

Abstract
The expression of inducible nitric oxide synthase (iNOS) in two different murine wound models was investigated. Animals were subjected to either full-thickness linear skin incision with subcutaneous implantation of sterile polyvinyl alcohol sponges, or to 1.5 × 1.5-cm dorsal skin excision. Reverse transcriptase-polymerase chain reaction detected iNOS mRNA in all cell samples retrieved from the sponges. Immunoblotting of lysates of inflammatory cells harvested from the sponges failed to detect iNOS protein, and immunohistochemistry of the incisional wound was mildly positive. Inflammatory cells of excisional wounds stained strongly positive for iNOS. Cutaneous wounds were found to be colonized with Staphylococcus aureus. The detection of iNOS in cells from sponges inoculated in vivo with heat-killed bacteria and the reduction of immunohistochemical signal for iNOS in excisional wounds of animals treated with antibiotics support a role of bacteria in the induction of iNOS in wounds. The expression of iNOS in excisional wounds requires interferon-γ and functional lymphocytes because interferon-γ knockout and SCID-Beige mice exhibited attenuated iNOS staining in excisional wounds. The expression of iNOS in the inflammatory cells of murine wounds is a response to bacterial colonization and not part of the normal repair process elicited by sterile tissue injury.
Previous work from this laboratory first described the expression of inducible nitric oxide synthase (iNOS) in a rat wound model. Using molecular and biochemical techniques, it was shown that iNOS is prominently expressed only early during wound repair, specifically the first 72 hours. Macrophages contributed the bulk of the iNOS protein to the wound, with the remainder found in polymorphonuclear leukocytes. 1 Based on such findings, it was proposed that NO generated by iNOS could participate in the regulation of the inflammatory phase of wound healing.
Other investigators have provided conflicting reports as to the role of iNOS-derived NO in the later stages of wound healing and the completion of the repair process. Yamasaki and colleagues 2 reported delayed healing in iNOS knockout (KO) mice that had undergone cutaneous excisional wounds, as well as in wild-type mice treated with an iNOS inhibitor. Additionally, they demonstrated that local adenoviral transfer of an iNOS expression vector normalized healing times in the iNOS-deficient animals. In contrast, Most and colleagues 3 found no difference between wild-type mice and iNOS KO mice in either the tensile strength of incisional wounds or the collagen content of subcutaneously implanted sterile polyvinyl alcohol (PVA) sponges.
Thus, although iNOS expression seems to be unnecessary for the development of normal tensile strength or for the accumulation of collagen in incisional wounds, it has been shown to be required for the normal healing of excisional wounds. Attempting to reconcile these differences, experiments were performed to compare the expression of iNOS in incisional and excisional wounds in mice at the molecular and histological levels. Results demonstrate that the iNOS protein is present in sebaceous glands and skeletal muscle in both wound models. Furthermore, iNOS is strongly expressed in the inflammatory cellular infiltrate of excisional skin wounds. In contrast, iNOS is minimally expressed in the cellular infiltrate of sterile cutaneous incisional wounds and absent in the inflammatory cellular infiltrate that congregates in implanted sterile PVA sponges. Evidence to be presented demonstrates that extracellular bacterial colonization of the wound is required for the expression of iNOS, and that interferon (IFN)-γ as well as functional lymphocytes are necessary for the full expression of iNOS in murine cutaneous wounds.
Materials and Methods
Animals
B6D2F1 and C.B.-Igh-1-GbmsTac-Prkdc-scid-Lyst−bg-N7 (SCID-Beige) mice (Taconic Laboratories, Germantown, NY) as well as C57BL/6-ifng−tmlTs (IFN-γ KO) mice (Jackson Laboratories, Bar Harbor, ME) at 6 to 8 weeks of age were used in these experiments. The mice were housed in a 12-hour light-dark cycle with access to food and water ad libitum. All animals were handled in compliance with the National Institutes of Health Guide for the care and use of laboratory animals. The Rhode Island Hospital/Brown Medical School Institutional Animal Care and Use Committee approved all experimental protocols.
Wound Model
Mice of both sexes were anesthetized with 45 mg/kg of sodium pentobarbital intraperitoneally (Abbott Laboratories, North Chicago, IL). The dorsum of each animal was clipped free of hair and prepped with 70% ethanol and 10% povidone-iodine solution (Purdue Frederick, Norwalk, CT) and then rinsed with 70% ethanol. For the incisional wound model, a 2-cm midline full-thickness skin incision was made in the dorsum of each mouse, through which bloodless subcutaneous pockets were developed. Four PVA sponges (PVA Unlimited, Warsaw, IN) 6 × 6 × 3 mm in size were implanted into the pockets through the skin incision, which was then closed with skin clips. All PVA sponges were sterile unless otherwise indicated.
