

Abstract
Monocyte chemotactic protein-3 (MCP-3/CCL7) has potent eosinophil chemoattractant properties. The present study determined its relative contribution to the formation of Th2 cytokine-mediated (type-2) eosinophil-rich interstitial lung granulomas induced by antigens of Schistosoma mansoni eggs. Both MCP-3 transcripts and protein levels were more strongly expressed in lungs with type-2 than with type-1 (mycobacterial antigen-elicited Th1-mediated) granulomas. In vivo treatment with neutralizing antibodies demonstrated that MCP-3 abrogated eosinophil accumulation in type-2 lesions by 40 to 50%. Immunohistochemical staining revealed that MCP-3 localized to vessels in or near granulomas suggesting that endothelial cells were an important in situ source of MCP-3. Maximal MCP-3 transcript expression was abrogated by anti-interleukin-4 treatment. Furthermore, cultured mouse lung endothelial cells displayed augmented MCP-3 production in response to interleukin-4. Together, these results suggest that MCP-3 contributes to a significant component of eosinophil recruitment in the type-2 interstitial granuloma formation and Th2 cytokines promote its production.
Because of their participation in hypersensitivity-related diseases, eosinophils and factors that direct their function have been the subject of extensive study. 1 A wide variety of factors can induce eosinophil chemotaxis and mobilization but recently investigative efforts have focused on the role of chemokines. It is recognized that chemokine ligands for C-C chemokine receptor 3 (CCR3) are important eosinophil chemotactic factors because this receptor is strongly expressed by eosinophils. 2-5 Reported ligands for this receptor include RANTES (CCL5), MCP-3 (CCL7), MCP-2 (CCL8), eotaxin-1 (CCL11), and MCP-4 (CCL13). 6,7 We recently demonstrated the induction of several of these chemokines in a murine model of hypersensitivity-type, eosinophil-rich, type-2 pulmonary granuloma formation elicited by Schistosoma mansoni egg Ags. 8 Induction of CCL7, CCL8, and CCL11 ligands occurred during the eosinophil recruitment phase of the lesion and we undertook a systematic analysis to test the hypothesis that these chemokines might contribute to eosinophil recruitment. In a previous report analyzing the contribution of eotaxin-1 (CCL11), we showed that it did not seem to contribute significantly to local eosinophil recruitment within type-2 lung granulomas. 9 In the present study, we similarly tested the potential of MCP-3 (CCL7) as an alternative mediator of eosinophil recruitment in this model. The findings support the notion that MCP-3 is at least one important eosinophil recruitment factor generated during hypersensitivity-type, T-cell-mediated, schistosomal egg Ag-elicited granuloma formation with endothelial cells appearing to be important local sources of this chemokine.
Materials and Methods
Animals
Female, CBA/J mice were obtained from Jackson Laboratories, Bar Harbor, ME. Eotaxin −/− and eotaxin +/+ controls on a B6 × 129 background were kindly provided by Dr. R. Bravo (Department of Oncology and Experimental Pathology, Bristol-Myers Squibb, Princeton, NJ). All mice were maintained under specific pathogen-free conditions and provided food and water ad libitum.
Sensitization and Granuloma Induction
Type-1 and type-2 granulomas were generated as previously described. 10 Briefly, mice were sensitized by subcutaneous injection of 20 μg of purified protein derivative (PPD) (Department of Agriculture, Veterinary Division, Ames, IA) of Mycobacteria bovis incorporated in 0.25 ml of completed Freund’s adjuvant (product no. F-5881; Sigma, St. Louis, MO) or 3000 S. mansoni eggs suspended in 0.5 ml of phosphate-buffered saline (PBS). Fourteen to 16 days later PPD and schistosome egg-sensitized mice were respectively challenged by tail vein injection with 6000 Sepharose 4B beads (in 0.5 ml of PBS) covalently coupled to PPD or to soluble schistosome egg antigens (SEA).
Antibodies and in Vivo Chemokine and Cytokine Depletion Protocol
Polyclonal, anti-MCP-3 antibodies were generated in rabbits by multisite immunization as previously described. 9 Preparations were used when titers reached 1:106 or greater. The anti-MCP-3 antibody (Ab) was highly specific, reacting only with MCP-3 when tested against the following panel of cytokines: eotaxin, MCP-1, MCP-3, MCP-5, MDC, MIP-1α, C10, RANTES, TARC, TCA3, KC, MIP-2, MIG, IP-10, 6Ckine, fractalkine, interleukin (IL)-4, interferon-γ, tumor necrosis factor-α, IL-1β. Neutralizing polyclonal anti-eotaxin and monoclonal anti-IL-4 antibodies were previously described. 9,11 Briefly, anti-murine eotaxin was prepared by multiple site immunization of rabbits with recombinant murine eotaxin. The antibody titer by direct enzyme-linked immunosorbent assay (ELISA) was 1:106 and other than eotaxin did not react with a panel of chemokines and cytokines as described above. In an in vitro chemotaxis assay, 150 μl of a 1:200 dilution of anti-eotaxin neutralized 300 ng of recombinant mouse eotaxin. Rat mAb with specificity for murine IL-4 was obtained from of the hybridoma line 11B11 (American Type Culture Collection, Rockville, MD) as previously described. 9 11B11 is of IgG isotype and has known neutralizing activity.
For in vivo neutralization, mice were given an intraperitoneal injection of 10 mg of purified rabbit anti-murine eotaxin or anti-murine MCP-3 IgG in 1 ml of PBS at the time of bead challenge. Nonimmune rabbit IgG served as a control. For IL-4 depletion, 2 mg of rat anti-murine IL-4 antibodies were administered intraperitoneally in 1 ml of PBS. Nonimmune rat IgG served as a control. Four days after challenge, lungs were excised. One lobe was analyzed for chemokine transcript expression by real-time polymerase chain reaction (PCR) whereas remaining lobes were used for granuloma morphometry or dispersal for differential analysis.
