

Abstract
Circadian rhythms in living organisms have long been attributed solely to a transcription-translation loop comprising a negative or positive feedback. The rhythms in cyanobacteria are known to be modulated by kaiC, kaiA and kaiB genes. It was recently shown, however, that their product proteins KaiC, KaiA and KaiB are sufficient to reconstitute the circadian rhythm in the phosphorylation level of KaiC in vitro. It has since been unclear why such an oscillatory behavior can occur in the absence of the apparent transcription-translation feedback. In the meantime, it has been reported that the monomer exchange between KaiC hexamers occurs in a phosphorylation-dependent manner, which suggests that the monomer shuffling is also involved in the circadian rhythm (H. Kageyama et al., Mol. Cell, 23, 161 (2006)). To further clarify the role of the monomer shuffling, we have performed a computational modeling of interactions among Kai proteins assuming the allosteric transition of KaiC hexamer as well as the monomer shuffling. The results show that the existence of both monomer shuffling and allosteric transition can synchronize the phosphorylation level of the KaiC hexamers, and stabilizes its oscillation.
Introduction
Biological activities of diverse organisms have circadian rhythms to fit to the day/night cycle of the environment [1]. The simplest organisms known to show circadian rhythms are cyanobacteria [2]. Three genes, kaiA, kaiB, and kaiC, play essential roles in the circadian clockwork of a cyanobacterium, Synechococcus elongatus PCC 7942 [3], in which the expressed protein, KaiC, represses the expression of kaiB and kaiC and thus constitutes a negative feedback loop [3], [4]. Various feedback regulations among clock genes were found in many other organisms, which led to a view that the transcription-translation oscillation is the source of circadian rhythmicity [5]. The generality of this view, however, is now being questioned at least for cyanobacteria [6]. Recent report of Nakajima et al. showed that the temperature-compensated robust circadian oscillation of KaiC phosphorylation can be reconstituted by incubating three Kai proteins, KaiA, KaiB, and KaiC, with ATP in vitro [7]. In this experiment, the reaction mixture contained neither mRNA nor DNA and the amount of protein molecules remained constant, which indicates that not the transcriptional regulation but Kai protein complexes are the pacemaker of cyanobacterial circadian clock [8].
As there is no transcription-translation feedback loop in this system, it is not clear at present why the number of phosphorylated KaiC can exhibit such a stable oscillation pattern. There have been a few preceding studies using theoretical models aiming at elucidating how this protein-only system can generate the robust circadian oscillation. With the limited experimental information, however, the theoretical model builders were forced to make some basic assumptions at their starting point [9]–[11]. Emberly and Wingreen assumed that KaiC monomers are shuffled during the period of phosphorylation of KaiC and that KaiC hexamers aggregate to form mesoscopic clusters during the period of KaiC dephosphorylation [9]. Kurosawa et al. assumed that both KaiA and KaiB have multiple states and regulate the KaiC phosphorylation process in multiple ways [10]. Mehra et al. assumed that the phosphorylated KaiC molecule catalyzes phosphorylation of other KaiC molecules [11]. Takigawa-Imamura et al. assumed that there are two sequential states of phosphorylated KaiC and that KaiA can bind to unphosphorylated KaiC and the second, but not to the first, state of phosphorylated KaiC [12]. Miyoshi et al. assumed that KaiB have two states which can be converted to each other with the help of KaiC hexamers at the different levels of phosphorylation [13]. These assumptions have not yet been confirmed experimentally and the consensus is not reached at the moment.
More recently, Kageyama et al. examined the detailed features of KaiC oscillation in vitro [14]. Their experimental data provided a new opportunity for testing the validity of theoretical models. Shuffling of monomers, for example, was shown to be most frequent during the dephosphorylation period [14], showing that the scenario of Emberly and Wingreen does not hold in its original form. In this paper, we exploit the data of Ref.14 to propose a consistent model of Kai protein complex dynamics. Our model presented here conforms to these experimental findings as well as other known structural and physicochemical information.
KaiA catalyzes the phosphorylation of KaiC [15] and KaiB attenuates the kinase activity of KaiA [16], where KaiA interacts with KaiC mostly in the KaiC phosphorylation phase and the KaiB activity is most evident in the KaiC dephosphorylation phase [14], [17]. We assume that the difference in binding affinities toward KaiA and KaiB can be attributed to the presence of two structural states of KaiC hexamer: In the phosphorylation phase KaiC has higher binding affinity to KaiA than to KaiB and in the dephosphorylation phase KaiC has higher binding affinity to KaiB than to KaiA. Such assumption of the concerted conformational change of monomers within homooligomers is well known to successfully describe the cooperativity in the ligand binding of hemoglobin [18] and chaperonin [19], both of which are correlated to changes in their quaternary structures [20], [21].
Another key assumption in our model is on the shuffling of KaiC monomers. KaiC is known to form a hexamer both in solution and crystals [22], [23], [14]. We assume that KaiC monomers are quickly interchanged when two hexamers interact, which is based on the experimental result [14] that the shuffling proceeds without accumulation of smaller or greater species than hexameric KaiC. The fact that the shuffling is most frequent in the dephosphorylation phase [14] indicates that intermonomer interactions are destabilized in the structural state in the dephosphorylation phase compared to that in the phosphorylation phase. Therefore, we denote the structural state which dominates in the dephosphorylation phase by relaxed (R) state, whereas the structural state which dominates in the phosphorylation phase is denoted by tense (T) state.
