

Abstract
Circadian rhythms are endogenous cellular programs that time metabolic and behavioral events to occur at optimal times in the daily cycle. Light and dark cycles synchronize the endogenous clock with the external environment through a process called entrainment. Previously, we identified the bacteriophytochrome-like circadian input kinase CikA as a key factor for entraining the clock in the cyanobacterium Synechococcus elongatus PCC 7942. Here, we present evidence that CikA senses not light but rather the redox state of the plastoquinone pool, which, in photosynthetic organisms, varies as a function of the light environment. Furthermore, CikA associates with the Kai proteins of the circadian oscillator, and it influences the phosphorylation state of KaiC during resetting of circadian phase by a dark pulse. The abundance of CikA varies inversely with light intensity, and its stability decreases in the presence of the quinone analog 2,5-dibromo-3-methyl-6-isopropyl-p-benzoquinone (DBMIB). The pseudo-receiver domain of CikA is crucial for sensitivity to DBMIB, and it binds the quinone directly, a demonstration of a previously unrecognized ligand-binding role for the receiver fold. Our results suggest that resetting the clock in S. elongatus is metabolism-dependent and that it is accomplished through the interaction of the circadian oscillator with CikA.
Keywords: biological rhythms, photosynthetic electron transport, pseudo-receiver, redox, Synechococcus elongatus
Circadian rhythms are oscillations of biological activities that show a periodicity of ≈24 h (1). This program of cellular metabolism allows organisms to anticipate predictable daily environmental changes, and it provides an adaptive advantage (2, 3). The circadian mechanism has been studied extensively in eukaryotic models such as fruit flies, fungi, and mice (4). The cyanobacterium Synechococcus elongatus PCC 7942 is the only prokaryotic organism whose circadian clock has been elucidated (5, 6).
All circadian systems share three major divisions (4). A central oscillator generates the fundamental rhythm of ≈24 h. In S. elongatus, the oscillator consists of the KaiA, KaiB, and KaiC proteins (6, 7). An input pathway conveys environmental signals, such as light and temperature, to the oscillator, and it modifies the oscillation to synchronize it precisely with the external daily cycle. The proteins CikA and LdpA are key components of this pathway in S. elongatus (8, 9). The third division is an output pathway that relays temporal information from the oscillator to a variety of downstream biochemical processes in diverse organisms (4). In S. elongatus, the kinase SasA acts close to the oscillator in an output pathway (10, 11).
In eukaryotic systems, circadian rhythmicity is generated by interlocked transcription–translation feedback loops (4). However, cyanobacteria use a posttranslational oscillator (12, 13) in which the phosphorylation state of the oscillator protein KaiC plays a critical role (7). KaiC has both autophosphorylation and autodephosphorylation activities; KaiA enhances the autophosphorylation activity, and KaiB weakens the effect of KaiA (14). The three Kai proteins and ATP can reconstitute a circadian oscillation of the phosphorylation pattern in vitro, similar to that in vivo, that is sustained for at least 72 h (7).
In contrast to the depth of understanding of oscillator performance (7), the pathway of signal transduction from the environment to the Kai proteins has not been clearly established. An environmental signal that universally affects circadian clocks is light. Usually, light is sensed by a photoreceptor, which affects the cellular level of a clock component (15, 16). In cyanobacteria, the bacteriophytochrome-like protein CikA was identified as a key constituent of the input pathway because cikA mutants lack the ability to reset the phase of the rhythm after a dark pulse (8). However, it is not clear what kind of signal CikA receives. A potential chromophore-binding GAF domain lacks the conserved residues expected for adduct formation, and in vivo chromophore-binding assays are negative (17). Previously, we demonstrated that CikA abundance varies with light intensity, and it is sensitive to an electron transport inhibitor, 2,5-dibromo-3-methyl-6-isopropyl-p-benzoquinone (DBMIB), which affects the redox state of the cellular plastoquinone (PQ) pool (18). These data suggested that CikA might sense light indirectly through the cellular redox state and then pass this information to the circadian oscillator.
The goal of this work was to establish the relationship between CikA and the oscillator, the basis of CikA sensitivity to light, and the mechanism of signal transfer from CikA to the oscillator. Here, we demonstrate that CikA directly binds a quinone ligand and interacts with the circadian oscillator. The results identify a molecular mechanism by which metabolism regulates the circadian clock.
Results
CikA Is Present in a High-Molecular-Mass Complex and Copurifies with Oscillator Proteins.