Preparation of mRNA and Protein Extracts from Lungs
Four days after bead challenge, mice were euthanized and exsanguinated before lung harvest. After perfusion with cold RPMI, lungs, excluding the trachea and major bronchi, were excised. The left lower lung lobe of each mouse was postinflated with buffered formalin. The right lobe was snap-frozen in liquid nitrogen for immunohistochemical staining or mRNA isolation, and remaining lobes were used for protein extraction or granuloma dispersal. mRNA was isolated from frozen tissues using Poly(A) pure mRNA isolation kits (Ambion, Austin, TX). Purity of mRNA was confirmed by failure to detect gene PCR amplification in nonreverse-transcribed samples up to 37 amplification cycles.
For protein extraction, snap-frozen lung lobes were suspended in 2 ml of PBS and homogenized for 20 seconds using a Tissue Tearor (Biospec Products, Inc., Bartlesville, OK). Next, 0.1 ml of fetal bovine serum was added as a protein stabilizer. The homogenate was centrifuged at 300 × g for 20 minutes and then the supernate was collected, aliquoted, and frozen at −80°C before ELISA.
Chemokines were measured in the whole lung aqueous extracts by specific ELISA using commercial reagents (R&D Systems, Minneapolis, MN) as previously described. 8 Next, total lung protein concentration was determined in experimental and control samples, then chemokine levels were normalized to mg of lung protein after subtraction of fetal bovine serum protein component.
Differential and Morphometric Analysis of Granuloma Cell Composition
Dispersed granulomas were prepared as previously described. 10,12 A 200-cell differential analysis was performed on duplicate Wright-stained cytospin preparations of dispersed granulomas. As a further measure of eosinophil mobilization, direct morphometric analysis of eosinophils was performed on digitized images of five granuloma histological cross sections from each of the treated mice. A blinded observer counted six 50-square μm fields from each granuloma.
Quantitation of Chemokine mRNA Expression in Lungs
Approximately 1 μg of mRNA was reverse-transcribed in a 20-μl reaction in a PCR reaction tube using Reverse Transcription System kits (Promega, Madison, WI). Five reactions were conducted to minimize variability between tubes. The products from each tube were pooled and subjected to real-time PCR using a TaqMan 7000 light cycler (Applied Biosystems, Foster City, CA.). For the current study the manufacturer’s method (see User Bulletin no. 2, Perkin-Elmer) was adopted with GAPDH as endogenous reference. The generating PCR products of the target gene and GAPDH were monitored simultaneously in real-time in the reaction with a fluorescence amplification factor measured for each gene relative to GAPDH. Data are expressed as arbitrary units (AU), which are calculated from the fluorescence amplication factor as measured by the real-time PCR fluorescent detection unit. The original gene copy number (Co) is related to fluorescence of the generated signal as follows: Co = F × E−1 × I × 2−n, where F is an arbitrary conversion constant, E−1 is amplification efficiency constant (approximately = 1 for manufacturer’s real-time primers sets), I is the fluorescent intensity reading, and n is the amplification cycle number. Hence, F × E−1 × I × 2−n constitutes an arbitrary measure of originally copy number that is directly related to the fluorescent product and inversely related to cycle number.
For MCP-3 gene analysis, TaqMan predeveloped reaction kits (Perkin-Elmer) were used. Oligonucleotides for PCR primers were obtained from Operon Technologies (Alameda, CA.) and TaqMan probes were purchased from Applied Biosystems. In all cases, TaqMan Universal PCR Master Mix (Perkin-Elmer) was used, and the thermal cycling condition was programmed according to the manufacturer’s instructions. Before being used in the actual mRNA expression analysis, each primer-probe set was pretested with an undiluted positive control sample and at 1:4 and 1:16 dilutions. Water served as a negative control.
Immunohistochemistry
Five- to 7-μm-thick frozen tissue sections were mounted on poly-l-lysine-coated slides, fixed with acetone, then rehydrated in PBS. Sections were preblocked with avidin, biotin, and 2% fetal bovine serum. The sections were covered with 10 μg/ml of goat anti-mouse eotaxin or anti-mouse MCP-3 IgG (R&D Systems). Nonimmune goat IgG served as a control. Slides were incubated for 30 minutes at 37°C, then rinsed and overlaid with biotinylated anti-goat IgG (supersensitive reagent 1:20; Biogenex, San Ramon, CA). After a further 30-minute incubation at 37°C, the slides were rinsed, then overlaid with streptavidin-alkaline-phosphatase (1:20, Biogenex,). The slides were rinsed again in PBS and overlaid with alkaline-phosphatase substrate (Fast red) solution (Biogenex) for 30 minutes at room temperature. Finally, the sections were rinsed, counterstained with Gill’s hematoxylin, and mounted with coverslips.
Lung Endothelial Culture and Stimulation
Mouse lung endothelial cells were isolated and cultured as described by Murphy and colleagues. 13 Briefly, strips of peripheral lungs from 4- to 5-week-old mice were removed, minced, and incubated in gelatin-coated tissue culture flasks in growth media consisting of RPMI-1640, supplemented with 20% fetal calf serum, penicillin-streptomycin, and endothelial cell growth supplement. After 65 hours, tissues were removed leaving cell monolayers that consisted uniformly of endothelial cells. Cells were used at 80 to 90% confluence at passage 1 for all experimental studies. Experiments described below were repeated on several separate isolates of endothelial cells.