At various intervals after wounding, the animals were sacrificed with CO2, the incisional wounds were reopened, and all of the sponges were retrieved. Single cell suspensions were obtained from the sponges by repeated compression as previously described. 4 Viable cell number was determined by trypan blue exclusion and found to be uniformly >90%. Cells were stored pelleted at −70°C until analysis. Bacteriological examination of the sponges confirmed their sterility at the time of harvest.
In the excisional wound model, an area of 1.5 × 1.5 cm of skin was resected from the dorsum of each mouse. The wounds were either allowed to remain open to air or were covered with a sterile bio-occlusive dressing (Tegaderm, 3 mol/L; Health Care, St. Paul, MN). The dressings were changed daily using sterile technique under pentobarbital anesthesia. The wounds were inspected for signs of infection, as well as swabbed for bacterial culture during dressing changes.
All wounds were harvested with an approximate 7-mm margin of normal skin and fixed in 10% formalin then stored at 4°C until histological and immunohistochemical examination. In the case of the excisional wound model, skeletal muscle underlying the wounds was harvested en bloc with the wound sample.
Bacteriology
Cultures demonstrated that the normal mouse skin flora includes Staphylococcus aureus, coagulase-negative Staphylococcus, Enterococcal species, and Proteus mirabilis. A polymicrobial stock in 10% fetal bovine serum-RPMI was developed from these cultures. The bacterial suspension was heat inactivated (80°C for 1 hour) and refrigerated until use. When indicated, the PVA sponges were soaked in this bacterial suspension before implantation.
The surface of skin wounds were swabbed daily with sterile cotton-tipped applicators, which were plated on both blood agar and LB agar plates (Lennox L Broth base; Life Technologies, Rockville, MD). The plates were incubated at 37°C ambient air and monitored daily for colony growth.
Immunohistochemical Analysis for iNOS
Formalin-fixed skin samples were imbedded in paraffin and cut into 5-μm sections. On use, these samples were deparaffinized in xylene and rehydrated with ethanol and phosphate-buffered saline (PBS). The slides were incubated in 0.6% H2O2 for 30 minutes at room temperature to block endogenous peroxidase. Sections were then blocked in 1% goat serum with 0.1% saponin in PBS for 1 hour. After washing with PBS, sections were incubated overnight with an anti-iNOS rabbit polyclonal IgG (BD Transduction Laboratories, San Diego, CA). The final concentration of antibody was 4 μg/ml in 1% normal goat serum, 0.1% saponin, in PBS. Normal rabbit IgG served as a control. The next day, sections were washed with PBS, and Vector Universal Elite ABC kit (PK-6200) and Vector VIP Peroxidase Substrate kit (SK-4600; Vector Laboratories, Burlingame, CA) were used according to manufacturer’s directions. The slides were then dehydrated with ethanol and xylene before permanent mounting and microscopic evaluation. RAW 264.7 murine macrophages that had been stimulated with murine recombinant IFN-γ (10 U/ml; Invitrogen Life Technologies, Carlsbad, CA) and lipopolysaccharide (1 μg/ml Escherichia coli O55:B5; Difco Laboratories, Detroit, MI) served as a positive control for iNOS staining. All slides were examined by a dermatopathologist (LRB) who had been blinded to the identity of each slide. The staining was graded as no staining, mild, moderate, or strong.
Semiquantitative Reverse Transcriptase-Polymerase Chain Reaction Analysis for iNOS mRNA
Total RNA was extracted from frozen cell pellets isolated from PVA sponges using the RNeasy Mini Kit (Qiagen, Valencia, CA) per the manufacturer’s instructions. RNA isolated from RAW 264.7 cells stimulated with IFN-γ and lipopolysaccharide, as described above, served as the positive control. One μg of total RNA from each sample was added to 1 μl of Dnase I (Invitrogen Life Technologies) for 15 minutes to eliminate genomic DNA. cDNA was synthesized using Moloney-murine leukemia virus (M-MLV) and random hexadeoxynucleotides included in the Bulk First Strand cDNA Synthesis kit (Amersham Biosciences, Piscataway, NJ) per the manufacturer’s instructions. Polymerase chain reaction was performed using 2 μl of cDNA sample added to TAQ Polymerase Kit (Qiagen). The iNOS primers were synthesized using sequences previously published by Yamasaki 2 with sense 5′-ATGGACCAGTATAAGGCAAGC-3′ and anti-sense 5′-GCTCTGGATGAGCCTATATTG-3′, with a predicted 429-bp product (Integrated DNA Technologies, Inc., Coralville, IA). Ambion’s Quantum RNA 18S Internal Standards kit (Ambion, Austin, TX) was used for target mRNA-to-18S ratios. A hot start technique was used at 92°C, then 30 cycles including 92°C for 1 minute to denature, 47°C 2 minutes to anneal, and 72°C for 3 minutes for extension. The products were stored at 4°C overnight. Samples then underwent electrophoresis on a 6% polyacrylamide gel at 100 V for 1 hour. The gel was stained with SYBR Green I solution (Bio Whittaker Molecular Applications, Rockland, ME) 1:10,000 in TBE for 30 minutes. Digital images were obtained using a Kodak DC 120 camera and software (Eastman-Kodak, Rochester, NY). NIH Image was used for densitometric analysis. The corrected relative value was calculated using the formula:
