

Abstract
In addition to its classical calciotropic effects, the active form of vitamin D, 1,25-dihydroxyvitamin D3 (1,25(OH)2D3) is a potent anti-proliferative/immunomodulatory secosteroid. The enzyme that catalyzes the synthesis of 1,25(OH)2D3, 1α-hydroxylase (1α-OHase), is expressed in many human tissues, highlighting its possible role as an autocrine/paracrine activator of vitamin D. Immunohistochemical and RNA analyses were used to characterize the ontogeny of 1α-OHase expression in human placenta and decidua. Protein for 1α-OHase was detectable in trophoblast and decidua; the latter being stronger in decidualized stromal cells than macrophages, with no staining of lymphocytes. Quantitative reverse transcriptase-polymerase chain reaction was used to assess changes in mRNA expression for 1α-OHase at different gestations: first (mean, 9.1 ± 1.5 weeks); second (mean, 14 ± 1.8 weeks), and third trimester (mean, 39.3 ± 2.5 weeks). 1α-OHase expression in decidua was ∼1000-fold higher in first (95% confidence limits, 611 to 1376) and second (95% confidence limits, 633 to 1623) trimester biopsies when compared with the third trimester (95% confidence limits, 0.36 to 2.81) (both P < 0.001). In placenta, 1α-OHase expression was 80-fold higher in the first (range, 42 to 137) and second (range, 30 to 199) trimester when compared with third trimester biopsies (0.6 to 1.6) (both P < 0.001). Similar results were obtained by semiquantitative IHC. Parallel analysis of the receptor for 1,25(OH)2D3 (vitamin D receptor) indicated that, as with 1α-OHase, highest levels of expression occurred in first trimester decidua. However, changes in vitamin D receptor mRNA expression across gestation were less pronounced than 1α-OHase. These spatiotemporal data emphasize the potential importance of 1α-OHase during early fetoplacental life and, in particular, suggest an autocrine/paracrine immunomodulatory function for the enzyme.
The active form of vitamin D [1,25-dihydroxyvitamin D3 (1,25(OH)2D3)], plays a pivotal role in mineral homeostasis by facilitating calcium and phosphate uptake in the gut and by modulating bone cell development and function. 1 However, in recent years, studies in vivo and in vitro have shown that 1,25(OH)2D3 is able to influence a wide range of functions not immediately linked to calcium homeostasis. In particular, potent anti-proliferative and immunosuppressive responses to 1,25(OH)2D3 have led to the use of vitamin D analogs as therapy for diseases such as psoriasis and leukemia. 2-4 The importance of 1,25(OH)2D3 as a pleiotropic modulator of tissue function is strengthened by the presence of key components of vitamin D metabolism and function at these sites. The cognate nuclear receptor for 1,25(OH)2D3, the vitamin D receptor (VDR), is ubiquitous in proliferating cells. Its function is therefore dependent primarily on the local availability of ligand. Synthesis of 1,25(OH)2D3 from the major circulating form of vitamin D, 25-hydroxyvitamin D3 (25(OH)D3), is catalyzed by the enzyme 25-hydroxyvitamin D3-1α-hydroxylase (1α-OHase) that is located in the kidney, 5,6 and at several key extra-renal sites. 2,7,8 Thus, the widespread co-expression of VDR and 1α-OHase emphasizes a putative role for 1,25(OH)2D3 as an autocrine/paracrine prodifferentiation and immunosuppressive agent, with diverse physiological functions.
Despite the fact that renal and extra-renal synthesis of 1,25(OH)2D3 is differentially regulated, the expression of 1α-OHase in different tissues seems to be because of the same gene product. 9-11 Based on this observation we have developed novel polyclonal antisera to the renal 1α-OHase that have allowed us to define for the first time the precise cellular distribution of the enzyme along the nephron 5 and within key peripheral tissues. 7 Our data have confirmed the immunolocalization of 1α-OHase in tissues such as lymph nodes and skin but also indicate that 1α-OHase is expressed in placentae from normal pregnancies. 7 In view of this, and the fact that reproduction in females is markedly diminished in states of vitamin D deficiency, 12-14 we have postulated that local synthesis of 1,25(OH)2D3 may play a role in implantation and/or placentation. To investigate further the role of local 1,25(OH)2D3 production in fetal-placental development we have performed immunohistochemical and quantitative reverse transcriptase-polymerase chain reaction (RT-PCR) analyses of 1α-OHase using placental and decidual tissue from different gestational ages and from cases of intrauterine growth restriction (IUGR).
Materials and Methods
Tissue Samples
The study had the approval of the local hospital ethics committee. Placenta and decidua were either collected from elective first or second trimester termination of pregnancy (in accordance with the Polkinghorne report 15 ) or from elective cesarean sections. For immunohistochemistry (IHC) a total of 58 placental and 25 decidual samples were fixed in 10% neutral buffered formalin for 24 to 48 hours, routinely processed, and embedded in paraffin wax. For RNA extraction placental and decidual samples were collected, snap-frozen in liquid nitrogen (−80°C), and grouped as shown in Table 1 ▶ . Thirteen cases of IUGR were diagnosed prospectively, having at least three of the four following characteristics determined from ultrasound examination: 1) fetal abdominal circumference (AC) ≤ third centile for gestational age; 2) abnormal fetal growth velocity (ΔAC < 1.5 SD throughout 14 days); 3) severe oligohydramnios (amniotic fluid index ≤ third centile for gestational age); 4) absent or reversed velocities in umbilical artery Doppler waveforms. The IUGR group ranged from 25 to 38 weeks of gestational age and was gestationally matched to a group of pregnancies with appropriately grown babies born between 27.3 and 34 weeks but delivered for maternal reasons and not associated with preterm labor (n = 5). Placental biopsies were taken from up to five randomly allocated areas; in previous studies we have shown relatively little variation in the expression of genes such as 11β-hydroxysteroid dehydrogenase within individual placentae. 16,17 Decidua was identified from termination samples with both macroscopical and histological confirmation performed. In third trimester samples decidua was mechanically scraped from the uterine cavity at cesarean section. Again, histological confirmation was obtained. Semiquantitative grading of immunohistochemical staining was performed blind by a single individual using randomly selected areas of each section.
Table 1.