Each KaiC monomer has two phosphorylation sites at Ser431 and Thr432 [24]. Although the close distance (7.4Å) between these two sites in crystal structure [25] indicates that phosphorylation at these two sites should not be independent, the detailed mechanism of the correlation between the two sites is unknown at present. For simplicity, we do not distinguish two sites explicitly and assume that phosphorylation on one of the two sites is essential for the circadian oscillation in the present paper. When we also neglect the difference in the location of phosphorylated monomer within the hexamer, phosphorylation states of KaiC hexamer is specified by the total number of phosphorylated KaiC monomers within hexamer. Therefore there are 14 distinct states for KaiC hexamer denoted by Ti or Ri, where T or R denotes the structural state and i is the number of phosphorylated monomers from 0 to 6. Analyses of thus simplified model should provide a starting point to clarify the essential mechanism of the circadian oscillation produced by dynamically interacting proteins. The reaction scheme assumed in our model is shown in Fig. 1, where KaiA and KaiB are denoted by A and B, respectively. We consider Kai protein complexes, KaiC-KaiA (denoted by TiA or RiA), KaiC-KaiB (RiB), KaiC-KaiA-KaiB (TiAB or RiAB) and KaiC-KaiB-KaiA (RiBA).
Figure 1. Reaction scheme assumed in the model.

Association/dissociation, kinase/phosphatase reactions, and allosteric transitions are indicated by black arrows. Monomers of KaiC can be shuffled between T and R6 (red arrows) and between two Rs (pink area).
In the phosphorylation phase, KaiC hexamer should be mostly in TiA states with i increases from 0 to 6 as time proceeds due to the kinase activity of KaiA. In the dephosphorylation phase, KaiC hexamer should be mostly in RiB states with i decreases from 6 to 0 as time proceeds due to the effect of KaiB. We assume that the main routes of transition between T and R states are R0+A→T0A and T6+B→R6B. As these reactions constitute a closed loop, it may seem trivial for this system to show the oscillatory behavior of KaiC states. However, if we start from the initial condition in which the population of KaiC hexamer is localized at any single state, population rapidly diverges to multiple states by stochastic fluctuation of the timing of each step and eventually the system is dominated by the steady flow of the population.
We circumvent this problem by introducing the following two modes of shuffling: 1) Monomers can be exchanged between two KaiC hexamers in R states as Ri+Rj→Rk+Rl with i+j = k+l, and can not be exchanged between two KaiC hexamers in T states. This assumption is consistent with the observation [14] that the shuffling is most frequent during the dephosphorylation phase. 2) Since shuffling is most frequent for the period of several hours in the beginning of the dephosphorylation phase [14], we introduce the additional shuffling reactions involving the R6 state, which is populated at this period, and the T state as R6+Tj→Rk+Rl with 6+j = k+l. This assumption implies that R state is more accessible than T state for a newly formed hexamer after shuffling. With these assumptions, the oscillatory behavior can be brought about as follows. If the transition from T6 to R6 is slow compared to the phosphorylation/dephosphorylation, the population of KaiC initially accumulates at the T6 state. When the amount of hexamers in the T6 state exceeds some threshold, the R6 state begins to be populated. Subsequently, this further triggers the autocatalytic increase of KaiC in R states via the shuffling reaction R6+Tj→Rk+Rl in a relatively short time. Overall, this autocatalytic allosteric transition acts as a pacemaker which makes the KaiC population coherent enough to maintain the oscillatory behavior. Further details of reactions in Fig. 1 are explained in Model and Methods section and kinetic constants for those reactions are summarized in Supporting Information.
Reactions in vivo appear to be essentially same as those reconstituted in vitro [14]. Reactions in vivo, however, involve only small number of molecules in a single cell: Considering the cyanobactrium cell size of several µm, concentrations of KaiA, KaiB, and KaiC in the reaction mixture in Ref.14 (1.2 µM, 3.5 µM, and 3.5 µM of monomers, respectively) correspond to about 104 to 105 molecules of each Kai monomer in a cell. In order to check whether such smallness in number of molecules affects the reactions, we simulate the stochastic dynamics of reactions in the model of Fig. 1 by using the Gillespie algorithm [26]. The reaction mixture confined in a µm vessel is simulated with 10000 dimers of KaiA, 30000 dimers of KaiB, and 10000 hexamers of KaiC, which corresponds to 1:3:3 ratio of monomers. Since the oscillation should be less stable in a more noisy system with smaller numbers of molecules, the stable persistent oscillation of reactions observed in this stochastic simulation will assure both the stability of oscillation in a cell of µm size and the stability of oscillation in vitro with the macroscopic number of molecules.
