

Abstract
ADAM 12 (meltrin-α) is a member of the ADAM (a disintegrin and metalloprotease) family. ADAM 12 functions as an active metalloprotease, supports cell adhesion, and has been implicated in myoblast differentiation and fusion. Human ADAM 12 exists in two forms: the prototype membrane-anchored protein, ADAM 12-L, and a shorter secreted form, ADAM 12-S. Here we report the occurrence of adipocytes in the skeletal muscle of transgenic mice in which overexpression of either form is driven by the muscle creatine kinase promoter. Cells expressing a marker of early adipogenesis were apparent in the perivascular space in muscle tissue of 1- to 2-week-old transgenic mice whereas mature lipid-laden adipocytes were seen at 3 to 4 weeks. Moreover, female transgenics expressing ADAM 12-S exhibited increases in body weight, total body fat mass, abdominal fat mass, and herniation, but were normoglycemic and did not exhibit increased serum insulin, cholesterol, or triglycerides. Male transgenics were slightly overweight and also developed herniation but did not become obese. Transgenic mice expressing a truncated form of ADAM 12-S lacking the prodomain and the metalloprotease domain did not develop this adipogenic phenotype, suggesting a requirement for ADAM 12 protease activity. This is the first in vivo demonstration that an ADAM protease is involved in adipogenesis.
Obesity is a medical problem of increasing concern. 1-4 In Europe, 15 to 20% of the middle-aged population is clinically obese (defined by a body mass index greater than 30 kg/m2). As a result the health care service reports high morbidity and mortality from weight-related conditions such as cardiovascular disease and certain forms of cancer, and has seen a dramatic increase in type 2 diabetes mellitus. Body weight is determined by an interaction of genetic, environmental, and psychosocial factors acting through the physiological mediators of energy intake and expenditure. Thus, understanding the molecular mechanisms underlying obesity, a complex disorder of appetite regulation and energy metabolism, has become the object of an intense research effort.
Adipocytes arise from mesodermal stem cells that can also differentiate into other mesenchymal cells including muscle cells and osteoblasts. 5,6 Once committed to an adipocyte lineage, preadipocytes can either remain quiescent or proliferate and undergo differentiation into fully mature lipid-laden adipocytes. It has been shown that insulin, insulin-like growth factors, growth hormone, glucocorticoids, and catecholamines affect fat cell proliferation and differentiation in vivo and in vitro. 5,6 In addition, a role for the metalloprotease stromelysin-1 has recently been demonstrated in adipogenesis during mammary gland involution. 7 In vitro, adipocytes can also arise through transdifferentiation of myoblasts. 8,9
ADAM 12 is a member of the family of proteins known as ADAMs (a disintegrin and metalloprotease). The ADAMs constitute a large family of multidomain membrane-anchored proteins with cell adhesion and protease activities. 10-12 Human ADAM 12, which is located on chromosome 10q26, exists in two forms that arise from alternate splicing. Both forms, ADAM 12-L, the prototype membrane-anchored protein and ADAM 12-S, a shorter secreted form, 13 contain a prodomain, and metalloprotease, disintegrin, cysteine-rich, and epidermal growth factor-like domains. At the COOH-terminus, ADAM 12-L contains a transmembrane domain and a cytoplasmic tail, whereas ADAM 12-S is not membrane-anchored and contains a unique stretch of 33 amino acids.
Recently, we and others have shown that ADAM 12 (also called meltrin-α) is involved in myoblast fusion and myogenesis. 13-15 In the present study we have generated transgenic mice that we used to provide the first evidence that ADAM 12 is a proadipogenic factor and that its metalloprotease activity is required for this function.
Materials and Methods
Production of ADAM 12 Transgenic Mice
Two constructs were generated by inserting either full-length human ADAM 12-S cDNA (GenBank no. AF023477) or a truncated minigene lacking the prodomain and entire metalloprotease domain at the HindIII site of pCCLMCK-II. The minigene included a heterologous signal peptide (nucleotides 1560 to 2528, described in Gilpin et al 13 ). The plasmid pCCLMCK-II was used because it contains the muscle creatine kinase (MCK) promoter that confers striated muscle-specific expression of the transgenes. 16 The NotI and SalI sites of pCCLMCK-II were changed to SwaI sites to allow excision of vector-free transgene fragments containing the MCK promoter, ADAM 12-S coding sequence, and SV-40 splicing and polyadenylation sequences. Two vector-free ADAM 12-MCK fragments were generated; ASFM (ADAM 12-S full-length under the MCK promoter, 9 kb) and ASMM (ADAM 12-S minigene under the MCK promoter, 6.7 kb). These fragments were microinjected into the male pronuclei of fertilized zygotes isolated from superovulated donor mice, according to standard procedures. 17 Viable embryos were implanted into pseudopregnant recipients and allowed to develop to term. B6CBAF1 (C57BL/6J X CBA, F1) mice (M&B, Copenhagen, Denmark) were used as donors and recipients as well as for breeding transgenic hemizygous lines. All experiments involving animals were conducted according to the guidelines of the Animal Inspectorate, Denmark.
Genomic DNA derived from mouse tails was screened by PCR using two primers specific for human ADAM 12 (sense, 5′-CAGAATTCAAGGAGGCCGGATTCTGTG-3′; anti-sense, 5′-CCATCTAGATCAGATGAGTGTCAGTGA-3′) to identify transgenic mice. A 392-bp PCR product indicated a transgenic mouse. Southern blots were performed to verify the genotyping results, confirm a single integration site of the transgenes, and determine copy number. Northern blots were performed as described previously. 13
Generation of transgenic mice overexpressing full-length human ADAM 12-L, the membrane-anchored form containing a cytoplasmic tail (GenBank AF023476), will be described elsewhere (Kronqvist and colleagues, manuscript in preparation).
Western Blotting, Immunoprecipitation, and Enzyme-Linked Immunosorbent Assay
Polyclonal antisera, rb 122 and rb 132, that bind the cysteine-rich domain and the prodomain, respectively, and a rat monoclonal antibody (2F7) recognizing the disintegrin domain were used. 18 Cell lysates of Cos-7 cells transiently transfected with the relevant cDNA served as positive controls. Serum samples and muscle cell extracts were examined by Western blotting as described previously 19,20 or by immunoprecipitation using the Seize X Protein A Immunoprecipitation Kit (Pierce, Bie and Berntsen, Rødovre, Denmark).
