

Abstract
The abuse of anabolic androgenic steroids (AS) is a growing problem; however, the effects and mechanisms underlying their addictive effects are not well understood. Research findings regarding androgen abuse in people and hedonic effects of androgens in laboratory rats are reviewed. Androgens, like other steroids, can have traditional actions via cognate intracellular steroid receptors, as well as other substrates. Our recent results indicate that testosterone (T) metabolites may have actions in part via γ–aminobutyric acid (GABA)A/benzodiazepine receptor complexes (GBRs) and/or dopaminergic neurons in the nucleus accumbens, to mediate T’s positive hedonic states. This may provide the basis for positive reinforcing effects of androgen seeking and use behavior. Following a comprehensive review of the background literature, our findings are presented that have explored the extent to which metabolites of T mediate euphorogenic effects of androgens by acting in the nucleus accumbens. Then results regarding whether GBRs are necessary substrates for androgens’ positive hedonic effects are discussed. Lastly, research that addresses if dopaminergic neurons in the nucleus accumbens are necessary for these effects of androgens are discussed. This review provides a comprehensive examination of the hedonic properties and abuse/addiction potential of androgens and the putative mechanisms underlying these effects.
Keywords: testosterone, reward, reinforcement, hedonic, conditioning, GABAA receptors, dopamine receptors, anxiety, affect, learning
1. Introduction-Overview
Anabolic-androgenic steroids (AS) are the synthetic variants of the primary masculinizing androgen, testosterone (T). They are abused by growing numbers of individuals in this country ranging from adolescents, seeking to improve their appearance, to professional athletes attempting to elevate their performance. The costs associated with AS abuse are substantial. For the individual, AS abuse is associated with many adverse physical and behavioral consequences. For society at large, AS abuse has spawned a significant black market which has promoted criminal behavior and placed strain on law enforcement agencies. The aim of this review is to explore more fully the causes of androgen abuse in people by describing research from our laboratory and others investigating hedonic effects of androgens.
In contrast to our understanding of the classic drugs of abuse like cocaine, heroin and alcohol, relatively little is known about the causes of AS abuse. Indeed, the principal question surrounding most drugs of abuse is to what extent they produce euphorogenic (positive hedonic and rewarding) effects, which can maintain and/or exacerbate future drug seeking behavior. Our earlier findings, and those from other laboratories, are mixed. While positive hedonic effects of T have been reported by some, the results have been compromised by serious procedural problems and significant variability. Our recent research suggests that a portion of this variability may be due to differences in the metabolism of T and the resulting availability of androgen metabolites at receptor sites in the brain. Our results suggest that the actions of T metabolites, perhaps in part through γ-aminobutyric acid (GABA)A/benzodiazepine receptor complexes (GBRs) and/or dopaminergic neurons in the nucleus accumbens (NA), may mediate T’s production of positive hedonic states and consequently provide the basis for positively reinforcing effects on drug seeking and use behaviors.
This review paper initially summarizes background research that has prompted our investigations aimed at answering four key questions geared towards elucidating the mechanism of androgens’ actions in the NA to produce hedonic effects. First, we have investigated whether the ability of T to produce positive hedonic effects involve its metabolites. Comparisons between T and its metabolites and effects of pharmacological blockade of T metabolism have been used to ascertain whether such manipulations inhibit the positive hedonic effects produced by T. Second, because some of T’s metabolites do not bind well to intracellular androgen receptors (ARs), we have examined effects of blocking ARs, on hedonic effects of androgens. Third, the importance of GBRs in the NA as substrates for androgens’ hedonic effects have been addressed using intra-accumbens infusions of antagonists and agonists to investigate the importance of GBRs for androgens’ production of positive hedonic state. Fourth, whether dopaminergic neurons in the NA are required for androgens’ hedonic effects has been investigated using intra-accumbens 6-OHDA infusions. These questions have been examined in rats by using classic methods of behavioral pharmacology and neuroendocrinology to manipulate androgens’ actions and examining hedonic effects using a traditional Pavlovian conditioning technique, conditioned place preference (CPP), to assess the ability of androgens to produce positive hedonic states. The results of this research from our laboratory comprise the second half of this review paper. Taken together, the information presented in this review expands our understanding of the biochemical pathways through which androgens work to produce euphorogenic effects which may lead to their abuse.
2. Background & Significance Definition of AS
Anabolic steroids are androgenic and/or synthetic variants of T, which is the primary masculinizing hormone secreted by the mammalian testes, and is also secreted by ovaries and adrenals, albeit in lower concentrations. The original goal in producing these drugs was to promote the well-known anabolic (tissue building) effects of T without its androgenic (masculinizing) properties. Importantly, although the androgenic effects of some of the presently marketed anabolic steroids have been reduced, they have not been eliminated. Therefore, to date, no exclusively anabolic substance has yet been found (Bahrke, Yesalis, and Wright, 1996).
2.1. Medical Use of AS
It has been asserted that the German government under Hitler developed and were the first to use AS, allegedly in an attempt to create an army of supermen (e.g., Marshall, 1988). After the war, AS were used in legitimate medicine to treat female breast cancer (by reducing estrogen), to combat two different forms of anemia, and to reduce the effects of hereditary angioedema (Wright, 1980; Bahrke, Yesalis, and Brower, 1998). While other drugs currently are more effective in the treatment of the above ailments, AS presently are used in initiating delayed puberty in preadolescents, growth promotion, treatment of micropenis, and treatment of hypogonadism (Moore, 1988; Wilson and Griffin, 1980; Bahrke, Yesalis, and Wright, 1996).
2.2. Elicit Use of AS
As the medical need for AS declined, their popularity among athletes soared dramatically. Soviet weight lifters began using them as power boosters in Olympic competitions in the 1950s, followed by the Americans in the 1960s (Wade, 1972; Yesalis and Cowart, 1998). Recent confirmations indicate the East German government sanctioned a massive experimental program in which their top athletes were administered AS for years in order to enhance athletic performance and medal counts in the Olympic games (Franke and Berendonk, 1997).
2.3. Incidence of AS use among athletes
While AS initially were used by super-athletes as a means for adding an edge to a performance already close to perfection, the word gradually spread that they could be effective for any sport that required strength. Although actual figures on the incidence and prevalence of AS use among elite, amateur, and recreational athletes is just beginning to emerge, it is generally agreed that AS currently are used widely by professional, college, and high school football and baseball players, shot-putters, discus throwers, swimmers, sprinters, tennis players, and bicyclists (Yesalis and Cowart, 1998; Bahrke, Yesalis, and Wright, 1996). For example, studies commissioned by the National Collegiate Athletic Association (NCAA) (Anderson, Albrecht, McKeag, Hough, and McGrew 1991; Albrecht, Anderson, and McKeag, 1992), indicate that roughly 5 % of male and female athletes reported using AS, with the highest user rates (10%) seen in collegiate football players. In a survey of track and field athletes in the 1972 Olympics, 68% reported prior steroid use (Silvester, 1973). Among elite power lifters, 55% of those interviewed conceded prior AS use (Yesalis, Herrick and Buckley, 1988). In a study of amateur competitive body builders, over half of the men and 10% of the woman reported that they had used AS at some point in their life (Tricker, O’Neil, and Cook, 1989).
2.4. Effects of AS use Among Athletes
Although AS are widely used today to enhance strength and athletic performance, there is little scientific documentation of their reputed benefits. An older review concluded that AS were beneficial only to strength and athletic performance if the individuals were in a continuing program of intensive exercise coupled with a high protein diet (Haupt and Rovere, 1984). However, a more recent study (Bhasin, Storer, Berman, Callegari, Clevenger, Phillips, Bunnell, Tricker, Shirazi, and Casaburi, 1996) controlled dietary factors, type of exercise, weight lifting experience, and duration and dose of AS exposure and was the first to demonstrate that supraphysiological dosages of T, with or without strength training, increase fat-free mass, muscle size, and strength in normal men. The results further showed that men who received 10 weeks of strength training or 10 weeks of T experienced significant increases in muscle size and strength; however, those that received both T and strength training had increased strength, tricep and quadricep size and fat-free mass that was well beyond that of men who received placebos, just AS with no strength training, or just strength training with no AS. These data demonstrate that T and exercise produced additive increases in performance. Also, a review of confiscated documents following the collapse of the German Democratic Republic clearly shows that AS had a positive effect on athletic performance in adult women and children (Franke and Berendonk, 1997).
A credibility and information gap between athletes and the medical/scientific community has resulted in the banning of AS by a number of athletic organizations (cf., Haupt and Rovere, 1984; Yesalis and Cowart, 1998) so as to protect athletes from potentially harmful side effects (see below). While sophisticated detection procedures have been developed in order to enforce these bans, athletes continue to abuse AS, often times by utilizing masking agents, or new AS which as yet can not be detected.
2.5. Incidence of AS use among Adolescents
Anabolic-androgenic steroids abuse is no longer confined to elite Olympic, professional, college or high school athletes. Lifetime use incidence is 5% for males and 3% for females, which indicates that more than 1 million Americans have taken AS illicitly. Although it may not be surprising that 55% of 27-year-old male and 10% of 24-year-old female body builders admit to using AS, and that the incidence in college athletes is estimated at 20%, AS abuse is now a problem that influences a much broader population including adolescents and young adults. The first nationwide study of AS abuse in 1987 particularly is revealing (Buckley, Yesalis, Friedl, Anderson, Streit, and Wright, 1988) since other more recent studies generally confirm these findings. The study showed that 7% of 12th grade male and 3% of female students were using or had used AS and that two thirds of the user group initiated use when they were 16 years of age or younger. Importantly, the survey also showed a number of intriguing trends in AS abuse. 47% of users reported that the main reason for using the drug was to improve athletic performance but an alarming 27% of the user group listed appearance as the main reason. Also, many users in the study could be described as habitual because (1) 40% of the self-identified users reported using AS for five or more cycles with each cycle usually lasting six to 12 weeks, and (2) 44% of users responded that they used more than one AS drug at the same time (“stacking”). More recent surveys of 9th through 12th grade public and private high school students confirm the earlier results. For example, the 1995 Youth Risk and Behavior Surveillance System data showed that 4.9% of boys and 2.4% of girls have used AS at least once in their lives (Kann, Warren, Harris, Collins, Williams, Ross, and Kolbe, 1996). More recently, it was reported that 4 % of Massachusetts junior high school students have used AS (Faigenbaum, Zaichkowsky, Gardner, and Micheli, 1998). These findings indicate that the epidemiology of AS abuse is following a classic pattern similar to that of other drugs of abuse. Namely, abuse by elite groups is soon followed by widespread use of the drug by many other segments of the general population. Extrapolating from all of these findings, it is estimated that approximately 375,000 adolescent boys and 175,000 adolescent girls are steroid users. This represents a significant population at risk as the potential for adverse effects (see below) may be greatest in this age group.
2.6. Traffic in AS
The alarming figures reported above are even more striking in view of legislation in 1990 (Anabolic Steroids Control Act) that attempted to control by reclassifying AS in the United States as Schedule III controlled substances. Distribution without a prescription is a felony punishable by up to 5 years in prison and a $250,000 fine. Possession of AS in the US is a misdemeanor punishable by up to l year imprisonment and a minimum of a $1000 fine. AS use is a pervasive international problem (other countries are reporting problems similar to those seen in the U.S. (Bahrke, 1995).
Despite government attempts to control AS, their demand has resulted in rapidly escalating black market, which was estimated to involve $500 million in sales in 1994 (Kouri, Pope and Katz, 1994). A growing concern is the nutritional supplement field in which products such as androstenedione (Andro) and dehydroepiandrosterone (DHEA) contain AS. Not covered by the Anabolic Steroids Control Act, over-the-counter and internet sales of Andro and DHEA represent a sizable and growing portion of the $15 billion nutritional supplement market (Yesalis and Cowart, 1998).
2.7. Health Risks Associated with AS
The health risks associated with AS abuse are considerable (e.g., Haupt and Rovere, 1984; Yesalis and Cowart, 1998; Bahrke, Yesalis, and Wright, 1996; Yesalis and Bahrke, 1995). The androgenic (masculinizing) risk is particularly acute for women, in whom it may be impossible to reverse masculine traits, such as facial hair, deepening of voice, and male physique, once they appear. In adult males, AS abuse can lead to kidney and liver damage, liver cancer, heart disease, and hypertension. It also can cause suppression of T production, enlargement of mammary tissue (gynecomastia), testicular atrophy, and decreased spermatogenesis. In adolescent males, AS abuse can hasten the onset of adulthood, promote early baldness, limit stature, and cause premature growth plate closing.