Cells were characterized as endothelial cells by their uniform cobblestone morphology and by the flow cytometric determination of a single population of cells with uniform uptake of fluorescently-labeled acetylated low-density lipoprotein (1,1′-diotadecyl-3,3,3′,3′-tetramethyl-indo-carbocyanine perchlorated) (DiI-Ac-LDL) (Biomedical Technologies Inc., Stoughton, MA). Human umbilical vein endothelial cells served as positive controls and dermal keratinocytes served as negative controls for low-density lipoprotein uptake studies. Cells demonstrated significant angiotensin-converting enzyme activity when compared to human serum with known angiotensin-converting enzyme activity using a spectrophotometric method using the hydrolysis of the synthetic peptide FAPGG by angiotensin-converting enzyme (Sigma Chemical Co.).
The lung endothelial cells were cultured in 24-well plates and were stimulated with graded doses of recombinant cytokine. Triplicate wells were prepared for each dose in each experiment the experiment was repeated with fresh endothelial cells.
Statistics
The Student’s t-test (two-tailed) was used to compare control with experimental groups. Values of P < 0.05 were considered to indicate significance.
Results
MCP-3 Expression Is Enhanced in Lungs with Eosinophil-Rich, Type-2 Pulmonary Granulomas
Initially, we compared MCP-3 protein and transcript levels in lungs with Th1-mediated type-1- (PPD) and Th2-mediated type-2 (SEA) bead granulomas. Type 1 lesions contain only a minor component of eosinophils, whereas the type-2 lesions are comprised of 20 to 30% eosinophils. As shown in Figure 1 ▶ , although not exclusive to the type-2 response, both MCP-3 transcripts and protein levels were higher in the lungs of CBA mice with type-2 as compared to those with type-1 bead granulomas.
Figure 1.

Enhanced MCP-3 (CCL7) expression during pulmonary, type-2 (SEA) bead granuloma formation. A: Relative component of eosinophils in dispersed type-1 (PPD) and type-2 (SEA) bead granulomas. Bars are means ± SD of three to four separate experiments. B: MCP-3 protein and mRNA expression in granulomatous lungs. 0 = prechallenge lung. Bars are means ± SD from two experiments with five mice per point in each experiment.
MCP-3 Contributes to Eosinophil Recruitment in Type-2 Pulmonary Granulomas
To assess the potential contribution of MCP-3 to eosinophil recruitment, mice were administered Abs to MCP-3 at the time of type-2 granuloma elicitation. As additional controls, anti-eotaxin Abs either alone or in combination with anti-MCP-3 were administered to parallel groups of animals. At 4 days of granuloma development, lesion composition was assessed by dispersal and morphometric analysis. As shown in Table 1 ▶ and as we previously reported, in vivo eotaxin depletion did not significantly reduce eosinophil accumulation in type-2 lesions. In contrast, depletion of MCP-3 selectively reduced eosinophil recruitment in type-2 lesions (by 40 to 50% in three separate experiments) with no further reduction of eosinophils observed by combining anti-eotaxin and anti-MCP-3 treatment. In addition, we similarly tested eotaxin-1 knockout mice, which showed no reduction of eosinophils unless treated with anti-MCP-3 antibodies (Table 2) ▶ .
Table 1.
Effect of in Vivo Chemokine Depletion on Cell Composition of Dispersed Type-2 Granulomas in Wild-Type CBA Mice
| Treatment | Lymphocytes | Macrophages | Eosinophils | Neutrophils |
|---|---|---|---|---|
| Control IgG | 45.0 ± 4.2 | 30.0 ± 1.4 | 21.0 ± 4.2 | 3.5 ± 0.7 |
| Anti-ETX | 41.0 ± 7.9 | 36.3 ± 2.5 | 19.0 ± 5.2 | 3.6 ± 1.5 |
| Anti-MCP-3 | 54.5 ± 3.5 | 29.0 ± 1.4 | 12.0 ± 4.2* | 3.5 ± 2.1 |
| Anti-ETX+ anti-MCP-3 | 48.5 ± 2.1 | 34.5 ± 4.9 | 12.0 ± 1.4* | 4.5 ± 0.7 |
Values are mean ± SD of a representative experiment of three separate experiments with similar results; six to eight mice per group. Mice with type-2 pulmonary granuloma formation were treated with 10 mg of anti-chemokine antibodies or control IgG. On day 4, pulmonary granulomas were isolated, dispersed, and duplicate cytospin preparations were subjected to a 200-cell differential.
*P < 0.05 compared with control IgG group.
Table 2.
Effect of Eotaxin Knockout and MCP-3 Depletion on Cell Composition of Dispersed Type-2 Granulomas
| Mouse strain | Treatment | Lymphocytes | Macrophages | Eosinophils | Neutrophils |
|---|---|---|---|---|---|
| Eotaxin +/+ | None | 53.0 ± 5.6 | 23.0 ± 2.1 | 20.5 ± 0.8 | 3.5 ± 2.1 |
| Eotaxin−/− | None | 49.5 ± 0.7 | 22.5 ± 0.7 | 27.0 ± 1.4 | 1.5 ± 0.8 |
| Eotaxin−/− | IgG | 43.0 ± 4.0 | 27.0 ± 1.4 | 25.0 ± 2.0 | 6.0 ± 2.1 |
| Eotaxin−/− | Anti-MCP-3 | 53.0 ± 2.1 | 24.0 ± 1.4 | 15.0 ± 2.0* | 5.0 ± 2.9 |
Values are mean ± SD. Type-2 pulmonary granulomas were induced in wild-type (eotaxin +/+) and eotaxin knockout mice (eotaxin −/−). Additional knockout groups were treated with 10 mg of anti-chemokine antibodies or control IgG at the time of bead challenge. On day 4, pulmonary granulomas were isolated, dispersed, and duplicate cytospin preparations were subjected to a 200-cell differential. Five to six mice per group.
*P < 0.05 compared with control IgG group.
Figure 2 ▶ shows the histological appearance of type-2 granulomas and the average eosinophil density for control and treated groups as determined by morphometric analysis. Similar to the dispersal analysis, anti-MCP-3 antibodies abrogated eosinophil accumulation in type-2 granulomas and again eotaxin depletion had no statistically significant effect.