 |
Western Blot Analysis
Cells isolated from PVA sponges (5 × 106 cells/sample) were boiled with 2× ESB (125 mmol/L Tris pH 6.8, 4% SDS, 10% glycerol, 0.006% bromphenol blue, 2% β-mercaptoethanol). Aliquots corresponding to 1.5 × 106 cell equivalents were electrophoresed in 7.5%-SDS acrylamide gel for 65 minutes at 150 V. The proteins were transferred to a nitrocellulose membrane and blocked (5% dry milk in Tris buffer) overnight at 4°C. The membrane was incubated in primary antibody (anti-iNOS rabbit polyclonal IgG; BD Transduction Laboratories, San Diego, CA) diluted 1:1000 in blocking buffer for 1 hour. After washing, the membrane was immersed in a 1:2000 dilution of secondary antibody (horseradish peroxidase-conjugated anti-mouse IgG; Amersham Biosciences). The membrane then underwent enhanced chemiluminescent detection (ECL, Amersham Biosciences)
Treatment with Intramuscular Ceftriaxone
When so indicated, mice were treated with ceftriaxone 1.5 mg (n = 3) or 3.0 mg (n = 3) intramuscularly/animal/day (Roche Laboratories, Baxter Health Care, Deerfield, IL). The first dose was administered 24 hours before excisional wounding, and treatment was then continued daily for 3 days. Ceftriaxone was chosen based on published sensitivities of the bacterial strains cultured from murine pelts. Dosages were chosen to meet or exceed human neonatal dosing recommendations. 5
IFN-γ Assay
Wound fluid from sterile and bacteria-inoculated sponges harvested 1, 2, and 3 days after implantation was analyzed for IFN-γ by enzyme-linked immunosorbent assay using the kit OptEIA Mouse IFN-γ Set (BD Pharmingen, San Diego, CA) and following the manufacturer’s directions. The limit of detection for this assay is 30 pg/ml.
Amino Acid Analysis
Citrulline concentrations in plasma and extracellular fluid samples from PVA sponges were determined using high pressure liquid chromatography (Dionex BioLC; Dionex, Sunnyvale, CA), as previously reported. 6
Data Presentation
Experiments included at least three mice per experimental group. All experiments were repeated at least three times. Data shown are from a representative experiment.
Results
iNOS Protein Is Absent in the Cellular Infiltrate Isolated from Sterile PVA Sponges and Minimally Detectable in Incisional Wounds
Following the procedures described by Most 3 and as detailed in Materials and Methods, mice underwent a 2-cm full-thickness linear incision with subcutaneous implantation of sterile PVA sponges. Immunohistochemical analysis of the skin wounds harvested on day 3 after injury demonstrated positive staining for iNOS protein in the sebaceous glands and the striated muscle adjacent to, but not immediately underlying, the wound. The inflammatory cellular infiltrate present at the most superficial aspect of the wound stained moderately positive for iNOS; whereas inflammatory cells located deeper within the tissues only showed minimal specific staining (Figure 1) ▶ . The composition of the cellular infiltrate was identified by hematoxylin and eosin staining as mostly polymorphonuclear cells with interspersed macrophages. Cultures of the surface of incisional wounds were positive for S. aureus, whereas those of subcutaneously implanted PVA sponges confirmed their sterility (data not shown).
Figure 1.

Immunohistochemical analysis of sterile incisional wound on day 3 after injury. Asterisk indicates edge of wound. A: iNOS immunostain with long arrow indicating the inflammatory cellular infiltrate at the superficial aspect of the wound staining moderately positive for iNOS. Short arrow indicates inflammatory cellular infiltrate in the deeper tissue with diminished iNOS staining. B: H&E. C: IgG control. D–F: iNOS specimen. D: Strongly positive sebaceous gland staining. E: Moderately positive skeletal muscle staining. F: Inflammatory cellular infiltrate identified by the short arrow in A. Original magnifications: ×10 (A–C); ×60 (D–F).
Western blots of the cellular infiltrate obtained from the subcutaneously implanted PVA sponges harvested on days 1, 3, and 5 after wounding failed to demonstrate iNOS protein (Figure 2) ▶ . Moreover, the citrulline content of the extracellular wound fluid harvested from the PVA sponges remained at plasma levels (40 to 60 μmol/L) at all times (data not shown).