Many studies indicate that androgens modulate aggressive and copulatory behavior in rodents (Svare, Mann, Broida, Kinsley, Ghiraldi, Miele, and Konen, 1983; Kinsley and Svare, 1988; Frye, van Keuran, and Erskine, 1996; Frye, Duncan, Basham, and Erskine, 1996). There are far fewer studies documenting a positive relationship between endogenous androgens and aggressive and copulatory behavior in humans. However, in men, there is ample evidence for an association between androgens and these behaviors (Bahrke, Yesalis and Wright, 1990; 1996).
2.8. Hedonic Effects of Androgens Among People
There is now little doubt that AS can have significant effects upon mood and mental disorders (cf., Bahrke, Yesalis and Wright, 1990; 1996). Clinical reports showing increases in affective and psychotic syndromes, a number of which are very violent and suicidal in nature, are associated with AS use in individuals seeking to improve their performance or appearance (e.g., Pope and Katz, 1987 e.g., Pope and Katz, 1988; Pope, Katz, and Aizley, 1994). Indeed, there are now several legal cases in which defendants have claimed that AS’ effects upon their behavior promoted their criminal acts (e.g., Bahrke, Yesalis and Wright, 1990). Data from the National Household Survey on Drug Abuse have shown a strong association between AS use and self-acknowledged acts of violence against people and property crimes. Based upon this accumulating information, some researchers (Orchard and Best, 1994) have suggested that violent offenders should be tested routinely for AS so as to further document the relationship and develop methods to control steroid abuse.
Case studies and anecdotal reports show AS abuse has been associated with changes in depression, euphoria, hypomania, increased aggression, libido, alertness, irritability, anger, anxiety, energy, hostility, mood swings, psychotic episodes, and violent rages (cf., Bahrke, Yesalis, and Brower, 1998; Yesalis and Cowart, 1998). Self-reported changes in mood, behavior, and somatic perceptions also have been associated with AS abuse (Bahrke, Wright, Strauss, and Catlin, 1992; Wilson, 1988). Up to 43% of AS users report feeling “high” or feeling extreme pleasure from using AS over extended periods of time (Brower, Blow, Young, and Hill, 1991). Similarly, many other uncontrolled studies have reported euphoric effects among athletes who have taken AS (reviewed in Taylor, 1987). Interestingly, some studies show that AS elicit electroencephlographic changes similar to those seen with amphetamines and tricyclic antidepressants (Bahrke, Yesalis and Wright, 1990). Indeed, these findings are consistent with reports that T was used to treat depression in the 1930’s (Altschule and Tilletson, 1948).
2.9. Dependence
Some researchers have raised the possibility that dependence may result from prolonged AS abuse (Tennant, Black, and Vou, 1988; Wright, 1980; Kashkin and Kleber, 1989). They note that AS abusers often experience a stimulant-like withdrawal syndrome characterized by depressive symptoms. Moreover, a number of studies and case reports have documented behavior, perceptions, and attitudes in some AS abusers that are indicative of dependence (e.g., Brower, Eliopulos, Blow, Catlin, and Beresford, 1990; Brower, Blow, Young, and Hill, 1991; Brower, Blow, Eliopulos, and Beresford, 1989; Corcoran and Longo, 1992; Pope and Katz, 1994). Since 1988, there have been at least four case reports of AS dependence in the medical literature. There is the case of a 24-year-old noncompetitive weight lifter, who met 6 of the 9 DSM criteria for dependence and experienced suicidal depression upon cessation of AS use (Brower, Blow, Beresford, and Fuelling, 1989). Another 22-year-old noncompetitive weight lifter reported low self-esteem and AS cravings so severe after AS cessation (Hays, Littleton, and Stillner, 1990) that he was unable to discontinue their use. As well, 3 weightlifters report initiating AS use to enhance performance but maintaining use to prevent withdrawal (Tennant, Black, and Vou, 1988). A 30-year-old woman was also described who had been taking AS for four years; she met 5 of 7 of the DSM criteria for dependence (Copeland, Peters, and Dillon, 1998).
These findings suggest that the positive hedonic effects of AS are very powerful and may be a primary mitigating factor for their continued use. Findings from survey research are consistent with the notion of dependence as demonstrated by intention for continued AS use despite adverse consequences (a DSM criteria for dependence). One-fourth of high school seniors that admitted to using AS indicated that they would not stop using them even if the drugs led to permanent sterility, increase in the risk of cancer, or heart attacks (Yesalis, Vicary, Buckley, Streit, Katz, and Wright, 1990); the response rate for heavy users of AS was as high as 50%. The incidence of AS users that meet the DSM criteria for abuse or dependence are as high as 100% and 75%, respectively (Brower, Blow, Young, and Hill, 1991; Brower, Elioulos, Blow, Catlin, and Beresford, 1990). Other studies that have used the DSM criteria for substance dependence report rates of 15 to 69% (Malone, Dimeff, Lombardo, and Sample, 1995; Pope and Katz, 1994; Clancy and Yates, 1992).
2.10. Hedonic Effects of Androgens in Animal Models
While these findings are interesting, their unsystematic and anecdotal nature limits their significance in understanding the mechanisms underlying AS dependence. Evidence from animal studies suggests T can have positive hedonic effects. Testosterone (Olds, 1958; Caggiula, 1970; Campbell, 1970), like many drugs of abuse (Kornetsky, 1995), will increase rates of bar pressing for electrical brain stimulation, which is considered an indication of a drugs’ rewarding effects. AS administration will increase the rate of bar pressing to deliver electrical brain stimulation to the mesolimbic system (Kornestsky, 1995; Cagguila and Hoebel, 1966; Herberg, 1963; Clark, Lindenfield and Gibbons, 1996). As well, animals can be made to be physically dependent on AS (Bonson, Garrick, and Murphy, 1994). Male hamsters preferentially self-administer testosterone orally (Johnson & Wood, 2001; Wood, 2002). In many studies of CPP, which is used to examine hedonic effects of drugs (Scoles and Siegel, 1986), T conditions a place preference (Alexander, Packard, and Hines, 1994; Caldarone, Stock, Abrahamsen, Boechler, Svare, and Rosellini, 1996; De Beun, Jansen, Slangen, and Van de Poll, 1992; Kashkin and Kleber, 1989; Packard, Cornell, and Alexander, 1997; Packard, Schroeder, and Alexander, 1998; Schroeder and Packard, 2000), when administered systemically (Alexander et al., 1994) or when applied centrally to the NA (Packard et al., 1997) or to the medial preoptic area (King, Packard, and Alexander, 1999). However, there is considerable variability in this effect. In some studies, CPP with T was seen only with very high systemic dosages and not with lower dosages; in others, no effect was observed (Caldarone et al., 1996). The requirement for high dosages of T in order to produce effects on CPP is consistent with the notion that tolerance can be seen following repeated administration.
2.11. Tolerance and Withdrawal
There is evidence from findings with people and animals that tolerance to AS can develop, which also leads to escalating and continued use. As early as 1950, Kochakian reported that the anabolic effect of AS were attenuated in rats repeatedly administered AS unless the dosages were increased. Up to 18% of AS users report tolerance (Brower, Blow, Young, and Hill, 1991). Withdrawal symptoms are also reported in rats given daily injections of T for 10, but not 3, weeks. For two weeks after T cessation rats had tremors, ataxic effects, and ptosis (Foltin, 1992). Although the findings discussed above indicate that androgens can produce interoceptive effects, what the nature of these effects are and the neurobiological substrates that mediate their actions remain to be elucidated.
3. Physiology & Pharmacology of Androgens
Steroids are vital for cell life. Early in evolution, hormones served as primitive growth regulators, and diversified later to sex steroids, gluco- and mineralo-corticoids with remarkable preservation of structure-activity relationships (Rousseau and Baxter, 1979). In mammals, all steroid hormones derive from cholesterol and are synthesized in steroidogenic organs, such as adrenals (mineralo-and gluco-corticoids), gonads and placenta (sex hormones including androgens), before being secreted into circulation. High lipophilicity of steroids facilitates penetration of biological membranes, securing access to all cells and organs, including the Central Nervous System (CNS).
The traditional view of how steroids exert their effects is through actions at specific intracellular steroid receptors. Briefly, once a steroid binds to its specific (cognate) intracellular steroid receptor, structural changes occur in the receptor that facilitate its binding to complementary regions of DNA in the cell nucleus. The receptor binding activates transcription of the gene(s), producing messenger RNA transcripts that encode a wide array of enzymatic, structural and receptor proteins (Rogozkin, 1991).
3.1. Genomic Actions of Androgens
Some of the effects of T and other AS may be mediated through intracellular androgen receptors (Janne, Palvimo, Kallio and Mehto, 1993). Androgen receptors are widely but selectively distributed throughout the brain (Stumpf and Sar, 1976). In the rat, the brain regions containing the highest levels of androgen receptors are the lateral septum, some areas in the hippocampus, the bed nucleus of the stria terminalis, the medial preoptic nucleus, the ventromedial hypothalamus, and the medial amygdaloid nucleus (Kritzer, 1997; Lieberburg, MacLusky, and McEwen, 1977). High dosages of AS lead to upregulation of androgen receptors in these areas (e.g., the ventral tegmental area, the CA-I region of the hippocampus), as well as in several non-classical target sites such as the locus ceruleus and the periaqueductal grey (Telegdy, 1987).
3.2. Non-Genomic Actions of Androgens
Steroids may also influence cellular activity in a “non-genomic” fashion or through means other than traditional actions at intracellular steroid receptors (Brann, Hendry, and Mahesh, 1995). Indeed, T metabolites and many AS do not bind with a high affinity at cognate intracellular androgen receptors (Cunningham, Tindall, and Means, 1979; Verhoeven, Heyns, and DeMoor, 1975). Some possible mechanisms for these non-classical actions are: changes in membrane fluidity; actions on receptors on plasma membranes; regulation of GBRs on plasma membranes; and, activation of steroid receptors by factors such as EGF, IGF-I and dopamine. These diverse intracellular and non-genomic modes of action provide for integrated actions of hormones which may be rapid and of short duration, or prolonged and of long duration.
There are well-preserved structure/activity relationships that are associated with genomic and non-genomic actions of steroid hormones. Hence, how a steroid works at a substrate may depend upon small differences in the steroids’ metabolism and structure. To a great extent, steroid effects may be determined by relatively few enzymatic steps performed by a small group of enzymes that result in the generation of all steroid hormones. In many respects, the metabolic pathway of a steroid determines the nature of the steroid signal and its degree of amplification (Rubinow and Schmidt, 1996). The final effect of a steroid hormone is determined by enzymatic activity in the target cell. For example, the composition of body hair is to a much higher extent related to 5α-reductase activity in the hair follicles than to plasma T levels (Lookingbill, Demers, Wang, Leung, Rittmaster and Santen, 1991).
3.3. Androgen Metabolism
Testosterone and many other androgens (i.e., those possessing a 3-keto 4-A configuration and a methyl group at the 19th carbon) may, via aromatization to estrogens (Akhtar, Njar and Wright, 1993; Graham-Lorence, Amameh, White, Peterson and Simpson, 1995; Korzekva, Trager, Mancewics and Osawa, 1993; Korzekva, Trager, Smith, Osawa and Gilette, 1991), also stimulate estrogen receptors. Indeed, in situ aromatization of androgens represents is an important metabolic event. Connolly, Roselli, and Resko (1990) have demonstrated that adult male guinea pig brains contained higher quantities of androgen aromatase than female brains. Androgen aromatase is concentrated in the limbic system and hypothalamus (amygdala, preoptic area, septum, hippocampus, and medial basal hypothalamus), whereas low levels were consistently found in cortical tissue (McEwen, 1980). Furthermore, female rats treated with T show a more marked enhancement of social aggression than rats treated with the nonaromatizable androgen dihydrotestosterone (DHT). Combined treatment with DHT and estrogen resulted in the same degree of increased aggression as T treatment, suggesting that the activation of estrogen receptors and androgen receptors may work synergistically (van de Poll, van Zanten, and de Jonge, 1986). Many of the synthetic AS are aromatizable (Bahrke, Yesalis, and Wright, 1990), a quality responsible for gynecomastia. Thus, central effects of AS may be mediated through genomic actions at androgen and estrogenic receptors. Furthermore, synthetic AS and their metabolites do not only bind to androgen or estrogen receptors but also to glucocorticoid and progestin receptors (Janne, 1990). Consequently, the effects of AS are far from purely androgenic and may involve actions at multiple genomic and non-genomic substrates.
