

Abstract
Syntactic priming is the facilitation of processing that occurs when a sentence has the same syntactic form as a preceding sentence. Such priming effects have been less consistently demonstrated in comprehension than in production, and those that have been reported have depended on the repetition of verbs across sentences. In an ERP experiment, participants read target sentences containing reduced-relative clauses. Each was preceded by a sentence that contained the same verb and either a reduced-relative or a main-clause construction. Reduced-relative primes elicited a larger positivity than did main-clause primes. Reduced-relative targets that were preceded by a main-clause prime were more positive than the same target sentences following a reduced-relative prime. In addition, syntactic priming effects were dissociated from effects of lexical repetition at the verb.
Language comprehension involves the activation of stored representations of different types of knowledge. An important question in research on language processing concerns the nature of these representations. What types of information are represented, and in what form? One way to address this question is through priming. Priming occurs when processing one stimulus facilitates processing of a subsequent stimulus. Structural priming can provide evidence regarding the representation of and access to linguistic knowledge because, when priming occurs, it demonstrates that some element of representation is shared across the prime and target stimuli. Repetition priming, in which a word is processed more quickly upon subsequent presentations than on the first, may result from residual activation of a lexical representation following its initial presentation (Morton, 1969). Evidence of repetition priming has been demonstrated in word lists (Scarborough, et al., 1977) and in sentence and discourse contexts (Sereno & Rayner, 1992; Traxler, et al., 2000).
Syntactic priming is the facilitation of processing that occurs when a sentence has the same syntactic form as a preceding sentence. Syntactic priming occurs reliably in studies of language production. In an early demonstration, participants tended to produce the syntactic form of a sentence they had just heard and repeated aloud (Bock, 1986). Subsequent studies have extended this basic finding to a broad range of participants, sentence types, and languages (Branigan, et al., 2000; Hartsuiker & Kolk, 1998a,b; Hartsuiker, et al., 2004; Loebell & Bock, 2003; Pickering & Branigan, 1998; Savage, et al., 2003).
Until recently, such priming effects had been less consistently demonstrated in on-line studies of language comprehension. However, several studies have now demonstrated priming effects that seem truly syntactic in nature (Frazier, et al., 1984; 2000). For example, Branigan, Pickering, and McLean (2005) had participants read sentence fragments and choose from two pictures the one that corresponded to the fragments. Participants were more likely on target trials to choose a given interpretation of a syntactically ambiguous phrase if they had read and seen a similar interpretation on the preceding prime trial.
Notably, while syntactic priming effects generally are affected by lexical overlap between primes and targets, priming can occur robustly in production, but not in comprehension, without such overlap (Cleland & Pickering, 2003; Pickering & Branigan, 1998). For example, Branigan and colleagues' (2005) syntactic priming effects were abolished when lexical overlap between prime and target stimuli was eliminated. Likewise, eye-tracking experiments have shown priming of target sentences for a variety of related prime types (Pickering & Traxler, 2004, 2006; Traxler & Pickering, 2005), but critically, only for prime-target pairs that contain the same verb form. Why is lexical overlap crucial to priming in comprehension? One possibility is that the kind of priming observed in comprehension is really semantic rather than syntactic. When primes and targets have overlapping lexical information, they also have greater semantic overlap relative to prime-target pairs that do not share words.
To test this possibility, we modified the experimental paradigm used by Pickering and Traxler (2004) to measure the effects of grammatical repetition on the comprehension of difficult reduced-relative (RR) clause sentences, as in (1).
(1) The defendant examined by the lawyer was unreliable
People initially interpret the defendant as the subject of the verb examined. They are “led down the garden-path” and interpret the sentence as a main-clause (MC) structure (as in the defendant examined the evidence). They must revise this interpretation when they encounter the disambiguating phrase by the lawyer, which forces the understanding of the verb as a past participle modifier of the defendant. Readers have difficulty with this kind of sentence, and that difficulty emerges when they first encounter the disambiguating phrase (Clifton, et al., 2003; Ferreira & Clifton, 1986; Trueswell, et al., 1994).
Pickering and Traxler (2004) monitored participants' eye movements while they read RR target sentences that had been preceded by either an RR or an MC prime sentence. In their first experiment, in which verbs were repeated across items, they found effects of syntactic priming on reading times: target-sentence processing was facilitated after a prime sentence with the same syntactic structure. In their second experiment, in which verbs were not repeated, no evidence of syntactic priming was observed.
The current experiment used event-related potentials (ERPs) to identify the source of priming for sentences like (1). ERPs are sensitive enough to detect differences in processing that may not consistently manifest behaviorally. Additionally, ERPs can be time-locked to different critical words, resulting in electrophysiological changes that are reliably associated with different aspects of processing. Most importantly, ERPs can dissociate effects of lexical repetition priming and syntactic priming.