Figure 2.

Immunoblot of iNOS in protein derived from cellular infiltrate within subcutaneously implanted sponges. Sponges were either sterile (S) or inoculated with heat-killed bacteria (B) as described in Materials and Methods.
iNOS mRNA was detected in the cellular infiltrate on days 1, 2, 3, 5, and 10 after injury by semiquantitative reverse transcriptase-polymerase chain reaction (Figure 3) ▶ . Peak iNOS mRNA was detected on day 1, with a rapid decline on the subsequent days (day 1 corrected relative value = 21.8, nadir day 5 corrected relative value = 0.24).
Figure 3.

Semiquantitative reverse transcriptase-polymerase chain reaction for iNOS mRNA isolated from cellular infiltrate of subcutaneously implanted PVA sponges. 3+B represents infiltrate isolated on day 3 after injury from sponges inoculated with heat-killed bacterial suspension as described in Materials and Methods. Each sample represents pooled specimens of at least three animals. CRV, corrected relative value.
iNOS Antigen Is Present in Excisional Wounds
B6D2F1 mice underwent 1.5 × 1.5-cm full-thickness excisional wounds of the dorsal skin as described by Yamasaki. 2 Immunohistochemical staining of the excisional wounds harvested on day 3 after wound demonstrated strong iNOS protein expression in the cellular infiltrate, the sebaceous glands, as well as in the panniculum carnosus (Figure 4) ▶ . Muscle staining in excisional wounds showed a pattern similar to that found in incisional wounds; that is, negative staining of muscle immediately underlying the wound but positive in that just lateral to the wound. The epidermis was mildly positive. All staining was markedly attenuated on days 14 and 21 after injury (data not shown).
Figure 4.

Immunohistochemical analysis of excisional wound on day 3 after injury. Asterisk indicates edge of wound. A: iNOS immunostain with short arrow indicating the inflammatory cellular infiltrate with strong staining for iNOS. B: H&E. C: IgG control. D–F: iNOS specimen. D: Strongly positive sebaceous gland. E: Strongly positive skeletal muscle. F: Inflammatory cellular infiltrate identified by short arrow in A. Original magnifications: ×10 (A–C); ×60 (D–F).
Bacterial Colonization of Wounds
Skin wounds were evaluated daily for signs of infection. There was no visible purulence or erythema in any of the wounds 1 to 21 days after injury. As just mentioned, superficial cultures of both incisional and excisional wounds were positive for S. aureus by day 3 after injury in all animals. This was true even in those animals whose wounds were covered with a bio-occlusive dressing. Microscopic evaluation of the excisional wounds demonstrated visible colonies of staphylococcal species within the scab overlying the wound (Figure 5) ▶ .
Figure 5.

Excisional wound, arrows indicate colonies of staphylococcal species. H&E, original magnification, ×60.
Inoculation with Heat-Killed Bacteria Induces iNOS Expression in the Inflammatory Cellular Infiltrate in PVA Sponges
The role of bacteria in the induction of iNOS was tested by implanting PVA sponges inoculated with heat-killed bacteria subcutaneously in mice having undergone an incisional wound. Western blot analysis of the cellular infiltrate harvested from these sponges on days 1, 3, and 5 after wounding and reverse transcriptase-polymerase chain reaction analysis of day 3 infiltrate demonstrated iNOS expression (Figures 2 and 3) ▶ ▶ . Notably, inoculation led to a 24-fold increase in iNOS mRNA compared to that in cells from sterile sponges harvested on the same day [day 3 sterile CRV, 0.44 versus day 3 bacterial inoculated (3+B) CRV, 10.7]. Moreover, immunohistochemical analysis of the incisional wounds overlying the inoculated sponges revealed markedly enhanced iNOS staining of the inflammatory cells when compared to sterile incisional wounds (Figure 6) ▶ .
Figure 6.