Findings from our laboratory demonstrate that chronic administration of T and/or its 5α-reduced metabolites may have actions via both genomic and non-genomic substrates. Administration of 1 mg of T, DHT or its metabolite 3α-Androstanediol (3α-diol) daily for 3 days reduced seminal vesicle weight (an androgen receptor dependent measure) and decreased androgen receptor binding in the hypothalamus. Concomitant with these androgen receptor mediated effects, the sensitivity of GBRs in the hippocampus was increased (Frye, Park, Tanaka, Rosellini, and Svare, 2001). As these androgen regimen are capable of having actions at both genomic (intracellular androgen receptors) and non-genomic (GBRs or other) substrates, an important question is what are the actions of androgens that underlie its hedonic effects.
4. Hypothesis
Our research to begin to address this question has focused on the role of T’s 5α-reduced metabolite, 3α-diol, which typically has actions at GBRs, rather than intracellular androgen receptors. The purpose of these studies is to better understand the abuse liability and potential of androgens’ by investigating the effects and mechanisms in mediating their interoceptive effects. First, research is summarized that has investigated the extent to which T’s 5α-reduced metabolites may have actions in the nucleus accumbens (NA) to mediate hedonic effects. Second, experiments that address whether these effects of androgens require actions at intracellular androgen receptors is presented. Third, effects of facilitating and/or blocking androgens actions at GBRs in the NA is discussed. Finally, the results of effects of dopaminergic lesions to the NA on androgen’s effects are described. These findings that will be reviewed here suggest that 3α-diol’s actions in the NA via GBRs and/or dopaminergic neurons, rather than intracellular androgen receptors, may underlie hedonic effects (Figure 1).
Figure 1.
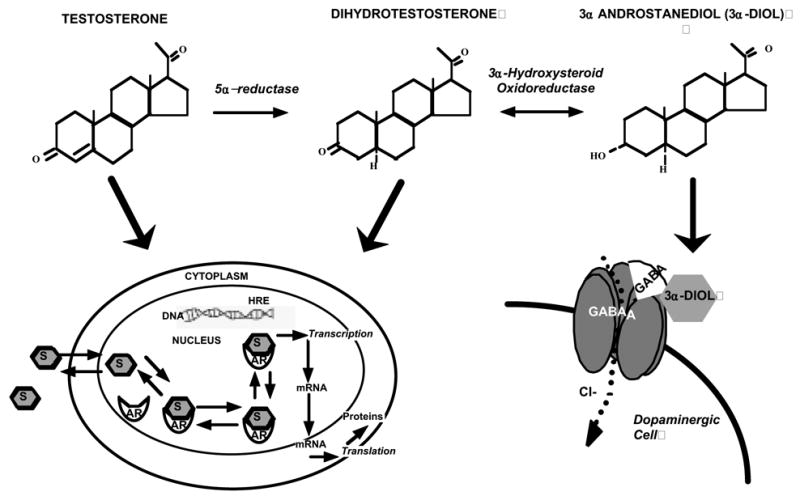
Metabolism pathway and substrates for androgen action. Testosterone (T) is metabolized to dihydrotesterone (DHT) via 5α-reductase, which is converted to 3α-androstanediol (3α-diol) via 3α-hydroxysteroid oxidoreductase. T and DHT bind to intracellular androgen receptors(ARs) and 3α-diol binds GBRs, which maybe localized to dopaminergic neurons in the nucleus accumbens.
4.1. Are effects of T on conditioned place preference mediated by actions of its 5α-reduced metabolites?
Conditioned place preference (CPP) has been used in many experiments to examine hedonic effects of drugs of abuse (Scoles and Siegel, 1986). In several studies of CPP and androgens, T did condition a place preference (Packard, Cornell, and Alexander, 1997; Alexander, Packard and Hines, 1994). However, there is considerable variability in this effect, with some studies reporting it only with extremely high dosages of T and not with lower dosages (Caldarone, Stock, Abrahamsen, Boechler, Svare, and Rosellini, 1996), and in males but not in females (DeBeun, Jansen, Slangen, and Van de Poll, 1992). In some experiments rats were tested soon (30 min) after T administration, which may be insufficient time to enable T to be metabolized by 5α-reductase to DHT and by 3α-oxidoreductase to 3α-diol. As discussed below, the manner in which androgens are given, e.g., dosage, bioavailability, route of administration, and/or vehicle may underlie some of T’s variability on CPP.
Evidence from the literature to support the notion that variability in T’s positive hedonic properties may be related to capacity to form 3α-diol include the following. First, when administered systemically in oil vehicle 30 minutes prior to CPP chamber exposure only a high dosage (1 mg) of T was effective at inducing a CPP and lower dosages (10 or 100 μg) were not (Caldarone, Stock, Abrahamsen, Boechler, Svare, and Rosellini, 1996). Higher dosages of T would more readily facilitate the metabolism of T even when administered in oil vehicle, compared to the lower dosages. Second, when administered systemically in a non-oil, encapsulization vehicle, 30 minutes prior to CPP chamber exposure, a CPP was observed in rats administered 1200 or 800, but not 400 μg/kg T (Alexander, Packard, and Hines, 1994). These dosages of T produced supraphysiological levels of circulating T in male rats (Taylor, Weiss, and Pitha, 1989), suggesting that the rewarding affective properties of T depend upon circulating levels of hormones that are markedly above baseline concentrations. Third, when the bioavailability of T is further increased by intrabrain infusion of T in a molecular encapsulization vehicle, 0.25 or 0.50, but not 0.125 μg/kg of T into the NA of male rats immediately prior to CPP chamber exposure enhanced place preference (Packard et al., 1997). The metabolism enzymes, 5α-reductase and 3α-hydroxysteroid dehydrogenase, have been localized to the NA (Mellon, 1994) and could have rapidly converted the higher dosages of T to 3α-diol to produce these effects. The studies described below summarize our research progress to empirically address the question that some of T’s hedonic effects may be related to formation of 3α-diol.
4.1.1. Systemic 3α-diol Regimen that Enhance CPP Increase 3α-diol > DHT> T in Plasma and NA
We have shown that systemic 3α-diol administration conditions a place preference more effectively than does systemic administration of DHT or T (Frye et al., 2001). Briefly, administration of 3α-diol, but neither T nor DHT, to intact male Long-Evans rats, 1.0 mg daily for 6 days, 30 minutes prior to exposure to the non-preferred side of the CPP chamber, produced significant increases in preference for the non-preferred side of the chamber at testing (Figure 2, right), compared to baseline preference (which did not differ across groups-Figure 2, left). Notably, circulating concentrations of 3α-diol were increased most in 3α-diol > DHT > T-administered rats, compared to vehicle-administration (Figure 2-inset).
Figure 2.
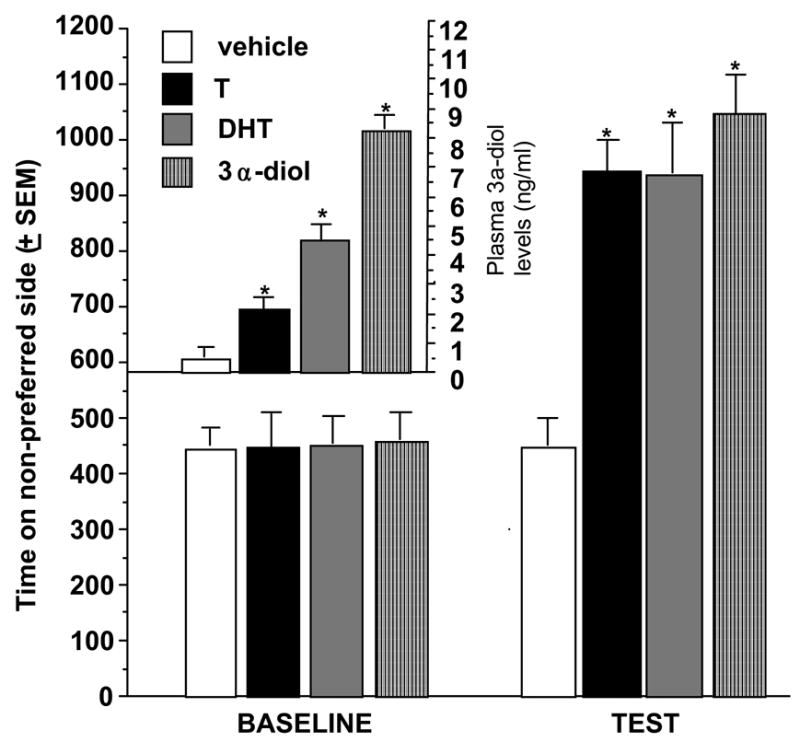
Rats administered subcutaneous T, DHT, or 3α-diol spend an increased amount of time on the non-preferred side of the conditioned place chamber and have increased levels of 3α-diol in the nucleus accumbens (inset). * P<0.05
Testosterone implants to the NA can condition a place preference (Packard et al., 1997); however, it is unclear whether this may be due to actions of its 5α-reduced metabolites. We have investigated effects of systemic androgens on CPP and levels of androgens in the NA (Rosellini et al. 2001; Frye, Rhodes, Rosellini and Svare, 2002). Rats were systemically administered 1 mg of T, DHT, or 3α-diol, 30, 90, or 180 mins prior to exposure on conditioning days to the non-preferred side of a CPP chamber. All rats administered 3α-diol demonstrated CPP and had the highest concentrations of 3α-diol in the NA at each of the temporal pairings tested. The percentage of rats spending more time on the non-preferred side of the CPP chamber on the test day was greatest with androgen regimens that increased levels of 3α-diol in the NA. These findings demonstrate that 3α-diol concentrations are increased in the NA by androgen regimens that produce CPP (3α-diol >DHT>T≫>vehicle) (Rosellini et al. 2001).
4.1.2. Administration of T, DHT or 3α-diol Directly to the Shell of the NA Enhances CPP
We have also investigated effects on CPP of directly stimulating the NA with androgens (Frye, Rhodes, Rosellini and Svare, 2002). Rats were administered implants of T, DHT or 3α-diol to the NA immediately prior to placement in the CPP apparatus on conditioning days. Implants of T, DHT, or 3α-diol immediately prior to exposure to the non-preferred side of chamber significantly increased time spent on the non-preferred side of the chamber on the test day (Figure 3, right), as compared to baseline (Figure 3, left),. Notably, this effect was only produced by androgenic stimulation of the shell, but not the core, of the NA (Figure 3, inset). Results of this experiment and the latter suggest that androgen regimen that increase 3α-diol concentrations in the accumbens can enhance CPP and direct implants of T, DHT, or 3α-diol to the shell of the NA elicit CPP; however, whether formation of 3α-diol is required for these effects was not established. The data presented below, which heretofore have not been published, address the importance of androgen metabolism, actions at androgen receptors, GBRs, and dopamine targets.
Figure 3.