Lexical repetition decreases the amplitude of the N400 component elicited by a word in a list, sentence or discourse context (Besson, et al., 1992, 1993; Paller & Kutas, 1992; Rugg, 1985, 1990). The N400 is a negative deflection in the ERP waveform that peaks approximately 400ms post-stimulus onset, and is maximal over posterior electrode sites (Kutas & Hillyard, 1980). A reduction of the amplitude of the N400 is found for words that can be easily integrated into the preceding context (e.g., Brown & Hagoort, 1993; Holcomb, 1993).
Syntactic processing has been shown to induce positive shifts in the ERP waveform. Processing of syntactic ambiguities has been found to affect the P600 component of the ERP (Osterhout & Holcomb, 1992), also known as the syntactic positive shift (SPS; Hagoort, et al., 1993). Generally, this broadly-distributed component onsets at approximately 500ms poststimulus onset; however, some studies have observed positive shifts with an onset latency as early as 200ms (Friederici, et al., 1998; 2001; see also Kutas et al., in press).1
We examined the contribution of repetition priming and syntactic priming during the comprehension of MC and RR sentences with repeated verbs, as shown in Table 1. To the extent that RR prime sentences are more difficult to parse than MC prime sentences, we should see a greater positivity to RR primes than to MC primes. Target sentences were always of the more difficult RR type, and were the same across comparison groups in the experiment; the only difference was the type of prime sentence (MC or RR) that preceded the target. If prime structure facilitates the syntactic processing of target structure, then participants' ERPs should show a reduced positivity when RR primes precede RR targets (relative to when those targets follow MC primes).
Table 1.
Example of the stimulus sets used in the experiment. Each participant saw one RR target, preceded by either an RR prime or an MC prime. Across participants, RR sentences could occur as either a prime or a target (dotted arrows). The ERPs were time-locked to the presentation of the verb (in italics) or the noun in postverbal phrase (underlined).
| RR Prime | MC Prime | (RR) Target |
|---|---|---|
| The speaker proposed by the group would work perfectly for the program. | The speaker proposed the solution to the group at the space program. | The manager proposed by the directors was a bitter old man. |
| The manager proposed by the directors was a bitter old man. | The manager proposed the changes to the bitter old man. | The speaker proposed by the group would work perfectly for the program. |
We also designed the experiment to allow a comparison of the processing of a given RR sentence when it appeared as a prime and as a target. That is, across participants, each RR sentence appeared as both a prime and as a target. When seen as a target, these sentences should benefit from the prior exposure to a sentence with the same verb and a similar syntactic structure: ERPs to the critical disambiguating noun as part of a target sentence, relative to a prime, should show a reduced positivity due to syntactic priming.
Finally, across prime and target sentences, we incorporated lexical repetition of the verb. By time-locking the ERPs to the first and second presentation of the verb, we could look for N400 effects of lexical repetition as dissociated from any observed effects of the repetition of syntactic structure.
Method
Participants
Participants were 30 right-handed native speakers of English recruited from the student population of UC Davis. None reported neurological impairment or medication.
Materials
The stimuli were adapted from those used by Pickering and Traxler (2004). Examples are presented in Table 1. Four versions of each of 80 experimental prime/target sentence pairs were constructed by manipulating two factors: type of prime sentence and position in a pair of a given RR sentence (prime or target). The two types of sentences (MC and RR) were exactly the same up to and including the first verb; disambiguation occurred at the phrase following the verb2. Target sentences were always of the RR construction, and always used the same verb as the paired prime sentence. Across lists, a given RR sentence occurred as both a prime sentence and a target sentence, thus serving as its own control.
Each participant read 80 prime sentences (40 MC primes and 40 RR primes) and 80 target sentences (all of which were RR form). In addition, participants read 190 filler sentences. Eighty of these were of the MC construction; as a result, participants read an equal number of MC and RR sentences (120 of each). The other 110 fillers were selected to include a wide variety of syntactic forms and semantic content.
Design and Procedure
Prime sentence type (MC or RR) was manipulated within-subjects; RR position (as prime or as target) was manipulated between-subjects. Fourteen filler sentences formed a practice block; the 160 experimental sentences were pseudo-randomly mixed with the remaining 176 filler sentences into 8 blocks of 42 sentences each. The prime and target sentences of a pair were presented sequentially; at least two filler sentences intervened between pairs. Each experimental and filler sentence was followed by a comprehension question. For the experimental sentences, a correct response to this question required the proper resolution of the MC and RR constructions. Four counterbalanced lists were created such that each experimental prime/target pair appeared in only one condition in a list, and across lists every pair occurred in all conditions. Across participants, each pair occurred equally often in each condition.
Each participant was tested individually in a dimly lit, electrically shielded, sound-attenuating booth. They were asked to silently read the sentences and to answer each question using a button box. Each trial began with a fixation cross (presented for 1000ms) in the center of the screen. Each sentence was presented one word at a time using rapid serial visual presentation with a duration of 300ms and an interstimulus interval of 200ms. The comprehension question appeared all at once 1000ms after the offset of the last word of each sentence, and remained on the screen until a response was recorded.