The concentration of ADAM 12-S was determined by enzyme-linked immunosorbent assay. Rabbit immunoglobulin raised against recombinant human ADAM 12-S (rb 134) was diluted 10 μg/ml with phosphate-buffered saline (PBS) and used to coat Nunc MaxiSorp immunoplates overnight. Plates were blocked with Seablock (Pierce) and purified human ADAM 12-S at 0 to 100 ng/ml. 20 Serum samples were added to the wells followed by alkaline phosphatase-conjugated rabbit immunoglobulin against ADAM 12-S (prepared using activated alkaline phosphatase from Roche Molecular Biochemicals, Hvidovre, Denmark). The reaction was visualized using p-nitrophenylphosphate (Pierce) as the substrate, and the absorbance at 405 nm was measured after 30 to 60 minutes.
Histological Analysis and Immunohistochemistry
Tissue specimens were snap-frozen in liquid nitrogen or fixed with formalin and embedded in paraffin. Deparaffinized sections were stained with hematoxylin and eosin. Frozen sections or deparaffinized sections were incubated with affinity-purified antiserum to peroxisome proliferator activated receptor (PPAR)γ (N-20, SC-1984; Santa Cruz Biotechnology, Inc., AH Diagnostics, Aarhus, Denmark) or to PPARγ-2 (PAI-824; Affinity Bioreagents, Inc., Golden, CO) diluted 1:400 in PBS using the DAKO LSAB2 kit as recommended by the manufacturer (DAKO, Glostrup, Denmark). After the immunostaining reaction, selected sections were stained with 4,6-diamidino-2-phenylindole-methanol (1 μg/ml, Roche) for 1 minute at room temperature. Oil red O staining was performed as previously described. 21
Dual Emission X-ray Absorptiometry of Body Composition
The in vivo body fat mass was determined using a peripheral densitometer (PIXImus; Lunar Corporation, Scanex, Copenhagen, Denmark) according to the manufacturer’s instructions and as described. 22,23
In Vivo Matrigel Plug Assay
Three-month-old mice (transgenic and littermate controls) were injected on the forehead subcutaneously with 100 μl of growth factor-reduced Matrigel (Becton Dickinson Labware, Copenhagen, Denmark) as described. 24 Five weeks after injection mice were sacrificed and the amount of triglycerides in the plugs was quantitated using a commercially available kit (Sigma Biotech, Hørsholm, Denmark).
Serum Metabolite Levels in Transgenic and Wild-Type Mice
Blood was collected from mice after 16 hours of fasting. Serum glucose, triglyceride, cholesterol, and creatine kinase levels were measured using commercially available kits (Sigma). Insulin levels were determined by radioimmunoassay as described. 25
Statistical Analysis
All data are expressed as means ± SE. Two groups were compared with the unpaired Student’s t-test or the Mann-Whitney test. A value of P ≤ 0.05 was considered statistically significant.
Results
Generation of Transgenic Mice to Study the in Vivo Role of ADAM 12 in Adipogenesis
To study the potential role of ADAM 12 protease in vivo, we generated transgenic mice overexpressing ADAM 12 driven by the MCK promoter. Two constructs were used to express ADAM 12-S in mice. One construct (ASFM) encoded full-length human ADAM 12-S cDNA consisting of the prodomain, the metalloprotease, disintegrin, cysteine-rich, and epidermal growth factor-like domains, and the 33-amino acid COOH-tail, whereas the other (ASMM) encoded a truncated minigene lacking the prodomain and the metalloprotease domain (Figure 1A) ▶ . Several transgenic founders were obtained for each of the transgenes. Northern blot analysis confirmed that ADAM 12 mRNA was expressed only in the striated muscle of transgenic mice and that the level of expression in skeletal muscle was much higher than that observed in the heart (Figure 1B) ▶ . Four independent ASFM 7,9,19,26 and three ASMM 20,32,33 lines were selected for further analysis. The number of copies of the transgene in ASFM mice, as estimated by Southern blot analysis, was 2 (line 7), 2 (line 9), 4 (line 19), and 32 (line 26).
Figure 1.

Generation of ADAM 12 transgenic mice. A: Schematic representation of the ASFM and ASMM transgenes and the resulting proteins. ADAM 12-S is expressed as a 92-kd proform, which is cleaved by furin to yield the 68-kd mature protein and a 24-kd propeptide. B: Northern blot showing that the 3.6-kb ADAM 12-S mRNA is specifically expressed in skeletal muscle and to a lesser extent in the heart of transgenic mice (TG) (lanes 1 to 4) but not in their littermate controls (LC) (lanes 5 to 8).
Western blot analysis showed that ADAM 12-S protein was present in skeletal muscle (not shown) and in serum from transgenic mice but not from littermate controls (Figure 2) ▶ . Antibodies recognizing the disintegrin (mAb 2F7; Figure 2A ▶ , lanes 1 to 3) or the cysteine-rich domain (rb 122, not shown) of ADAM 12 detected the expected Mr 68,000 protein band representing the mature form of human full-length ADAM 12-S in which the prodomain is cleaved intracellularly by a furin-peptidase activity before secretion. We have recently demonstrated that the cleaved propeptide co-purifies with mature recombinant ADAM 12-S mature protein. 20 To determine whether the propeptide remains bound to ADAM 12-S in transgenic mice, Western blotting of serum samples with antisera to the cysteine-rich domain (rb 122) combined with antibodies to the prodomain (rb 132) was undertaken and revealed the presence of both the mature 68-kd protein and the 24-kd prodomain in transgenic mouse serum but not in serum from littermate controls (Figure 2B ▶ , lanes 1 and 2). Immunoprecipitation with an antiserum to the prodomain (rb132) followed by Western blotting with the monoclonal antibody (2F7) to the disintegrin domain further demonstrated that the prodomain, although cleaved from the mature protein, forms a complex with the mature ADAM 12 protein in serum from transgenic mice (Figure 2B ▶ , lane 3). In contrast, the ASMM mice expressing the truncated minigene form of ADAM 12-S lacking the prodomain and the metalloprotease domain exhibited a Mr 45,000 protein band in serum (Figure 2C) ▶ . As estimated by enzyme-linked immunosorbent assay, the concentration averaged 4.3 μg/ml for ASFM line 26 and 0.8 μg/ml for ASFM lines 7 and 19. Together these results demonstrate that the ASFM transgenic mice express ADAM 12-S protein that is potentially proteolytically active in muscle and serum, whereas ASMM transgenic mice express a minigene of ADAM 12-S lacking protease activity.