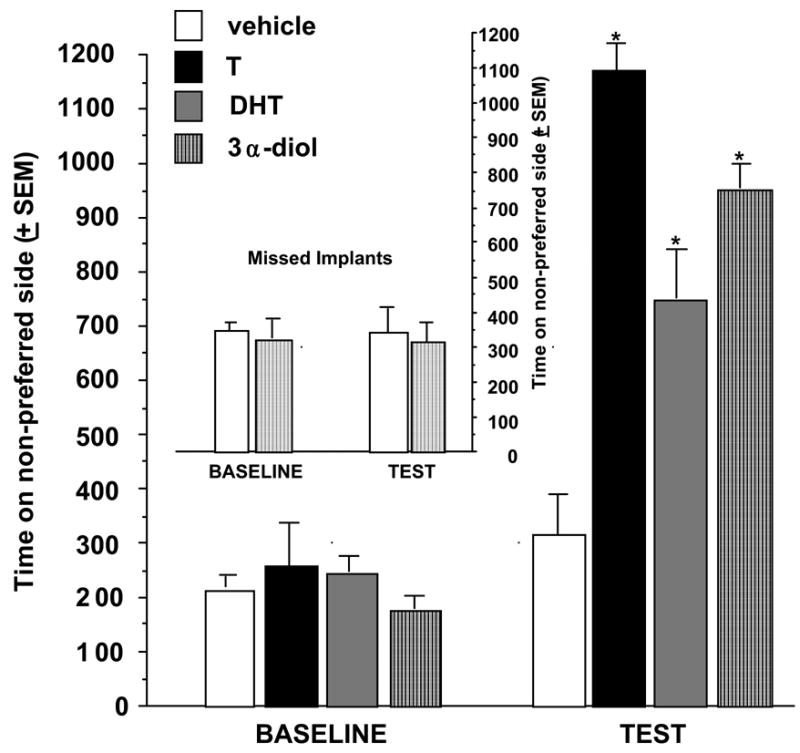
Rats administered T, DHT, or 3α-diol to the shell of the nucleus accumbens spend an increased amount of time on the non-preferred side of the conditioned place chamber. 3α-diol administered to the core of the nucleus accumbens does not produce a place preference (i.e. no difference from vehicle administration to the core of the nucleus accumbens (inset). * P<0.05
4.1.3. Systemic Metabolism Inhibitors Attenuates T-Induced CPP & Levels of 3α-diol in Whole Brain
Testosterone is metabolized by 5α-reductase to DHT; additional conversion by 3α-hydroxysteroid dehydrogenase forms 3α-diol. Findings above suggest systemic administration of 3α-diol > DHT > T paired with exposure to the non-preferred side of the CPP chamber produces the greatest effects on CPP and is associated with increases in 3α-diol in the NA during conditioning. These findings are consistent with the hypothesis that metabolism of T or DHT to 3α-diol in the NA is essential in mediating the hedonic effects of androgens. However, we have also directly tested the hypothesis that blocking T or DHT’s metabolism to 3α-diol will attenuate effects of the androgens on CPP. Systemic administration of 5α-reductase (finasteride) or by 3α-hydroxysteroid dehydrogenase (indomethacin) inhibitors, prior to T and/or DHT administration, attenuates CPP and whole brain 3α-diol concentrations (Rosellini et al., 2003; Figure 4, left). However, this does not address whether metabolism in the NA alone is required for hedonic effects of androgens.
Figure 4.
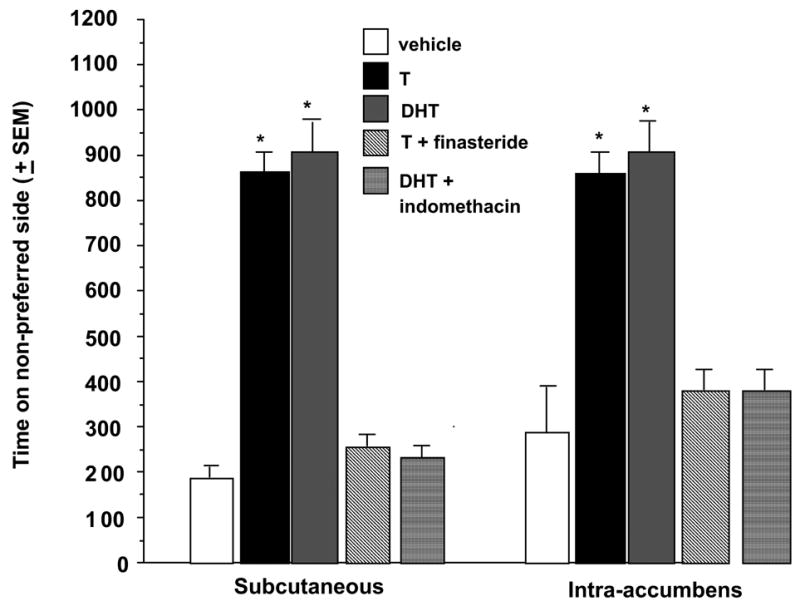
Rats administered T or DHT and subcutaneous (left) or intra-accumbens (right) metabolism inhibitors spend less time on the non-preferred side of the conditioned place chamber compared to rats administered T or DHT and vehicle. * P<0.05
4.1.4. Intra-accumbens Metabolism Inhibitors Attenuates T-Induced CPP & 3α-diol Levels in NA
We have investigated whether androgen metabolism inhibitors to the NA can attenuate the hedonic effects of systemic T. First, finasteride applied to the NA prior to SC T paired with exposure to the non-preferred side of the chamber attenuates DHT and 3α-diol formation in the NA during conditioning trials as well and T’s facilitation of CPP at test time (Figure 4, right). Second, indomethacin administered directly to the NA prior to SC DHT attenuates 3α-diol formation in the NA during conditioning and subsequently DHT’s facilitation of CPP at testing (Figure 4, right). Third, the similar effects of systemic or intra-accumbens administration of metabolism inhibitors is very similar which implies that blocking formation of 3α-diol in the NA is sufficient to attenuate T’s effects on CPP. Fourth, similar effects of blocking T’s and/or DHT’s metabolism suggest that 3α-diol, rather than DHT, in the accumbens is the active androgen underlying some effects on CPP.
4.2. Are actions at androgen receptors in the NA necessary for androgens effects on CPP?
The findings described above that 3α-diol and its prohormones T and DHT can induce CPP when applied to the NA, a region of the brain with few intracellular androgen receptors (Stumpf and Sar, 1976), suggests that these androgens may exert some of their hedonic effects via non-genomic actions. For example, implants of T to the medial preoptic area or the NA condition a place preference (Packard, Cornell, and Alexander, 1997). As well, implants of T or 3α-diol also condition a place preference when applied to the NA (Frye et al., 2002; Rosellini et al., 2001). Although there are many intracellular androgen receptors in the medial preoptic area that could be substrates for T’s effects, there are few intracellular androgen receptors in the NA (Stumpf and Sar, 1976). As well, we have shown that implants of T or 3α-diol and immediate pairing with the non-preferred site of the chamber enhances CPP. Testosterone and DHT both bind readily to intracellular androgen receptors, while 3α-diol is devoid of affinity for androgen receptors in physiological concentrations (Cunningham, Tindall and Means, 1979; Verhoeven, Heyns and DeMoor, 1975). Although there are few androgen receptors that have been identified in the NA and the rapid effects of 3α-diol to enhance CPP would seem to preclude sufficient time for androgen receptor mediated changes in transcription, it is necessary to investigate whether 3α-diol‘s actions at for androgen receptors are necessary for its hedonic effects. Pharmacological concentrations of 3α-diol that enhance CPP could override 3α-diol’s selective low affinity for androgen receptors and thereby potentially produce effects produce effects via the few androgen receptors in the NA or in other regions of the brain such as the hippocampus.
4.2.1. Systemic Administration of an Androgen Receptor Blocker Does Not Attenuate 3α-diol -Induced CPP
We investigated whether 3α-diol has actions via intracellular androgen receptors to mediate CPP. During conditioning, rats received SC flutamide (10 mg) or vehicle (sesame oil) two hours prior to SC 3α-diol (10 mg/kg) or vehicle (propylene glycol), which were administered immediately before placement in the non-preferred side of the CPP chamber. As we have previously demonstrated, rats that received vehicle and 3α-diol spent a significantly increased amount of time on the non-preferred side of the chamber at test time compared to vehicle-vehicle or vehicle-flutamide controls. Co-administration of flutamide with 3α-diol does not attenuate 3α-diol-induced CPP, and did not have intrinsic effects (Figure 5, left). Although, these findings suggest that 3α-diol’s actions to enhance CPP occur independent of actions at intracellular androgen receptors in any part of the brain, the flutamide regimen utilized has been demonstrated to block physiological, rather than pharmacological regimen of androgens. As such, we also wanted to investigate effects of a pharmacological flutamide regimen to counter pharmacological effects of 3α-diol.
Figure 5.
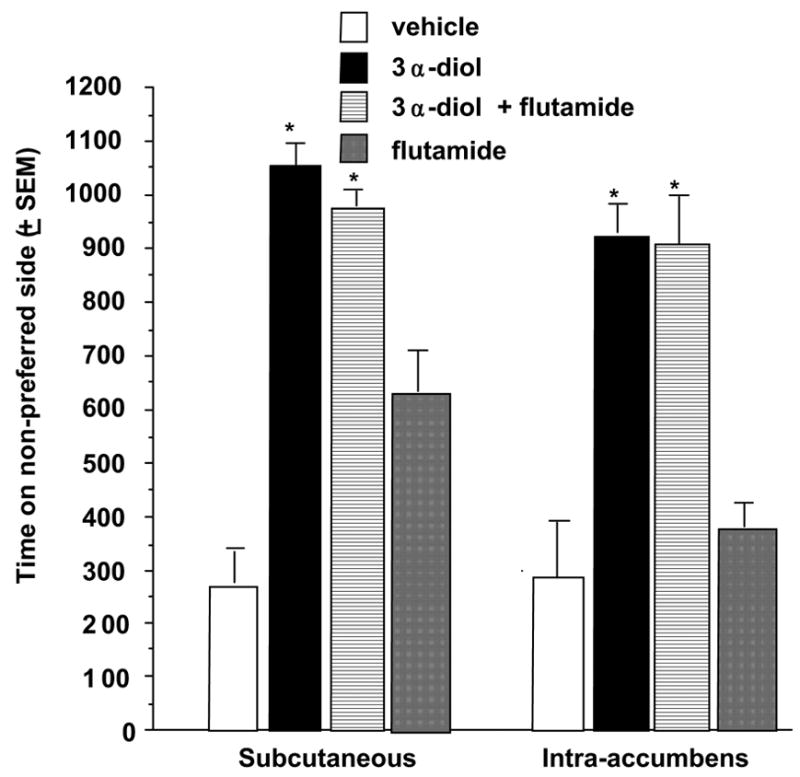
Rats administered subcutaneous 3α-diol and subcutaneous (left) or intra-accumbens (right) androgen receptor antagonist (fluamide) did not spend less time on the non-preferred side of the conditioned place chamber compared to rats administered 3α-diol and vehicle. * P<0.05
4.2.2. Intra-accumbens Administration of an Androgen Receptor Blocker Does Not Attenuate T-Induced CPP & 3α-diol Levels in NA
We investigated whether 3α-diol has actions via intracellular androgen receptors in the NA to mediate CPP. Rats were administered flutamide or empty control implants to the NA 2 hours prior to SC 3α-diol or vehicle immediately prior to placement in the non-preferred side of the CPP chamber. As previously demonstrated, rats administered 3α-diol exhibited a place preference for the non-preferred side of the chamber on test day compared to vehicle. Administration of flutamide implants directly to the NA with 3α-diol did not attenuate 3α-diol-induced CPP, nor did it effect CPP itself in the absence of 3α-diol (Figure 5, right). Although these data indicate that actions of 3α-diol in the NA to mediate CPP may be independent of its actions at ARs, the neurobiological substrates in the NA that mediate 3α-diol’s hedonic effects need to be revealed.
4.3. Are actions at GBRs in the NA necessary for androgens effects on CPP?
There are data to suggest that steroids also may influence cellular activity in a “non-genomic” fashion or through means other than traditional actions at intracellular steroid receptors (Brann, Hendry and Mahesh, 1995). Indeed, T, its metabolites, and many AS, do not bind with a high affinity to traditional intracellular androgen receptors (Cunningham, Tindall, and Means, 1979; Verhoeven, Heyns, and DeMoor, 1975). Anabolic steroids upregulate androgenic substrates in several non-classical target sites, e.g., the CA- I region of the hippocampus and the VTA (Teledgy 1987). Likely mechanisms for these non-classical actions are: (a) changes in membrane fluidity; (b) steroid hormones acting on receptors on plasma membranes; (c) steroid hormones regulating GBRs on plasma membranes; and (d) activation of dopamine substrates. The GABA system has been implicated as a substrate for illicit drug use, and steroids have been shown to interact with GBRs (Majewska, Harrison, Schwartz, Barker, and Paul, 1986; Gee, 1988; Frye, van Keuran, and Erskine, 1996) and dopamine systems (Mani et al. 1996; Frye, Bayon and Vongher, 1999). Although the former (a and b) possibilities cannot be ruled out, we have not investigated these possibilities to date because such substrates have not been sufficiently defined as to warrant extensive investigation at this time. Our research to begin to address the possibility that GBRs (this section) or dopaminergic neurons (d; next section) are possible “non-genomic” substrates for androgens’ hedonic effects is described below.