Figure 2.
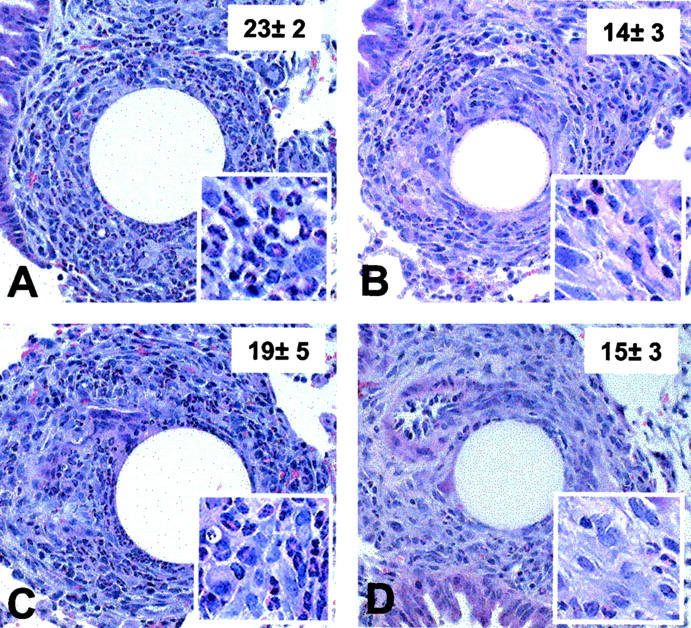
Histological appearance of type-2 (SEA) bead granulomas in mice treated with antibodies to eotaxin and MCP-3. Type-2 (SEA) lesions were generated in presensitized CBA mice, then at the time of bead challenge they were administered 10 mg of purified IgG (nonimmune rabbit, anti-eotaxin, anti-MCP-3, or 10 mg each of anti-eotaxin plus anti-MCP-3 Abs). On day 4 after Ab treatment and bead challenge, lung tissues were harvested and histological sections were prepared as described in Materials and Methods. A, Nonimmune IgG treated; B, anti-MCP-3 treated; C, anti-eotaxin treated; D, combined anti-MCP-3 and anti-eotaxin treatment. Granulomas from each of the five to six mice per treatment group were analyzed morphometrically and the inset numbers indicate mean eosinophil counts per 50 × 50 μm area ± SD. Four such areas were measured from each granuloma. Original magnifications: ×200 (A to D); ×400 (insets).
In Vivo Treatment with IL-4 Antibodies Abrogates MCP-3 Transcript Expression
We and others have previously reported that IL-4 is an important mediator of schistosomal antigen-elicited granuloma formation. 11,14,15 We also previously reported reduced MCP-3 transcripts and eosinophil recruitment in IL-4 knockout mice. 10,12 To further detect a possible link between IL-4 and MCP-3, we administered anti-IL-4 antibodies to wild-type mice and assessed effects on MCP-3 transcript expression. As shown in Figure 3 ▶ , anti-IL-4 profoundly reduced levels of MCP-3 transcripts, which correlated with a significant reduction of eosinophils within lesions. These findings indicated that IL-4 was either directly or indirectly promoting MCP-3 production in vivo.
Figure 3.

In vivo IL-4 neutralization abrogates MCP-3 (CCL7) transcript expression and eosinophil recruitment in lungs with type-2 (SEA) bead granuloma formation. Just before bead challenge mice were treated with anti-cytokine Abs as described in the Materials and Methods, then 4 days later lungs were harvested for analysis. A: MCP-3 (CCL7) transcript expression. Bars are means ± SD of three experiments. B: Relative component of eosinophils in isolated, dispersed granulomas. Bars are means ± SE derived from two experiments with a total of six to eight mice per group.
MCP-3 Displays a Vascular Localization Pattern in Lungs with Type-2 Granuloma Formation
The above studies indicated that MCP-3 contributes to eosinophil recruitment during type-2 cytokine-mediated granuloma formation. Therefore, it was of interest to determine the physical relationship of chemokine expression to the inflammatory lesion. To this end, we performed immunohistochemical staining for MCP-3, which was consistently associated with the luminal side of small vessels in or adjacent to type-2 granulomas (Figure 4 ▶ ; A, C, and E). Inflammatory cells were often found marginating and transmigrating these vessels, which included both terminal pulmonary arterioles and venules. Staining was not observed in normal lungs but was minimally detected in lungs challenged with PPD or non-Ag beads (Figure 4, G and H) ▶ . Staining was also performed for eotaxin, another potential eosinophil chemotactic agent, and a different staining pattern was identified. Eotaxin was not observed on endothelia but was expressed in bronchoepithelial cells and focally in alveolar septal interstitial cells (Figure 5, A and C) ▶ . The distribution of the latter was suggestive of interstitial macrophages. Surprisingly, eotaxin was also not identified in granulomas (Figure 5E) ▶ . Interestingly, eotaxin was found in occasional bronchoepithelial cells within normal lungs and lungs challenged with PPD or non-Ag coated beads (Figure 5F) ▶ , albeit to a lesser extent. The latter was fully consistent with reports of baseline constitutive eotaxin expression in mouse lungs. 16,17 These patterns of expression seemed to provide an explanation for the different contribution of eotaxin and MCP-3 to local eosinophil recruitment to type-2 granulomas noted in our chemokine depletion studies. Specifically, eotaxin was generally remote from granulomas, whereas MCP-3 was associated with vessels within and immediately adjacent to lesions.
Figure 4.