Number of Individual Samples of Placenta or Decidua Tissue in Each Trimester and Intrauterine Growth Restriction (IUGR)
| Trimester (mean gestational age ±SD) | Placenta | Decidua |
|---|---|---|
| First (9.1± 1.5 wks) | 35 | 27 |
| Second (14.0± 1.8 wks) | 67 | 10 |
| Third (39.3± 1.8 wks) | 33 | 10 |
| IUGR (30.8± 1.8 wks) | 13 | |
| IUGR gestation-matched controls (30.6± 2.6 wks) | 5 |
Mean gestational age in weeks ± standard deviation (SD).
Immunohistochemistry
1α-OHase
Immunohistochemical (IHC) analysis of 1α-OHase expression was performed using previously described methods. 5,7 Briefly, dewaxed and rehydrated paraffin-embedded sections were processed in 0.01 mol/L of sodium citrate buffer (pH 6.0) in a pressure cooker at 103 kPa for 2 minutes for antigen retrieval. Slides were then incubated with methanol-hydrogen peroxide (1:100) to block endogenous peroxidase activity, and then washed in Tris-buffered saline, pH 7.6. The slides were then incubated with 1α-OHase antiserum (1:150) in 10% normal swine serum for 45 minutes at room temperature (25°C). After rinsing with Tris-buffered saline for 15 minutes, donkey anti-sheep IgG peroxidase conjugate (1:100; The Binding Site, Birmingham, UK) was added to sections for 45 minutes. Staining was developed using 3,3′-diaminobenzidine (2.5 mg/ml) followed by counterstaining with Mayer’s hematoxylin. Negative control sections included: 1) omission of primary antibody; 2) use of primary antibody preabsorbed with a 100-fold excess of immunizing peptide. Immunolabeled slides were assessed semiquantitively blind to the gestational age of the samples. Reactivity was graded as negative, weak, moderate, or strong, assessed against a known strongly positive sample (kidney) that was included in each staining run.
Double-Immunohistochemical Characterization of 1α-OHase-Positive Cells
Selected first and second trimester decidual samples (n = 10) were subjected to double-immunoenzymatic labeling to further characterize the 1α-OHase immunoreactive cells. Firstly, sections were labeled for 1α-OHase using an indirect immunoperoxidase method as described above, except that the reaction was developed in aminoethylcarbazole (Vector NovaRed; Vector Laboratories, Peterborough, UK) rather than in 3,3′-diaminobenzidine to give a red-brown reaction product. Sections were then incubated for 60 minutes with monoclonal antibodies directed against CD3 (T lymphocytes, NCL-CD3-PS1 1/20; Novocastra Laboratories, Newcastle on Tyne, UK), CD56 (NCAM, natural killer cells including endometrial granulated lymphocytes, NCL-CD56–1B6 1/50; Novocastra), CD14 (monocytes/macrophages, NCL-CD14–223; Novocastra), CD68 (monocytes/macrophages, KP1 1/50; DAKO, Ely, UK), and CD10 (endometrial stromal cells, NCL-CD10–270 1/40; Novocastra). The second antibody reaction was detected using an avidin-biotin-alkaline phosphatase technique (Vectastain alkaline phosphatase kit, Vector Laboratories) and the reaction was developed for 20 minutes in the dark at room temperature with the alkaline-phosphatase substrate kit III (Vecta Blue, Vector Laboratories). The reaction was stopped with excess water. Sections were not counterstained and were initially mounted in Supermount (Biogenix, San Ramon, CA), air-dried overnight, and then mounted in synthetic resin (DPX; Raymond Lamb, London, UK). Negative controls included omission of the first and second primary antibody in turn to exclude nonspecific binding and spurious double labeling.
RNA Extraction
RNA was extracted from placental and decidual samples using the TRI reagent mRNA extraction method (Sigma, Poole, UK) or the StrataPrep total RNA miniprep kit (Stratagene, Amsterdam, Netherlands). An aliquot (2 μg) of RNA from each sample was reverse-transcribed (RT) using AMV reverse transcriptase (Promega, Madison, WI) and random hexamers in 40-μl reaction volumes according to the manufacturer’s instructions.
Quantitative RT-PCR Analysis of 1α-OHase mRNA Expression
1α-OHase mRNA levels were analyzed using an ABI 7700 sequence detection system (PE Biosystems, Warring, UK). Amplification of 1α-OHase cDNA was performed in 25-μl volumes on 96-well plates, in a reaction buffer containing TaqMan Universal PCR Master Mix, 3 mmol/L Mn(Oac)2, 200 μmol/L dNTPs, 1.25 U AmpliTaq Gold polymerase, 1.25 U AmpErase uracil-N-glycosylase (UNG), 150 nmol TaqMan probe, 900 nmol primers, and 50 ng cDNA. All reactions were multiplexed with the housekeeping gene 18S rRNA, provided as a preoptimized control probe (PE Biosystems) enabling data to be expressed in relation to an internal reference to allow for differences in sampling and RT efficiency. Data were obtained as Ct values (the cycle number at which logarithmic PCR plots cross a calculated threshold line) according to the manufacturer’s guidelines, and used to determine ΔCt values (ΔCt = Ct of the target gene − Ct of the housekeeping gene). Measurements were performed in triplicate and data were reported as the mean of three values from separate experiments. All target gene probes were labeled with five-carboxy fluorescein (FAM), and the housekeeping gene with VIC. All PCR reactions were performed with the primers and probes outlined below using the following reaction conditions: 50°C for 2 minutes, 95°C for 10 minutes, 44 cycles of 95°C for 15 seconds, 60°C for 1 minute.
Primers and Probes for PCR Reactions
PCR primer and probe sequences for 1α-OHase were as follows: forward primer, CACCCGACACGGAGACCTT; reverse primer, TCAACAGCGTGGACACAAACA; and TaqMan probe, TCCGCGCTGTGGGCTCGG. For cytokeratin-8 (CK-8): forward primer, 5′-ATGTGGATGAAGCATACATGAACA-3′; reverse primer, 5′-TCCCGGATCTCCTCTTCAT-3′; and TaqMan probe, 5′-CCGACGAGATCAA-CTTCCTCAGGCA-3′. For VDR: forward primer, 5′-CTTCA-GGCGAAGCATGAAGC-3′; reverse primer, 5′-CCTTCAT-CATGCCGATGTCC-3′; and TaqMan probe, 5′-AAGGCA-CTATTCACCTGCCCCTTCAA-3′.