EEG Recording
EEG was recorded from 29 tin electrodes fitted in an elastic cap, referenced to the right mastoid; left mastoid was actively recorded for later re-referencing. Vertical eye movements were monitored by a sub-orbital electrode, and horizontal eye movements via left and right external canthus montages. Impedance was kept below 5 kOhm. Prior to off-line averaging, all single-trial waveforms were automatically screened for amplifier blocking, muscle artifacts, horizontal eye movements and blinks over epochs of 1200ms, starting 200ms before the onset of the critical words. For each participant, average ERPs were computed over artifact-free trials for critical words in all conditions. Off-line the waveforms were rereferenced to the algebraic average of both mastoids. The bandpass was 0.01 to 30 Hz at a sampling rate of 250 Hz.
Results
Behavioral results
Mean comprehension accuracy was .89 (SD=.05). Comprehension accuracy did not differ significantly by condition (RR primes: M=.88, SD=.09; MC primes: M=.88, SD=.08; targets after RR primes: M=.89, SD=.05; targets after MC primes: M=.89, SD=.07).
ERP results
ERP data were analyzed using repeated-measures analyses of variance (ANOVAs) performed on the mean amplitude of the ERPs to the critical words over the 29 electrode sites in 100ms overlapping (by 50ms) windows to determine the onset and offset of effects. The results of these analyses between 300ms and 800ms post-stimulus onset are presented in Table 23. For evaluating effects with more than one degree of freedom in the numerator, the Greenhouse-Geisser correction was used to compensate for inhomogeneous variances and covariances across treatment levels (Greenhouse & Geisser, 1959); the adjusted p-values are reported.
Table 2.
Results of ANOVAs in 100ms overlapping (by 50ms) time windows.
| 300-400ms | 350-450ms | 400-500ms | 450-550ms | 500-600ms | 550-650ms | 600-700ms | 650-750ms | 700-800ms | |
|---|---|---|---|---|---|---|---|---|---|
| Primes (RR vs. MC) | F(1,29)=2.89 | F(1,29)=5.94* | F(1,29)=7.22* | F(1,29)=6.73* | F(1,29)=2.37 | F(1,29)=2.03 | F(1,29)=8.02** | F(1,29)=9.58** | F(1,29)=8.71** |
| Targets (after RR vs. after MC) | F(1,29)=6.91* | F(1,29)=6.41* | F(1,29)=4.67* | F(1,29)=2.91 | F(1,29)=1.52 | F < 1 | F < 1 | F < 1 | F < 1 |
| RR Prime vs. Target after RR Prime | F(1,29)=6.24* | F(1,29)=6.87* | F(1,29)=6.37* | F(1,29)=5.53* | F(1,29)=4.12* | F(1,29)=3.00 | F(1,29)=4.05 | F(1,29)=2.78 | F(1,29)=1.21 |
| RR Prime vs. Target after MC Prime | F < 1 | F < 1 | F(1,29)=1.72 | F(1,29)=1.22 | F < 1 | F(1,29)=1.30 | F(1,29)=2.22 | F(1,29)=1.08 | F(1,29)=1.15 |
| Verb Repetition (Overall) | F < 1 | F(1,29)=1.38 | F(28,812)=2.47* | F(28,812)=4.18** | F < 1 | F < 1 | F < 1 | F < 1 | F < 1 |
| ant: F < 1 post: F(1,29)=12.28** | ant: F < 1 post: F(1,29)=14.23** | ||||||||
| Verb Repetition (MC Prime) | F < 1 | F(1,29)=2.18 | F(28,812)=2.70* | F(28,812)=4.69** | F(28,812)=2.38* | F < 1 | F(1,29)=1.65 | F(1,29)=3.67 | F(1,29)=2.16 |
| ant: F < 1 post: F(1,29)=9.14** | ant: F < 1 post: F(1,29)=9.60** | ant: F < 1 post: F(1,29)=3.29 | |||||||
| Verb Repetition (RR Prime) | F(1,29)=1.72 | F < 1 | F < 1 | F < 1 | F < 1 | F < 1 | F(28,812)=2.60* | F(28,812)=2.92* | F(28,812)=2.52* |
| ant: F < 1 post: F(1,29)=5.75* | ant: F < 1 post: F(1,29)=9.22** | ant: F < 1 post: F(1,29)=7.89** |
= p < .05, p(rep) > .88, ηp2 > .08
= p < .01, p(rep) > .95, ηp2> .13
Prime sentences
As shown in Figure 1 (left panel), ERPs to the critical noun in the postverbal phrase in RR prime sentences were more positive (from approximately 350-850ms post-stimulus onset) than those to the critical noun in MC prime sentences.
Figure 1.