Results
As shown in Fig. 2, the model exhibits a stable persistent oscillation in the level of KaiC phosphorylation, p(t) = Σi i(Xi R+Xi T)/6X, where X = ) = Σi (Xi R+Xi T), Xi R = [Ri]+[RiA]+[RiB]+[RiAB]+[RiBA], Xi T = [Ti]+[TiA]+[TiAB], and […] denotes the concentration of each of the chemical species. With the parameters used (summarized in Supplementary Table S1), the phosphorylation level of KaiC oscillates between 0.2 and 0.8 and the period length is about 22 hours. These findings are consistent with the experimental observations (Nakajima et al Science, Kageyama et al Mol Cell). This oscillatory behavior is necessarily accompanied by the autocatalytic increase of KaiC population in R states described above. To illustrate how the mechanism works, typical examples of distributions of KaiC states at time t, P(Ri,t) = Xi R/X and P(Ti,t) = Xi T/X, are plotted in Fig. 3. When the rate of transition from T6 to R6 is 100 times faster than its standard value, KaiC hexamers are not caught at the T6 state and leak to the R6 state, which promotes active monomer shuffling between T and R. Through this active shuffling the population spreads over the R states, which suppresses the oscillation as shown in Fig. 3b. The oscillation is also suppressed when the shuffling rates are low. With the lower shuffling rates, the small amount of KaiC hexamers appeared at the R6 state do not trigger further increase at the R states via reactions R6+Tj→Rk+Rl (data not shown).
Figure 2. The simulated oscillation in the phosphorylation level p(t).

Figure 3. Typical examples of distributions of KaiC states, P (R i,t ) = Xi R/X and P (T i , t ) = Xi T/ X .

(a) Distributions at t = 18 hours (black line) and t = 22 hours (red line) calculated with the standard parameter set consistent with the autocatalytic increase of KaiC population in R states. (b) Distributions at t = 10 hours (black line) and t = 14 hours (red line) calculated with the transition rate from T6 to R6 increased 100-fold from its standard value.
The model also predicts the dependence of the oscillation on various rate constants. For example, kp TA[TiA], which is the rate of phosphorylation from TiA to Ti +1A, should affect the period length of oscillation, and [TiA] is strongly dependent on the binding rate kb T-A of A to Ti. (See Model and Methods for the precise definition of these constants.) In Fig. 4a, the oscillation period length is plotted against kp TA and kb T-A. The persistent oscillation exists in the parameter region of kp TA kb T-A ∼8.6×10−5sec−2, and in the region of kb T-A∼5 sec−1. The oscillation should also depend on kdp RB[Ri +1B], which is the rate of dephosphorylation from Ri +1B to RiB. In Fig. 4b, the oscillation period is plotted against kdp RB and kb R-B, where kb R-B is the rate constant of binding of B to Ri. The stable oscillation is found in the parameter region of kdp RB kb R-B∼1.1×10−4 sec−2. In wide areas of Figs. 4a and 4b, the oscillation period length does not vary much as far as the parameters are consistent with the autocatalytic increase of KaiC population in R states. The period varies, however, on the line of kp TA kb T-A∼8.6×10−5 sec−2. and the period length tends to be extended when the parameters are close to the boundary of the stable oscillation.
Figure 4. The parameter dependence of the period length of oscillation in the phosphorylation level p(t).

The period length is shown in gray scale against (a) logkp TA and logkb T-A or (b) logkdp RB and logkb R-B. The oscillation dies out in the black area.
The oscillation can be further characterized by examining the association dynamics among Kai proteins. In the following, the simulated association dynamics are compared with the experimental results of Ref.14 from several aspects.
Dependence on concentrations of Kai proteins
It was experimentally observed that the oscillation is persistent when concentrations of Kai proteins are increased from their standard values, but the oscillation dies out when the concentrations are decreased to 1/10 or less [14]. When the concentration of KaiC is fixed, the oscillation is stable against a small change in concentration of KaiA or KaiB, but the oscillation dies out when the KaiA concentration is decreased to 1/4 or the KaiB concentration is decreased to 1/3 of their standard values [14]. These features are well reproduced in the simulation as shown in Fig. 5. In the model, these features are largely explained by the Michaelis-Menten kinetics of binding among Kai proteins. With the standard values of concentrations of Kai proteins, the binding reaction rates are saturated and insensitive to the small changes in concentrations of Kai proteins. With the decreased concentration, however, the binding rates leave this saturated regime and begin to sensitively depend on the concentrations, which disturbs the balance among the reaction rates required for the oscillation.
Figure 5. Concentration dependence of the simulated time course of KaiC phosphorylation level, p(t).

(a) p(t) for the standard mixture (red) and p(t) calculated with the concentrations of KaiA, KaiB, and KaiC increased 2.5-fold (blue), 5-fold (purple) or decreased to 1/10 (orange), 1/20 (yellow), and 1/40 (green) of their standard values. (b) p(t) calculated with the KaiA concentration decreased to 2/3 (blue) or 1/4 (green) of its standard value (red). (c) p(t) calculated with the KaiB concentration decreased to 2/3 (blue) or 1/3 (green) of its standard value (red).
Kai protein complex formation
We refer to the probability for a KaiC hexamer to be in complex with KaiA or with KaiB as P AC and P BC, respectively, and the probability for a KaiC to be in complex with KaiA and KaiB as P ABC. Then, the ratio of the free KaiC hexamer is expressed as P free = 1−(P AC+P BC+P ABC). In Ref.14, it was experimentally observed that P free oscillates between 55–80% and P BC oscillates between 7–18%, which are 180° out of phase with each other. P AC oscillates between 8–12% in phase with that of P free, and P ABC oscillates between 10–14% in phase close to that of P BC. Temporal changes of corresponding values in our simulation are plotted in Fig. 6, which are obtained by calculating P AC = Σi([TiA]+[RiA])/X, P BC = Σi[RiB]/X, and P ABC = Σi([TiAB]+[RiAB]+[RiBA])/X. The overall features of the experiment are mostly reproduced in Fig. 6: The simulated oscillations of P free and P BC resemble the experimental data, and P AC oscillates with the same oscillation phase as was observed in the experiment. The range of oscillation of P AC, however, is 18–30% in the model, which is consistently higher than the experimentally observed range of 8–12%, and P ABC oscillates only weakly in Fig. 6. Such disagreements may be due to the inappropriate description of binding of B to AC in the model. Further experimental information on the binding mode of KaiB to KaiC-KaiA complex is desired.