Data Analysis
To exclude potential bias because of averaging data that had been transformed through the equation 2−ddCt, all statistics were performed at the ΔCt stage. Statistical analysis was performed using one-way analysis of variance, with posthoc testing using Student-Newman-Keuls multiple comparison posttest (Instat version 2.04a computer program; GraphPad Software, Inc., San Diego, CA).
Results
Tissue Distribution of 1α-OHase
Analysis of 1α-OHase expression by IHC demonstrated the presence of the enzyme in both placental and decidual tissue (Figure 1) ▶ . Strong reactivity was observed in villous syncytiotrophoblast from first and second trimester placentae with less intense immunostaining of the villous cytotrophoblast (Figure 1A) ▶ . Cytotrophoblast columns and cytotrophoblast islands showed comparable staining with villous cytotrophoblast. Lower (moderate) levels of 1α-OHase expression were detected in third trimester villous syncytiotrophoblast (Figure 1B) ▶ . Third trimester villous cytotrophoblast was generally scanty but was weakly positive for 1α-OHase. Extravillous trophoblast cells in the basal plate were weakly positive. Relatively high levels of 1α-OHase protein were also observed in most samples of first trimester decidua (Figure 1C) ▶ , with reduced expression in decidua basalis from third trimester samples (Figure 1D) ▶ . Parallel analysis of cell surface markers for trophoblast (cytokeratin) and decidualized stromal cells (vimentin) confirmed that 1α-OHase was expressed by both cell types (data not shown). In addition, further IHC analyses showed that 1α-OHase was also expressed in glandular tissue. Figure 1E ▶ shows double-labeling IHC for 1α-OHase (red-brown staining) and the decidual stromal cell marker CD10 (blue staining), with glands expressing 1α-OHase only and stromal cells showing co-expression of the enzyme with CD10 (purple staining).
Figure 1.
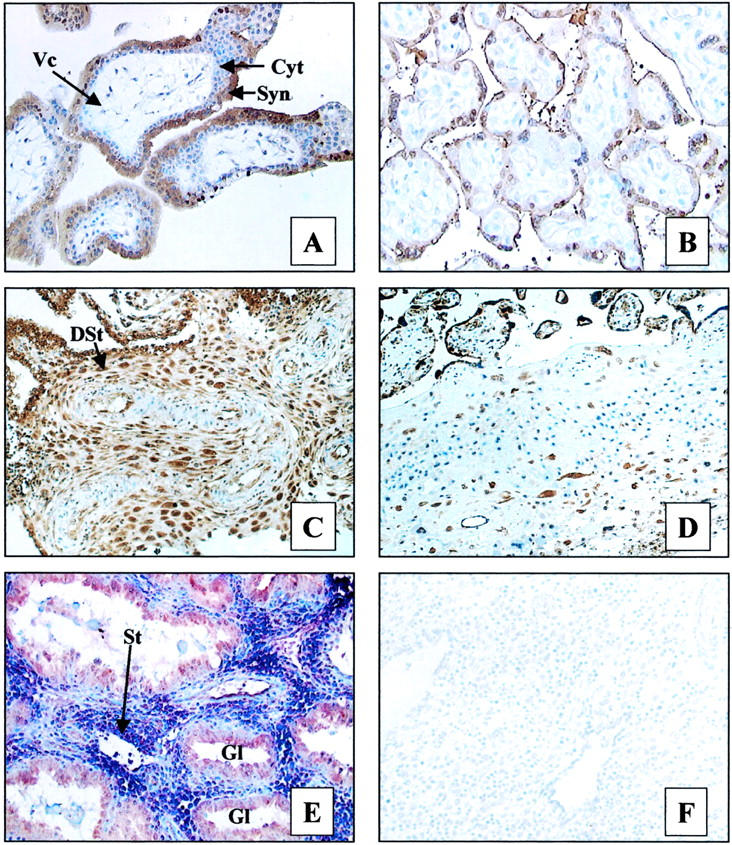
Immunohistochemical analysis of 1α-OHase expression in human placenta and decidua (brown staining). A: First trimester placenta. B: Third trimester placenta. C: First trimester decidua. D: Third trimester decidua. E: Double labeling of decidua with 1α-OHase antiserum (brown staining) and antibody to the stromal cell marker CD10 (blue staining). F: Negative control for specificity of 1α-OHase expression (preabsorption of antiserum with 200-fold excess of immunizing peptide). Tissue features: Syn, syncytiotrophoblast; Cyt, cytotrophoblast; Vc, villous core; St, stromal cells; DSt, decidual stromal cells; GL, gland. Original magnifications: ×200 (A, B, E, F); ×150 (C, D).
Expression of 1α-OHase in Decidual Cell Subgroups
In view of the relatively high levels of 1α-OHase expression in early pregnancy, double-labeling studies were performed to determine the specific decidual cell subgroups associated with the enzyme (Figure 2) ▶ . Both decidualized and nondecidualized endometrial stromal cells showed immunoreactivity for CD10, although staining was more intense in areas without prominent decidual change in stromal cells. Reactivity for 1α-OHase was more intense in decidualized stromal cells that showed weak CD10 immunostaining, whereas strongly CD10-positive nondecidualized or poorly decidualized endometrial stromal cells showed weaker reactivity for the enzyme (Figures 1E and 2B) ▶ ▶ . The second major decidual leukocyte population, macrophages, showed double labeling for 1α-OHase with either CD14 (Figure 2C) ▶ or CD68 (data not shown). In all first and second trimester decidua studied, up to 25% of the CD14-positive and CD68-positive population co-expressed the enzyme. CD68 was also expressed, albeit less intensely, by decidualized stromal cells. However, in contrast to CD10, CD68 was detected in decidualized stromal cells that were 1α-OHase-positive, but was not detected in the nondecidualized stromal cells that showed weaker 1α-OHase immunostaining (data not shown). Double-labeling studies using antibodies to CD56 (Figure 2D) ▶ and CD3 (data not shown) confirmed that decidual T lymphocytes and endometrial granulated lymphocytes, respectively, did not co-express 1α-OHase.
Figure 2.