The three Kai proteins and SasA have been shown to form a periodosome, a high-molecular-mass protein complex essential for sustaining circadian rhythmicity in S. elongatus (19, 20). Previously, we demonstrated that LdpA copurifies with KaiA, SasA, and CikA proteins, suggesting that LdpA and CikA might also be a part of the periodosome (18). To test directly for evidence that CikA forms a complex with components of the circadian oscillator, we used both copurification and gel-filtration methods.
Either CikA or KaiC was affinity-tagged with 6 His residues, expressed in cyanobacterial cells at wild-type (WT) levels, and recovered under mild conditions that allow copurification of proteins with which they interact. The fraction that coeluted with His-tagged KaiC contained KaiA (whose interaction with KaiC is known) and CikA (Fig. 1A, lane 2). None of these proteins was present in eluate from extracts from the WT cells, which do not express a His-tagged protein (Fig. 1 A and B, lanes 1). The sigma factor RpoD1, which was not expected to interact with Kai proteins, did not copurify with any of these samples either when the tagged proteins were expressed at native levels or when overexpressed (see Fig. 6, which is published as supporting information on the PNAS web site). KaiA coeluted with tagged CikA (Fig. 1B, lane 2). We did not detect KaiC in the fraction that copurifies with CikA-His expressed at the WT level (21), probably because the yield of eluted CikA-His is an order of magnitude lower than that of KaiC-His expressed at its WT level based on silver-stained SDS/PAGE (data not shown). However, KaiC was found in the eluted fraction when CikA-His was overexpressed (Fig. 6, lane 3). Overexpression of His-tagged KaiC to a similar extent reproduced the associations that were detected when it was expressed at WT levels (Fig. 1A, lane 2, and Fig. 6, lane 1), suggesting that the data obtained with overexpressed CikA are reliable. KaiB and SasA, known to interact with KaiC, were not tested.
Fig. 1.

CikA forms high-molecular-mass complexes and copurifies with oscillator proteins. Antisera used for immunoblot analysis are shown to the right of each panel. (A and B) Immunoblot analysis of the eluate fraction from the strains whose KaiC-His (A, lane 2) or CikA-His (B, lane 2) was expressed at a level similar to that of the corresponding native protein. Tagged proteins were purified under mild conditions, allowing for copurification of interacting proteins. WT strain (lanes 1) did not contain any tagged proteins, and it was used as a negative control. Lanes tested with anti-RpoD1, which were all blank, are not included. (C) Immunoblot analysis of gel-filtration fractions. Cyanobacterial cells were collected at ZT5 and ZT17. Soluble protein extracts were fractionated according to the size of proteins and protein complexes. The molecular masses of the protein standards are indicated.
To test whether the composition of native complexes in vivo is consistent with the physical interaction of Kai proteins and CikA detected in the copurification assay, cyanobacterial proteins and protein complexes were separated by gel filtration. Two samples were analyzed from cells grown in a 12-h light/12-h dark cycle (LD), where time points are designated by zeitgeber time (ZT5 and ZT17), indicating 5 or 17 h after light onset. In both samples, CikA is present in fractions that correspond to a molecular mass of just under 440 kDa (Fig. 1C); this value is larger than the active CikA dimer of 166 kDa (21), which indicates that CikA participates in a protein complex.
KaiC fractionated in two populations as reported previously (19). The first is that in which CikA was found, ≈440 kDa; KaiC forms a hexamer (≈360 kDa) as a part of the periodosome (19). At ZT17, this population of KaiC molecules shifted to fractions that correspond to just under 669 kDa. The second population corresponds to an estimated molecular mass <100 kDa, which likely includes monomeric KaiC (≈61 kDa).
At ZT5, KaiA, whose dimer is 65 kDa, mostly eluted in the fractions that include monomeric KaiC; however, a small amount of KaiA was present across most fractions analyzed. At ZT17, most KaiA eluted in high-molecular-mass fractions that also contain CikA and some KaiC. Taken as a whole, the data from gel-filtration analysis and copurification experiments allow us to suggest that CikA forms a high-molecular-mass complex and, along with Kai proteins, is a part of the periodosome.
CikA Affects the Phosphorylation State of KaiC.