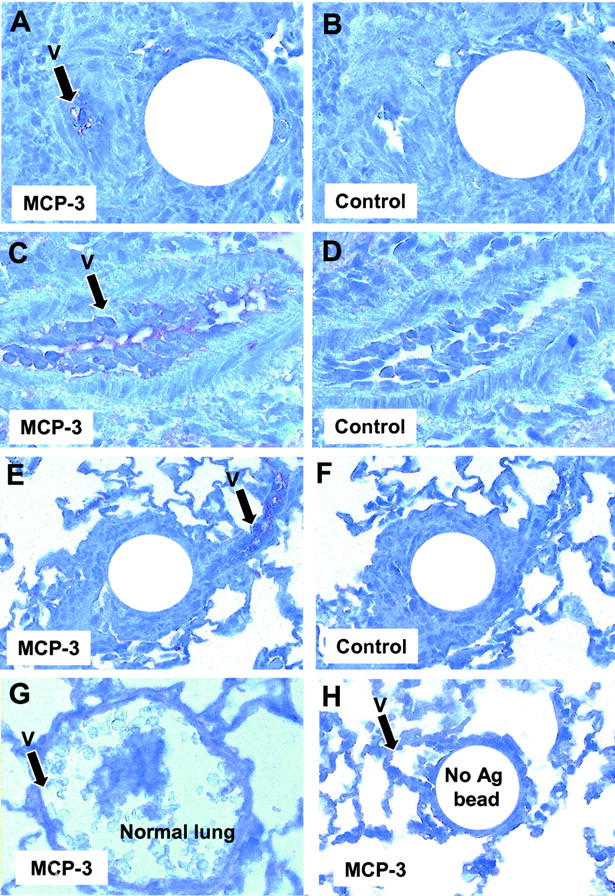
Immunohistochemical localization of MCP-3. Frozen sections of lungs were prepared and stained with anti-MCP-3 Abs as described in Materials and Methods. Note staining of vessels (V) in A, C, and E. A: Type-2 (SEA) granulomas, MCP-3 stain. B: Serial section of A, nonimmune Ab control. C: Vessel in proximity to type-2 (SEA) granuloma, MCP-3 stain. D: Serial section of C, nonimmune Ab control. E: Type-2 (SEA) bead granuloma, MCP-3 stain, low-power magnification. F: Serial section of E, nonimmune Ab control. G: Parenchyma and vessel in normal lung, MCP-3 stain. H: Control bead lesion in lung challenged with antigen-free beads, low magnification. Original magnifications: ×200 (A, G);×400 (C); ×100 (D, E, H).
Figure 5.

Immunohistochemical localization of eotaxin. Frozen sections of lungs were prepared and stained with anti-eotaxin Abs as described in Materials and Methods. A: Bronchus from lung with type-2 (SEA) granulomas, eotaxin stain. B: Serial section of A, nonimmune Ab control. C: Lung parenchymal alveoli in lung with type-2 (SEA) granulomas, eotaxin stain. D: Lung parenchymal alveoli in lung with type-2 (SEA) granulomas, nonimmune Ab control; E, Type-2 (SEA) bead granuloma, eotaxin stain. F: Bronchus and nearby control bead lesion in lung challenged with antigen-free beads. Original magnifications: ×200 (A, C, F); ×400 (D, E).
Mouse Lung Endothelial Cells Express MCP-3 in Response to Type-2 Cytokines
Our immunohistochemical studies suggested that vascular endothelial cells were potential sources of MCP-3. Human endothelial cells have been reported to produce MCP-3 in response to Th2 cytokines and are capable of binding chemokines to their surfaces by promiscuous membrane-binding receptors. 18,19 To determine whether such a cytokine-chemokine network could occur in the mouse lung, we isolated mouse lung endothelial cells and tested their capacity to produce MCP-3 and eotaxin in response to graded doses of IL-4. As shown in Figure 6 ▶ , IL-4 induced MCP-3 production by mouse pulmonary endothelial cells in a dose-response manner but did not elicit significant amounts of eotaxin.
Figure 6.

Interleukin-4 induced MCP-3 (CCL7) production by lung endothelial cells. Lung endothelial cells were cultured with graded doses of IL-4 as described in the Materials and Methods and then 24 hours supernates were assayed for MCP-3 and eotaxin by ELISA. Bars are means ± SD of triplicate cultures.
Discussion
This study represents the first analysis of the role of MCP-3 (CCL7) in eosinophil recruitment during schistosomal Ag-elicited granuloma formation. MCP-3 is an agonist for the CCR3 chemokine receptor, a functional receptor expressed by eosinophils. 2,3,5,20 MCP-3 is a promiscuous chemokine and reportedly binds to other receptors such as CCR1 and CCR2. 21 MCP-3 is expressed by the mucosa of asthmatics and is associated with significantly increased numbers of bronchial mucosal eosinophils in these patients. 22-24 Likewise, MCP-3 is implicated in the pathogenesis of ulcerative colitis. 25 In addition, Stafford and colleagues 26 directly demonstrated a role for MCP-3 in eosinophil recruitment in a model of experimental asthma and provided circumstantial evidence for IL-4-induced expression. In view of these data, we directly tested the contribution of MCP-3 to the eosinophil rich type-2 hypersensitivity lung granulomatous response induced by bead-immobilized schistosomal egg Ags and evaluated its relationship to IL-4.
We previously observed that Ab-mediated eotaxin (CCL11) depletion did not seem to significantly reduce local eosinophil recruitment to type-2 hypersensitivity lung granulomas despite eotaxin induction within the whole lung. 9 That study suggested that other mechanisms were involved in eosinophil recruitment and indeed the present study demonstrated that MCP-3 is an important alternative mediator. Moreover, combining eotaxin and MCP-3 depletion did not result in significant further reduction of eosinophils, indicating that MCP-3 was the more significant contributor. It should be noted that eosinophil recruitment into type-2 lung granulomas was likely not mediated solely by MCP-3, because in vivo MCP-3 depletion failed to completely eliminate eosinophils from the lesions. Many eosinophil chemotactic activities have been described. In fact, eotaxin-2 has been recently demonstrated in the mouse and this isoform may participate where eotaxin-1 may not. 27 It should also be noted that although MCP-3 was enhanced in the type-2 response it probably cannot dictate selective eosinophil accumulation by itself. The degree of eosinophil infiltration likely depends as much on eosinophilopoietic and activating factors such as IL-5, which is also expressed to a greater degree in the type-2 response.