Figure 6. Dynamics of protein association as shown by the amount of the KaiC-KaiA complex (P AC, green), the KaiC-KaiB complex (P BC, blue), the KaiC-KaiA-KaiB and KaiC-KaiB-KaiA complexes (P ABC, red), and the free KaiC (P free, black).

The amounts are expressed as percentage of the total number of KaiC hexamers.
Kinetics of KaiC-KaiA and KaiC-KaiB association
In the experiment, when KaiC with the phosphorylation level ∼95% was incubated in the presence of KaiA, the phosphorylation level was kept constant at ∼95% and the KaiC-KaiA complex was readily formed to give P AC ∼10% [14]. On the other hand, when KaiC with the phosphorylation level ∼5% was incubated in the presence of KaiA, it took ∼6 hours for the phosphorylation level to increase to ∼95%, while the KaiC-KaiA complex was readily formed to make P AC ∼10% from the beginning [14]. These observations indicate that the binding affinity of KaiA to KaiC does not depend on the phosphorylation level and the rate of KaiC-KaiA binding is much faster than that of phosphorylation. As shown in Figs. 7a and 7b, these features are qualitatively reproduced by our model: Starting from the solution of KaiA and KaiC with p(0) ∼95%, p(t) approaches ∼75% in the model and P AC stays at ∼30%. Starting from the solution of KaiA and KaiC with p(0) ∼5%, it took 12 hours for p(t) to increase to ∼75%, while P AC stays at ∼30% from the beginning.
Figure 7. Interplay of the KaiC-KaiA and KaiC-KaiB association and the phosphorylation level of KaiC.

The amount of associated KaiC is shown in solid lines as percentage of the total number of KaiC hexamers, whereas the phosphorylation level, p(t), is shown in dashed lines. (a) KaiC with p(0) ∼95% is incubated with KaiA, (b) KaiC with p(0) ∼5% is incubated with KaiA, (c) KaiC with p(0) ∼95% is incubated with KaiB, and (d) KaiC with p(0) ∼5% is incubated with KaiB.
Kinetics of KaiC-KaiB association was also measured experimentally [14]. When KaiC with the phosphorylation level ∼95% was incubated with KaiB, it took 8 hours for the phosphorylation level to decrease to 5% and the phosphorylation level further decreased to less than 5%. P BC increased to 20% within the first 8 hours and then gradually decreased to 10%. When KaiC with the phosphorylation level ∼5% was incubated with KaiB, on the other hand, the phosphorylation level decreased to ∼0%, while P BC increased to 10% [14]. Thus, KaiB binds strongly to highly phosphorylated KaiC but only weakly to KaiC with the phosphorylation level<5%. Most of these features are qualitatively reproduced by the model as shown in Figs. 7c and 7d: As the phosphorylation level decreases, KaiC accumulates in the T0 state, which in turn decreases the binding affinity of KaiB to KaiC.
Shuffling of KaiC monomers
It has been shown experimentally that two KaiC hexamers can interact with each other and exchange monomers between them [14]. After the KaiC hexamers composed of tagged monomers and those without a tagged monomer were mixed at a ratio of 1:1, the kinetics of shuffling was followed by the increase of tagged KaiC hexamers. In the absence of KaiA or KaiB or in the presence of KaiB, the number of tagged KaiC hexamers became twice of the initial amount in about 4 hours, showing that all the KaiC hexamers interact and exchange monomers within this time interval. In the presence of KaiA, however, the amount of tagged KaiC monomers did not increase as much as that in the above cases. In the presence of KaiA and KaiB, the shuffling activity was low at the beginning but rose rapidly after the phosphorylation level had reached a maximum [14]. These features are also qualitatively reproduced in the model as shown in Fig. 8.
Figure 8. Monomer shuffling between KaiC hexamers.

(a) KaiC hexamers composed of tagged monomers and those composed of untagged monomers were mixed in the ratio of 1:1 in the absence of KaiA or KaiB (real line), in the presence of KaiB (dashed line), or in the absence of KaiA (dotted line). The amounts of tagged KaiC hexamers normalized by its initial amount are shown. (b) The phosphorylation level p(t) (dashed line) and the amount of tagged KaiC hexamers normalized by its initial amount (solid line) in the presence of KaiA and KaiB.
Temperature compensation
It was observed that, with the change of temperature from 25°C to 35°C, the oscillation period length was shortened only by 5% (from 22 h to 21 h) [7]. The model provides a possible explanation of this weak temperature dependence. As shown in Fig. 1, the model has two opposite pathways of phosphorylation and dephosphorylation within the both T and R states. In T state, for example, when the dephosphorylation process has higher activation energy than the phosphorylation process, the dephosphorylation rate approaches the phosphorylation rate at an elevated temperature. This effect should counteract the acceleration of reactions due to the temperature increase in both T and R states, yielding the weaker temperature dependence of the oscillation period. The simulation results of Fig. 9 show that the period is indeed temperature-compensated through this mechanism, so that the period is shortened only by 10% in the temperature increase from 300K to 309K, even though time constant of each reaction decreases by 26–32%. See Model and Methods for the temperature dependence of individual reactions defined in the model.