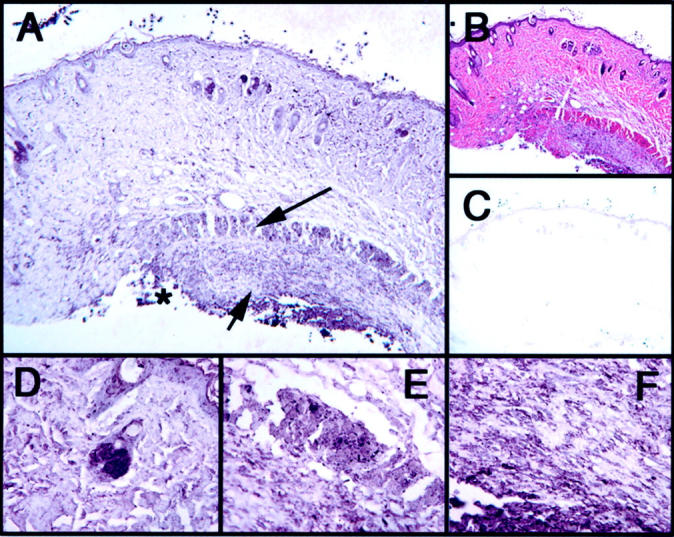
Immunohistochemical analysis of incisional wounds in animals implanted with PVA sponges inoculated with heat-killed bacteria on day 3 after injury. Asterisk indicates location of inoculated sponge implantation. A: iNOS immunostain with long arrow indicating skeletal muscle staining moderately positive for iNOS and short arrow indicating strongly positive inflammatory cellular infiltrate at the location of inoculated sponge insertion. B: H&E. C: IgG control. D–F: iNOS specimen. D: Strongly positive sebaceous gland. E: Skeletal muscle identified by long arrow. F: Inflammatory cellular infiltrate identified by short arrow in A. Original magnifications: ×10 (A–C); ×60 (D–F).
Antibiotics Attenuate iNOS Expression in Excisional Wounds
Additional mice undergoing excisional wounds were injected with ceftriaxone as described in Materials and Methods. Analysis of wounds harvested on day 3 after wounding from these animals revealed a markedly attenuated iNOS staining of the inflammatory infiltrate in all but the most superficial aspect of the wound when compared to untreated mice (Figure 7) ▶ . Furthermore, the muscular staining was now only minimally positive. The reduction in signal intensity was present with both doses of ceftriaxone.
Figure 7.

Immunohistochemical analysis of antibiotic-treated excisional wound on day 3 after injury. Asterisk indicates edge of wound. A: iNOS immunostain with long arrow indicating skeletal muscle immediately underlying the wound absent for iNOS and the short arrow indicating positive inflammatory cellular infiltrate adjacent to the wound. B: H&E. C: IgG control. D–F: iNOS specimen. D: Strongly positive sebaceous gland. E: Skeletal muscle identified by long arrow. F: Inflammatory cellular infiltrate identified by short arrow in A. Original magnifications: ×10 (A–C); ×60 (D–F).
Ceftriaxone failed to suppress iNOS induction or activity when added to the cultures of RAW 264.7 macrophages before activation with IFN-γ and lipopolysaccharide (data not shown). The attenuation of iNOS signal in vivo was, then, not because of any direct effect of the antibiotic on iNOS expression.
Attenuation of iNOS Signal in Excisional Wounds of IFN-γ Knockout and SCID-Beige Mice
To explore the mechanism for iNOS activation in excisional wounds, these wounds were inflicted on animals with targeted genetic mutations, namely IFN-γ KO and SCID-Beige mice. Although there was strong iNOS staining of the infiltrate at the superficial, exposed portion of the wound in each of the animals, specific staining for iNOS was markedly reduced in the infiltrating cells immediately adjacent to the wounds of both the IFN-γ KO and SCID-Beige mice (Figures 8 and 9 ▶ ▶ , respectively). Sebaceous glandular staining was not reduced in either the IFN-γ KO or the SCID-Beige mice. Finally, muscular staining was abrogated in the IFN-γ KO animals, but minimally reduced in the SCID-Beige mice. IFN-γ was not detected in wound fluid harvested from sterile or bacterial-inoculated sponges harvested 1 to 3 days after implantation.
Figure 8.

Immunohistochemical analysis of excisional wound in IFN-γ KO animal on day 3 after injury. Asterisk indicates edge of wound. A: iNOS immunostain with short arrow indicating inflammatory cellular infiltrate with minimal iNOS staining. B: H&E. C: IgG control . D–F: iNOS specimen. D: Strongly positive sebaceous gland. E: Mildly positive skeletal muscle. F: Inflammatory cellular infiltrate identified by short arrow in A. Original magnifications: ×10 (A–C); ×60 (D–F).
Figure 9.

Immunohistochemical analysis of excisional wound in SCID-Beige animal on day 3 after injury. Asterisk indicates edge of wound. A: iNOS immunostain with short arrow indicating the inflammatory cellular infiltrate with mildly positive iNOS staining. B: H&E. C: IgG control. D–F: iNOS specimen. D: Strongly positive sebaceous gland. E: Strongly positive skeletal muscle. F: Inflammatory cellular infiltrate identified by the short arrow in A. Original magnifications: ×10 (A–C); ×60 (D–F).
