, Otera A. J Appl Bacteriol. 1994;77:549. doi: 10.1111/j.1365-2672.1994.tb04400.x. [DOI] [PubMed] [Google Scholar]
- 43.Gimenez MA, Solanes RE, Gimenez DF. Rev Argent Microbiol. 1988;20:17. [PubMed] [Google Scholar]
- 44.Davis LE, Shen J, Royer RE. Planta Med. 1994;60:546. doi: 10.1055/s-2006-959568. [DOI] [PubMed] [Google Scholar]
- 45.Hughes BG, Lawson LD. Photother Res. 1991;5:154. [Google Scholar]
- 46.Yamada Y, Azuma K. Antimicrob Agents Chemother. 1997;11:743. doi: 10.1128/aac.11.4.743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Tsai Y, Cole LL, Dav Lockwood SJ, Simmons V, Wild GC. Planta Med. 1985;5:460. doi: 10.1055/s-2007-969553. [DOI] [PubMed] [Google Scholar]
- 48.Tatarintsev AV, Vrzhets PV, Ershov DE, Turgiev AS, Karamov EV, Kornilaeva GV, Makarova TV, Federov NA, Varfolomeev SD. Vestn Ross Akad Med Nauk. 1992;11:6. [PubMed] [Google Scholar]
- 49.Mirelman D, Monheit D, Varon S. J Infect Dis. 1987;156:243. doi: 10.1093/infdis/156.1.243. [DOI] [PubMed] [Google Scholar]
- 50.Ankri S, Miron T, Rabinkov A, Wilchek M, Mirelman D. Antimicrob Agents Chemother. 1997;10:2286. doi: 10.1128/aac.41.10.2286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Adetumbi M, Javor GT, Lau BH. Antimicrob Agents Chemother. 1986;30:499. doi: 10.1128/aac.30.3.499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Ghannoum MA. J Gen Microbiol. 1988;134:2917. doi: 10.1099/00221287-134-11-2917. [DOI] [PubMed] [Google Scholar]
- 53.Neuwirth Z, Sundstrom DC, Thompson NH. Antimicrob Agents Chemother. 1988;32:1763. doi: 10.1128/aac.32.12.1763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Augusti KT, Mathew PT. Experentia. 30:468. doi: 10.1007/BF01926297. [DOI] [PubMed] [Google Scholar]
- 55.Feldberg RS, Chang SC, Kotik AN, Nadler M, Neuwirth Z, Sundstrom DC, Thompson NH. Antimicrob Agents Chemother. 1988;32:1763. doi: 10.1128/aac.32.12.1763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Focke M, Feld A, Lichtenthaler HK. FEBS Lett. 1990;261:106. doi: 10.1016/0014-5793(90)80647-2. [DOI] [PubMed] [Google Scholar]
- 57.Ozolin ON, Uteshev TA, Kim IA, Deev AA, Kamzolova SG. Mol Biol (Mosk) 1990;24:1057. [PubMed] [Google Scholar]
- 58.Urbina JA, Marchan E, Lazardi K, Visbal G, Apitz-Castro R, Gil F, Aguirre T, Piras MM, Piras R. Biochem Pharm. 1993;45:2381. doi: 10.1016/0006-2952(93)90217-k. [DOI] [PubMed] [Google Scholar]
- 59.San-Blas G, Urbina JA, Marchan E, Contreras LM, Sorais F, San-Blas F. Microbiology. 1997;143:1583. doi: 10.1099/00221287-143-5-1583. [DOI] [PubMed] [Google Scholar]