A striking phenotypic feature of the cikA mutant is its inability to reset the phase of the circadian rhythm after a 5-h dark pulse, whereas WT cells respond in the next cycle with a peak that is offset by up to 12 h relative to the initial daily peak time (8). The greatest difference in circadian resetting between the WT and cikA strains is detected when the dark pulse is given at ZT8 (21). To establish the role CikA plays during the dark pulse, cikA-null and WT cells were collected before, during, and after a pulse administered at ZT8, and protein extracts were subjected to immunoblot analysis. The amount of CikA increased in the dark, and it decreased once cells were returned to light (Fig. 2A), but it did not change in cells not subjected to the dark pulse (ZT8–ZT14; data not shown). KaiA was constant both during the dark pulse and in constant light conditions (LL, data not shown). KaiC is known to autophosphorylate, and the phosphorylation pattern varies during the circadian cycle (22). This pattern differed strikingly between WT and cikA strains. In WT, the KaiC pool was mostly phosphorylated at ZT8, and it became progressively dephosphorylated during the dark pulse and after the return to light (Fig. 2B). When WT cells were kept in LL, the proportion of unphosphorylated KaiC increased more slowly. In the cikA strain at ZT8, KaiC was equally divided between phosphorylated and unphosphorylated forms. The proportion of phosphorylated KaiC slowly increased whether or not cells were subjected to a dark pulse. These data indicate that CikA affects the phosphorylation state of KaiC and its dynamics in response to an environmental stimulus.
Fig. 2.

CikA affects phosphorylation state of KaiC. Immunoblot analysis of total protein fractions from cyanobacterial cells collected before, during, and after a dark pulse or in LL is shown. Cells were put in the dark at ZT8, and the samples were collected at 1 h, 3 h, and 5 h from the start of the dark pulse and at 1 h, 2 h, and 4 h after the dark pulse ended (+1, +2, +4). At the same time, samples were collected from the cultures kept in LL. (A and B) Immunoblot analysis using antiserum raised against CikA (A) or KaiC (B). Upper and lower bands correspond to phosphorylated and unphosphorylated forms of KaiC, respectively (47), and they were verified by lambda phosphatase treatment (data not shown). ▿cikA is a mutant defective for CikA.
CikA Is Sensitive to Light and an Inhibitor That Affects Redox State.
Immunoblot analysis was performed to see whether the abundance of CikA is regulated during the LD cycle. In WT cells, the CikA level decreased in the light and increased in the dark, and in LL it decreased in the subjective day and increased in subjective night, indicating that abundance of CikA is regulated by the circadian clock (Fig. 3A). To separate circadian from environmental control, CikA abundance was examined in a kaiC mutant that lacks a functional clock. In the absence of the clock, CikA levels were still regulated in an LD cycle but not in LL. Thus, the abundance of CikA is modulated by light and by the circadian clock, decreasing in light (day) and increasing in the dark (night).
Fig. 3.

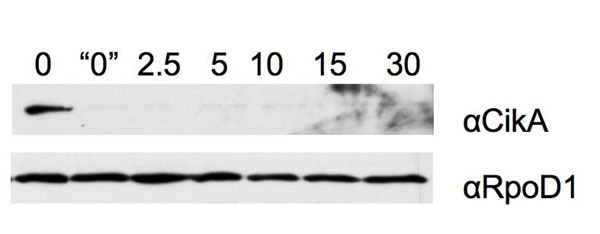
CikA is sensitive to environmental or chemical signals that affect the redox state of the cell. Immunoblot analysis of total protein samples is shown. (A) Samples collected from cells in the LD cycle and LL and probed with CikA antiserum. Lanes are marked with time in hours. ▿kaiC is a mutant defective for KaiC. Because of differences in film-exposure times, the results are not quantitative among panels. (B) Cyanobacterial cells were treated for 15 min with inhibitors of electron transport or the proton gradient as indicated above each lane. Immunoblots were probed with antiserum directed against CikA or RpoD1. DCMU, 3-(3,4-dichlorophenyl)-1,1-dimethylurea; CCCP, carbonylcyanide m-chlorophenylhydrazone. (C) Cyanobacterial cells were treated for 15 min with DBMIB in the presence or absence of chloramphenicol, and protein extracts were subjected to immunoblot analysis.
In photosynthetic organisms, some processes that appear to be regulated by light are actually regulated by the redox state of the cell, particularly by that of the PQ pool, which varies in relation to the intensity of light (23). To establish whether the amount of CikA is regulated by redox state, cyanobacterial cells were treated briefly with commonly used photosynthetic electron transport (PET) inhibitors and proton ionophores (24). Previously, we demonstrated that the quinone analog DBMIB affects the levels of CikA, LdpA and, to a lesser extent, KaiA, but it does not affect other proteins, such as PsaC (an iron–sulfur cluster protein involved in PET within photosystem I) and D1 (a key component of photosystem II) (18). Fig. 3B demonstrates that after 15 min of treatment with the DBMIB, CikA was undetectable, whereas treatment with other classes of inhibitors (DCMU, CCCP, methylamine, and hydroxylamine) had no effect. The level of RpoD1, which is not specifically related to the clock, was not affected by any of the treatments. Additional experiments showed that CikA disappears within the 3-min sample preparation time when cells are harvested immediately after 10 μM DBMIB treatment (Fig. 7, which is published as supporting information on the PNAS web site).