Evidence for GABA involvement in AS mechanism of action include that withdrawal is characteristic of drugs of abuse that act at GABA. Withdrawal from chronic exposure to other psychoactive GABA-active agents, such as ethanol, benzodiazepines, and barbiturates, can result in psychomotor disruption and anxiogenic effects (Buck, McQuillen, and Harris, 1991; Hauser, Devinsky, DeBellsi, Theodore, and Post, 1989; McCaslin and Morgan, 1988). Cessation or diminished use of AS has been associated with depressive symptoms (Corrigan 1996). Other symptoms related to the loss of positive psychological effects of AS include listlessness; apathy; loss of appetite, libido and self-esteem; feelings of anxiety; difficulty in concentrating; and mood swings (Bahrke, Yesalis, and Wright 1990; Corrigan, 1996, Uzych 1992).
There is evidence that androgens, particularly 3α-diol, may have actions through GBRs. Several steroids can alter GBR function, and the 5α-reduced, 3α-hydroxylated structure of 3α-diol meets the requirements of the most potent steroid modulators of GBRs (Belelli, Lan, and Gee, 1990; Beyer, Gonzalez-Mariscal, Equibar, and Gomora, 1988). Although T is not particularly effective at altering GBRs, 3α-diol is (Gee, 1988; Frye, Duncan, Basham, and Erskine, 1996) and GBRs have been localized to the NA (Zhang et al., 1991). Chronic administration of T and metabolites alters the sensitivity of GBRs in the hippocampus (Frye, Park, Tanaka, Rosellini, and Svare, 2001). AS administration stabilizes GBRs in a moderate affinity state for benzodiazepine binding and reduces the EC50 for GABA-stimulated chloride influx (Masonis and McCarthy, 1995, 1996). GABA-stimulated chloride influx and muscimol binding are increased in animals administered 3α-diol (Frye, van Keuran, Rao, and Erskine, 1996a, b). Co-administration of a GBR antagonist, bicuculline, counters 3α-diol-induced changes in social (Frye, van Keuran, Rao, and Erskine, 1996b; Frye, van Keuran, and Erskine, 1996), and affective (Frye, van Keuran, Rao, and Erskine, 1996a) behavior. Furthermore, rats administered the AS, dianabol, demonstrated increases in levels of 3α-diol in the NA and greater GABA-stimulated chloride influx in cortical synnaptoneurosomes than did vehicle controls or rats that had dianabol discontinued and were withdrawing (see Table 1).
Table 1.
The AS Dianabol Enhances 3α-Diol Levels, and GABA-stimulated Chloride Influx
| Treatment Group | 3α-Diol Levels | Emax | EC50 |
|---|---|---|---|
| Vehicle | 2.25 + 0.5 ng/ml | 24.4 + 1.4 | 11.2 + 0.7 |
| Dianabol | 3.00 + 0.2 ng/ml * | 34.8 + 3.3* | 9.3 + 0.7* |
| Dianabol | 1.40 + 0.2 ng/ml* | 28.1 + 4.0 | 11.8 + 1.0 |
| Withdrawal |
4.3.1. Intra-accumbens GBR Antagonist Attenuates 3α-diol-Induced CPP
These background data that show that 3α-diol has actions via GBRs suggest that androgens’ hedonic effects may be mediated in part via GBRs in the NA. To further determine the role of GBRs in the NA to mediate some of the effects of androgens on CPP, the following experiments were conducted. Rats received either a high dosage of 3α-diol (1 mg) that has been used previously to induces CPP or vehicle priming, in conjunction with infusions of vehicle or the GBR antagonist, bicuculline (50 ng) to the NA immediately prior to exposure to the conditioning chamber. As previously demonstrated, rats administered 1 mg of systemic 3α-diol showed an increase in time spent on the non-preferred side of the conditioning chamber on test day compared to rats that received vehicle. Co-administration of bicuculline, a GBR antagonist, to the NA, attenuated 3α-diol-induced CPP. Neither bicuculline administration alone, nor vehicle administration alone, had effects on CPP (Figure 6). Although these findings suggest that blocking 3α-diol’s actions at GBRs can attenuate CPP; a truer measure of whether 3α-diol can work through GBRs to produce hedonic effects would involve facilitation of such effects.
Figure 6.
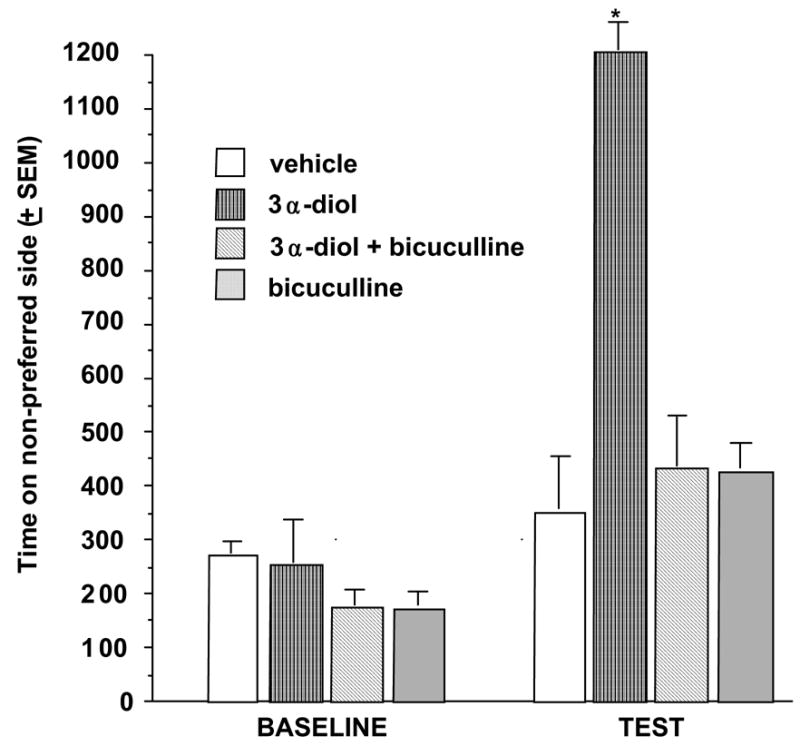
Rats administered subcutaneous 3α-diol and intra-accumbens administration of the GBR antagonist, bicuculline, decreased amount of time on the non-preferred side of the conditioned place chamber. * P<0.05
4.3.2. Intra-accumbens GBR Agonist Enhances 3α-diol-Induced CPP
In this experiment, we examined whether activating GBRs could enhance effects of subthreshold 3α-diol stimulation to produce CPP. Rats were systemically administered a lower dosage of 3α-diol (1 mg/kg) that alone is insufficient to induce CPP or vehicle-priming. In addition, the GBR agonist, muscimol, or saline vehicle was infused to the NA immediately prior to being put in the conditioning chamber. As expected, rats administered the lower dosage of 3α-diol (1 mg/kg) spend a similar amount of time on the non-preferred side of the chamber on test day as did vehicle-primed rats. However, co-administration of muscimol, a GBR agonist, to the NA together with this sub-threshold dosage of 3α-diol, was sufficient to induce a CPP.
Muscimol alone did not produce a CPP and was no more effective than vehicle at enhancing CPP (Figure 7). These data, which show that 3α-diol and muscimol may have synergistic actions as GBR agonists, suggest that 3α-diol may have effects in the NA on CPP in part through agonist-like actions at GBRs.
Figure 7.
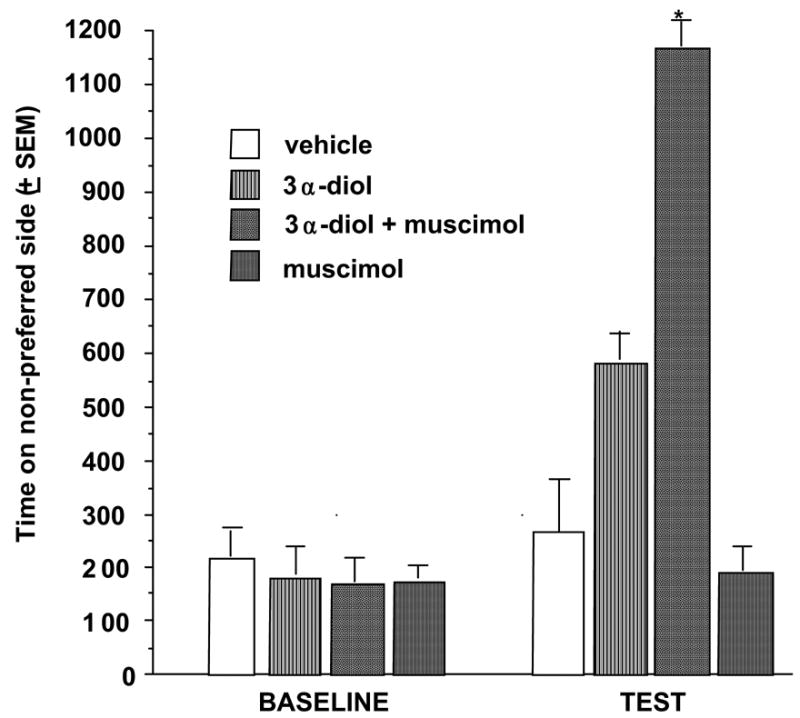
Rats administered subcutaneous 3α-diol and intra-accumbens administration of the GBR agonist, muscimol, increased amount of time on the non-preferred side of the conditioned place chamber. * P<0.05
4.4. Are actions of 3α-diol at dopaminergic neurons essential for 3α-diol-enhanced CPP?
Interoceptive effects of androgens may involve direct or indirect actions at dopaminergic neurons. Evidence in support of this is as follows. We, and others (Packard, Cornell, and Alexander, 1997), have found that androgens can condition a place preference when applied directly to the NA, an area of the brain containing dopamine neurons. Administration of AS can produce EEG activity similar to that of psychostimulants (Itil, 1976; Itil, Cora, Akpinar, Herrmann, and Patterson, 1974; Stenn, Kaliber, Vogel, and Broverman, 1972). Androgens can act on the dopamine reward system in a manner similar to cocaine or other stimulants (Alderson, and Baum, 1981; Goudsmit, Feenstra, and Swaab, 1990; Jalilian-Tehrani, Karakiulakis, LeBlond, Powell, and Thomas, 1982; Mitchell and Stewart, 1989; Kaskin and Kleber, 1989; Vermes, Varszegi, Toth, and Telegdy, 1979). Notably, the mesolimbic dopamine system is often considered the final common pathway for many dependence-producing drugs (Koob and LeMoal, 1997; Koob, 1992; Wise and Bozarth, 1987; Robinson and Berridge, 1993). There are some reports of AS use enhancing sexual desire and pleasure in people (Greenblatt and Karpas, 1983; Taylor, 1987), effects that are known in rats to be associated with increased levels of dopamine in the NA (Moses, Loucks, Watson, Matuszewich, and Hull, 1995; Lorrain, Riolo, Matuszewich, and Hull, 1999). We have begun to address whether another possible non-genomic substrate through which androgens may have their hedonic effects is via dopaminergic neurons in the NA.