Figure 2.
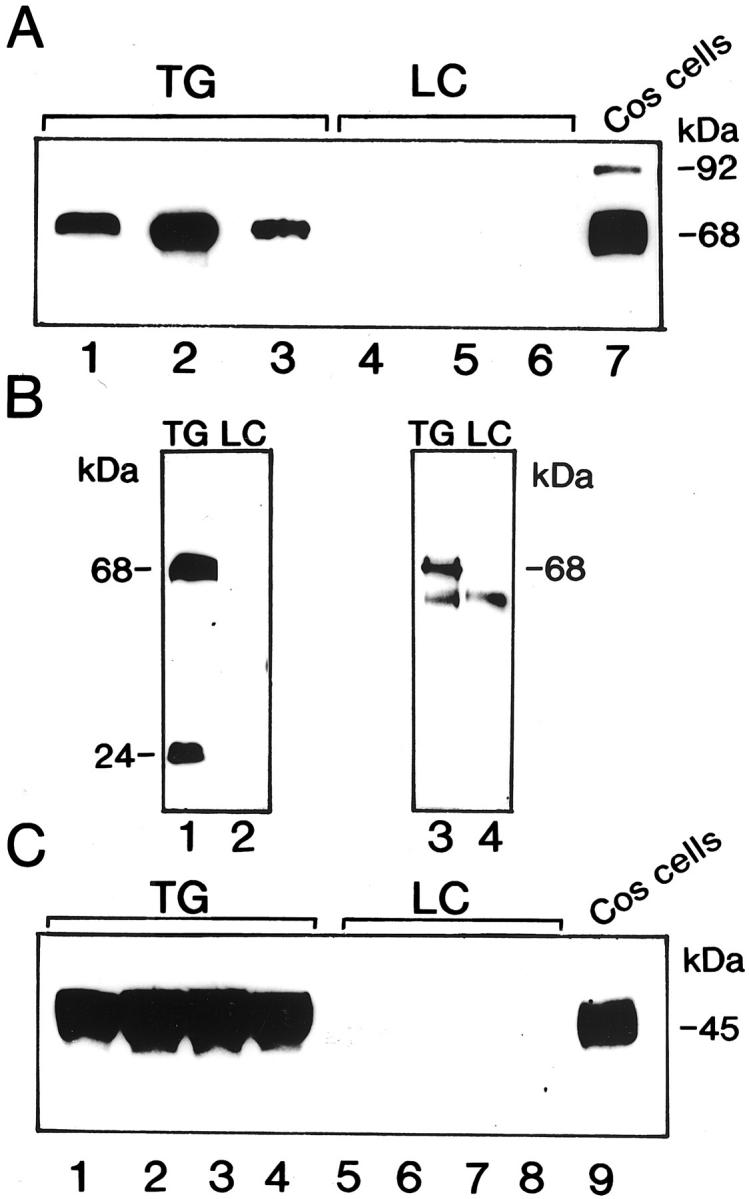
ADAM 12 protein expression in transgenic mice. A: Western blot of serum from three different ASFM transgenic mice (TG) with littermate controls (LC) shows a 68-kd protein representing the mature form of ADAM 12-S. Lane 7 shows Western blot of cell lysates of Cos-7 cells transfected with full-length ADAM 12-S that express the 92-kd proform and the 68-kd mature form. The 2F7 monoclonal antibody, which recognizes both forms of the protein, was used in this assay. B: Western blot of serum from ASFM mice using antiserum to the cysteine-rich domain (rb 122) and to the prodomain (rb 132) shows the 68-kd mature form and the 24-kd prodomain of ADAM 12-S in transgenic mice (lane 1) but not in littermate controls (lane 2). Immunoprecipitation with the prodomain antiserum (rb 132) followed by Western blotting with antiserum to the cysteine-rich domain (rb122) demonstrates that the 24-kd prodomain interacts with the mature form in transgenic mice (lane 3). The 50-kd band seen in both transgenic and littermate controls (lanes 3 and 4) most likely represents the Ig heavy chain. C: Western blot of serum from ASMM mice shows the 45-kd band in transgenic mice (lanes 1 and 2, line 32; lanes 3 and 4, line 33), but not in their respective littermate controls (lanes 5 to 8). Lane 9 shows the 45-kd protein in cell lysates from Cos-7 cells transfected with a truncated ADAM 12-S construct.
ADAM 12 Transgenic Mice Develop Abdominal Hernias and Increased Fat Tissue
ADAM 12-S transgenic mice did not differ from their littermate controls in overall health, fertility, or longevity. They exhibited no obvious abnormal phenotypic traits until 3 to 6 months of age. At this time lateral swellings developed in ASFM transgenic mice but not in their nontransgenic littermates (Figure 3) ▶ . Males and females of all four ASFM transgenic lines 7,9,19,26 developed these abdominal hernias. The protruding masses consisted of fat tissue (Figure 3B) ▶ and in most cases could be freely moved back and forth into the abdominal cavity. Microscopic analysis confirmed the presence of increased amounts of fat tissue. Individual adipocytes in the fat tissue of transgenic mice were similar in size and morphology to those from the littermate controls (not shown).
Figure 3.

Phenotype of ADAM 12-S transgenic mice. A 1-year-old female ASFM transgenic (TG) mouse with hernia formation (A) composed mainly of white adipose tissue (B), as indicated by arrows. The littermate controls (LC) did not exhibit this adipogenic phenotype (C and D).