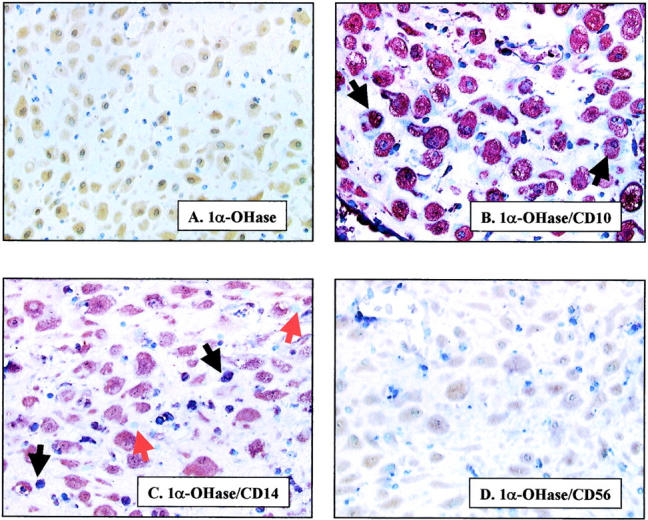
Co-localization of 1α-OHase with markers of stromal cell subtypes in first trimester decidua. A: Staining for 1α-OHase protein only (brown staining). B–D: Double labeling with 1α-OHase antiserum (brown staining) and antibodies to: stromal cell marker CD10 (blue staining) (B); monocyte marker CD14 (blue staining) (C); and lymphocyte marker CD56 (blue staining) (D). Blue staining only is shown by red arrows. Co-localization (purple staining) is shown by black arrows. Original magnifications, ×250.
Quantitative Analysis of Gestational Changes in 1α-OHase mRNA Expression
Data from real-time RT-PCR analyses revealed strong expression of 1α-OHase mRNA in first and second trimester placenta and decidua (Figure 3) ▶ . Furthermore, decidual samples showed 10-fold higher levels of mRNA when compared to placenta. In both types of tissue, expression of 1α-OHase decreased dramatically in third trimester and IUGR placentae. The latter showed a small but significant decrease in 1α-OHase expression when compared to normal third trimester placenta but IUGR values were not significantly different to the gestationally matched placental samples. To assess the extent to which changes in 1α-OHase expression were because of the altered cellular composition of placental and decidual tissue, further RT-PCR studies were performed for CK-8, a trophoblast/endometrial epithelium cell marker (Figure 4) ▶ . CK-8 levels in the first and second trimester placenta were not significantly different from third trimester or IUGR samples (including gestational controls). In contrast, decidual CK-8 levels paralleled changes in 1α-OHase with 100-fold lower levels of expression in third trimester samples. Based on these observations, further analysis of 1α-OHase expression was performed after correction for CK-8 mRNA (Figure 5) ▶ . Data revealed comparable levels of 1α-OHase mRNA in first trimester placenta and decidual tissue, although in both types of tissue expression of 1α-OHase remained lower in third trimester samples. Importantly, after correction for CK-8, the expression of 1α-OHase in IUGR samples did not differ significantly from normal third trimester placentae or gestationally matched samples.
Figure 3.

Expression of 1α-OHase mRNA in human placenta and decidua. A: Placental and decidual 1α-OHase mRNA expression normalized for 18S rRNA levels in first, second, and third trimester samples, and placentae from patients with IUGR. All data are reported relative to a third trimester placenta value of 1. *, Significantly different from third trimester placenta, P < 0.001; **, significantly different from third trimester placenta, P < 0.05; #, significantly different from equivalent placenta, P < 0.01. B: Data expressed with 95% confidence limits. IUGR controls, gestationally matched placentae.
Figure 4.

Expression of mRNA for CK-8 in human placenta and decidua. A: Placental and decidual CK-8 mRNA expression normalized for 18S rRNA levels in first, second, and third trimester samples and placentae from patients with IUGR. All data are reported relative to a third trimester placenta value of 1. *, Significantly different from third trimester placenta, P < 0.001; #, significantly different from equivalent placenta, P < 0.001. B: Data expressed with 95% confidence limits. IUGR controls, gestationally matched placentae.
Figure 5.

Changes in the expression of 1α-OHase in human placenta and decidua with gestation are independent of variations in trophoblast/endometrial epithelial cell status. A: Expression of 1α-OHase mRNA in human placenta and decidua corrected for 18S rRNA and CK-8 mRNA expression. All data are reported relative to a third trimester placenta value of 1. *, Significantly different from third trimester placenta, P < 0.001; **, significantly different from third trimester placenta, P < 0.05. B: Data expressed with 95% confidence limits. IUGR controls, gestationally matched placentae.
Further studies were performed to compare spatiotemporal changes in 1α-OHase mRNA levels with gestational variations in VDR expression. Data, again corrected for CK-8 expression (Figure 6) ▶ , indicated that, as with 1α-OHase, VDR expression is highest in early gestation. However, in contrast to 1α-OHase, changes in VDR expression across gestation were less pronounced with only first trimester decidua and second trimester placenta expressing mRNA at levels that were significantly higher than third trimester placental tissue.
Figure 6.

Changes in the expression of VDR mRNA in human placenta and decidua with gestation. A: Expression of VDR mRNA in human placenta and decidua corrected for 18S rRNA and CK-8 mRNA expression. All data are reported relative to a third trimester placenta value of 1. *, Significantly different from third trimester placenta, P < 0.05. B: Data expressed with 95% confidence limits. IUGR controls, gestationally matched placentae.