Figure 9. Temperature compensation of the oscillation period length of the phosphorylation level p(t).

The period length is normalized by its value at 300K.
Discussion
In this paper, we constructed a model of oscillation of KaiC phosphorylation and showed that the model can qualitatively reproduce the hitherto published experimental data. However, this model has some quantitative disagreements with the experimental data, such as the oscillation range of P AC in Fig. 6 or the phosphorylation level in Fig. 7. Although some of these points may be remedied by introducing further reaction channels in the model, such revision of the model would not strengthen the plausibility of the model at this point with the lack of the corresponding experimental information. Instead, we here demonstrated that a simple model is enough to capture the essential features of the oscillation observed in Ref. 7 and those of the association dynamics among Kai proteins observed in Ref. 14.
The model, however, still has a room for improvement. Possible extensions include the introduction of the process of ATP hydrolysis and phosphorylation at two phosphorylation sites, Ser431 and Thr432. Since the ATP binding site and two phosphorylation sites are close in space in the crystal structure of KaiC hexamer [25], the timing of phosphorylation or dephosphorylation at two sites should be correlated with each other, as well as with the timing of hydrolysis, capture, or release of the nucleotide. This correlation may well have significant effect on the stability and robustness of the the autocatalytic increase of KaiC population in R states discussed in this paper.
The present model is based on the two main assumptions, the shuffling of KaiC monomers, and the allosteric transition between R and T states of KaiC hexamer. It should be noted that both of these assumptions are natural consequences from the experimental observations. Overall qualitative agreement between the present model and the experimental data suggests that the monomer shuffling and the allosteric transition constitute a sufficient condition for the circadian oscillation of KaiC phosphorylation. Furthermore, the analysis on the population dynamics provides an explanation on why KaiC hexamers can synchronize in their phosphorylation level: the autocatalytic increase of KaiC population in R states can adjust the otherwise incoherent levels of phosphorylation. Comparison of the further improved model and the detailed experiments will provide a deeper understanding of a core mechanism of circadian rhythmicity.
Methods
Model and Methods
In this section, the kinetic scheme of each reaction in the model is explained.
Autokinase/phosphatase activity
In the absence of KaiA or KaiB, the phosphorylation level is low at equilibrium at room temperature, showing that the weak autophosphatase activity of KaiC is dominant [14], [16], [27] over the even weaker autokinase activity [28]. Thus, we assume that KaiC is dephosphorylated with the rate kdp T[Ti +1] or kdp R[Ri +1] for i = 0 to 5. When complexed with KaiA, KaiC is phosphorylated [14] with the rate kp TA[TiA] for i = 0 to 5 or kp RA[RiA] for i = 1 to 5. When complexed with KaiB, KaiC in the R state is dephosphorylated with the rate kdp RB[Ri +1B] for i = 0 to 5. We require that the catalyzed dephosphorylation should be faster than the autodephosphorylation; kdp RB>kdp T≈kdp R. Reactions in the KaiABC complex have not yet been specified experimentally, but we assume that KaiC in TiAB or RiAB is phosphorylated as in TiA or RiA with the rate kp TAB[TiAB] for i = 0 to 5 or kp RAB[RiAB] for i = 1 to 5, and that KaiC in RiBA is dephosphorylated as in RiB with the rate kdp RBA[Ri +1BA] for i = 0 to 5. If we assume equal phosphorylation rates for all unphosphorylated monomers, the phosphorylation rate for hexamer decreases as phosphorylation proceeds. Likewise, if we assume equal dephosphorylation rates for all phosphorylated monomers, the dephosphorylation rate for hexamer decreases as dephosphorylation proceeds. This assumption, however, centralize KaiC hexamer population around the states with i = 3 to 4, which suppresses the oscillation. Instead, we here assume that the reaction takes place at a specific monomer in each KaiC hexameric state, so that kdp T, kdp R, kp TA, kp RA, kdp RB, kp TAB, kp RAB, and kdp RBA are assumed to be independent of i.
Association/dissociation of Kai proteins
Homooligomers of KaiC (Ti or Ri), and KaiA in solution are assumed to be a hexamer [14], [22], [23] and a dimer [14], [27], [29]–[31], respectively. KaiA is supposed to be dimeric when bound to KaiC hexamer [14], [32]. We assume that KaiA binds to KaiC with the rate kb T-A[A][Ti]/(KM T-A+[A]) or kb R-A[A][Ri]/(KM R-A+[A]), where [A] is the concentration of KaiA dimer. This rather ad hoc assumption of Michaelis-Menten kinetics is required for explaining the invariability of the period length over the broad range of concentration of Kai proteins observed experimentally (Fig. 5). This suggests the possibility that the formation of KaiC-KaiA complex is preceded by the weak encounter complex which is in equilibrium with the uncomplexed KaiA and KaiC. KaiA dissociates from the KaiC-KaiA complex with the rate kd TA[TiA] or kd RA[RiA]. We assume that R0A is rapidly brought into T0A as will be explained later and we omit the KaiA dissociation from R0A. As the binding kinetics between KaiA and KaiC does not depend on the phosphorylation level of KaiC [14], we assume that the constants kb T-A, kb R-A, KM T-A, KM R-A, kd TA, and kd RA are independent of i.