The total body weight and the amount of abdominal fat tissue were measured to confirm the impression that female ASFM mice were more obese than littermate controls. As shown in Table 1 ▶ , 1-year-old female ASFM mice weighed significantly more than nontransgenic littermate controls (47.1 ± 1.4 g and 34.0 ± 0.8 g, respectively; P < 0.0001) and they contained significantly more abdominal fat (21.9% compared to 13.2%; P < 0.0001). Younger female ASFM mice (4 months old) also contained more abdominal fat than littermate controls (5.5% compared to 3.7%; P = 0.05). Among males a slight increase in total body weight between 1-year-old ASFM mice and littermate controls was observed (47.1 ± 1.0 g and 43.5 ± 1.2 g, respectively; P < 0.05), but there was no difference in the amount of abdominal fat (6.8% compared to 7.6%; P = 0.3). ASMM mice did not develop abdominal hernias (not shown), were not obese, and did not develop increased abdominal fat masses (Table 1) ▶ .
Table 1.
The Effect of ADAM 12-S on Abdominal Fat Mass, Total Body Weight, and Total Body Fat Mass
| Mice | Abdominal fat (g) | Total body weight (g) | Abdominal fat (%) | Total body fat mass (%) |
|---|---|---|---|---|
| ASFM (full-length ADAM12-S) | ||||
| Female/TG, 4 mo | 1.4 ± 0.2 | 25.5 ± 0.7 (n = 18) | *5.5 | 17.7 ± 1.6 (n = 18) |
| Female/LC, 4 mo | 0.9 ± 0.2 | 24.1 ± 0.7 (n = 17) | 3.7 | 15.9 ± 0.8 (n = 18) |
| Male/TG, 4 mo | 1.3 ± 0.2 | 32.3 ± 1.3 (n = 12) | 4.0 | 17.0 ± 1.7 (n = 10) |
| Male/LC, 4 mo | 0.9 ± 0.2 | 30.7 ± 0.9 (n = 11) | 2.9 | 16.4 ± 2.0 (n = 11) |
| Female/TG, 1 yr | 10.3 ± 0.7 | 47.1 ± 1.4 (n = 14) | **21.9 | ***48.5 ± 1.1 (n = 13) |
| Female/LC, 1 yr | 4.5 ± 0.4 | 34.0 ± 0.8 (n = 13) | 13.2 | 39.8 ± 1.1 (n = 12) |
| Male/TG, 1 yr | 3.2 ± 0.2 | 47.1 ± 1.0 (n = 8) | 6.8 | 36.2 ± 1.8 (n = 10) |
| Male/LC, 1 yr | 3.3 ± 0.4 | 43.5 ± 1.2 (n = 9) | 7.6 | 33.1 ± 1.4 (n = 8) |
| ASMM (truncated ADAM 12-S) | ||||
| Female/TG, 5 mo | 1.8 ± 0.3 | 27.0 ± 0.9 (n = 12) | 6.4 | 27.6 ± 2.7 (n = 14) |
| Female/LC, 5 mo | 1.2 ± 0.3 | 24.9 ± 0.7 (n = 8) | 4.8 | 24.5 ± 2.6 (n = 13) |
| Male/TG, 5 mo | 1.9 ± 0.2 | 38.1 ± 2.0 (n = 6) | 5.0 | 26.9 ± 1.2 (n = 11) |
| Male/LC, 5 mo | 1.8 ± 0.2 | 35.5 ± 1.3 (n = 10) | 5.1 | 26.2 ± 1.6 (n = 12) |
| Female/TG, 1 yr | 4.5 ± 0.5 | 34.6 ± 1.4 (n = 12) | 13.0 | 35.5 ± 2.1 (n = 12) |
| Female/LC, 1 yr | 4.8 ± 1.2 | 36.6 ± 3.0 (n = 6) | 13.1 | 33.9 ± 4.9 (n = 6) |
| Male/TG, 1 yr | 2.6 ± 0.4 | 42.9 ± 2.0 (n = 7) | 6.0 | 29.8 ± 1.1 (n = 8) |
| Male/LC, 1 yr | 2.0 ± 0.6 | 39.5 ± 3.4 (n = 5) | 5.1 | 29.2 ± 2.2 (n = 5) |
TG, transgenic; LC, littermate control.
Abdominal fat mass % was calculated as abdominal mass (g)/total body weight (g) × 100%. Total body fat mass % was measured by DXA scanning.
*P = 0.05;
**P < 0.0001;
***P < 0.0001 (Mann-Whitney test).
Dual emission X-ray absorptiometry was used to analyze total body fat mass in ADAM 12-S transgenic mice. Age- and sex-matched ASFM and ASMM transgenic mice were compared to their respective littermate controls. As shown in Table 1 ▶ , the total body fat mass (%) was significantly increased in 1-year-old female ASFM transgenic mice compared to littermate controls (48.5 ± 1.1% and 39.8 ± 1.1%, respectively; P < 0.0001). Younger (4-month-old) ASFM female mice did not exhibit increased total body fat mass nor did male ASFM mice at any age. Neither female nor male ASMM transgenic mice showed any increase in total body fat mass.
Because female ASFM mice showed increased abdominal fat mass and increased total body fat mass, we asked whether these mice exhibited changes in the physiological regulation of energy balance. Serum samples from fasting female mice were examined; no increase in serum glucose, insulin, triglyceride, or cholesterol levels relative to controls was detected in transgenic mice (not shown).
Adipogenesis in ADAM 12 Transgenic Skeletal Muscle
Inspection of ASFM transgenic mice revealed that increased amounts of fat tissue surrounded and infiltrated the muscle mass of the hindlimb (Figure 4A) ▶ . Consequently, skeletal muscles from the abdominal wall, forelimb, hindlimb, and diaphragm of transgenic and control mice were investigated in detail at the microscopic level. No apparent pathological changes of the ASFM muscle fibers were observed; however, dispersed groups of adipocytes were observed intermingled with the muscle fibers (Figure 4C) ▶ . This accumulation of adipocytes was not observed in littermate controls (Figure 4D) ▶ . No fat was seen inside the individual muscle fibers. We also undertook a morphological analysis of skeletal muscles from transgenic mice expressing full-length ADAM 12-L. ADAM 12-L transgenic mice, but not littermate controls, also contained groups of adipocytes in muscle (Figure 4, E and F) ▶ . In contrast, no accumulation of adipocytes was observed in ASMM mouse muscle (not shown).
Figure 4.