Our finding that MCP-3, a CCR3 agonist, contributes strongly to eosinophil recruitment fully agrees with observations that CCR3 is an important functional chemotactic receptor expressed by eosinophils. 3,5,20 Although a direct eosinophil recruitment effect of MCP-3 is the simplest interpretation of our findings, alternative indirect effects of MCP-3 depletion must also be considered. Type-2 helper T (Th2) clones reportedly express CCR3 28-31 and MCP-3 depletion could conceivably impair local type-2 cytokine production by reducing CD4+ Th2-derived migration to inflammatory sites. 32 However, this possibility is unlikely as more recent evidence indicates that CCR4 is in fact the more dominant Th2 cell-associated receptor and hence other chemokines such as monocyte-derived chemokine (MDC/CCL22 and thymus activation-related chemokine (TARC/CCL17) are likely more important in Th2 cell recruitment. 33-37 Indeed, we have demonstrated both of these chemokines to be produced during type-2 bead granuloma formation. 8
Our immunohistochemical studies indicated that neither eosinophils nor other granuloma leukocytes stained for MCP-3 but rather pointed to endothelial cells as potential sources. In addition, the staining patterns provided an explanation for the different contribution of eotaxin and MCP-3 to eosinophil recruitment by revealing that MCP-3 but not eotaxin was present within granuloma-associated vessels. MCP-3 staining has been reported in mucosal epithelial cells and macrophages in biopsies of patients with allergic rhinitis. 38 In the present study we found MCP-3 associated with endothelium of small vessels in or adjacent to type-2 granulomas, which is fully consistent with reports that endothelial cells synthesize MCP-3 in response to Th2-associated cytokines. 18 In addition, our results may have relevance to reports that MCP-3 and other chemokines appear to coat the luminal surface of endothelial cells by chemokine receptor-independent interactions, which potentially allows for adherence by classic chemokine receptor-dependent interactions with circulating leukocytes. 19 In support of this notion, eosinophil adherence to endothelial cells via the CCR3 receptor has been directly demonstrated in vitro. 39 Thus, our findings would support the hypothesis that at least some chemokines contribute to recruitment by promoting leukocyte adherence.
Finally, as demonstrated for human endothelial cells, 18 we showed that the type-2 cytokine IL-4 directly stimulated MCP-3 production by primary mouse lung endothelial cell cultures, providing an in vitro correlate to our in vivo cytokine depletion and immunolocalization studies. Also in support of our immunolocalization studies was the finding that these cells did not produce eotaxin and we have also been unable to elicit eotaxin synthesis in mouse lung endothelia with type-1-associated cytokines, interferon-γ, and tumor necrosis factor-α (data not shown). The failure of these cells to produce eotaxin was unexpected because interferon-γ reportedly induced eotaxin in murine endothelial cells, 40 however those studies were performed using immortalized, virally transformed endothelial cells and not primary cell cultures, possibly explaining the observed differences. In addition, endothelial cells can display functional differences based on their organ site. 13
In summary, our findings indicate that murine MCP-3 contributes to eosinophil accumulation during type-2 hypersensitivity-type pulmonary granuloma formation, endothelial cells likely represent a major source, and its production in this model is promoted by IL-4. These findings potentially are relevant to eosinophilic conditions such as hypersensitivity pneumonitis and asthma. Moreover, they further emphasize the apparent complexity and redundancy of leukocyte recruitment mechanisms.
Acknowledgments
We thank Pamela Lincoln for her expert technical assistance and Aron Pollack and Stacey Haller for their histological skills.
Footnotes
Address reprint requests to Stephen W. Chensue, M.D., Ph.D., Pathology and Laboratory Medicine 113, Veterans Affairs Medical Center, 2215 Fuller Rd., Ann Arbor, MI 48105. E-mail: schensue@med.umich.edu.
Supported by the Department of Veterans Affairs and the National Institutes of Health-National Institute of Allergy and Infectious Diseases (grant AI43460). Schistosome life stages or materials for this work were supplied through National Institutes of Health-National Institute of Allergy and Infectious Diseases contract NO1-AI-55270.