Discussion
Detailed analysis of the nonclassical actions of vitamin D has shown that its active form, 1,25(OH)2D3, is a pluripotent secosteroid with immunomodulatory and anti-proliferative properties. The clinical importance of this is most clearly illustrated by the development of synthetic analogs of 1,25(OH)2D3 as therapy for autoimmune disease, psoriasis, and cancer. 2-4,18 In contrast, our understanding of the impact of 1,25(OH)2D3 on reproduction is much less clear despite evidence linking vitamin D status with reproductive function. Spermatogenesis is compromised by vitamin D deficiency but this can be corrected by calcium supplementation, suggesting an indirect effect of 1,25(OH)2D3 on male reproduction. 19 Female fertility also seems to be markedly reduced in vitamin D deficiency but may involve both calcium-dependent or -independent actions depending on the animal model used. In vitamin D-deficient rats decreased fertility and litter size seems to be independent of calcium status. 12-14 Conversely female reproductive efficiency in VDR-null mice seems to be a direct consequence of altered calcium status. 20,21 In either model the precise mechanism for this has yet to be determined, although ovarian cells express VDR and are an established target tissue for 1,25(OH)2D3. 22-24 Recent studies have suggested that 1,25(OH)2D3 is able to influence ovarian function by modulating local estrogen synthesis. VDR-null mice show decreased expression of the estrogenic enzyme aromatase in ovaries and testis, although this seemed to be because of, in part, altered calcium homeostasis. 21
Previous reports have also implicated 1,25(OH)2D3 in fetoplacental development. In particular, maternal circulating levels of 1,25(OH)2D3 more than double with advancing gestation, resulting in a concomitant increase in calcium uptake by the maternal intestine. 25 Although 1,25(OH)2D3 is the principal stimulator of calcium uptake in the gut, its role in placental calcium handling has yet to be clearly defined. Perfusion studies in animals using pharmacological doses of 1,25(OH)2D3 have demonstrated an increase in fetal blood calcium, placental calcium transport, and fetal calcium status. 26 Furthermore, previous nephrectomy of animals has been shown to result in decreased placental calcium transport. 27 However, these observations are further complicated by the fact that decidual and placental tissues are themselves a key site for peripheral 1,25(OH)2D3 production. Both decidua and trophoblast cells express VDR as well as 1α-OHase, suggesting an autocrine or paracrine role for 1,25(OH)2D3 within these tissues. 28-33 In particular, we have postulated that local activation of vitamin D may influence implantation, either through the established immunomodulatory effects of 1,25(OH)2D3 or via the regulation of specific target genes associated with implantation. To test this hypothesis we have characterized spatiotemporal variations in 1α-OHase expression in human decidua and placenta.
IHC data showed that 1α-OHase protein is expressed by both syncytio- and cytotrophoblast layers of the placental chorionic villus. These observations are similar to previous reports of 1α-OHase mRNA expression. 31,33 However, the intensity of immunoreactivity (as measured using a blinded semiquantitative scoring system), seemed to be greater in first and second trimester pregnancies. In particular, decidual samples from early pregnancy showed strong immunolocalization of 1α-OHase within the decidualized endometrial stromal cells. Although there are no specific markers for these cells, CD10 and CD68 were both detectable in these cells, with the intensity of expression seeming to be related to the degree of decidualization. Lymphocytes were negative for 1α-OHase but double-labeling IHC studies demonstrated reactivity for 1α-OHase in a substantial proportion of decidual macrophages. Macrophages are less abundant than granulated lymphocytes in the first half of pregnancy but they persist throughout gestation and are often closely associated with extravillous trophoblast in the decidua basalis. 34
The presence of 1α-OHase in decidual macrophages is analogous to that previously described for lymphatic tissues in diseases such as sarcoidosis and tuberculosis, in which local synthesis of 1,25(OH)2D3 has the potential to influence several elements of the local immune system. The hormone may act in an autocrine/intracrine manner to enhance cellular immune function, including cytokine production, 35,36 antigen presentation, 37 and cytotoxic function. 38 Local production of 1,25(OH)2D3 by macrophages may also influence lymphocyte development and function, 39 as well as T-cell-mediated B-cell immunoglobulin production. 40 Modulation of specific T-cell subsets may also occur. Specifically, 1,25(OH)2D3 promotes a shift away from cell-mediated (Th1-type) responses toward humoral immunity (Th2-type) with accompanying changes in cytokine expression. 41 In this way, local activation of vitamin D seems to act by maximizing the ability of the host to control the activating antigen while minimizing the potentially self-destructive effects of unregulated lymphocyte activity. The immunological nature of decidual tissue suggests that the synthesis 1,25(OH)2D3 by decidual cells may act in a paracrine manner to modulate local lymphocyte activity. Evidence is accumulating from both mouse and humans that successful pregnancy is associated with a bias toward Th2-type cytokines, Th1-type responses being suppressed both systemically and locally. 42 We can therefore postulate that vitamin D may play a role in promoting this shift away from cell-mediated immune responses in pregnancy. It would be of interest to examine levels of 1α-OHase in pregnancy disorders such as recurrent miscarriage and primary infertility that have been associated with abnormalities of this Th1/Th2 shift.
The importance of local 1,25(OH)2D3 production in early decidual function is emphasized by the variations in 1α-OHase and VDR expression across gestation. Levels of 1α-OHase mRNA were maximal in first and second trimester placentae and deciduae, compared with the third trimester, even after adjustment for changes in the cellular population (as determined by CK-8 levels). VDR expression also seems to be relatively high in early gestation although cross-gestational changes in mRNA for the receptor were less pronounced than for 1α-OHase. Collectively, these observations suggest that in early gestation decidual cells are primed for both synthesis of and response to 1,25(OH)2D3. This is consistent with a more localized function for the hormone with 1α-OHase being the most likely rate-limiting mechanism. During early pregnancy the extravillous trophoblast invades uterine decidua, myometrium, and spiral arteries, establishing the uteroplacental blood supply. This invasive trophoblast expresses the nonclassical HLA-G class I MHC antigen as well as HLA-C. 43 At this stage of pregnancy the uterine decidua contains an abundant population of granulated lymphocytes, capable of natural killer cell activity, as well as CD8-positive T lymphocytes. 44 The mechanisms that allow the potentially semiallogeneic fetally derived extravillous trophoblast cells to migrate through maternal uterine tissues remain to be established but suggest a local modification of immune responses.