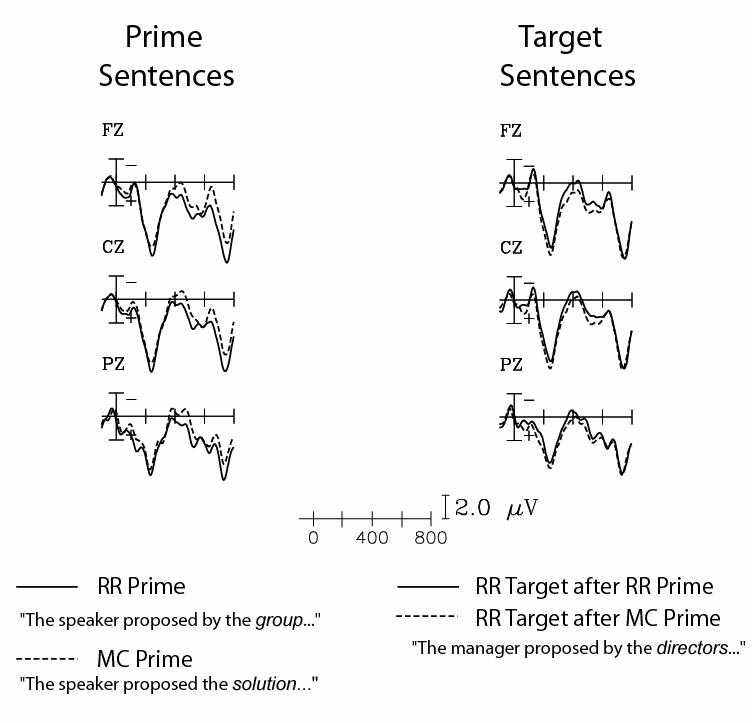
Prime and target sentences. For the primes (left), ERPs are shown to the noun following the verb in the disambiguating phrase of RR (solid) and MC (dashed) sentences. For the targets (right), ERPs are shown to the critical disambiguating noun of target (RR) sentences that followed RR (solid) or MC (dashed) prime sentences. The ERPs are grand averages across all participants, shown at midline (frontal: Fz, central: Cz, parietal: Pz) electrode sites.
Target sentences
The processing of the same RR target sentences differed as a function of the type of prime sentence (Figure 1, right panel). The response to the noun in an RR target sentence was more positive in the 300-500ms time window if its presentation had been preceded by an MC prime than if it had been preceded by an RR prime.
RR sentences as prime vs. target
Because of the counterbalancing of the RR stimuli, the ERPs to the sentences as primes could be compared to those to the sentences as targets. The ERPs to the critical noun when it appeared in an RR prime sentence and in a target sentence following an RR prime sentence are shown in Figure 2 (left panel). There was a significantly greater positivity when these words appeared in prime sentences, relative to targets sentences, during the 300-600ms time window.
Figure 2.
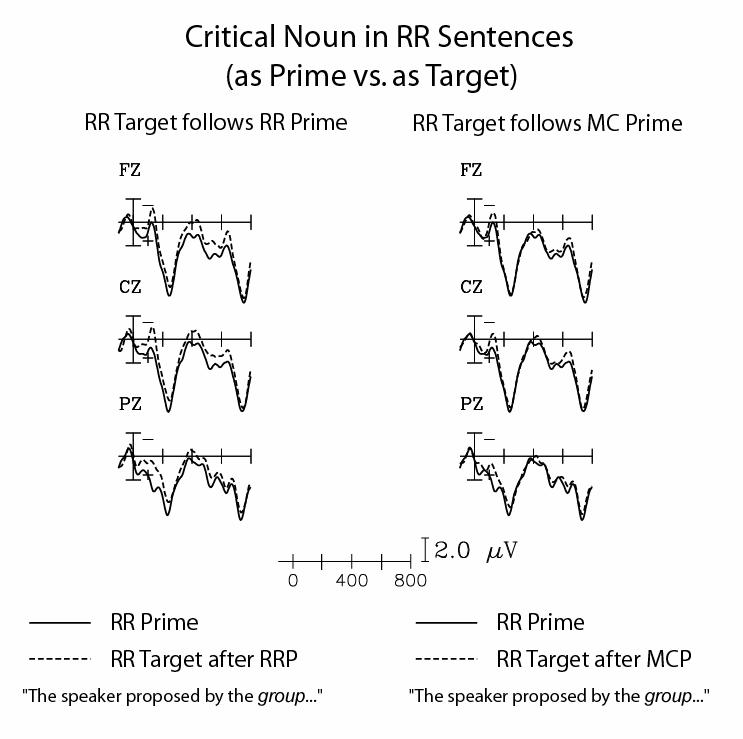
RR sentences: Primes vs. targets. The left panel shows ERPs to the noun following the verb in the disambiguating phrase of RR prime sentences (solid) and of target (RR) sentences that followed RR prime sentences (dashed). The right panel shows ERPs to the noun following the verb in the disambiguating phrase of RR prime sentences (solid) and of target (RR) sentences that followed MC prime sentences (dashed). The ERPs are grand averages across all participants, shown at midline (frontal: Fz, central: Cz, parietal: Pz) electrode sites.
In contrast, Figure 2 (right panel) shows the ERPs to the critical noun when it appeared in an RR prime sentence and in a target sentence following an MC prime. There was no significant difference in the ERPs to the words in the two conditions in any of the time windows tested.