References
- 1.Wardlaw AJ, Brightling C, Green R, Woltmann G, Pavord I: Eosinophils in asthma and other allergic diseases. Br Med Bull 2000, 56:985-1003 [DOI] [PubMed] [Google Scholar]
- 2.Gao JL, Sen AI, Kitaura M, Yoshie O, Rothenberg ME, Murphy PM, Luster AD: Identification of a mouse eosinophil receptor for the CC chemokine eotaxin. Biochem Biophys Res Commun 1996, 223:679-684 [DOI] [PubMed] [Google Scholar]
- 3.Heath H, Qin S, Rao P, Wu L, LaRosa G, Kassam N, Ponath PD, Mackay CR: Chemokine receptor usage by human eosinophils. The importance of CCR3 demonstrated using an antagonistic monoclonal antibody. J Clin Invest 1997, 99:178-184 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.White JR, Imburgia C, Dul E, Appelbaum E, O’Donnell K, O’Shannessy DJ, Brawner M, Fornwald J, Adamou J, Elshourbagy NA, Kaiser K, Foley JJ, Schmidt DB, Johanson K, Macphee C, Moores K, McNulty D, Scott GF, Schleimer RP, Sarau HM: Cloning and functional characterization of a novel human CC chemokine that binds to the CCR3 receptor and activates human eosinophils. J Leukoc Biol 1997, 62:667-675 [DOI] [PubMed] [Google Scholar]
- 5.Grimaldi JC, Yu NX, Grunig G, Seymour BWP, Cottrez F, Robinson DS, Hosken N, Ferlin WG, Wu XY, Soto H, O’Garra A, Howard MC, Coffman RL: Depletion of eosinophils in mice through the use of antibodies specific for C-C chemokine receptor 3 (CCR3). J Leukoc Biol 1999, 65:846-853 [DOI] [PubMed] [Google Scholar]
- 6.Zlotnik A, Morales J, Hedrick JA: Recent advances in chemokines and chemokine receptors. Crit Rev Immunol 1999, 19:1-47 [PubMed] [Google Scholar]
- 7.Zlotnik A, Yoshie O: Chemokines: a new classification system and their role in immunity. Immunity 2000, 12:121-127 [DOI] [PubMed] [Google Scholar]
- 8.Qiu B, Frait KA, Reich F, Komuniecki E, Chensue SW: Chemokine expression dynamics in mycobacterial (type-1) and schistosomal (type-2) antigen-elicited pulmonary granuloma formation. Am J Pathol 2001, 158:1503-1515 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ruth JH, Lukacs NW, Warmington KS, Polak TJ, Burdick M, Kunkel SL, Strieter RM, Chensue SW: Expression and participation of eotaxin during mycobacterial (type 1) and schistosomal (type 2) antigen-elicited granuloma formation. J Immunol 1998, 161:4276-4282 [PubMed] [Google Scholar]
- 10.Chensue SW, Warmington K, Ruth JH, Lukacs N, Kunkel SL: Mycobacterial and schistosomal antigen-elicited granuloma formation in IFN-gamma and IL-4 knockout mice: analysis of local and regional cytokine and chemokine networks [published erratum appears in J Immunol 1999, 162:3106]. J Immunol 1997, 159:3565-3573 [PubMed] [Google Scholar]
- 11.Ruth JH, Warmington KS, Shang X, Lincoln P, Evanoff H, Kunkel SL, Chensue SW: Interleukin-4 and -13 participation in mycobacterial (type-1) and schistosomal (type-2) antigen-elicited pulmonary granuloma formation: multiparameter analysis of cellular recruitment, chemokine expression and cytokine networks. Cytokine 2000, 12:432-444 [DOI] [PubMed] [Google Scholar]
- 12.Chensue SW, Warmington KS, Ruth JH, Lincoln P, Kunkel SL: Cytokine function during mycobacterial and schistosomal antigen-induced pulmonary granuloma formation. Local and regional participation of IFN-gamma, IL-10, and TNF. J Immunol 1995, 154:5969-5976 [PubMed] [Google Scholar]
- 13.Murphy HS, Bakopoulos N, Dame MK, Varani J, Ward PA: Heterogeneity of vascular endothelial cells: differences in susceptibility to neutrophil-mediated injury. Microvasc Res 1998, 56:203-211 [DOI] [PubMed] [Google Scholar]
- 14.Chensue SW, Terebuh PD, Warmington KS, Hershey SD, Evanoff HL, Kunkel SL, Higashi GI: Role of IL-4 and IFN-gamma in Schistosoma mansoni egg-induced hypersensitivity granuloma formation. Orchestration, relative contribution, and relationship to macrophage function. J Immunol 1992, 148:900-906 [PubMed] [Google Scholar]
- 15.Cheever AW, Williams ME, Wynn TA, Finkelman FD, Seder RA, Cox TM, Hieny S, Caspar P, Sher A: Anti-IL-4 treatment of Schistosoma mansoni-infected mice inhibits development of T cells and non-B, non-T cells expressing Th2 cytokines while decreasing egg-induced hepatic fibrosis. J Immunol 1994, 153:753-759 [PubMed] [Google Scholar]
- 16.Rothenberg ME, MacLean JA, Pearlman E, Luster AD, Leder P: Targeted disruption of the chemokine eotaxin partially reduces antigen-induced tissue eosinophilia. J Exp Med 1997, 185:785-790 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Rothenberg ME, Zimmermann N, Mishra A, Brandt E, Birkenberger LA, Hogan SP, Foster PS: Chemokines and chemokine receptors: their role in allergic airway disease. J Clin Immunol 1999, 19:250-265 [DOI] [PubMed] [Google Scholar]
- 18.Polentarutti N, Introna M, Sozzani S, Mancinelli R, Mantovani G, Mantovani A: Expression of monocyte chemotactic protein-3 in human monocytes and endothelial cells. Eur Cytokine Netw 1997, 8:271-274 [PubMed] [Google Scholar]
- 19.Hub E, Rot A: Binding of RANTES, MCP-1, MCP-3, and MIP-1alpha to cells in human skin. Am J Pathol 1998, 152:749-757 [PMC free article] [PubMed] [Google Scholar]
- 20.Fujisawa T, Kato Y, Nagase H, Atsuta J, Terada A, Iguchi K, Kamiya H, Morita Y, Kitaura M, Kawasaki H, Yoshie O, Hirai K: Chemokines induce eosinophil degranulation through CCR-3. J Allergy Clin Immunol 2000, 106:507-513 [DOI] [PubMed] [Google Scholar]
- 21.Baggiolini M, Loetscher P: Chemokines in inflammation and immunity. Immunol Today 2000, 21:418-420 [DOI] [PubMed] [Google Scholar]