In addition to its potent immunosuppressive actions 1,25(OH)2D3 may influence implantation by regulating key developmental target genes, such as homeobox (HOX) genes. In particular, the HOXA10 gene has been shown to be intimately associated with implantation. 45 Female homozygotes with HOXA10 knock outs ovulate and fertilize normally but demonstrate embryo lethality at days 2.5 to 3.5. This coincides spatially and temporally with expression of maternal HOXA10 in distal oviductal and uterine epithelium. 46 These results indicate a role for HOX genes in female fertility and suggest that maternal HOXA10 is required to regulate the expression of a factor that affects the viability of preimplantation embryos. 47 Human endometrium shows up-regulation of HOXA10 expression at the time of implantation, indicating a conserved role in women. 48 Recent reports have highlighted a link between vitamin D and HOXA10. Studies in vitro using myeloid leukemic cells have shown that 1,25(OH)2D3 stimulates a dose-dependent increase in HOXA10 expression. 49 This seems to be because of direct regulation of transactivation as other reports have documented a functional vitamin D response element in the promoter of the HOXA10 gene. 50
Temporal data indicate that 1α-OHase expression is much stronger in the first and second trimesters. However, the presence of detectable levels of the enzyme in third trimester placenta emphasizes that 1α-OHase may have functions across gestation. This is consistent with previous studies that have characterized 1α-OHase mRNA expression in human syncytiotrophoblast cells. 33 To investigate the possible impact of 1,25(OH)2D3 synthesis on later stages of gestation, we assessed 1α-OHase expression in placentae from IUGR pregnancies. IUGR is characterized by abnormal placentation and remains a major cause of perinatal morbidity and mortality. The specific cohort of IUGR pregnancies with uteroplacental failure described in this article have a particularly poor prognosis. The villous placentae of these pregnancies are significantly smaller than normal, with reduced cytotrophoblast proliferation and abnormal vasculature. Also, in common with preeclampsia, there is a paucity of extravillous trophoblast invasion of the maternal spiral arteriolar circulation. However, in contrast to previous studies of preeclampsia that have highlighted an association with decreased circulating levels of 1,25(OH)2D3, 51-53 relatively little is known about the impact of vitamin D on IUGR. Our observations indicated that 1α-OHase levels were lower in IUGR samples compared to normal third trimester placentae. However, this was not statistically significant after normalization for changes in CK-8 expression and, more importantly, the IUGR samples showed no difference in 1α-OHase expression compared to gestationally matched controls.
Data presented here indicate that both mRNA and protein for 1α-OHase are abundantly expressed in placenta and decidua from first and early second trimester pregnancies. The relatively high levels of 1α-OHase in decidua raises the possibility that the enzyme plays a local role in human implantation and/or placentation. In particular, expression of the enzyme by decidualized stromal cells and macrophages suggests an autocrine/paracrine modulatory role for 1,25(OH)2D3 within this immune privileged site. A potential role for vitamin D at the maternofetal interface was first postulated by Rebut-Bonneton and Demingnon 54 who showed that 1,25(OH)2D3 inhibited the proliferation of allogenically-stimulated lymphocytes and cytotoxic T cells in vitro. Analysis of placental/decidual 1α-OHase expression suggests that, in vivo, the effects of 1,25(OH)2D3 are more likely to be a consequence of locally synthesized hormone rather than circulating levels. In this respect it is important to recognize that, unlike its kidney counterpart, extra-renal 1α-OHase is not subject to tight autoregulation. 2,8 As such the amount of 1,25(OH)2D3 synthesized by placenta and decidua will primarily be a reflection of the availability of substrate; in other words dietary/environmental access to precursor vitamin D. We can therefore postulate that the decreased fertility associated with vitamin D deficiency 12-14,19 is because of suboptimal synthesis of 1,25(OH)2D3 in placenta/decidua, with concomitant loss of immune regulation in these tissues. Further analysis of implantation and placentation in the recently reported 1α-OHase knockout mouse 55 will help to clarify this mechanism.
Footnotes
Address reprint requests to Dr. Martin Hewison, Division of Medical Sciences, The University of Birmingham, Queen Elizabeth Hospital, Birmingham, B15 2TH, UK. E-mail: m.hewison@bham.ac.uk.
Supported by Biotechnology and Biological Sciences Research Council project grant number 6/S14523.
References
- 1.Feldman D: Vitamin D, parathyroid hormone, and calcium: a complex regulatory network. Am J Med 1999, 107:637-639 [DOI] [PubMed] [Google Scholar]
- 2.Hewison M, Gacad MA, Lemire J, Adams JS: Vitamin D as a cytokine and hematopoietic factor. Rev Endocrinol Metab Disorders 2001, 2:217-227 [DOI] [PubMed] [Google Scholar]
- 3.Jones G, Strugnell SA, DeLuca HF: Current understanding of the molecular actions of vitamin D. Physiol Rev 1998, 78:1193-1231 [DOI] [PubMed] [Google Scholar]
- 4.Bouillon R, Garmyn M, Verstuyft A, Segaert S, Casteels K, Mathieu C: Paracrine role for calcitriol in the immune system and skin creates new therapeutic possibilities for vitamin D analogs. Eur J Endo 1995, 133:7-16 [DOI] [PubMed] [Google Scholar]