Discussion
As stated in the introduction, impetus to this study was given by contrasting findings from others regarding the effects of iNOS abrogation on the healing of excisional wounds versus incisional wounds with sponge implantation. Briefly, those reports indicated that iNOS deletion retarded healing of excisional wounds 2 but was without effect on the acquisition of tensile strength or the deposition of collagen in sterile incisional wounds with sponge implantation. 3 A key finding of this study is that bacterial colonization is required for iNOS expression by inflammatory cells in murine wounds. This requirement was demonstrated by the detection of iNOS protein in the inflammatory cells harvested from PVA sponges only when these were inoculated with heat-killed bacteria before implantation. Moreover, iNOS-positive inflammatory cells were found in both incisional and excisional wounds at their most superficial aspects, which were in turn shown to be culture-positive for S. aureus. Treatment of animals with colonized excisional wounds with systemic antibiotics significantly attenuated iNOS expression in all but those cells at the exposed portion of the wounds. It follows from these findings that the expression of iNOS by inflammatory cells in murine wounds is a response to bacterial colonization of the wounds.
A corollary of these findings is that iNOS protein is absent, and therefore not necessary for the repair of sterile wounds. In this connection, present findings provide a ready explanation for the observation that the deletion of the iNOS gene is without effect in the acquisition of tensile strength by sterile incisional wounds and for the accumulation of collagen in sterile implanted sponges in mice. This is so because iNOS protein is minimally expressed in the sterile incisional wound that was used for tensile strength testing and not at all in the PVA sponges used for the measurement of collagen accumulation by Most and colleagues. 3 The lack of iNOS activity in the sterile PVA sponge wound model was further emphasized by the fact that citrulline, a product of the metabolism of l-arginine by the nitric oxide synthases, remained at normal plasma concentrations in fluid harvested from sterile PVA sponges at all tested time points.
Although iNOS protein was not found in cells isolated from implanted sterile PVA sponges, iNOS mRNA was detectable by reverse transcriptase-polymerase chain reaction in all samples of sponge-infiltrating cells. This latter finding is in accordance with results by Yamasaki and colleagues 2 who demonstrated iNOS mRNA isolated from frozen excisional wound tissue. Present results demonstrate that the detection of iNOS mRNA cannot be equated quantitatively with the presence of iNOS protein in murine tissues, and suggests a role for posttranslational events. In this regard, although it is generally thought that iNOS is regulated mainly at the transcriptional level, 7 evidence has been provided for its posttranslational regulation. For example, TGF-β can decrease iNOS message stability, reduce iNOS mRNA translation, and increase the degradation of iNOS protein in macrophages. 8 Furthermore, IFN-γ has been shown to stabilize lipopolysaccharide-induced iNOS mRNA RAW 264.7 macrophage-like cells, determining a fivefold increase in the half-life of the iNOS mRNA. 9 Current results differ from those reported previously from this laboratory in rat incisional wounds. 1 In those wounds, iNOS protein and the metabolic products of the NOS reaction (eg, citrulline) were detectable in wound cells and wound fluids during the initial, inflammatory phase of the repair process. The difference in the expression of iNOS in wound cells of rats and mice is probably explained by the relative promiscuity of expression of iNOS in rat versus mouse inflammatory cells. More specifically, although nonstimulated rat peritoneal macrophages express iNOS in culture, their murine counterparts exhibit strict requirements for activating signals (eg, tumor necrosis factor-α, IFN-γ, lipopolysaccharide) for enzyme induction. 10
Lending further support to this conclusion, current findings demonstrate that IFN-γ is necessary for the full expression of iNOS in murine excisional wounds. These observations are in agreement with previous reports demonstrating markedly reduced iNOS expression in macrophages from mice deficient in IFN regulatory factor-1, a nuclear transcription activator of IFN-dependent gene expression. 11,12 Moreover, results obtained with SCID-Beige mice indicate that lymphocytes are the most likely source for IFN-γ in the wounds and define a requirement for such cells for the complete expression of iNOS in murine wounds.
No soluble IFN-γ was detected in wound fluids harvested from sterile and bacteria-inoculated subcutaneously implanted sponges. To some extent these results were predictable given that very few lymphocytes infiltrate the PVA sponge during the time frame here studied (data not shown). It is conceivable that IFN-γ is present in these fluids at concentrations below the limit of detection of the assay and that the cytokine exerts its effects in a paracrine manner. Alternatively, the diminished expression of iNOS in IFN-γ KO animals was observed in excisional wounds where cells from the skin-associated lymphoid tissue, unlikely to infiltrate subcutaneously implanted sponges, may contribute IFN-γ in a manner not duplicated in the sponge implantation model.
iNOS protein was detected immunohistochemically in tissue harvested from excisional wounds. Specific iNOS staining was found to be mild in the epidermis and strong in the sebaceous glands, the inflammatory cellular infiltrate, and in the striated skeletal muscle cells in these wounds. Because iNOS activity is required for normal healing in this model, 2 the expression of the enzyme in some or all of the positively stained cells and tissues is likely to be highly relevant to repair.