- 22.Ying S, Meng Q, Zeibecoglou K, Robinson DS, Macfarlane A, Humbert M, Kay AB: Eosinophil chemotactic chemokines (eotaxin, eotaxin-2, RANTES, monocyte chemoattractant protein-3 (MCP-3), and MCP-4), and C-C chemokine receptor 3 expression in bronchial biopsies from atopic and nonatopic (intrinsic) asthmatics. J Immunol 1999, 163:6321-6329 [PubMed] [Google Scholar]
- 23.Humbert M, Ying S, Corrigan C, Menz G, Barkans J, Pfister R, Meng Q, Van Damme J, Opdenakker G, Durham SR, Kay AB: Bronchial mucosal expression of the genes encoding chemokines RANTES and MCP-3 in symptomatic atopic and nonatopic asthmatics: relationship to the eosinophil-active cytokines interleukin (IL)-5, granulocyte macrophage-colony-stimulating factor, and IL-3. Am J Respir Cell Mol Biol 1997, 16:1-8 [DOI] [PubMed] [Google Scholar]
- 24.Powell N, Humbert M, Durham SR, Assoufi B, Kay AB, Corrigan CJ: Increased expression of mRNA encoding RANTES and MCP-3 in the bronchial mucosa in atopic asthma. Eur Respir J 1996, 9:2454-2460 [DOI] [PubMed] [Google Scholar]
- 25.Uguccioni M, Gionchetti P, Robbiani DF, Rizzello F, Peruzzo S, Campieri M, Baggiolini M: Increased expression of IP-10, IL-8, MCP-1, and MCP-3 in ulcerative colitis. Am J Pathol 1999, 155:331-336 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Stafford S, Li H, Forsythe PA, Ryan M, Bravo R, Alam R: Monocyte chemotactic protein-3 (MCP-3)/fibroblast-induced cytokine (FIC) in eosinophilic inflammation of the airways and the inhibitory effects of an anti-MCP-3/FIC antibody. J Immunol 1997, 158:4953-4960 [PubMed] [Google Scholar]
- 27.Zimmermann N, Hogan SP, Mishra A, Brandt EB, Bodette TR, Pope SM, Finkelman FD, Rothenberg ME: Murine eotaxin-2: a constitutive eosinophil chemokine induced by allergen challenge and IL-4 overexpression. J Immunol 2000, 165:5839-5846 [DOI] [PubMed] [Google Scholar]
- 28.Avrameas A, McIlroy D, Hosmalin A, Autran B, Debre P, Monsigny M, Roche AC, Midoux P: Expression of a mannose/fucose membrane lectin on human dendritic cells. Eur J Immunol 1996, 26:394-400 [DOI] [PubMed] [Google Scholar]
- 29.Sallusto F, Mackay CR, Lanzavecchia A: Selective expression of the eotaxin receptor CCR3 by human T helper 2 cells. Science 1997, 277:2005-2007 [DOI] [PubMed] [Google Scholar]
- 30.Sallusto F, Lanzavecchia A, Mackay CR: Chemokines and chemokine receptors in T-cell priming and Th1/Th2-mediated responses. Immunol Today 1998, 19:568-574 [DOI] [PubMed] [Google Scholar]
- 31.Sallusto F, Lenig D, Mackay CR, Lanzavecchia A: Flexible programs of chemokine receptor expression on human polarized T helper 1 and 2 lymphocytes. J Exp Med 1998, 187:875-883 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Gonzalo JA, Lloyd CM, Kremer L, Finger E, Martinez AC, Siegelman MH, Cybulsky M, Gutierrez-Ramos JC: Eosinophil recruitment to the lung in a murine model of allergic inflammation. The role of T cells, chemokines, and adhesion receptors. J Clin Invest 1996, 98:2332-2345 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lloyd CM, Delaney T, Nguyen T, Tian J, Martinez AC, Coyle AJ, Gutierrez-Ramos JC: CC chemokine receptor (CCR)3/eotaxin is followed by CCR4/monocyte-derived chemokine in mediating pulmonary T helper lymphocyte type 2 recruitment after serial antigen challenge in vivo. J Exp Med 2000, 191:265-274 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Imai T, Nagira M, Takagi S, Kakizaki M, Nishimura M, Wang J, Gray PW, Matsushima K, Yoshie O: Selective recruitment of CCR4-bearing Th2 cells toward antigen-presenting cells by the CC chemokines thymus and activation-regulated chemokine and macrophage-derived chemokine. Int Immunol 1999, 11:81-88 [DOI] [PubMed] [Google Scholar]
- 35.Panina-Bordignon P, Papi A, Mariani M, Di Lucia P, Casoni G, Bellettato C, Buonsanti C, Miotto D, Mapp C, Villa A, Arrigoni G, Fabbri LM, Sinigaglia F: The C-C chemokine receptors CCR4 and CCR8 identify airway T cells of allergen-challenged atopic asthmatics. J Clin Invest 2001, 107:1357-1364 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Yamamoto J, Adachi Y, Onoue Y, Adachi YS, Okabe Y, Itazawa T, Toyoda M, Seki T, Morohashi M, Matsushima K, Miyawaki T: Differential expression of the chemokine receptors by the Th1- and Th2-type effector populations within circulating CD4+ T cells. J Leukoc Biol 2000, 68:568-574 [PubMed] [Google Scholar]
- 37.Kim CH, Rott L, Kunkel EJ, Genovese MC, Andrew DP, Wu L, Butcher EC: Rules of chemokine receptor association with T cell polarization in vivo. J Clin Invest 2001, 108:1331-1339 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Christodoulopoulos P, Wright E, Frenkiel S, Luster A, Hamid Q: Monocyte chemotactic proteins in allergen-induced inflammation in the nasal mucosa: effect of topical corticosteroids. J Allergy Clin Immunol 1999, 103:1036-1044 [DOI] [PubMed] [Google Scholar]
- 39.Kitayama J, Mackay CR, Ponath PD, Springer TA: The C-C chemokine receptor CCR3 participates in stimulation of eosinophil arrest on inflammatory endothelium in shear flow. J Clin Invest 1998, 101:2017-2024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Rothenberg ME, Luster AD, Leder P: Murine eotaxin: an eosinophil chemoattractant inducible in endothelial cells and in interleukin 4-induced tumor suppression. Proc Natl Acad Sci USA 1995, 92:8960-8964 [DOI] [PMC free article] [PubMed] [Google Scholar]