- 5.Zehnder D, Bland R, Walker EA, Bradwell AR, Howie AJ, Hewison M, Stewart PM: Expression of 25-hydroxyvitamin D3-1α-hydroxylase in the human kidney. J Am Soc Nephrol 1999, 10:2465-2473 [DOI] [PubMed] [Google Scholar]
- 6.Bland R, Zehnder D, Hewison M: Expression of 25-hydroxyvitamin D3-1α-hydroxylase along the nephron: new insights into renal vitamin D metabolism. Curr Opin Nephrol Hypertens 2000, 9:17-22 [DOI] [PubMed] [Google Scholar]
- 7.Zehnder D, Bland R, Williams MC, McNinch RW, Howie AJ, Stewart PM, Hewison M: Extra-renal expression of 25-hydroxyvitamin D3-1α-hydroxylase. J Clin Endocrinol Metab 2001, 86:888-894 [DOI] [PubMed] [Google Scholar]
- 8.Bell NH: Renal and non-renal 25-hydroxyvitamin D-1α-hydroxylases and their clinical significance. J Bone Miner Res 1999, 13:350-353 [DOI] [PubMed] [Google Scholar]
- 9.Takeyama K, Kitanaka S, Sato T, Kobori M, Yanagisawa J, Kato S: 25-Hydroxyvitamin D3 1α-hydroxylase and vitamin D synthesis. Science 1997, 277:1827-1830 [DOI] [PubMed] [Google Scholar]
- 10.Fu KG, Lin D, Zhang MYH, Bikle DD, Shackleton CHL, Miller WL, Portale AA: Cloning of human 25-hydroxyvitamin D-1α-hydroxylase and mutations causing vitamin D-dependent Rickets type I. Mol Endocrinol 1997, 11:1961-1970 [DOI] [PubMed] [Google Scholar]
- 11.Smith SJ, Rucka AK, Berry JL, Davies M, Mylchreest S, Paterson CR, Heath DA, Tassabehji M, Read AP, Mee AP, Mawer EB: Novel mutations in the 1 alpha-hydroxylase (P450cl) gene in three families with pseudovitamin D-deficiency rickets resulting in loss of functional enzyme activity in blood-derived macrophages. J Bone Miner Res 1999, 14:730-739 [DOI] [PubMed] [Google Scholar]
- 12.Halloran BP, DeLuca HF: Effect of vitamin D deficiency on fertility and reproductive capacity in the female rat. J Nutr 1980, 110:1573-1580 [DOI] [PubMed] [Google Scholar]
- 13.Hickie JP, Lavigne DM, Woodward WD: Reduced fecundity of vitamin D deficient rats. Comp Biochem Physiol A 1983, 74:923-925 [DOI] [PubMed] [Google Scholar]
- 14.Kwiecinksi GG, Petrie GI, DeLuca HF: 1,25-Dihydroxyvitamin D3 restores fertility of vitamin D-deficient female rats. Am J Physiol 1989, 256:E483-E487 [DOI] [PubMed] [Google Scholar]
- 15.Polkingthorne Report. Review of the Guidance on the Research of Fetuses and Fetal Material. London, Her Majesty’s Stationery Office, 1989
- 16.Somerset DA, Li X-F, Afford S, Strain AJ, Ahmed AS, Sangha R, Whittle MJ, Kilby MD: Ontogeny of hepatocyte growth factor and its receptor in human pregnancy: reduced expression in IUGR. Am J Pathol 1998, 153:1139-1147 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.McTernan CL, Draper N, Nicholson H, Chalder SM, Driver P, Hewison M, Kilby MD, Stewart PM: Reduced placental 11β-hydroxysteroid dehydrogenase type 2 mRNA levels in human pregnancies complicated by intrauterine growth restriction: an analysis of possible mechanisms. J Clin Endocrinol Metab 2001, 86:4979-4983 [DOI] [PubMed] [Google Scholar]
- 18.Bouillon R, Okamura WH, Norman AW: Structure-function relationships in the vitamin D endocrine system. Endocr Rev 1995, 16:200-257 [DOI] [PubMed] [Google Scholar]
- 19.Kwiecinski GG, Petrie GI, Deluca HF: Vitamin D is necessary for reproductive functions in the male rat. J Nutr 1989, 119:741-744 [DOI] [PubMed] [Google Scholar]
- 20.Yoshizawa T, Handa Y, Uematsu Y, Takeda S, Sekine K, Yoshihara Y, Kawakami T, Arioka K, Sato H, Uchiyama Y: Mice lacking the vitamin D receptor exhibit impaired bone formation, uterine hypoplasia and growth retardation after weaning. Nat Genet 1997, 16:391-396 [DOI] [PubMed] [Google Scholar]
- 21.Johnson LE, Deluca HF: Vitamin D receptor null mice fed high levels of calcium are fertile. J Nutr 2001, 131:1787-1791 [DOI] [PubMed] [Google Scholar]
- 22.Dokoh SC, Donaldson A, Marion SL, Pike JW, Haussler MR: The ovary: a target organ for 1,25-dihydroxyvitamin D3. Endocrinology 1983, 112:200-206 [DOI] [PubMed] [Google Scholar]
- 23.Stumpf WE, Sar M, Reid FA, Tanaka Y, Deluca HF: Target cells for 1,25-dihydroxyvitamin D3 in intestinal tract, stomach, kidney, skin, pituitary and parathyroid. Science 1979, 206:1188-1190 [DOI] [PubMed] [Google Scholar]
- 24.Kinuta K, Tanaka H, Moriwake T, Aya K, Kato S, Seino Y: Vitamin D is an important factor in estrogen biosynthesis of both female and male gonads. Endocrinology 2000, 141:1317-1324 [DOI] [PubMed] [Google Scholar]
- 25.Whitehead M, Lane G, Young O, Campbell S, Abeyasekara G, Hillyard CJ, MacIntyre I, Phang KG, Stevenson JC: Interrelations of calcium-regulating hormones during normal pregnancy. Br Med J 1981, 283:10-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Durand D, Braithewaite GD, Barlet JP: The effect of 1α-hydroxycholecalciferol on the placental transfer of calcium and phosphate in sheep. Br J Nutr 1983, 49:475-480 [DOI] [PubMed] [Google Scholar]
- 27.Care AD: The placental transfer of calcium. J Dev Physiol 1991, 15:253-257 [PubMed] [Google Scholar]
- 28.Weisman Y, Varjs A, Duckett G, Reiter E, Root AW: Synthesis of 1,25-dehydroxyvitamin D in the nephrectomized pregnant rat. Endocrinology 1978, 103:1992-1996 [DOI] [PubMed] [Google Scholar]
- 29.Wiseman Y, Harell A, Edelstein S, David M, Spirer Z, Golander A: 1α,25-dehydroxyvitamin D3 and 24, 25-dehydroxyvitamin D3 in-vitro synthesis by decidua and placenta. Nature 1979, 281:317-319 [DOI] [PubMed] [Google Scholar]
- 30.Tanaka Y, Halloran B, Schnoes HK, DeLuca HF: In vitro production of 1,25-dihydroxyvitamin D3 by rat placental tissue. Proc Natl Acad Sci USA 1979, 76:5033-5035 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Gray TK, Lester GE, Lorenc RS: Evidence for extra-renal 1α-hydroxylation of 25-hydroxyvitamin D3 in pregnancy. Science 1979, 204:1311-1313 [DOI] [PubMed] [Google Scholar]