4.4.1. 6-OHDA Lesions That Completely Eradicate the Shell of the NA Prevent 3α-diol’s Enhancement of CPP
In this study, we hypothesized that 6-hydroxydopamine (6-OHDA) lesions to the dopamine neurons of the NA would decrease the conditioning affects normally seen with 3α-diol administration. 3α-diol was administered subcutaneously (SC) 1 mg daily for the six days of pairings with the non-preferred side of the chamber to 6-OHDA lesioned male Long-Evans rats, 30 minutes prior to exposure to the non-preferred side of the CPP chamber. 3α-diol conditioned a place preference in rats with partial lesions to the shell and core of the NA, but no conditioning was seen in rats that had complete lesions to the shell of the NA (Figure 8). These data suggest 3α-diol’s enhancing effects on CPP may require actions at dopaminergic neurons in the shell of the NA. This suggestion is consistent with the conclusion reached by McBride, Murphy, and Ikemoto (1999) in their thorough review of the literature on brain reinforcement mechanisms that intracranial self administration and conditioned place preference studies with “psychostimulants, morphine, and PCP all produce reinforcing effects within the NA and that their (reinforcing) effects are observed mainly in the shell” (p 141). As such, these data, and the findings discussed above that 3α-diol implants to the shell, but not the core, of the NA enhance CPP, also underscore other findings presented, which indicated that 3α-diol can have positive hedonic effects and that such effects may involve actions at GBRS and dopamine neurons.
Figure 8.

Rats administered subcutaneous 3α-diol with 60HDa lesions to the shell (but not core; see inset) of the nucleus accumbens spend less time on the non-preferred side of the conditioned place chamber. * P<0.05
5. Other substrates for AS
This review of potential targets for andogens’ actions for hedonic effects is by no means exhaustive. There are clearly numerous substrates at which androgens and AS could have actions to produce positive hedonic effects. One that may be particularly relevant that is beyond the scope of the present review, but warrants further consideration, is the opioid system. Many dopamine neurons in the mesolimbic pathways are juxtaposed with opioid containing neurons. Among the reported interoceptive effects of AS in people are effects similar to that seen in narcotic users, such as euphoric mood (Pope and Katz, 1988, 1994) and elevation of mood in depressed patients (Bahrke, Yesalis, and Wright, 1990). Indeed, there is evidence that AS use may be increasingly considered a “gateway” drug for later use of opiates (McBride, Williamson, and Petersen, 1996; Pope, Phillips, and Olivardia, 2000). In 1999, 10% of men being treated for opiate addiction in a rehabilitation program indicated that AS use preceded their opiate use. In 1990, only 1% of men in treatment for dependence in the same rehabilitation program reported any preceding AS use (Arvary and Pope, 2000). There is also evidence that withdrawal from AS may share many common characteristics of opiate withdrawal, such as the initial phase of withdrawal characterized by increased pulse and blood pressure, sweats, chills, nausea, and dizziness (Kashkin and Kleber, 1989; Tennant, Black, and Voy, 1988).
6. Summary
In summary, androgen and AS use is widespread and increasing. The background literature regarding effects of AS underscore the notion that AS can have effects that are akin to that of other drugs of abuse. Use of AS is associated with adverse illicit use of other drugs of abuse, as well as physiological and behavioral consequences (including violence and aggression). Anabolic-androgenic steroids use may lead to addiction, dependence, and withdrawal such that use is often continued despite short- and long-term health risks. Understanding the pharmacological effects of AS is important, but attempts have been confounded both by the complex receptor-and non-receptor-bound actions of AS and the lack of basic understanding of androgens’ actions and their hedonic effects and neurobiological mechanisms. The research described addresses whether T, a widely used AS, has positive euphorogenic effects which may contribute to their abuse liability in part through non-genomic actions. The implications of this research are that AS and/or T may produce some of their positive hedonic effects by enhancing 3α-diol production, which in turn have actions at GBRs in the NA, which synapse on dopaminergic neurons, to produce positive hedonic effects. Further research is needed to ascertain other hedonic effects and mechanisms of androgens.
Acknowledgments
Grant support was provided by the National Science Foundation (IBN03-16083). Technical assistance provided by Luigi DiRienzo, Frances Melendez, Laurie Spofford, Kanako Sumida, Nicole Schmidt, and Alicia Walf is greatly appreciated. This research has also benefited from the input of colleagues including Drs. Rhodes, Rosellini, and Svare.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Akhtar M, Njar V, Wright J. Mechanistic studies of aromatese and related c-c-bonds cleaving P450 enzymes. J Steroid Biochem Mol Biol. 1993;44:375–387. doi: 10.1016/0960-0760(93)90241-n. [DOI] [PubMed] [Google Scholar]
- Albrecht RR, Anderson WA, McKeag DB. Drug testing of college athletes. The issues Sports Med. 1999;14:349–352. doi: 10.2165/00007256-199214060-00002. [DOI] [PubMed] [Google Scholar]
- Alderson LM, Baum MJ. Differential effects of gonadal steroids on dopamine metabolism in mesolimbic and nigro-striatal pathways of male rat brain. Brain Research. 1981;218:189–206. doi: 10.1016/0006-8993(81)91300-7. [DOI] [PubMed] [Google Scholar]
- Alexander G, Packard M, Hines M. Testosterone has rewarding affective properties in male rats: implications for the biological basis of sexual motivation. Behav Neurosci. 1994;108:424–428. doi: 10.1037//0735-7044.108.2.424. [DOI] [PubMed] [Google Scholar]
- Altschule MD, Tilletson KJ. The use of testosterone in the treatment of depression . New England J Med. 1948;239:1036–1038. doi: 10.1056/NEJM194812302392704. [DOI] [PubMed] [Google Scholar]
- Anderson WA, Albrecht RR, McKeag DB, Hough DO, McGrew CA. A national survey of alcohol and drug use by college athletes. Phys Sports med. 1991;19:91–104. [Google Scholar]
- Arvary D, Pope HG. Anabolic-androgenic steroids as a gateway to opioid dependence. N Engl J Med. 2000;342:1532. doi: 10.1056/NEJM200005183422018. [DOI] [PubMed] [Google Scholar]
- Bahrke M. International conference on abuse and trafficking of anabolic steroids. International J Drug Policy. 1995;5:23–26. [Google Scholar]
- Bahrke M, Wright JE, Strauss RH, Catlin DH. Psychological moods and subjectively perceived behavioral and somatic changes accompanying anabolic-androgenic steroid use. American J Sports Med. 1992;20:717–724. doi: 10.1177/036354659202000613. [DOI] [PubMed] [Google Scholar]
- Bahrke M, Yesalis C, Brower K. Anabolic-androgenic steroid abuse and performance-enhancing drugs among adolescents. Child and adol psych clinics N America. 1998;1:821–838. [PubMed] [Google Scholar]
- Bahrke M, Yesalis C, Wright J. Psychological and behavioral effects of endogenous testosterone and anabolic-androgenic steroids. Sports Med. 1996;22(6):367–390. doi: 10.2165/00007256-199622060-00005. [DOI] [PubMed] [Google Scholar]
- Bahrke M, Yesalis C, Wright J. Psychological and behavioral effects of endogenous testosterone levels and anabolic-androgenic steroids among males. Sports Med. 1990;10:303–337. doi: 10.2165/00007256-199010050-00003. [DOI] [PubMed] [Google Scholar]
- Belelli D, Lan NC, Gee KW. Anticonvulsant steroids and the GABA benzodiazepine receptor-chloride ionophore complex. Neurosci Biobehav Rev. 1990;14:315–322. doi: 10.1016/s0149-7634(05)80041-7. [DOI] [PubMed] [Google Scholar]
- Beyer C, Gonzalez-Mariscal G, Equibar JR, Gomora P. Lordosis facilitation in estrogen primed rats by intrabrain injection of pregnanes. Pharmacol Biochem Behav. 1988;31:919–926. doi: 10.1016/0091-3057(88)90405-4. [DOI] [PubMed] [Google Scholar]
- Bhasin S, Storer TW, Berman N, Callegari C, Clevenger B, Phillips J, Bunnell TJ, Tricker R, Shirazi A, Casaburi R. The effects of supraphysiologic doses of testosterone on muscle size and strength in normal men. NE J Med. 1996;335:1–7. doi: 10.1056/NEJM199607043350101. [DOI] [PubMed] [Google Scholar]
- Bonson KR, Garrick NA, Murphy DL. Evidence for a withdrawal syndrome following chronic administration of an anabolic steroid to rats. Society for Neurosci Abstr. 1994;20:1527. [Google Scholar]
- Brann DW, Hendry LB, Mahesh VB. Emerging diversities in the mechanism of action of steroid hormones. Journal of Steroid Biochem Molec Bio. 1995;52:113–133. doi: 10.1016/0960-0760(94)00160-n. [DOI] [PubMed] [Google Scholar]
- Brower KJ, Blow FC, Beresford TP, Fuelling C. Anabolic-androgenic steroid dependence. Clin Psych. 1989;50:31–33. [PubMed] [Google Scholar]
- Brower KJ, Blow FC, Eliopulos GA, Beresford TP. Anabolic androgenic steroids and suicide. Am J Psychiat. 1989;146:1075. doi: 10.1176/ajp.146.8.1075a. [DOI] [PubMed] [Google Scholar]
- Brower KJ, Blow FC, Young JP, Hill EM. Symptoms and correlates of anabolic-androgenic steroid dependence. Br J Addict. 1991;86:759–768. doi: 10.1111/j.1360-0443.1991.tb03101.x. [DOI] [PubMed] [Google Scholar]
- Brower KJ, Eliopulos GA, Blow FC, Catlin DH, Beresford TP. Evidence for physical and psychological dependence on anabolic androgenic steroids in eight weight lifters. Am J Psych. 1990;147:510–512. doi: 10.1176/ajp.147.4.510. [DOI] [PubMed] [Google Scholar]
- Buck KJ, McQuillen SJ, Harris RA. Modulation of GABA receptor operated chloride channels by benzodiazepine inverse agonists is related to genetic differences in ethanol withdrawal seizure severity. J Neurochem. 1991;57:2100–2105. doi: 10.1111/j.1471-4159.1991.tb06428.x. [DOI] [PubMed] [Google Scholar]
- Buckley WE, Yesalis CE, Friedl KE, Anderson WA, Streit AL, Wright JE. Estimated prevalence of anabolic steroid use among male high school seniors. JAMA. 1988;260:3441–3445. [PubMed] [Google Scholar]
- Caggiula AR. Analysis of copulation-reward properties of posterior hypothalamic stimulation in male rats. J Comparative and Physio Psych. 1970;70:399–412. doi: 10.1037/h0028706. [DOI] [PubMed] [Google Scholar]
- Caggiula AR, Hoebel BG. Copulation reward site” in the posterior hypothalamus. Sci. 1966;153:1284–1285. doi: 10.1126/science.153.3741.1284. [DOI] [PubMed] [Google Scholar]
- Caldarone B, Stock H, Abrahamsen G, Boechler M, Svare B, Rosellini R. Nonassociative processes and place preferences conditioned by testosterone. The Psych Record. 1996;46:373–390. [Google Scholar]