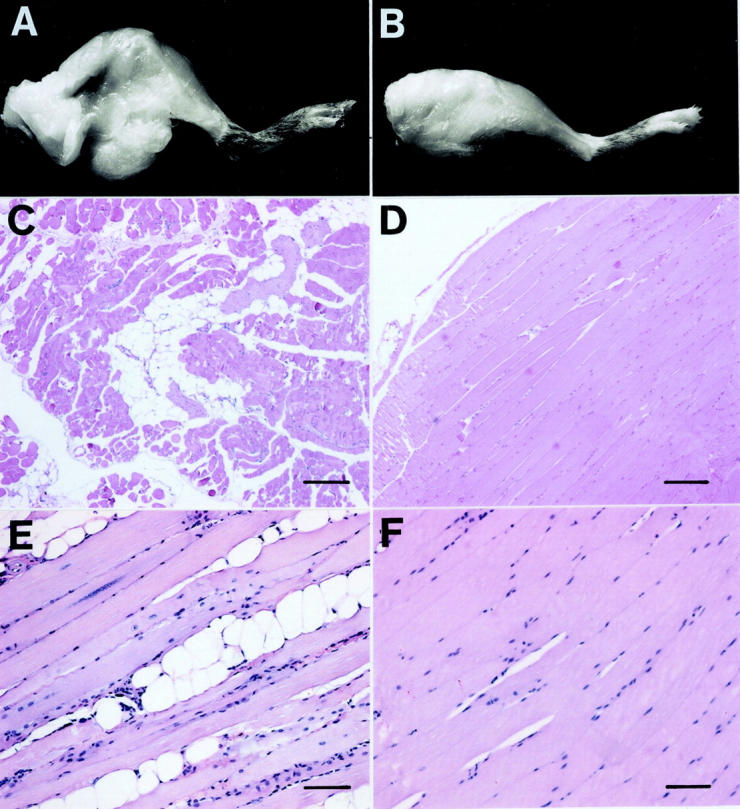
Accumulation of adipocytes in muscle tissue of ADAM 12 transgenic mice. Macroscopic demonstration of increased fat tissue surrounding the thigh muscle of ASFM mice (A) but not in littermate controls (B). C–F: H&E-stained sections of m. quadriceps femoris muscle from transgenic and control mice. C: Muscle from a 3-month-old ASFM transgenic mouse with adipocytes interspersed among muscle cells. D: Littermate control with no adipocyte accumulation. E: Muscle from 1-year-old transgenic mouse expressing ADAM 12-L with adipocytes interspersed among muscle cells. F: Littermate controls with no adipocyte accumulation. Scale bars: 160 μm (C and D); 80 μm (E and F).
To determine which cell types are affected by ADAM 12 overexpression, muscle specimens from mice of different ages were investigated to learn when and where adipogenesis could first be detected. It is thought that adipocyte differentiation is controlled by the PPAR and the CCAAT/enhancer binding protein (C/EBP) families of transcription factors. 5,6 PPARγ, a nuclear hormone receptor, is known to be induced early in the adipogenesis process. 5,6 We, therefore, immunostained using antibodies to PPARγ (not shown) and PPARγ-2 (Figure 5) ▶ as markers. Scattered PPARγ-positive fibroblastic cells could be detected within connective tissue of the perivascular space in the muscles of 1-week-old transgenic mice (not shown). In 2- to 3-week-old mice PPARγ-positive cells were readily detected in the muscle of transgenic mice but not in littermate controls (Figure 5 ▶ ; A to C). At this time small oil red O-positive droplets were observed in cells in the connective tissue (Figure 5D) ▶ indicating that intracellular fat had accumulated in these cells. Distinct adipocytes first became apparent around 3 to 4 weeks of age (Figure 5E) ▶ . Indicators of adipogenesis in the muscle connective tissue were consistently found near blood vessels (Figure 5 ▶ ; A, D, and E) suggesting that the adipocytes originate from mesenchymal progenitor/preadipocytes in the perivascular space.
Figure 5.
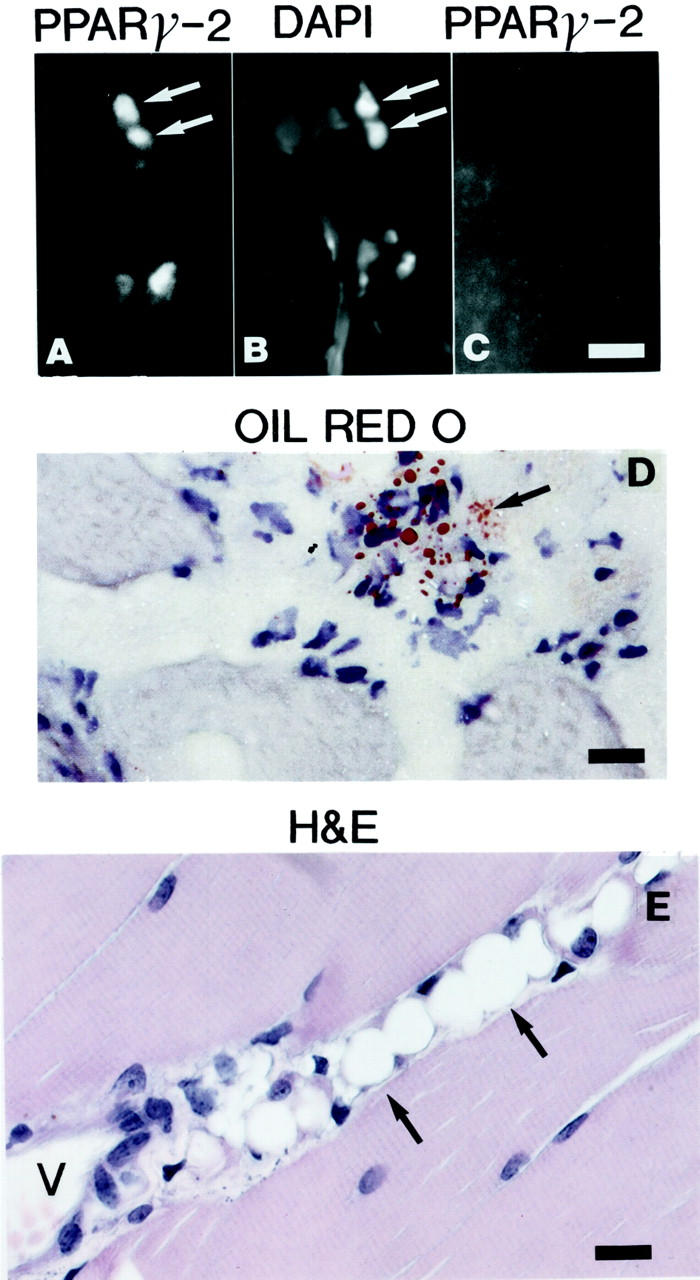
Early onset of adipogenesis in the perivascular space of ADAM 12 transgenic mice. A: Immunofluorescence staining using antibodies to PPARγ-2 on frozen sections of a 3-week-old ADAM 12-L transgenic mouse. Note the presence of scattered immunoreactive nuclei in the perivascular space. B: The same section was stained with 4,6-diamidino-2-phenylindole (DAPI) to stain the nuclei. C: Immunofluorescence staining using antibodies to PPARγ-2 on frozen sections of a 3-week-old littermate control mouse did not reveal any immunoreactive nuclei. D: Oil red O staining demonstrates small fat droplets in cells in frozen sections of a 3-week-old ADAM 12-L mouse. E: H&E-stained paraffin section of m. quadriceps femoris from a 3-week-old ASFM transgenic mouse shows several mature adipocytes. Arrows mark cells with PPARγ-2-immunoreactive nuclei and 4,6-diamidino-2-phenylindole-stained nuclei in A and B, respectively. Cells labeled by oil red O are marked with arrow in D. Mature adipocytes are marked with arrows in E. P, perivascular space; V, blood vessel. Scale bars: 60 μm (A–C); 40 μm (D and E).