- 32.Delvin EE, Arabian A, Glorieux FH, Mamer OA: In vitro metabolism of 25-hydroxycholecalciferol by isolated cells from human decidua. J Clin Endocrinol Metab 1985, 60:880-885 [DOI] [PubMed] [Google Scholar]
- 33.Diaz L, Sanchez I, Avila E, Halhali A, Vilchis F, Larrea F: Identification of a 25-hydroxyvitamin D3 1-α-hydroxylase gene transcription product in cultures of human syncytiotrophoblast. J Clin Endocrinol Metab 2000, 85:2543-2549 [DOI] [PubMed] [Google Scholar]
- 34.Bulmer JN, Smith J, Morrison L, Wells M: Maternal and fetal cellular relationships in the human placental basal plate. Placenta 1988, 9:237-246 [DOI] [PubMed] [Google Scholar]
- 35.Bhalla AK, Amento EP, Krane SM: Differential effects of 1,25-dihydroxyvitamin D3 on human lymphocytes and monocyte/macrophages: inhibition of interleukin-2 and augmentation of interleukin-1 production. Cell Immunol 1986, 98:311-322 [DOI] [PubMed] [Google Scholar]
- 36.Ucla C, Roux-Lombard P, Dayer J-M, Mach B: IFN-gamma drastically modifies the regulation of IL-1 genes by endotoxin in U937 cells. J Clin Invest 1990, 85:185-191 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Morel PA, Manolagas SC, Provvedini DM, Wegmann DR, Ciller JM: Interferon-gamma-induced IA expression in WEHI-3 cells is enhanced by 1,25-dihydroxyvitamin D3. J Immunol 1986, 57:159-163 [PubMed] [Google Scholar]
- 38.Rook GAW, Steele J, Fraher LJ, Barker S, Karmali R, O’Riordan JLH: Vitamin D3, gamma-interferon and control of proliferation of mucobacterium tuberculosis by human monocytes. Immunology 1986, 57:159-163 [PMC free article] [PubMed] [Google Scholar]
- 39.Lemire JM, Adams JS, Kermani-Arab V, Bakke AC, Sakai R, Jordan SC: 1,25-Dihydroxyvitamin D3 suppresses human T helper/inducer lymphocyte activity in vitro. J Immunol 1985, 134:3032-3035 [PubMed] [Google Scholar]
- 40.Lemire JM, Adams JS, Sakai R, Jordan SC: 1,25-Dihydroxyvitamin D3 suppresses proliferation and immunoglobulin production by normal human peripheral blood mononuclear cells. J Clin Invest 1984, 74:657-661 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Lemire JM, Archer DC, Beck L, Spiegelberg HL: Immunosuppressive actions of 1,25-dihydroxyvitamin D3: preferential inhibition of Th1 functions. J Nutr 1995, 125:1704S-1708S [DOI] [PubMed] [Google Scholar]
- 42.Piccinni MP, Scaletti C, Maggi E, Romagnani S: Role of hormone-controlled Th1- and Th2-type cytokines in successful pregnancy. J Neuroimmunol 2000, 109:30-33 [DOI] [PubMed] [Google Scholar]
- 43.King A, Hiby SE, Gardner L, Joseph S, Bowen JM, Verma S, Burrows TD, Loke YW: Recognition of trophoblast HLA class I molecules by decidual NK cell receptors—a review. Placenta 2000, 21(Suppl A):S81-S85 [DOI] [PubMed] [Google Scholar]
- 44.Johnson PM, Christmas SE, Vince GS: Immunological aspects of implantation and implantation failure. Hum Reprod 1999, 14(Suppl 2):S26-S36 [DOI] [PubMed] [Google Scholar]
- 45.Daftary GS, Taylor HS: Implantation in the human: the role of HOX genes. Semin Reprod Med 2000, 18:311-320 [DOI] [PubMed] [Google Scholar]
- 46.Satokata I, Benson G, Maas R: Sexually dimorphic sterility phenotypes in Hoxa10-deficient mice. Nature 1995, 374:460-463 [DOI] [PubMed] [Google Scholar]
- 47.Benson GV, Lim H, Paria BC, Satokata I, Dey SK, Maas RL: Mechanisms of reduced fertility in Hoxa-10 mutant mice: uterine homeosis and loss of maternal HOXA-10 expression. Development 1996, 122:2687-2696 [DOI] [PubMed] [Google Scholar]
- 48.Taylor HS, Arici A, Olive D, Igarashi P: HOXA10 is expressed in response to sex steroids at the time of implantation in the human endometrium. J Clin Invest 1998, 101:1379-1384 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Rots NY, Liu M, Anderson EC, Freedman LP: A differential screen for ligand-regulated genes: identification of HoxA10 as a target of vitamin D3 induction in myeloid leukemic cells. Mol Cell Biol 1998, 18:1911-1918 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Lalwani S, Bagot M, Troy P, Pando S, Taylor HS: Vitamin D regulates HOXA10 gene expression via an intronic vitamin D response element. Proc Soc Gynecol Invest 2000, 7:44 [Google Scholar]
- 51.Seely EW, Wood RJ, Brown EM, Graves SW: Lower serum ionized calcium and abnormal calciotropic hormone levels in preeclampsia. J Clin Endocrinol Metab 1992, 74:1436-1440 [DOI] [PubMed] [Google Scholar]
- 52.Lalau JD, Jans I, el Esper N, Bouillon R, Fournier A: Calcium metabolism, plasma parathyroid hormone, and calcitriol in transient hypertension of pregnancy. Am J Hypertens 1993, 6:522-527 [DOI] [PubMed] [Google Scholar]
- 53.Halhali A, Tovar AR, Torres N, Bourges H, Garabedian M, Larrea F: Preeclampsia is associated with low circulating levels of insulin-like growth factor I and 1,25-dihydroxyvitamin D in maternal and umbilical cord compartments. J Clin Endocrinol Metab 2000, 85:1828-1833 [DOI] [PubMed] [Google Scholar]
- 54.Rebut-Bonneton C, Demignon J: Effects of 1,25-dihydroxyvitamin D3 on in vitro lymphocyte reactions: arguments for a role at the maternofetal interface. Gynecol Obstet Invest 1991, 32:134-138 [DOI] [PubMed] [Google Scholar]
- 55.Panda DK, Miao D, Tremblay ML, Sirois J, Farookhi R, Hendy GN, Goltzman D: Targeted ablation of the 25-hydroxyvitamin D 1α-hydroxylase enzyme: evidence for skeletal, reproductive, and immune dysfunction. Proc Natl Acad Sci USA 2001, 98:7498-7503 [DOI] [PMC free article] [PubMed] [Google Scholar]