Present findings do not clarify the specific relevance of iNOS expression in each specific tissue to wound repair, but they do allow for cautious speculation when interpreted along with previously published reports. iNOS expression in the inflammatory cells and sebaceous glands of excisional wounds colonized by bacteria may contribute to the bactericidal capacity of the wound. In this regard, NO has been shown to be toxic for extracellular bacteria. Specifically, it has been shown that NO is produced at the skin surface 13 where it can be bactericidal against staphylococcal species. 14 iNOS expression by keratinocytes may contribute to enhanced closure rate of the wound because NO has been shown to promote keratinocyte proliferation and re-epithelialization. 15
It is somewhat harder to connect the strong expression of iNOS in skeletal muscle cells with a possible function in wound repair. Although it has been repeatedly shown that proinflammatory cytokines and lipopolysaccharide induce iNOS expression in muscle, 16-18 little is known regarding a functional role for NO in skeletal muscle. It has been shown that NO results in decreased contractile force of the skeletal muscle. 19,20 In cardiac myocytes, this effect has been shown to be related to iNOS-dependent depression of β-adrenergic-stimulated calcium release from the sarcoplasmic reticulum 21 as well as to decreased mitochondrial ATP production. 22 Along lines of previous findings from this laboratory on the impact of NO on glucose metabolism in other cells types, 23,24 nitric oxide production has also been shown to increase expression of the Glut 1 glucose transporter in rat L6 skeletal muscle cells. 25 Nonetheless, the potential connection between muscle iNOS and accelerated repair remains undefined. Taken together, these findings demonstrate that the expression of iNOS in the inflammatory cells of murine wounds is a response to bacterial colonization and not part of the inflammatory process in sterile wound repair.
Acknowledgments
We thank Jill Rose, Thomas Blaney, Liz Lavigne, and Brian LeBlanc for their instruction, assistance, and technical support; and Ginny Havanesian for the photography.
Footnotes
Address reprint requests to Dr. Jorge E. Albina, Department of Surgery, Rhode Island Hospital, 593 Eddy St., Providence, RI 02903. E-mail: jorge_albina@brown.edu.
Supported by the National Institute of General Medical Sciences (grant GM-42859), the Rhode Island Hospital (funds to the Division of Surgical Research, Department of Surgery), and by the Carter Family Charitable Trust (Armand D. Versaci Research Scholar in Surgical Sciences Award to E. J. M.).
References
- 1.Reichner JS, Meszaros AJ, Louis CA, Henry WL, Jr, Mastrofrancesco B, Martin BA, Albina JE: Molecular and metabolic evidence for the restricted expression of inducible nitric oxide synthase in healing wounds. Am J Pathol 1999, 154:1097-1104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Yamasaki K, Edington HD, McClosky C, Tzeng E, Lizonova A, Kovesdi I, Steed DL, Billiar TR: Reversal of impaired wound repair in iNOS-deficient mice by topical adenoviral-mediated iNOS gene transfer. J Clin Invest 1998, 101:967-971 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Most D, Efron DT, Shi HP, Tantry US, Barbul A: Healing in inducible nitric oxide synthase knockout mice. Surgery, in press [DOI] [PubMed]
- 4.Albina JE, Mills CD, Barbul A, Thirkill CE, Henry WL, Jr, Mastrofrancesco B, Caldwell MD: Arginine metabolism in wounds. Am J Physiol 1988, 254:E459-E467 [DOI] [PubMed] [Google Scholar]
- 5.The Sanford Guide to Antimicrobial Therapy, ed 31. Edited by D Gilbert, R Moellering, M Sande. Hyde Park, VT, Antimicrobial Therapy, Inc. Publishers, 2001, p 119
- 6.Albina JE, Abate JA, Mastrofrancesco B: Role of ornithine as a proline precursor in healing wounds. J Surg Res 1993, 55:97-102 [DOI] [PubMed] [Google Scholar]
- 7.Kleinert H, Boissel J-P, Schwarz PM, Förstermann U: Ignarro LJ eds. Nitric Oxide Biology and Pathobiology. 2000:p 108 Academic Press, San Diego