ADAM 12 Stimulates Adipogenesis in an in Vivo Matrigel Plug Assay
The adipogenic potential of ADAM 12 was assessed using an in vivo Matrigel assay for adipogenesis as described. 24 Adipogenesis was demonstrated morphologically by the presence of adipocytes containing numerous small lipid droplets (not shown). In addition, triglyceride levels in the Matrigel plugs were significantly higher in the ADAM 12 transgenic mice than in the littermate controls (0.58 mg/pad ± 0.08; n = 8 compared to 0.31 mg/pad; n = 9; P < 0.05, Mann-Whitney test).
Discussion
The data presented here suggest that both the membrane-anchored (ADAM 12-L) and secreted (ADAM 12-S) forms of ADAM 12 can stimulate adipogenesis. Adipocytes accumulated in the striated muscle of transgenic mice overexpressing either full-length human ADAM 12-S or ADAM 12-L under the control of the mouse MCK promoter. In addition, adult female transgenic mice that overexpressed ADAM 12-S developed an increased amount of abdominal fat and showed an increased total body fat mass compared to sex- and age-matched nontransgenic littermate controls. Further support for ADAM 12 as a proadipogenic factor was demonstrated by in vivo Matrigel plug assays.
Immunochemical assays demonstrated that ADAM 12 protein was present in both the muscle and serum of mice expressing ADAM 12-S. The MCK promoter, which was used to drive muscle-specific expression of ADAM 12 transgenes in this study, is known to be highly tissue-specific and restricted in expression to striated skeletal and cardiac muscle. 16 It is likely that local overexpression of ADAM 12 in adult muscle results in de novo adipogenesis, whereas ADAM 12-S secreted into the serum by muscle cells has additional systemic effects leading to an increase in abdominal fat and overall body fat mass in female mice. ADAM 12-L female transgenic mice, which do not express ADAM 12 in serum and exhibited only local muscle-specific expression, showed no increase in abdominal fat. This finding supports the hypothesis of a local versus a systemic effect of ADAM 12.
It is not clear why the tendency to become obese with increased serum ADAM 12-S is restricted to females. Although the ADAM 12 transgenic female mice described here exhibit increased body fat, they appeared healthy, were normoglycemic, and did not exhibit increased levels of insulin, cholesterol, or triglycerides. In this respect, ADAM 12 mice are distinctly different from several mouse models exhibiting a much more severe phenotype with either an increase (ob/ob, db/db mice) or a decrease (A-ZIP/F-1, and SREBP-1c mice) in body fat mass that is associated with diabetes. 26
The occurrence of clusters of adipocytes dispersed among muscle fibers in the transgenic mice was a striking finding. Active adipogenesis in muscle connective tissue was first apparent in the perivascular spaces. Distinct PPARγ-positive fibroblastic cells and cells with small droplets of oil red O material were observed in muscle specimens from mice as young as 1 to 2 weeks of age. Clusters of mature adipocytes were seen in mice at 3 weeks and older. These results strongly suggest that ADAM 12 can function as a paracrine regulator of the differentiation of mesenchymal progenitor cells/preadipocytes present in the perivascular space. It should be noted, however, that adipocytes can arise through trans-differentiation of myoblasts in vitro. 8,9 Disruption of Wnt signaling resulted in trans-differentiation of cultured myoblasts into adipocytes. Furthermore, the ectopic expression of Wnt in 3T3-L1 stabilized free β-catenin and blocked adipogenic differentiation. 9 To test the idea that ADAM 12 protease could interfere with Wnt signaling, we added purified ADAM 12-S protease to 3T3-L1 preadipocytes cells in culture, however, we did not observe any obvious change in their capacity to differentiate into adipocytes (our unpublished results). It should also be noted that transdifferentiation was not reported in the study by Yagami-Hiromasa and colleagues 14 in which mouse ADAM 12-L was transfected into cultured myoblasts. These cumulative data suggests that the most likely targets of both ADAM 12-S and ADAM 12-L are cells of the adipocyte lineage located in the perivascular space.