Verb repetition
Figure 3 (left panel) shows the ERPs timelocked to the verb, on first presentation (in prime sentences) and on second presentation (in target sentences). At points between 400 and 550ms, we found an interaction between presentation (first vs. second) and electrode site. Subsequent comparisons indicated a significant reduction of the N400 to the verb on second presentations at posterior electrode sites, but not at anterior electrode sites.
Figure 3.
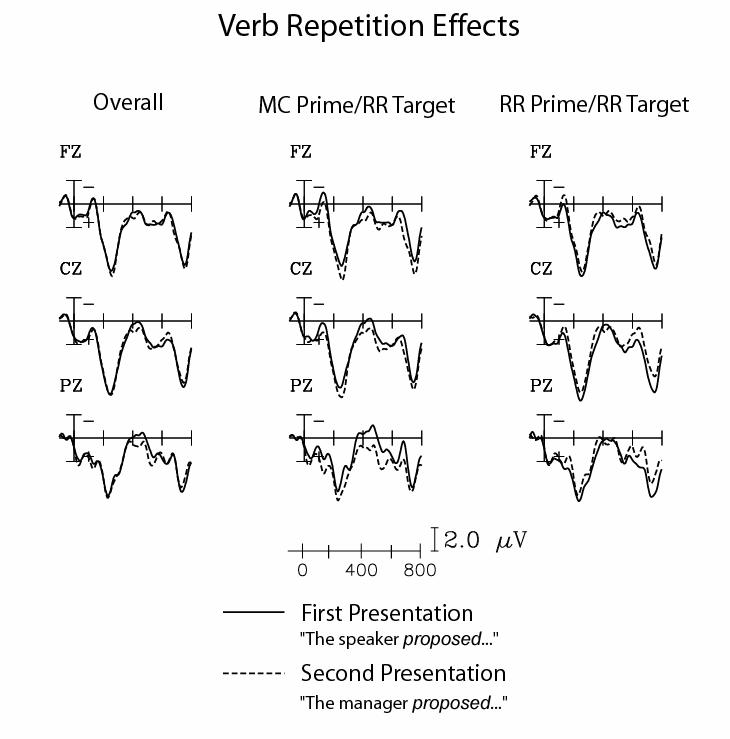
Verb repetition effects. The left panel shows ERPs to the verb before the disambiguating phrase of all prime sentences (first presentation; solid) and of all target sentences (second presentation; dashed). The middle panel shows ERPs to the verb before the disambiguating phrase of MC prime sentences (first presentation; solid) and of target sentences that followed MC prime sentences (second presentation; dashed). The right panel shows ERPs to the verb before the disambiguating phrase of RR prime sentences (first presentation; solid) and of target sentences that followed RR prime sentences (second presentation; dashed). The ERPs are grand averages across all participants, shown at midline (frontal: Fz, central: Cz, parietal: Pz) electrode sites.
We looked separately at the effect of verb repetition when the prime sentence was an MC or an RR. Figure 3 (middle panel) shows the response to the verb on first and second presentations, when the first presentation was an MC sentence. There was a clear repetition benefit for the verb on second presentations: a significant N400 repetition effect to the verb at posterior electrode sites was observed in this time window.
In contrast, Figure 3 (right panel) shows the response to the verb on first and second presentations when the first presentation was an RR sentence. In this case, there was no evidence of the N400 lexical repetition effect; instead, there was evidence of a decreased positivity at posterior electrodes to the verb on the second presentation in the 600-800ms time window.
Discussion
We used ERPs to examine lexical repetition and syntactic priming effects in language comprehension. Participants read target RR sentences that had been preceded by an MC or RR prime sentence that used the same verb. In the prime sentences, we found a greater positive shift to the critical disambiguating noun in the RR sentences than in the MC sentences. We believe that this shift resembles other positive deflections (P600/SPS) whose amplitude has been shown to increase in response to syntactic ambiguity and parsing demands, and suggests a greater difficulty in parsing RR primes relative to MC primes (mirroring prior behavioral and electrophysiological findings).
RR target sentences differed across conditions only as a function of the type of prime sentence (MC or RR) that came before. We found evidence of the differential processing of these target sentences, depending on the preceding prime: the ERP response to the critical noun was more positive if it occurred in a sentence that followed an MC prime than if it followed an RR prime. Given that the only difference between our conditions was the syntactic structure of the prime sentence, this effect likely represents an advantage in syntactic ambiguity resolution of the target sentences that was conferred by the experience of having recently read another sentence with the same syntactic structure (and the same verb).
Our experimental design allowed us to examine the reading of a given RR sentence as a prime or as a target. We looked at our target sentences as a function of whether they had been preceded by an MC or an RR prime sentence, and compared these results to the processing of RR sentences that appeared as primes (those which had not been preceded by another sentence with the same verb). The comparison of RR targets that had been preceded by MC primes to RR primes revealed no differences in the processing of the sentences in the two conditions. Having seen an MC prime sentence had no effect on the processing of a following target sentence that contained the same verb but a different syntactic structure; these targets were processed in the same way as an RR sentence that had no prime. Lexical overlap between primes and targets did not alone lead to differential syntactic processing of the target sentences (as might be the case if readers were developing processing strategies). On the other hand, comparison of RR targets that had been preceded by RR primes to RR primes revealed differences between these two conditions. The reduced positivity to the disambiguating noun in the target sentences relative to the same word in the prime sentences suggests that exposure to the RR prime had an effect on the processing of the RR target. Taken together, these results represent a demonstration of syntactic priming effects in comprehension.