- Campbell HJ. The effect of steroid hormones on self-stimulation, central and peripheral. Steridologia. 1970;1:8–24. [PubMed] [Google Scholar]
- Clancy GP, Yates WR. Anabolic steroid use among substance abusers in treatment. Clin Psych. 1992;53:97–100. [PubMed] [Google Scholar]
- Clark AS, Lindenfield RC, Gibbons CH. Anabolic-androgenic steroids and brain reward. Pharmacol Biochem Behav. 1996;53:741–745. doi: 10.1016/0091-3057(95)02082-9. [DOI] [PubMed] [Google Scholar]
- Connolly PB, Roselli CE, Resko JA. Aromatase activity in adult guinea pig brain is androgen dependent. Biol Reprod. 1990;43:698–703. doi: 10.1095/biolreprod43.4.698. [DOI] [PubMed] [Google Scholar]
- Copeland J, Peters R, Dillon P. Anabolic-androgenic steroids dependence in a woman. Aust New Zeal J Psych. 1998;32:589. [PubMed] [Google Scholar]
- Corcoran JP, Longo E. Psychological treatment of anabolic androgenic steroids dependent individuals. J Subst Abuse Treat. 1992:228–235. doi: 10.1016/0740-5472(92)90065-v. [DOI] [PubMed] [Google Scholar]
- Corrigan B. Anabolic steroids and the mind. Med J Aust. 1996;165:222–223. doi: 10.5694/j.1326-5377.1996.tb124932.x. [DOI] [PubMed] [Google Scholar]
- Cunningham GR, Tindal DJ, Means AR. Differences in steroid specificity for rat androgen binding protein and the cytoplasmic receptor. Steroids. 1979;33:261–276. doi: 10.1016/0039-128x(79)90003-5. [DOI] [PubMed] [Google Scholar]
- DeBeun R, Jansen E, Slangen JL, Van de Poll NE. Testosterone as appetitive and discriminative stimulus in rats: sex- and dose-dependent effects. Physiol Behav. 1992;52:629–634. doi: 10.1016/0031-9384(92)90389-j. [DOI] [PubMed] [Google Scholar]
- Faigenbaum AD, Zaichkowsky LD, Gardner DE, Micheli LJ. Anabolic steroid use by male and female middle school students. Pediatrics. 1998;101:398–407. doi: 10.1542/peds.101.5.e6. [DOI] [PubMed] [Google Scholar]
- Foltin RW. The importance of drug self-administration studies in the analysis of abuse liability: An analysis of caffeine, nicotine, anabolic steroids, and designer drugs. Amer J Addictions. 1992;1:139–149. [Google Scholar]
- Franke WW, Berendonk B. Hormonal doping and androgenization of athletes: a secret program of the German Democratic Republic government. Clin Chem. 1997;43:1262–1279. [PubMed] [Google Scholar]
- Frye CA, Bayon LE, Vongher JM. Intravenous progesterone elicits a more rapid induction of lordosis in rats than does SKF38393. Psychobiology. 2000;28:99–109. [Google Scholar]
- Frye CA, Duncan JE, Basham M, Erskine MS. Behavioral effects of 3α-Androstanediol II: Hypothalamic and preoptic area actions via a GABAergic mechanism. Behavioral Brain Res. 1996;79:119–130. doi: 10.1016/0166-4328(96)00005-8. [DOI] [PubMed] [Google Scholar]
- Frye CA, Park D, Tanaka M, Rosellini R, Svare B. The neurosteroids 3α-Androstandiol may mediate the effects of testosterone on conditioned place preference. Psychoneuroendocrinol. 2001;26:731–750. doi: 10.1016/s0306-4530(01)00027-0. [DOI] [PubMed] [Google Scholar]
- Frye CA, Rhodes ME, Rosellini R, Svare B. The nucleus accumbens as a site of action for rewarding properties of testosterone and its 5alpha-reduced metabolites. Pharmacol Biochem Behav. 2002;74:119–27. doi: 10.1016/s0091-3057(02)00968-1. [DOI] [PubMed] [Google Scholar]
- Frye CA, van Keuran KR, Erskine MS. Behavioral effects of 3α-Androstanediol: I. Modulation of sexual receptivity and promotion of GABA-stimulated chloride flux. Behavioral Brain Res. 1996;79:109–118. doi: 10.1016/0166-4328(96)00004-6. [DOI] [PubMed] [Google Scholar]
- Frye CA, Van Keuran KR, Rao PN, Erskine MS. Analgesic effects of the neurosteroid 3α-Androstanediol. Brain Res. 1996a;709:1–9. doi: 10.1016/0006-8993(95)01118-8. [DOI] [PubMed] [Google Scholar]
- Frye CA, Van Keuran KR, Rao PN, Erskine MS. Progesterone and 3α-Androstanediol conjugated to bovine serum albumin affects estrous behavior when applied to the MBH and POA. Behav Neurosci. 1996b;96:603–612. doi: 10.1037//0735-7044.110.3.603. [DOI] [PubMed] [Google Scholar]
- Gee KW. Steroid modulation of the GABA/benzodiazepine receptor-linked chloride ionophore. Mol Neurobiol. 1988;2:291–317. doi: 10.1007/BF02935636. [DOI] [PubMed] [Google Scholar]
- Goudsmit E, Feenstra MG, Swabb DF. Central monoamine metabolism in the male Brown-Norway rat in relation to aging and testosterone. Brain Res Bull. 1990;25:755–763. doi: 10.1016/0361-9230(90)90054-4. [DOI] [PubMed] [Google Scholar]
- Graham-Lorence S, Amameh B, White R, Peterson J, Simpson E. A three dimensional model of aromatase cytochrome. Protein Sci. 1995;4:1065–1080. doi: 10.1002/pro.5560040605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greenblatt RB, Karpas A. Hormone therapy for sexual dysfunction. The only “true aphrodisiac. Postgrad Med. 1983;74:78–80. 84–9. doi: 10.1080/00325481.1983.11698379. [DOI] [PubMed] [Google Scholar]
- Haupt HA, Rovere GD. Anabolic steroids: A review of the literature. American J Sports Med. 1984;12:469–484. doi: 10.1177/036354658401200613. [DOI] [PubMed] [Google Scholar]
- Hauser P, Devinsky O, DeBellis M, Theodore WH, Post RM. Benzodiazepine withdrawal delirium with catatonic features. Occurences in patients with partial seizure disorders. Arch Neurol. 1989;46:696–699. doi: 10.1001/archneur.1989.00520420118033. [DOI] [PubMed] [Google Scholar]
- Hays LR, Littleton S, Stillner V. Anabolic steroid dependence. American J Psych. 1990;147:122. doi: 10.1176/ajp.147.1.122a. [DOI] [PubMed] [Google Scholar]
- Herberg IJ. Seminal ejaculation following positively reinforcing electrical stimulation of the rat hypothalamus. J Comparative Physio Psych. 1963;56:679–685. doi: 10.1037/h0041146. [DOI] [PubMed] [Google Scholar]
- Itil TM. Neurophysiological effects of hormones in humans: Computer EEG profiles of sex and hypothalamic hormones. In: Sachar EJ, editor. Hormones, behavior, and psychopathology. Raven Press; NY: 1976. pp. 31–40. [Google Scholar]
- Itil TM, Cora R, Akpinar S, Herrmann WM, Patterson CJ. Psychotropic” action of sex hormones: Computerized EEG in establishing the immediate CNS effects of steroid hormones. Current Therapeutic Res. 1974;16:1147–1170. [PubMed] [Google Scholar]
- Jalilian-Tehrani MH, Karakiulakis G, LeBlond CB, Powell R, Thomas PJ. Androgen-induced sexual dimorphism in high affinity dopamine binding in the brain transcends the hypothalamic-limbic region. British J Pharmaco. 1982;75:37–48. doi: 10.1111/j.1476-5381.1982.tb08755.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janne O. Androgen interaction through multiple steroid receptors. National Institute on Drug Abuse; Rockville: 1990. [PubMed] [Google Scholar]
- Janne OA, Palvimo JJ, Kallio P, Mehto M. Androgen receptor and mechanism of androgen action. Annals Med. 1993;25:83–89. doi: 10.3109/07853899309147863. [DOI] [PubMed] [Google Scholar]
- Johnson LR, Wood RI. Oral testosterone self-administration in male hamsters. Neuroendocrinology. 2001 Apr;73:285–92. doi: 10.1159/000054645. [DOI] [PubMed] [Google Scholar]
- Kann L, Warren C, Harris W, Collins JL, Williams BI, Ross JG, Kolbe LJ. Youth risk behavior surveillance: United States. J School Health. 1996;10:365–377. doi: 10.1111/j.1746-1561.1996.tb03394.x. [DOI] [PubMed] [Google Scholar]
- Kashkin K, Kleber H. Hooked on hormones? An anabolic steroid addiction hypothesis. J A M A. 1989;262(22):3166–3170. doi: 10.1001/jama.262.22.3166. [DOI] [PubMed] [Google Scholar]
- King BE, Packard MG, Alexander GM. Affective properties of intra-medial preoptic area injections of testosterone in male rats. Neurosci Lett. 1999;269:149–152. doi: 10.1016/s0304-3940(99)00440-1. [DOI] [PubMed] [Google Scholar]
- Kinsley C, Svare B. Prenatal stress alters maternal aggression in mice. Physiol Behav. 1988;42:7–13. doi: 10.1016/0031-9384(88)90252-1. [DOI] [PubMed] [Google Scholar]
- Koob F. Neural mechanisms of drug reinforcement. In: The neurobiology of drug and alcohol addiction. In: Kalixas PW, Samson HH, editors. Academy Sci, NY NY. Vol. 654. 1992. p. 91. [Google Scholar]
- Koob GF, LeMoal M. Drug abuse: hedonic homeostatic dysregulation. Sci. 1997;278:52–58. doi: 10.1126/science.278.5335.52. [DOI] [PubMed] [Google Scholar]
- Kornetsky C. Action of opioid drugs on the brain-reward system. NIDA Res Monogr. 1995;147:33–52. [PubMed] [Google Scholar]
- Korzekva K, Trager W, Mancewics J, Osawa Y. Studies in the mechanism of aromatase and other cytochrome P450 mediated deformylation reaction. J Steroid Biochem Molec Bio. 1993;44:367–373. doi: 10.1016/0960-0760(93)90240-w. [DOI] [PubMed] [Google Scholar]
- Korzekva K, Trager W, Smith S, Osawa Y, Gilette J. Theorectical studies on the mechanism of conversion of androgens to estrogens by aromatose. Biochem. 1991;30:6155–6162. doi: 10.1021/bi00239a011. [DOI] [PubMed] [Google Scholar]
- Kouri EM, Pope HG, Jr, Katz DL. Use of anabolic-androgenic steroids: We are talking prevalence rates. J A M A. 1994;271:347. [PubMed] [Google Scholar]
- Kritzer MF. Selective colocalization of immunoreactivity for intracellular gonadal hormone receptors and tyrosine hydroxylase in the ventral tegmental area, substantia nigra, and retrorubral fields in the rat. J Comp Neurol. 1997;379:247–60. doi: 10.1002/(sici)1096-9861(19970310)379:2<247::aid-cne6>3.0.co;2-3. [DOI] [PubMed] [Google Scholar]
- Lieberburg I, Maclusky NJ, McEwen BS. 5alpha-Dihydrotestosterone (DHT) receptors in rat brain and pituitary cell nuclei. Endocrinology. 1977;100:598–607. doi: 10.1210/endo-100-2-598. [DOI] [PubMed] [Google Scholar]
- Lookingbill D, Demers L, Wang C, Leung A, Rittmaster R, Santen R. Clinical and biochemical parameters of androgen action in normal healthy Caucasian versus Chinese subjects. J Clin Endocrinol Metab. 1991;72:1242–1248. doi: 10.1210/jcem-72-6-1242. [DOI] [PubMed] [Google Scholar]
- Lorrian DS, Riolo V, Matuszewich L, Hull EM. Lateral hypothalamic serotonin inhibits nucleus accumbens dopamine: implications for sexual satiety. J Neurosci. 1999;19:7648–7652. doi: 10.1523/JNEUROSCI.19-17-07648.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Majewska MD, Harrison NL, Schwartz RD, Barker JL, Paul SM. Steroid hormone metabolites are barbiturate-like modulators of the GABA receptor. Science. 1986;232:1004–1007. doi: 10.1126/science.2422758. [DOI] [PubMed] [Google Scholar]
- Malone DA, Jr, Dimeff RJ, Lombardo JA, Sample RH. Psychiatric effects and psychoactive substance use in anabolic-androgenic steroid users. Clin J Sport Med. 1995;5:25–31. doi: 10.1097/00042752-199501000-00005. [DOI] [PubMed] [Google Scholar]
- Mani SK, Allen JM, Lydon JP, Mulac-Jericevic B, Blaustein JD, DeMayo FJ, Conneely O, O’Malley BW. Dopamine requires the unoccupuied progesterone receptor to induce sexual behavior in mice. Mol Endocrinol. 1996;10:1728–1737. doi: 10.1210/mend.10.12.8961281. [DOI] [PubMed] [Google Scholar]