One potentially important function of the ADAM family of proteins is the catalysis of so-called “ectodomain shedding” in which the extracellular domains of transmembrane proteins are proteolytically shed from the cell surface. Examples include tumor necrosis factor-α and heparin-binding epidermal growth factor-like growth factor (HB-epidermal growth factor). 13-15 ADAMs have also been implicated in α-secretase cleavage of the β-amyloid precursor protein in various cell lines 27,28 and most recently in the cleavage of cellular prion protein (PrPc), a glycosylphosphatidylinositol-anchored protein. 29 The finding that transgenic mice expressing ADAM 12 lacking protease activity (ASMM) did not develop fat cells in skeletal muscle and did not exhibit increased body fat mass suggests that ADAM 12 protease activity is required for the development of adipocytes seen in the full-length ADAM 12 transgenics. The next logical step is to determine the substrate of ADAM 12 protease activity. We have recently demonstrated that human ADAM 12-S can degrade IGFBP-3 and -5. 20,30 ADAM 12-S is present in human pregnancy serum (10 μg/ml) where IGFBP-3 is partially degraded, but not in nonpregnancy serum where IGFBP-3 is present in an intact form, suggesting that IGFBP-3 may serve as a physiological substrate of ADAM 12. 20,30 IGFBPs are ubiquitously expressed inserum, other biological fluids, and in several tissues where they bind IGF I and II and modulate their bioavailability. 31,32,33 Overexpression of IGFBP-1 in transgenic mice results in a phenotype consistent with partial inhibition of the growth-stimulatory effect of IGF, including impaired adipogenesis. 32 Overexpression of IGFBP-3 is likewise associated with growth retardation and changes in adipogenesis. 33 We analyzed whether the presence of ADAM 12-S in the serum of transgenic mice led to degradation of IGFBP-3; however, we did not find any consistent correlation (data not shown). Although this result does not preclude the possibility that the effect of ADAM 12 on adipogenesis may be mediated through degradation of IGFBP-3 in the local microenvironment, we believe it is equally likely that ADAM 12 metalloprotease activity activates adipogenesis through other unidentified mechanisms. ADAM 12 protease could release a growth factor by ectodomain shedding. Alternately, ADAM 12 could increase the bioavailability of adipogenic growth factors through remodeling of the extracellular microenvironment.
Accumulations of adipocytes are not normally present in skeletal muscle. Their occurrence is a distinct morphological hallmark of muscular dystrophy, in which dying muscle cells are gradually replaced by fibrosis and adipocytes. 34 The exact mechanism of the initiation of adipogenesis in these pathologically altered muscles is unknown. The results presented here lead us to speculate that ADAM 12, which we have previously shown to be up-regulated in regenerating muscle fibers in the mdx mice, 15 could be a factor that is involved in this process. We conclude that ADAM 12 transgenic mice provide a useful model for studying adipocyte biology and pathology. In particular these mice represent a novel in vivo model for studying the molecular mechanisms of the early stages of adipogenesis.
Ongoing studies in our laboratory are intended to further explore the physiological relevance of ADAM 12 in adipogenesis. We have demonstrated that ADAM 12 is expressed by preadipocytes as well as adipocytes in cell culture, suggesting that ADAM 12 might function as a paracrine/autocrine factor during adipocyte differentiation (manuscript in preparation). In addition to ADAM 12’s role as a protease, we have shown that ADAM 12 interacts with syndecans and β1 integrins on mesenchymal cells. 18 Thus, it is conceivable that ADAM 12 modulates adipocyte differentiation through multiple pathways. Elucidating these pathways is a primary focus of our current research.
Acknowledgments
We thank Brit Valentin for expert technical assistance; Jens Juul Holst for performing the RIA on insulin; Reinhard Fässler, Eva Engvall, and Peter Bang for helpful discussions; and Elise Lamar and Maryellen M. Daston for comments on the manuscripts.
Footnotes
Address reprint requests to Ulla M. Wewer, Institute of Molecular Pathology, University of Copenhagen, Frederik V’s vej 11, 2100 Copenhagen, Denmark. E-mail: ullaw@pai.ku.dk.
Supported by the Neye Foundation; the Danish Medical Research Council; the Danish Cancer Society; the Velux, Novo Nordisk, and Haensch Foundations; the Japanese Society for the promotion of Science; and by an European Union grant, Quality of Life and Management of Living Resources [contract no. QLG1-CT-1999-00870, designated “Genetic Resolution of Myopathies: European Cluster (Myocluster)].
N. K. and X. X. contributed equally to this work.
References
- 1.Bjorntorp P: Obesity. Lancet 1997, 350:423-426 [DOI] [PubMed] [Google Scholar]
- 2.Rosenbaum M, Leibel RL, Hirsch J: Obesity. N Engl J Med 1997, 337:396-407 [DOI] [PubMed] [Google Scholar]
- 3.Rosenbaum M, Leibel RL: The role of leptin in human physiology. N Engl J Med 1999, 341:913-915 [DOI] [PubMed] [Google Scholar]
- 4.Nature insights. Nature 2000, 404:632-67210766249 [Google Scholar]
- 5.Cornelius P, MacDougald OA, Lane MD: Regulation of adipocyte development. Annu Rev Nutr 1994, 14:99-129 [DOI] [PubMed] [Google Scholar]
- 6.Gregoire FM, Smas CM, Sul HS: Understanding adipocyte differentiation. Physiol Rev 1998, 78:783-809 [DOI] [PubMed] [Google Scholar]
- 7.Alexander CM, Selvarajan S, Mudgett J, Werb Z: Stromelysin-1 regulates adipogenesis during mammary gland involution. J Cell Biol 2001, 152:693-703 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hu E, Tontonoz P, Spiegelman BM: Transdifferentiation of myoblasts by the adipogenic transcription factors PPAR gamma and C/EBP alpha. Proc Natl Acad Sci USA 1995, 92:9856-9860 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ross SE, Hemati N, Longo KA, Bennett CN, Lucas PC, Erickson RL, MacDougald OA: Inhibition of adipogenesis by Wnt signaling. Science 2000, 289:950-953 [DOI] [PubMed] [Google Scholar]