- 8.Vodovotz Y, Bogdan C, Paik J, Xie QW, Nathan C: Mechanisms of suppression of macrophage nitric oxide release by transforming growth factor beta. J Exp Med 1993, 178:605-613 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Weisz A, Oguchi S, Cicatiello L, Esumi H: Dual mechanism for the control of inducible-type NO synthase gene expression in macrophages during activation by interferon-gamma and bacterial lipopolysaccharide.Transcriptional and post-transcriptional regulation. J Biol Chem 1994, 269:8324-8333 [PubMed] [Google Scholar]
- 10.Albina JE: On the expression of nitric oxide synthase by human macrophages. Why no NO? J Leukoc Biol 1995, 58:643-649 [DOI] [PubMed] [Google Scholar]
- 11.Kamijo R, Harada H, Matasuyama T, Bosland M, Gerecitano J, Shapiro D, Le J, Koh SI, Kimura T, Green SJ, Mak TW, Taniguchi T, Vilcek J: Requirement for transcription factor IRF-1 in NO synthase induction in macrophages. Science 1994, 263:1612-1615 [DOI] [PubMed] [Google Scholar]
- 12.Shiraishi A, Dudler J, Lotz M: Role of IFN regulatory factor-1 in synovitis and nitric oxide production. J Immunology 1997, 159:3549-3554 [PubMed] [Google Scholar]
- 13.Weller R, Price RJ, Ormerod AD, Benjamin N, Leifert C: Antimicrobial effect of acidified nitrite on dermatophyte fungi, Candida and bacterial skin pathogens. J Appl Microbiol 2001, 90:648-652 [DOI] [PubMed] [Google Scholar]
- 14.Weller R, Pattullo S, Smith L, Golden M, Ormerod A, Benjamin N: Nitric oxide is generated on the skin surface by reduction of sweat nitrate. J Invest Dermatol 1996, 107:327-331 [DOI] [PubMed] [Google Scholar]
- 15.Stallmeyer B, Kampfer H, Kolb N, Pfeilshifter J, Frank S: The function of nitric oxide in wound repair: inhibition of inducible nitric oxide-synthase severely impairs wound reepithelialization. J Invest Dermatol 1999, 113:1090-1098 [DOI] [PubMed] [Google Scholar]
- 16.Ziolo MT, Katoh H, Bers DM: Expression of inducible nitric oxide synthase depresses beta-adrenergic-stimulated calcium release from the sarcoplasmic reticulum in intact ventricular myocytes. Circulation 2001, 104:2961-2966 [DOI] [PubMed] [Google Scholar]
- 17.Garcia YR, May JJ, Green AM, Krolick KA: Acetylcholine receptor-reactive antibody induces nitric oxide production by a rat skeletal muscle cell line: influence of cytokine environment. J Neuroimmunol 2001, 120:103-111 [DOI] [PubMed] [Google Scholar]
- 18.Adams V, Lenk K, Schubert A, Gielen S, Schuler G, Hambrecht R: Differentially expressed genes in L6 rat skeletal muscle myoblasts after incubation with inflammatory cytokines. Cytokine 2001, 13:342-348 [DOI] [PubMed] [Google Scholar]
- 19.el-Dwairi Q, Comtois A, Guo Y, Hussain SN: Endotoxin-induced skeletal muscle contractile dysfunction: contribution of nitric oxide synthases. Am J Physiol 1998, 274:C770-C779 [DOI] [PubMed] [Google Scholar]
- 20.Boczkowski J, Lanone S, Ungureanu-Longrois D, Danialou G, Fournier T, Aubier M: Induction of diaphragmatic nitric oxide synthase after endotoxin administration in rats: role on diaphragmatic contractile dysfunction. J Clin Invest 1996, 98:1550-1559 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ziolo MT, Katoh H, Bers DM: Expression of inducible nitric oxide synthase depresses beta-adrenergic-stimulated calcium release from the sarcoplasmic reticulum in intact ventricular myocytes. Circulation 2001, 104:2961-2966 [DOI] [PubMed] [Google Scholar]
- 22.Tatsumi T, Matoba S, Kawahara A, Keira N, Shiraishi J, Akashi K, Kobara M, Tanaka T, Katamura M, Nakagawa C, Ohta B, Shirayama T, Takeda K, Asayama J, Fliss H, Nakagawa M: Cytokine-induced nitric oxide production inhibits mitochondrial energy production and impairs contractile function in rat cardiac myocytes. J Am Coll Cardiol 2000, 35:1338-1346 [DOI] [PubMed] [Google Scholar]
- 23.Albina JE, Mastrofrancesco B, Reichner JS: Acyl phosphatase activity of NO-inhibited glyceraldehyde-3-phosphate dehydrogenase (GAPDH): a potential mechanism for uncoupling glycolysis from ATP generation in NO-producing cells. Biochem J 1999, 341:5-9 [PMC free article] [PubMed] [Google Scholar]
- 24.Albina JE, Mastrofrancesco B: Modulation of glucose metabolism in macrophages by products of nitric oxide synthase. Am J Physiol 1993, 264:C1594-C1599 [DOI] [PubMed] [Google Scholar]
- 25.Bedard S, Marcotte B, Marette A: Cytokines modulate glucose transport in skeletal muscle by inducing the expression of inducible nitric oxide synthase. Biochem J 1997, 325:487-493 [DOI] [PMC free article] [PubMed] [Google Scholar]