These differences we observed were manifest in the ERP waveforms as a reduced positivity (to RR targets that followed RR primes, relative to those that followed MC primes; or to RR sentences that were presented as targets, relative to the same sentences when presented as primes). This reduced positivity may be interpreted as a reduction of the P600/SPS that has been previously implicated in syntactic reanalysis or ambiguity resolution. The latency of the positive shift in our experiment is similar to that observed for the P600. The distribution of this effect in our experiment was broad, with some suggestion of an anterior locus (although this was not reliable in statistical analyses). Anterior distributions of P600 effects have been demonstrated in the past, leading some researchers to suggest that such a distribution is more likely in cases of syntactic ambiguity resolution (as in our experiment) and a posterior distribution in cases of syntactic violations (Hagoort, et al., 1999; Kaan & Swaab, 2003). The most likely source of this reduced positivity is a decrease in the syntactic parsing demands of difficult RR sentences when these sentences were preceded by sentences of similar syntactic structure.
Our experimental design also allowed an examination of the contribution of lexical or semantic repetition to this syntactic priming effect. When we compared the processing of the verb in primes and in targets, we found a reduction in the amplitude of the N400 to the verb upon repetition. This effect was modulated, however, by the structure of the sentence in which the first presentation of the verb appeared. If participants first read the verb in a less taxing MC prime sentence, there was a large N400 benefit from repetition priming at the verb in the target sentences (before the difficult RR clause was encountered). This N400 repetition effect was not observed when participants first read the verb in an RR prime sentence. Instead, a comparison of the ERPs to the repeated verb in RR prime sentences and in target sentences that followed RR prime sentences revealed a decreased positivity to the verb on second presentations in a late time window (600-850 ms). This difference reflects a reduction in the amplitude of the Late Positive Component (LPC) that has been reported to repeated words in a sentence or discourse context (Van Petten, et al., 1991). Van Petten and colleagues (1991) offered a functional interpretation of the LPC as an index of the retrieval and updating demands imposed by words in varying contexts. The LPC difference in our experiment may therefore reflect the maintenance of some type of information in working memory from the first presentation of the verb to the second (resulting in a decrease in the retrieval and updating processing required on the second presentation). It is interesting to note in this case that the modulation of the LPC emerges in the absence of the typical N400 repetition effect. Some kind of lexical information about the verb remains active in working memory from the first presentation to the second (as indexed by the reduction in the LPC), but this information does not seem to ease the semantic integration of the verb with its sentence context on the second presentation (as suggested by the lack of the N400 repetition effect).
What kind of information might be maintained in working memory, leading to this LPC effect? It seems likely, given the evidence of syntactic priming described above, that this information relates to the syntactic properties of the verb, and its use in different syntactic constructions. Most theories assume that the lexical representation of a verb incorporates information about the types of constructions in which that verb participates (its subcategorization frames; Ford, Bresnan, & Kaplan, 1982). The lexical representation of a given verb will include information about all such possible constructions, but may be biased toward one construction in which the verb often appears. In this experiment, readers may have generally defaulted to the MC interpretation of the verbs. Having recently seen the verb in an RR construction, however, might have increased the activation of this subcategorization frame, leading to a quicker diagnosis of the possibility of this construction with this verb on subsequent presentations. The reduction in the amplitude of the LPC to this verb on second presentations might reflect the maintenance of this subcategorization information in working memory following the first presentation of the verb. This explanation could help to explain the necessity of verb lexical repetition in syntactic priming effects in comprehension.
In sum, we have demonstrated ERP evidence of syntactic priming effects in comprehension. These effects were dissociable from effects of the repetition of verbs across prime and target sentences and do not likely reflect semantic facilitation effects. The repetition of syntactic form led to changes in the electrophysiological response that are associated with a facilitation of syntactic analysis or ambiguity resolution. Changes in the processing of the verb in target sentences that followed RR prime sentences suggest that syntactic priming effects might be at least partially localized to changes in the representation of syntactic information at the verb.
Acknowledgments
This research was supported by NIMH grant R01-MH066271. MT is supported by the National Institutes of Health (NICHD #R01-40865) and the National Science Foundation (NSF#446618). We thank Martin Pickering for useful discussion, and Jocelyn Sy and Jennifer Morris for assistance with data collection.
Footnotes
Two earlier negative responses in the ERP have been observed to syntactic manipulations: the Early Left Anterior Negativity (ELAN; 100-300ms post-stimulus onset) and the Left Anterior Negativity (LAN; 300-500ms). Both are observed primarily in response to syntactic violations. Because the sentences we used were all syntactically well-formed, we did not expect to find ELAN or LAN effects in our experiment.