- 10.Black RA, White JM: ADAMs: focus on the protease domain. Curr Opin Cell Biol 1998, 10:654-659 [DOI] [PubMed] [Google Scholar]
- 11.Blobel CP: Remarkable roles of proteolysis on and beyond the cell surface. Curr Opin Cell Biol 2000, 12:606-612 [DOI] [PubMed] [Google Scholar]
- 12.Primakoff P, Myles DG: The ADAM gene family. Surface proteins with adhesion and protease activity. Trends Genet 2000, 16:83-87 [DOI] [PubMed] [Google Scholar]
- 13.Gilpin BJ, Loechel F, Mattei MG, Engvall E, Albrechtsen R, Wewer UM: A novel, secreted form of human ADAM 12 (meltrin alpha) provokes myogenesis in vivo. J Biol Chem 1998, 273:157-166 [DOI] [PubMed] [Google Scholar]
- 14.Yagami-Hiromasa T, Sato T, Kurisaki T, Kamijo K, Nabeshima Y, Fujisawa-Sehara A: A metalloprotease-disintegrin participating in myoblast fusion. Nature 1995, 377:652-656 [DOI] [PubMed] [Google Scholar]
- 15.Galliano MF, Huet C, Frygelius J, Polgren A, Wewer UM, Engvall E: Binding of ADAM12, a marker of skeletal muscle regeneration, to the muscle-specific actin-binding protein, alpha-actinin-2, is required for myoblast fusion. J Biol Chem 2000, 275:13933-13939 [DOI] [PubMed] [Google Scholar]
- 16.Lee CC, Pons F, Jones PG, Bies RD, Schlang AM, Leger JJ, Caskey CT: Mdx transgenic mouse: restoration of recombinant dystrophin to the dystrophic muscle. Hum Gene Ther 1993, 4:273-281 [DOI] [PubMed] [Google Scholar]
- 17.Hogan B, Beddington R, Costantini F, Lacy E: Manipulating the Mouse Embryo: A Laboratory Manual. 1994. Cold Spring Harbor Laboratory Press, Cold Spring Harbor
- 18.Iba K, Albrechtsen R, Gilpin B, Frohlich C, Loechel F, Zolkiewska A, Ishiguro K, Kojima T, Liu W, Langford JK, Sanderson RD, Brakebusch C, Fassler R, Wewer UM: The cysteine-rich domain of human ADAM 12 supports cell adhesion through syndecans and triggers signaling events that lead to beta1 integrin-dependent cell spreading. J Cell Biol 2000, 149:1143-1156 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Loechel F, Gilpin BJ, Engvall E, Albrechtsen R, Wewer UM: Human ADAM 12 (meltrin alpha) is an active metalloprotease. J Biol Chem 1998, 273:16993-16997 [DOI] [PubMed] [Google Scholar]
- 20.Loechel F, Fox JW, Murphy G, Albrechtsen R, Wewer UM: ADAM 12-S cleaves IGFBP-3 and IGFBP-5 and is inhibited by TIMP-3. Biochem Biophys Res Commun 2000, 278:511-515 [DOI] [PubMed] [Google Scholar]
- 21.Ono M, Aratani Y, Kitagawa Y: Ascorbic acid phosphate stimulates type IV collagen synthesis and accelerates adipose conversion of 3T3–L1 cells. Exp Cell Res 1990, 187:309-314 [DOI] [PubMed] [Google Scholar]
- 22.Nagy TR, Clair AL: Precision and accuracy of dual-energy X-ray absorptiometry for determining in vivo body composition of mice. Obes Res 2000, 8:392-398 [DOI] [PubMed] [Google Scholar]
- 23.Iba K, Durkin M, Johnsen L, Hunziker E, Damgaard-Pedersen K, Zhang H, Engvall E, Albrechtsen R, Wewer UM: Mice with a targeted deletion of the tetranectin gene exhibit a spinal deformity. Mol Cell Biol 2001, 21:7817-7825 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kawaguchi N, Toriyama K, Nicodemou-Lena E, Inou K, Torii S, Kitagawa Y: De novo adipogenesis in mice at the site of injection of basement membrane and basic fibroblast growth factor. Proc Natl Acad Sci USA 1998, 95:1062-1066 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Brand CL, Jørgensen PN, Svendsen I, Kristensen JS, Knigge U, Warberg J, Holst JJ: The role of glucagon in the maintenance of euglycemia in the fed and the fasted state. Monoclonal antibody treatment as a tool in glucagon physiology in rats. Am J Physiol 1995, 269:E469-E477 [DOI] [PubMed] [Google Scholar]
- 26.Kahn BB, Flier JS: Obesity and insulin resistance. J Clin Invest 2000, 106:473-481 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.De Strooper B, Annaert W: Proteolytic processing and cell biological functions of the amyloid precursor protein. J Cell Sci 2000, 113:1857-1870 [DOI] [PubMed] [Google Scholar]
- 28.Lammich S, Kojro E, Postina R, Gilbert S, Pfeiffer R, Jasionowski M, Haass C, Fahrenholz F: Constitutive and regulated alpha-secretase cleavage of Alzheimer’s amyloid precursor protein by a disintegrin metalloprotease. Proc Natl Acad Sci USA 1999, 96:3922-3927 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Vincent B, Paitel E, Saftig P, Frobert Y, Hartmann D, De Strooper B, Grassi J, Lopez-Perez E, Checler F: The disintegrins ADAM10 and TACE contribute to the constitutive and phorbol ester-regulated normal cleavage of the cellular prion protein. J Biol Chem 2001, 276:37743-37746 [DOI] [PubMed] [Google Scholar]
- 30.Shi Z, Xu W, Loechel F, Wewer UM, Murphy LJ: ADAM 12, a disintegrin metalloprotease, interacts with insulin-like growth factor-binding protein-3. J Biol Chem 2000, 275:18574-18580 [DOI] [PubMed] [Google Scholar]
- 31.Ferry RJ, Jr, Katz LE, Grimberg A, Cohen P, Weinzimer SA: Cellular actions of insulin-like growth factor binding proteins. Horm Metab Res 1999, 31:192-202 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Rajkumar K, Modric T, Murphy LJ: Impaired adipogenesis in insulin-like growth factor binding protein-1 transgenic mice. J Endocrinol 1999, 162:457-465 [DOI] [PubMed] [Google Scholar]
- 33.Modric T, Silha JV, Shi Z, Gui Y, Suwanichkul A, Durham SK, Powell DR, Murphy LJ: Phenotypic manifestations of insulin-like growth factor-binding protein-3 overexpression in transgenic mice. Endocrinology 2001, 142:1958-1967 [DOI] [PubMed] [Google Scholar]
- 34.Dubowitz V: Muscle Disorders in Childhood. 1995. W.B. Saunders Company Ltd., London