DBMIB and other PET inhibitors block translation in S. elongatus (25). To determine whether DBMIB reduces CikA levels by blocking synthesis or by accelerating its degradation, cells were treated with the translation inhibitor chloramphenicol, which did not cause a reduction in CikA levels or alter the effect of DBMIB on CikA (Fig. 3C). The data indicate that the effect of DBMIB is posttranslational, that DBMIB probably actively affects the stability of CikA, and that translation of new proteins is not required for CikA degradation in the presence of the quinone analog.
The PsR Domain of CikA Is Crucial for DBMIB Sensitivity of the Protein.
To identify the domain of CikA responsible for CikA sensitivity to DBMIB, we examined a collection of strains that carry defective variants of CikA encoded by point or deletion mutations in cikA (21). When various concentrations of DBMIB were added to the panel of mutants, those CikA variants that lack the GAF domain (ΔGAF) or carry a substitution in the phosphorylation site of the kinase domain (H393A) were still sensitive to DBMIB, although to a lesser degree than the WT protein (Fig. 4). However, the CikA variant that lacks the pseudo-receiver domain (ΔPsR) was completely insensitive to DBMIB, indicating that this domain is crucial for DBMIB sensitivity. Moreover, the PsR domain, when expressed alone, was sensitive to the quinone analog (Fig. 4); thus, the PsR domain is necessary and sufficient for DBMIB sensitivity of CikA.
Fig. 4.

The pseudo-receiver (PsR) domain is necessary and sufficient for quinone sensitivity of CikA. Cyanobacterial cells that express different variants of CikA were treated for 15 min with DBMIB at the concentration shown above each lane. Total protein samples were analyzed by using immunoblotting and anti-CikA antiserum. Strains express WT CikA (WT), a variant missing the GAF domain (ΔGAF), a kinase-defective missense mutant (H393A), a variant missing the PsR domain (ΔPsR), or the PsR domain alone (PsR).
Membrane-Associated Factors Are Not Necessary for CikA Sensitivity to DBMIB.
Native quinones, including the PQ of cyanobacteria and other photosynthetic organisms, are lipid-soluble molecules, so all of their known targets are proteins associated with membranes (26). CikA is a soluble protein; however, in vivo CikA is localized to the poles of the cell (21). To see whether specific membrane-associated or soluble factors are necessary for the effect of DBMIB on CikA, we separated cyanobacterial proteins into membrane and soluble fractions; CikA was present in both (Fig. 8B, which is published as supporting information on the PNAS web site), as might be expected if it is peripherally associated with the membrane at the cell pole and some CikA molecules detach during the fractionation. CikA in both fractions was still sensitive to DBMIB, indicating that specific soluble or membrane-associated factors either are not necessary for the DBMIB effect or are present in both fractions (Fig. 8B). Furthermore, when CikA-His was purified under mild conditions that allow copurification of interactants (Fig. 6), CikA in the eluate fraction was also sensitive to DBMIB (Fig. 8C), suggesting that the specific factor responsible for the DBMIB effect on CikA is closely associated with CikA or that DBMIB affects CikA directly.
The PsR Domain of CikA Binds Quinones Directly.