It was not possible (for semantic reasons) to match the critical nouns across the MC and RR prime sentences.
We found significant early effects (between 0-300ms) for one comparison, that of RR sentences as primes vs. as targets (Figure 3). It is possible that these differences are carry-over effects from the previous word; however, when we examined the ERPs timelocked to the previous word, we did not observe any differences in the waveforms. It seems likely, then, that the differences observed here are due to perceptual or attentional processing of the sentences on first vs. second presentation.
References
- Besson M, Kutas M, Van Petten C. An event-related potential (ERP) analysis of semantic congruity and repetition effects in sentences. Journal of Cognitive Neuroscience. 1992;4(2):132–149. doi: 10.1162/jocn.1992.4.2.132. [DOI] [PubMed] [Google Scholar]
- Besson M, Kutas M. The many facets of repetition: A cued-recall and event-related potential analysis of repeating words in same versus different sentence contexts. Journal of Experimental Psychology: Learning, Memory, & Cognition. 1993;19(5):1115–1133. doi: 10.1037//0278-7393.19.5.1115. [DOI] [PubMed] [Google Scholar]
- Bock JK. Syntactic persistence in language production. Cognitive Psychology. 1986;18:355–387. [Google Scholar]
- Branigan HP, Pickering MJ, McLean JF. Priming prepositional-phrase attachment during comprehension. Journal of Experimental Psychology: Learning, Memory, & Cognition. 2005;31(3):468–481. doi: 10.1037/0278-7393.31.3.468. [DOI] [PubMed] [Google Scholar]
- Branigan HP, Pickering MJ, Stewart AJ, McLean JF. Syntactic priming in spoken production: Linguistic and temporal interference. Memory & Cognition. 2000;28:1297–1302. doi: 10.3758/bf03211830. [DOI] [PubMed] [Google Scholar]
- Brown CM, Hagoort P. The processing nature of the N400: evidence from masked priming. Journal of Cognitive Neuroscience. 1993;5:34–44. doi: 10.1162/jocn.1993.5.1.34. [DOI] [PubMed] [Google Scholar]
- Cleland AA, Pickering MJ. The use of lexical and syntactic information in language production: Evidence from the priming of noun phrase structure. Journal of Memory & Language. 2003;49:214–230. [Google Scholar]
- Clifton C, Jr., Traxler MJ, Mohammed MT, Williams RS, Morris RK, Rayner K. The use of thematic role information in parsing: Syntactic processing autonomy revisited. Journal of Memory & Language. 2003;49:317–334. [Google Scholar]
- Ferreira F, Clifton C. The independence of syntactic processing. Journal of Memory & Language. 1986;25:75–87. [Google Scholar]
- Ford M, Bresnan J, Kaplan R. A competence based theory of syntactic closure. In: Bresnan J, editor. The mental representation of grammatical relations. MIT Press; Cambridge, MA: 1982. pp. 727–796. [Google Scholar]
- Frazier L, Taft L, Roeper T, Clifton C, Erlich K. Parallel structure: A source of facilitation in sentence comprehension. Memory & Cognition. 1984;12(5):421–430. doi: 10.3758/bf03198303. [DOI] [PubMed] [Google Scholar]
- Frazier L, Munn A, Clifton C., Jr. Processing coordinate structures. Journal of Psycholinguistic Research. 2000;29:343–371. doi: 10.1023/a:1005156427600. [DOI] [PubMed] [Google Scholar]
- Friederici AD. Diagnosis and reanalysis: Two processing steps the brain may differentiate. In: Fodor JD, Ferreira F, editors. Reanalysis in sentence processing. Kluwer Academic Publishers; Dordrecht: 1998. pp. 177–200. [Google Scholar]
- Friederici AD, Mecklinger A, Spencer KM, Steinhauer K, Donchin E. Syntactic parsing preferences and their on-line revisions: A spatio-temporal analysis of event-related brain potentials. Cognitive Brain Research. 2001;11:305–323. doi: 10.1016/s0926-6410(00)00065-3. [DOI] [PubMed] [Google Scholar]
- Greenhouse SW, Geisser S. On methods in the analysis of profile data. Psychometrika. 1959;24:95–112. [Google Scholar]
- Hagoort P, Brown C, Groothusen J. The syntactic positive shift as an ERP-measure of syntactic processing. Language & Cognitive Processes. 1993;8(4):439–483. [Google Scholar]
- Hagoort P, Brown C, Osterhout L. The neurocognition of syntactic processing. In: Brown C, Hagoort P, editors. Neurocognition of language. Oxford University Press; Oxford, UK: 1999. pp. 273–316. [Google Scholar]
- Hartsuiker RJ, Kolk HHJ. Syntactic persistence in Dutch. Language & Speech. 1998a;41:143–184. doi: 10.1177/002383099804100202. [DOI] [PubMed] [Google Scholar]