KaiB is assumed to be a dimer in solution [14], [17], [30], and is supposed to be tetrameric when bound to KaiC, since the tetrameric form of KaiB was observed to be functionally important [33], [34]. Concentration of KaiB tetramer is proportional to [B]2, where [B] is the concentration of KaiB dimer, so that KaiB binds to KaiC with the rate kb
R-B[B]2[Ri]/(KM
R-B+[B]2) or  . The binding rate of KaiB is large when KaiC is phosphorylated [14], so that we simply put
. The binding rate of KaiB is large when KaiC is phosphorylated [14], so that we simply put  for i = 0 to 5 and
for i = 0 to 5 and  to be nonzero. KaiB dissociates from RiB with the rate kd
RB[RiB]. We assume that T6B is rapidly brought into R6B as will be explained later and we omit the KaiB dissociation from T6B. For simplicity we assume that kb
R-B, KM
R-B and kd
RBare independent of i.
to be nonzero. KaiB dissociates from RiB with the rate kd
RB[RiB]. We assume that T6B is rapidly brought into R6B as will be explained later and we omit the KaiB dissociation from T6B. For simplicity we assume that kb
R-B, KM
R-B and kd
RBare independent of i.
Likewise, the KaiC-KaiA-KaiB complex is formed by binding of KaiB to KaiC-KaiA and the KaiC-KaiB-KaiA complex is formed by binding of KaiA to KaiC-KaiB. Even though these two complexes are experimentally indistinguishable at present, we take them into account separately because it is proposed that KaiA and KaiB compete with each other for a common binding site on KaiC hexamer [28]. The binding reactions have the rate kb TA-B[B]2[TiA]/(KM TA-B+[B]2), kb RA-B[B]2[RiA]/(KM RA-B+[B]2) or kb RB-A[A][RiB]/(KM RB-A+[A]). KaiA dissociates from the KaiC-KaiB-KaiA complex with the rate kd RBA[RiBA] and two KaiB dimers dissociate from the KaiC-KaiA-KaiB complex with the rate kd TAB[TiAB] or kd RAB[RiAB]. We assume that the constants, kb TA-B, kb RA-B, kb RB-A, K M TA-B, K M RA-B, K M RB-A, kd RBA, kd TAB, and kd RAB are independent of i.
Allosteric transitions and shuffling
Conformational transition between the T state and the R state of KaiC hexamer is limited to the KaiC with i = 0 or 6, and other possible transitions are neglected. Allosteric transition between T0 state and R0 state occurs in three reactions: the dominant and catalyzed reaction R0+A→T0A with the rate  , the uncatalyzed reaction R0→T0 with the rate
, the uncatalyzed reaction R0→T0 with the rate  , and the uncatalyzed reverse reaction T0→R0 with the rate
, and the uncatalyzed reverse reaction T0→R0 with the rate  . Here, the rates of the uncatalyzed reactions are assumed to be similar and lower than that of the catalyzed reactions:
. Here, the rates of the uncatalyzed reactions are assumed to be similar and lower than that of the catalyzed reactions:  . Allosteric transition between T6 state and R6 state occurs in only one reaction T6+B→R6B with the rate
. Allosteric transition between T6 state and R6 state occurs in only one reaction T6+B→R6B with the rate  . As discussed above,
. As discussed above,  has to be low enough for the the autocatalytic increase of KaiC population in R states to occur so that
has to be low enough for the the autocatalytic increase of KaiC population in R states to occur so that 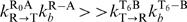 . We assume that R0A and T6B are the special states primed for conformational transition so that phosphorylation or dephosphorylation does not occur from R0A and T6B.
. We assume that R0A and T6B are the special states primed for conformational transition so that phosphorylation or dephosphorylation does not occur from R0A and T6B.
We assume that monomer shuffling occurs between two KaiC hexamers when both of them are in the R state as Ri+Rj→Rk+Rl with the rate ks RR[Ri][Rj]. Since shuffling is most frequent for several hours at the beginning of the dephosphorylation phase [14] we introduce the additional shuffling reactions involving the R6 state, which is populated at this interval, and the T state as R6+Rj→Rk+Rl with the rate ks RT[R6][Tj]. Note that the resulting two monomers are again assumed to be both in the R state. This amounts to an assumption that the R state is entropically stabilized and is more easily formed than the T state which takes more time to be realized after shuffling. For both types of shuffling reactions, we assume that monomers are randomly exchanged between two hexamers, which results in the binomial distribution of k and l with i+j = k+l. For simplicity, we assume ks RR and ks RTdo not depend on i or j.
Temperature dependence
We assume that the rate constants defined above depend on temperature as k∝exp(−ΔE/k B T), where k B is the Boltzmann constant. Though there is no available experimental data to estimate ΔE for each reaction, it would be reasonable to assume that ΔE should be within the range of typical biochemical processes of 10-20k B T. For most of the rate constants the activation energy was set to be ΔE = 10k B T 0 with T 0 = 300K, but 12k B T 0 for kd TA, kd TAB, kb R-A, kd RB, kd RAB, kb RA-B and 13k B T 0 for kdp T, kp RA, and kp RAB.