- Marshall E. The drug of champions. Science. 1988;242:183–184. doi: 10.1126/science.3175643. [DOI] [PubMed] [Google Scholar]
- Masonis AET, McCarthy MP. Direct effects of the anabolic/androgenic steroids, stanozolol and 17-methyltestosterone, on benzodiazepine binding to the γ-aminobutyric acidA receptor. Neurosci Lett. 1995;189:35–38. doi: 10.1016/0304-3940(95)11445-3. [DOI] [PubMed] [Google Scholar]
- Masonis AET, McCarthy MP. Effects of the adrogenic/anabolic steroid stanozolol on GABAA receptor function: GABA-stimulated 36Cl− influx and [35S] TBPS binding. J. Pharmacol. Exp Ther. 1996;279:186–193. [PubMed] [Google Scholar]
- McBride WJ, Murphy JM, Ikemoto S. Localization of brain reinforcing mechanisms: Intracranial self-administration and intra-cranial place-conditioning studies. Behav Brain Res. 1999;101:129–152. doi: 10.1016/s0166-4328(99)00022-4. [DOI] [PubMed] [Google Scholar]
- McBride AJ, Williamson K, Petersen T. Three cases of nalbuphine hydrochloride dependence associated with anabolic steroid use. Br J Sports Med. 1996;30:69–70. doi: 10.1136/bjsm.30.1.69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCaslin PP, Morgan WM. Anticonvulsant activity of several excitatory amino acids antagonists against barbital withdrawal-induced spontaneous convulsions. Eur J Pharm. 1988;147:381–386. doi: 10.1016/0014-2999(88)90172-0. [DOI] [PubMed] [Google Scholar]
- McEwen B. Binding and metabolism of sex steroids by the hypothalamic-pituitary unit: physiological implications. Reviews Physio. 1980;42:97–110. doi: 10.1146/annurev.ph.42.030180.000525. [DOI] [PubMed] [Google Scholar]
- Mellon SH. Neurosteroids: biochemistry, modes of action, and clinical relevance. J Clin Endocrinol Metab. 1994;78:1003–1008. doi: 10.1210/jcem.78.5.8175951. [DOI] [PubMed] [Google Scholar]
- Mitchell JB, Stewart J. Effects of castration, steroid replacement, and sexual experience on mesolimbic dopamine and sexual behaviors in the male rat. Brain Res. 1989;491:116–127. doi: 10.1016/0006-8993(89)90093-0. [DOI] [PubMed] [Google Scholar]
- Moore WV. Anabolic steroid use in adolescence. Jama. 1988;260:3484–3486. [PubMed] [Google Scholar]
- Moses J, Loucks JA, Watson HL, Matuszewick L, Hull EM. Dopaminergic drugs in the medial preoptic area and nucleus accumbens: effects on motor activity, sexual motication, and sexual performance. Phamacol Biochem Behav. 1995;51:681–686. doi: 10.1016/0091-3057(94)00437-n. [DOI] [PubMed] [Google Scholar]
- Olds J. Effects of hunger and male sex hormone of self-stimulation of the brain. J Comparative Physio Psych. 1958;51:320–324. doi: 10.1037/h0040783. [DOI] [PubMed] [Google Scholar]
- Orchard JW, Best JP. Test violent offenders for anabolic steroid use. Medical Journal of Australia. 1994;161:232. doi: 10.5694/j.1326-5377.1994.tb127408.x. [DOI] [PubMed] [Google Scholar]
- Packard M, Cornell A, Alexander G. Rewarding affective properties of intra-nucleus accumbens injections of testosterone. Behav Neurosci. 1997;111:219–224. doi: 10.1037//0735-7044.111.1.219. [DOI] [PubMed] [Google Scholar]
- Packard M, Schroeder J, Alexander G. Expression of testosterone conditioned place preference is blocked by peripheral or intra-accumbens injection of □-flupenthixol. Hormones and Behav. 1998;34:39–47. doi: 10.1006/hbeh.1998.1461. [DOI] [PubMed] [Google Scholar]
- Pope HG, Katz DL. Bodybuilder’s psychosis. Lancet. 1987;8537:863. doi: 10.1016/s0140-6736(87)91642-4. [DOI] [PubMed] [Google Scholar]
- Pope HG, Katz DL. Affective and psychotic symtoms associated with anabolic steroid use. Am J Psychiat. 1988;145:487–490. doi: 10.1176/ajp.145.4.487. [DOI] [PubMed] [Google Scholar]
- Pope HG, Katz DL. Psychiatric and medical effects of anabolic-androgenic steroid use: A controlled study of 160 athletes. Arch Gen Psychiat. 1994;51:375–382. doi: 10.1001/archpsyc.1994.03950050035004. [DOI] [PubMed] [Google Scholar]
- Pope HG, Katz D, Aizley H. Psychiatric and medical effects of anabolic steroids: A controlled study of 160 athletes. Archives Gen Psych. 1994;51:375–382. doi: 10.1001/archpsyc.1994.03950050035004. [DOI] [PubMed] [Google Scholar]
- Pope HG, Phillips KA, Olivardia R. The neural basis of drug craving Brain Research Reviews. Vol. 18. Free Press: New York. Robinson, Berridge; 1993. The Adonis Complex 2000; pp. 247–291. [DOI] [PubMed] [Google Scholar]
- Robinson TE, Berridge KC. The neural basis of drug craving: an incentive-sensitization theory of addiction. Brain Res Brain Res Rev. 1993;18:247–91. doi: 10.1016/0165-0173(93)90013-p. [DOI] [PubMed] [Google Scholar]
- Rogozkin . Metabolism of anabolic androgen steroid. Press; Boca Raton: 1991. [Google Scholar]
- Rosellini RA, Rhodes ME, Svare B, Frye CA. Testosterone’s hedonic effects may involve metabolism to 3□-diol and actions at GABAA receptors in the nucleus accumbens. Trabajos del Instituto Cajal. 2003;79:141–142. [Google Scholar]
- Rosellini RA, Svare B, Rhodes ME, Frye CA. The testosterone metabolite and neurosteroid 3alpha-androstanediol may mediate the effects of testosterone on conditioned place preference. Brain Res Brain Res Rev. 2001 Nov;37(1–3):162–71. doi: 10.1016/s0165-0173(01)00116-3. [DOI] [PubMed] [Google Scholar]
- Rousseau GG, Baxter JD. Glucocorticoids and the metabolic code. Monogr Endocrinol. 1979;12:613–29. doi: 10.1007/978-3-642-81265-1_34. [DOI] [PubMed] [Google Scholar]
- Rubinow D; Schmidt P. Androgens, brain, and behavior. Am J Psychiatry. 1996;153:974–984. doi: 10.1176/ajp.153.8.974. [DOI] [PubMed] [Google Scholar]
- Schroeder JP, Packard MG. Role of dopamine receptor subtypes in the acquisition of a testosterone conditioned place preference in rats. Neurosci Lett. 2000;282:17–20. doi: 10.1016/s0304-3940(00)00839-9. [DOI] [PubMed] [Google Scholar]
- Scoles M, Siegel S. A potential role of saline trials in morphine-induced place-preference conditioning. Pharmaco Biochem Behav. 1986;25:1169–1173. doi: 10.1016/0091-3057(86)90106-1. [DOI] [PubMed] [Google Scholar]
- Silvester L. Anabolic steroids at the 1972 Olympics. Scholastic Coach. 1973;43:90–92. [Google Scholar]
- Stenn PG, Klaiber EL, Vogel W, Broverman DM. Testosterone effects upon photic stimulation of the electroencephalogram (EGG) and mental performance of humans. Percept Mot Skills. 1972;34:371–378. doi: 10.2466/pms.1972.34.2.371. [DOI] [PubMed] [Google Scholar]
- Stumpf WE, Sar M. Steroid hormone target sites in the brain: the differential distribution of estrogin, progestin, androgen and glucocorticosteroid. J Steroid Biochem. 1976 Nov-Dec;7(11–12):1163–70. doi: 10.1016/0022-4731(76)90050-9. [DOI] [PubMed] [Google Scholar]
- Svare B, Mann M, Broida J, Kinsley C, Ghiraldi L, Miele J, Konen C. Intermale aggression and infanticide in aged C57BL/6J male mice: behavioral deficits are not related to serum testosterone (T) levels and are not recovered by supplemental T. Neurobio Aging. 1983;4:305–312. doi: 10.1016/0197-4580(83)90007-6. [DOI] [PubMed] [Google Scholar]
- Taylor WN. Synthetic anabolic-androgenic steroids: A plea for controlled substance status. The Physician Sports med. 1987;15:140–150. doi: 10.1080/00913847.1987.11709356. [DOI] [PubMed] [Google Scholar]
- Taylor GT, Weiss J, Pitha J. Testosterone in a cyclodextrin-containing formulation: behavioral and physiological effects of episode-like pulses in rats. Pharm Res. 1989;6:641–646. doi: 10.1023/a:1015922019038. [DOI] [PubMed] [Google Scholar]
- Teledgy G. Frontiers in hormone research. Vol. 15. Karger: Basel; 1987. [Google Scholar]
- Tennant F, Black DL, Vou R. Anabolic steroid dependence with opioid-type features. N Engl J Med. 1988;319:578. doi: 10.1056/NEJM198809013190910. [DOI] [PubMed] [Google Scholar]
- Tricker R, O’Neill MR, Cook D. The incidence of anabolic steroid use among competitive body builders. J Drug Educ. 1989;19:313–325. doi: 10.2190/EGT5-4YWD-QX15-FLKK. [DOI] [PubMed] [Google Scholar]
- Uzych L. Anabolic androgenic steroids and psychiatric-related effects: a review. Canadian J Psych. 1992;37:23–28. doi: 10.1177/070674379203700106. [DOI] [PubMed] [Google Scholar]
- Van de Poll NE, Van Zanten S, Jonge FH. Effects of testosterone, estrogen, and dihydrotestosterone upon aggressive and sexual behavior of female rats. Horm Behav. 1986;20:418–431. doi: 10.1016/0018-506x(86)90004-8. [DOI] [PubMed] [Google Scholar]
- Verhoeven G, Heyns W, DeMoor P. Ammonium sulfate precipitation as a tool for the study of androgens receptors proteins in rat prostrate and mouse kidney. Steroids. 1975;6:149–167. doi: 10.1016/s0039-128x(75)80016-x. [DOI] [PubMed] [Google Scholar]
- Vermes I, Varszegi M, Toth E, Telegdy G. Action of androgenic steroids on brain neurotransmitters in rats. Neuroendocrinology. 1979;28:386–393. doi: 10.1159/000122887. [DOI] [PubMed] [Google Scholar]
- Wade G. Anabolic steroids: doctors denounce them, but athletes aren’t listening. Science. 1972;176:1399–1403. doi: 10.1126/science.176.4042.1399. [DOI] [PubMed] [Google Scholar]
- Wilson J, Griffin JE. The use and misuse of androgens. Metabolism. 1980;29:1278–1295. doi: 10.1016/0026-0495(80)90159-6. [DOI] [PubMed] [Google Scholar]
- Wilson JD. Androgen abuse by athletes. Endocrine Reviews. 1988;8:181–199. doi: 10.1210/edrv-9-2-181. [DOI] [PubMed] [Google Scholar]
- Wise RA, Bozarth MA. A psychomotor stimulant theory of addiction. Psychol Rev. 1987;94:469–492. [PubMed] [Google Scholar]
- Wood RI. Oral testosterone self-administration in male hamsters: dose-response, voluntary exercise, and individual differences. Horm Behav. 2002 May;41(3):247–58. doi: 10.1006/hbeh.2002.1769. [DOI] [PubMed] [Google Scholar]
- Wright JE. Anabolic steroids and athletics. Exerc Sport Sci Rev. 1980;8:149–202. doi: 10.1249/00003677-198000080-00007. [DOI] [PubMed] [Google Scholar]
- Yesalis C, Bahrke M. Anabolic-androgenic steroids. Current issues Sports Med. 1995;19:326–340. doi: 10.2165/00007256-199519050-00003. [DOI] [PubMed] [Google Scholar]
- Yesalis C, Cowart VS. The steroid game. Champaign: Human Kinetics; 1998. [Google Scholar]
- Yesalis C, Herrick RT, Buckley WE. Self-reported use of anabolic-androgenic steroids by elite power lifters. Physician Sport Med. 1988;16:91–100. doi: 10.1080/00913847.1988.11709666. [DOI] [PubMed] [Google Scholar]
- Yesalis CE, Vicary JR, Buckley WE, Streit AL, Katz DL, Wright JE. Indications of psychological dependence among anabolic-androgenic steroid abusers. NIDA Research Monograph. 1990;102:196–214. [PubMed] [Google Scholar]
- Zhang JH, Sato M, Tohyama M. Region-specific expression of the mRNAs encoding beta subunits (beta 1, beta 2, and beta 3) of GABAA receptor in the rat brain. J Comp Neurol. 1991 Jan 22;303(4):637–57. doi: 10.1002/cne.903030409. [DOI] [PubMed] [Google Scholar]