- Hartsuiker RJ, Kolk HHJ. Syntactic facilitation in agrammatic sentence production. Brain and Language. 1998b;62:221–254. doi: 10.1006/brln.1997.1905. [DOI] [PubMed] [Google Scholar]
- Hartsuiker RJ, Pickering MJ, Veltkamp E. Is syntax separate or shared between languages? Cross-linguistic syntactic priming in Spanish/English bilinguals. Psychological Science. 2004;15:409–414. doi: 10.1111/j.0956-7976.2004.00693.x. [DOI] [PubMed] [Google Scholar]
- Holcomb PJ. Semantic priming and stimulus degradation: Implications for the role of the N400 in language processing. Psychophysiology. 1993;30:47–61. doi: 10.1111/j.1469-8986.1993.tb03204.x. [DOI] [PubMed] [Google Scholar]
- Kaan E, Swaab TY. Repair, revision, and complexity in syntactic analysis: An electrophysiological differentiation. Journal of Cognitive Neuroscience. 2003;15(1):98–110. doi: 10.1162/089892903321107855. [DOI] [PubMed] [Google Scholar]
- Kutas M, Hillyard SA. Reading senseless sentences: Brain potentials reflect semantic incongruity. Science. 1980;207:203–205. doi: 10.1126/science.7350657. [DOI] [PubMed] [Google Scholar]
- Kutas M, Van Petten C, Kluender R. Psycholinguistics electrified II: ERP research on language processing. In: Traxler MJ, Gernsbacher MA, editors. The handbook of psycholinguistics. 2nd edition. Elsevier; San Diego, CA: in press. [Google Scholar]
- Loebell H, Bock K. Structural priming across languages. Linguistics. 2003;41:791–824. [Google Scholar]
- Morton J. Interaction of information in word recognition. Psychological Review. 1969;76:165–178. [Google Scholar]
- Osterhout L, Holcomb PJ. Event-related brain potentials elicited by syntactic anomaly. Journal of Memory and Language. 1992;31:785–806. [Google Scholar]
- Paller KA, Kutas M. Brain potentials during memory retrieval provide neurophysiological support for the distinction between conscious recollection and priming. Journal of Cognitive Neuroscience. 1992;4:375–391. doi: 10.1162/jocn.1992.4.4.375. [DOI] [PubMed] [Google Scholar]
- Pickering MJ, Branigan HP. The representation of verbs: Evidence from syntactic priming in language production. Journal of Memory and Language. 1998;39:633–651. [Google Scholar]
- Pickering MJ, Traxler MJ. Syntactic priming in comprehension; Paper presented at the 17th Annual CUNY Conference on Human Sentence Processing; College Park, MD. Mar, 2004. [Google Scholar]
- Pickering MJ, Traxler MJ. Syntactic priming in comprehension. 2006 Manuscript in preparation. [Google Scholar]
- Rugg MD. The effects of semantic priming and word repetition on event-related potentials. Psychophysiology. 1985;22(6):642–647. doi: 10.1111/j.1469-8986.1985.tb01661.x. [DOI] [PubMed] [Google Scholar]
- Rugg MD. Event-related brain potentials dissociate repetition effects of high- and low-frequency words. Memory & Cognition. 1990;18(4):367–379. doi: 10.3758/bf03197126. [DOI] [PubMed] [Google Scholar]
- Savage C, Lieven E, Theakston A, Tomasello M. Testing the abstractness of children's linguistic representations: Lexical and structural priming of syntactic constructions in young children. Developmental Science. 2003;6:557–567. doi: 10.1111/1467-7687.00312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scarborough DL, Cortese C, Scarborough HS. Frequency and repetition effects in lexical memory. Journal of Experimental Psychology: Human Perception & Performance. 1977;3:1–17. [Google Scholar]
- Sereno SC, Rayner K. Fast priming during eye fixations in reading. Journal of Experimental Psychology: Human Perception & Performance. 1992;18:173–184. doi: 10.1037//0096-1523.18.1.173. [DOI] [PubMed] [Google Scholar]
- Traxler MJ, Foss DJ, Seely RE, Kaup B, Morris RK. Priming in sentence processing: Intralexical spreading activation, schemas, and situation models. Journal of Psycholinguistic Research. 2000;29(6):581–594. doi: 10.1023/a:1026416225168. [DOI] [PubMed] [Google Scholar]
- Traxler MJ, Pickering MJ. The lexical nature of syntactic priming in comprehension; Paper presented at the 19th Annual CUNY Conference on Human Sentence Processing; Tucson, AZ. Mar, 2005. [Google Scholar]
- Trueswell JC, Tanenhaus MK, Garnsey SM. Semantic influences on parsing: Use of thematic role information in syntactic disambiguation. Journal of Memory & Language. 1994;33:285–318. [Google Scholar]
- Van Petten C, Kutas M, Kluender R, Mitchiner M, McIsaac H. Fractionating the word repetition effect with event-related potentials. Journal of Cognitive Neuroscience. 1991;3:131–150. doi: 10.1162/jocn.1991.3.2.131. [DOI] [PubMed] [Google Scholar]