Parameters and initial conditions
Values of parameters are summarized in Supplementary Table S1. The oscillatory behavior in the model does not sensitively depend on the parameter values. As there is no direct experimental observation to determine these values, they were manually tuned to fit the experimental data semi-quantitatively in Figs. 4– 8. When the reaction mixture was prepared by mixing recombinant Kai proteins, KaiC seemed to be phosphorylated to some extent [14]. We therefore start our simulations from the initial condition that KaiC is populated at the T state with the distribution of phosphorylation level i to be pi = 6!/(i!(6−i)!)0.5i0.56−i except for the simulations in Fig. 7. In Figs. 7a and c the initial distribution is pi = 6!/(i!(6−i)!)0.95i0.056−i and in Figs. 7b and d pi = 6!/(i!(6−i)!)0.05i0.0956−i.
Supporting Information
(0.31 MB DOC)
Acknowledgments
We are grateful to Prof. Takao Kondo and members of his research group for fruitful discussions.
Footnotes
Competing Interests: The authors have declared that no competing interests exist.
Funding: This work was supported by grants from the Ministry of Education, Culture, Sports, Science, and Technology, Japan, and from Japan Society for Promotion of Science and by grants for the 21st century COE program for Frontiers of Computational Science.
References
- 1.Bell-Pedersen D, Cassone VM, Earnest DJ, Golden SS, Hardin PE, et al. Circadian rhythms from multiple oscillators: lessons from diverse organisms. Nat Rev Genet. 2005;6:544–556. doi: 10.1038/nrg1633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kondo T, Mori T, Lebedeva NV, Aoki S, Ishiura M, et al. Improving the palatability of oral rehydration solutions has implications for salt and water transport: a study in animal models. Science. 1997;275:224–227. doi: 10.1097/00005176-199610000-00012. [DOI] [PubMed] [Google Scholar]
- 3.Ishiura M, Kutsuna S, Aoki S, Iwasaki H, Andersson CR, et al. Expression of a gene cluster kaiABC as a circadian feedback process in cyanobacteria. Science. 1998;281:1519–1523. doi: 10.1126/science.281.5382.1519. [DOI] [PubMed] [Google Scholar]
- 4.Nakahira Y, Katayama M, Miyashita H, Kutsuna S, Iwasaki H, et al. Global gene repression by KaiC as a master process of prokaryotic circadian system. Proc Natl Acad Sci U S A. 2004;101:881–885. doi: 10.1073/pnas.0307411100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Young MW, Kay SA. Time zones: a comparative genetics of circadian clocks. Nat Rev Genet. 2001;2:702–715. doi: 10.1038/35088576. [DOI] [PubMed] [Google Scholar]
- 6.Naef F. Circadian clocks go in vitro: purely post-translational oscillators in cyanobacteria. Mol Syst Biol. 2005;1:2005.0019. doi: 10.1038/msb4100027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Nakajima M, Imai K, Ito H, Nishiwaki T, Murayama Y, et al. Reconstitution of Circadian Oscillation of Cyanobacterial KaiC Phosphorylation in Vitro. Science. 2005;308:414–415. doi: 10.1126/science.1108451. [DOI] [PubMed] [Google Scholar]
- 8.Takai N, Nakajima M, Oyama T, Kito R, Sugita C, et al. A KaiC-associating SasA-RpaA two-component regulatory system as a major circadian timing mediator in cyanobacteria. Proc Natl Acad Sci U S A. 2006;103:12109–12114. doi: 10.1073/pnas.0602955103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Emberly E, Wingreen NS. Hourglass model for a protein-based circadian oscillator. Phys Rev Lett. 2006;96:038303. doi: 10.1103/PhysRevLett.96.038303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kurosawa G, Aihara K, Iwasa Y. A model for the circadian rhythm of cyanobacteria that maintains oscillation without gene expression. Biophys J. 2006;91:2015–2023. doi: 10.1529/biophysj.105.076554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Mehra A, Hong CI, Shi M, Loros JJ, Dunlap JC, et al. Circadian rhythmicity by autocatalysis. PLoS Comput Biol. 2006;2:e96. doi: 10.1371/journal.pcbi.0020096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Takigawa-Imamura H, Mochizuki A. Predicting regulation of the phosphorylation cycle of KaiC clock protein using mathematical analysis. J Biol Rhythms. 2006;21:405–416. doi: 10.1177/0748730406291329. [DOI] [PubMed] [Google Scholar]
- 13.Miyoshi F, Nakayama Y, Kaizu K, Iwasaki H, Tomita M. A mathematical model for the Kai-protein-based chemical oscillator and clock gene expression rhythms in cyanobacteria. J Biol Rhythms. 2007;22:69–80. doi: 10.1177/0748730406295749. [DOI] [PubMed] [Google Scholar]
- 14.Kageyama H, Nishiwaki T, Nakajima M, Iwasaki H, Oyama T, et al. Cyanobacterial circadian pacemaker: Kai protein complex dynamics in the KaiC phosphorylation cycle in vitro. Mol Cell. 2006;23:161–171. doi: 10.1016/j.molcel.2006.05.039. [DOI] [PubMed] [Google Scholar]
- 15.Iwasaki H, Nishiwaki T, Kitayama Y, Nakajima M, Kondo T. KaiA-stimulated KaiC phosphorylation in circadian timing loops in cyanobacteria. Proc Natl Acad Sci U S A. 2002;99:15788–15793. doi: 10.1073/pnas.222467299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kitayama Y, Iwasaki H, Nishiwaki T, Kondo T. KaiB functions as an attenuator of KaiC phosphorylation in the cyanobacterial circadian clock system. EMBO J. 2003;22:2127–2134. doi: 10.1093/emboj/cdg212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kageyama H, Kondo T, Iwasaki H. Circadian formation of clock protein complexes by KaiA, KaiB, KaiC, and SasA in cyanobacteria. J Biol Chem. 2003;278:2388–2395. doi: 10.1074/jbc.M208899200. [DOI] [PubMed] [Google Scholar]
- 18.Monod J, Wyman J, Changeux JP. On the nature of allosteric transitions: a plausible model. J Mol Biol. 1965;12:88–118. doi: 10.1016/s0022-2836(65)80285-6. [DOI] [PubMed] [Google Scholar]
- 19.Yifrach O, Horovitz A. Nested cooperativity in the ATPase activity of the oligomeric chaperonin GroEL. Biochemistry. 1995;34:5303–5308. doi: 10.1021/bi00016a001. [DOI] [PubMed] [Google Scholar]
- 20.Perutz MF, Wilkinson AJ, Paoli M, Dodson GG. The stereochemical mechanism of the cooperative effects in hemoglobin revisited. Annu Rev Biophys Biomol Struct. 1998;27:1–34. doi: 10.1146/annurev.biophys.27.1.1. [DOI] [PubMed] [Google Scholar]
- 21.Horovitz A. Structural aspects of GroEL function. Curr Opin Struct Biol. 1998;8:93–100. doi: 10.1016/s0959-440x(98)80015-8. [DOI] [PubMed] [Google Scholar]
- 22.Mori T, Saveliev SV, Xu Y, Stafford WF, Cox MM, et al. Circadian clock protein KaiC forms ATP-dependent hexameric rings and binds DNA. Proc Natl Acad Sci U S A. 2002;99:17203–17208. doi: 10.1073/pnas.262578499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hayashi F, Suzuki H, Iwase R, Uzumaki T, Miyake A, et al. ATP-induced hexameric ring structure of the cyanobacterial circadian clock protein KaiC. Genes to Cells. 2003;8:287–296. doi: 10.1046/j.1365-2443.2003.00633.x. [DOI] [PubMed] [Google Scholar]
- 24.Nishiwaki T, Satomi Y, Nakajima M, Lee C, Kiyohara R, et al. Role of KaiC phosphorylation in the circadian clock system of Synechococcus elongatus PCC 7942. Proc Natl Acad Sci USA. 2004;101:13927–13932. doi: 10.1073/pnas.0403906101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Xu Y, Mori T, Pattanayek R, Pattanayek S, Egli M, et al. Identification of key phosphorylation sites in the circadian clock protein KaiC by crystallographic and mutagenetic analyses. Proc Natl Acad Sci U S A. 2004;101:13933–13938. doi: 10.1073/pnas.0404768101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Xu Y, Mori T, Johnson CH. Cyanobacterial circadian clockwork: roles of KaiA, KaiB and the kaiBC promoter in regulating KaiC. EMBO J. 2003;22:2117–2126. doi: 10.1093/emboj/cdg168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ye S, Vakonakis I, Ioerger TR, LiWang AC, Sacchettini JC. Crystal structure of circadian clock protein KaiA from Synechococcus elongatus. J Biol Chem. 2004;279:20511–20518. doi: 10.1074/jbc.M400077200. [DOI] [PubMed] [Google Scholar]
- 28.Garces RG, Wu N, Gillon W, Pai EF. Anabaena circadian clock proteins KaiA and KaiB reveal a potential common binding site to their partner KaiC. EMBO J. 2004;23:1688–1698. doi: 10.1038/sj.emboj.7600190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Uzumaki T, Fujita M, Nakatsu T, Hayashi F, Shibata H, et al. Crystal structure of the C-terminal clock-oscillator domain of the cyanobacterial KaiA protein. Nat Struct Mol Biol. 2004;11:623–631. doi: 10.1038/nsmb781. [DOI] [PubMed] [Google Scholar]
- 30.Pattanayek R, Williams DR, Pattanayek S, Xu Y, Mori T, et al. Analysis of KaiA-KaiC protein interactions in the cyano-bacterial circadian clock using hybrid structural methods. EMBO J. 2006;25:2017–2028. doi: 10.1038/sj.emboj.7601086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Iwase R, Imada K, Hayashi F, Uzumaki T, Morishita M, et al. Functionally important substructures of circadian clock protein KaiB in a unique tetramer complex. J Biol Chem. 2005;280:43141–43149. doi: 10.1074/jbc.M503360200. [DOI] [PubMed] [Google Scholar]
- 32.Hitomi K, Oyama T, Han S, Arvai AS, Getzoff ED. Tetrameric architecture of the circadian clock protein KaiB. A novel interface for intermolecular interactions and its impact on the circadian rhythm. J Biol Chem. 2005;280:19127–19135. doi: 10.1074/jbc.M411284200. [DOI] [PubMed] [Google Scholar]
- 33.Nishiwaki T, Iwasaki H, Ishiura M, Kondo T. Nucleotide binding and autophosphorylation of the clock protein KaiC as a circadian timing process of cyanobacteria. Proc Natl Acad Sci U S A. 2000;97:495–499. doi: 10.1073/pnas.97.1.495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Gillespie DT. Exact stochastic simulation of coupled chemical reactions. J Phys Chem. 1977;81:2340–2361. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(0.31 MB DOC)