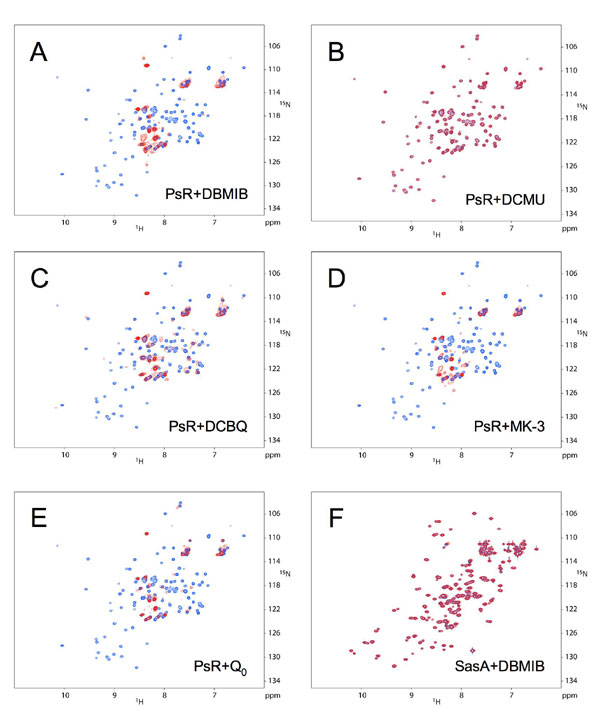
The sensitivity to DBMIB of partially purified CikA (Fig. 8C) and the fast disappearance CikA in the presence of DBMIB (Fig. 7) suggested that the quinone might affect CikA directly rather than through an intermediate step. Analysis of mutants (Fig. 4) pointed to a special role for the PsR domain in this process. To see whether DBMIB directly binds the PsR domain, purified recombinant PsR was subjected to two-dimensional NMR analysis in the presence or absence of equal molar DBMIB (Fig. 5). In the presence of DBMIB, a subset of NMR peaks changed significantly, and other peaks changed little or not at all, which suggests that the interaction with the PsR domain is specific at this 1:1 ratio. Addition of the PET inhibitor DCMU, which had no effect on CikA stability in vivo, did not affect the NMR spectrum (Fig. 9B, which is published as supporting information on the PNAS web site), nor did ethanol, which was used as solvent for DBMIB and DCMU stocks, at the relevant concentrations (data not shown). The NMR spectrum of another protein involved in circadian regulation (SasA) did not change in the presence of DBMIB, supporting specificity in the DBMIB–PsR interaction (Fig. 9F). To see whether other quinones affect the PsR domain, several were tested, such as 2,5-dichloro-1,4-benzoquinone (DCBQ, an artificial electron acceptor for photosystem II), menadione sodium bisulfate (MK-3), and coenzyme Q0; MK-3 and coenzyme Q0 are soluble analogs of the natural Escherichia coli quinones MK-8 and Q-8, respectively (27). PQ itself is not water-soluble or commercially available. The effect on the PsR NMR spectrum of all tested quinones is identical to that of DBMIB (Fig. 9 C–E). Of these quinones, only DCBQ, a PQ analog, affected CikA in vivo in a manner similar to that of DBMIB (data not shown). We do not know whether MK-3 and coenzyme Q0 enter the cyanobacterial cells.
Fig. 5.

NMR observation of PsR–DBMIB interaction. Superposition of two-dimensional NMR 1H,15N correlation spectra of the PsR domain alone (blue) and with added DBMIB at a 1:1 ratio (red) is shown. Boxed and labeled correlations are those of PsR alone, which apparently change significantly when DBMIB is added. Horizontal lines connect the amide protons of asparagine and glutamine side chains. Additional spectra are shown in Fig. 9. They show that PsR interacts with other quinones but not with a nonquinone PET inhibitor, DCMU.
Discussion
In the clocks of multicellular eukaryotes, the transcription–translation feedback loop requires cellular and sometimes tissue-level compartmentalization of input, oscillator, and output pathways. Here, we present additional evidence that, in contrast to eukaryotic systems, cyanobacteria colocalize most of the known clock components in a large protein complex, the periodosome. Previously, it was shown that the three Kai proteins of the oscillator, and SasA of the output pathway, are part of this complex (19). Recently, we demonstrated that Kai proteins also copurify with LdpA, a component of the input pathway responsible for adjusting circadian period length according to light intensity (18). Here, we provide evidence that CikA, a key element of the input pathway, is also a part of the periodosome.
We demonstrate that CikA affects the ratio of phosphorylated to unphosphorylated KaiC (Fig. 2). A kaiC mutant with altered phosphorylation rhythms has a resetting phenotype similar to that of the cikA mutant (28). It is still not clear how CikA affects the phosphorylation state of KaiC. Yeast two-hybrid assays do not detect direct interaction of CikA with any of the Kai proteins (S.R. Mackey and S.S.G., unpublished data). The addition of CikA to Kai proteins in vitro does not affect the phosphorylation rate of KaiC (S.B. Williams and S.S.G., unpublished data). Thus, the interaction of CikA and Kai proteins probably includes one or more yet-unknown proteins, which serves as a bridge between them and which is absent from both yeast and in vitro assays.
It was proposed initially that CikA acts similarly to plant phytochromes by sensing light with a chromophore bound in the GAF domain. The data shown here pose an alternative paradigm in which the cyanobacterial clock receives information about light and darkness not through a photoreceptor (as is done in most circadian systems) but by CikA detecting quinones, small molecules whose presence and redox state depend not only on light but also on the metabolic state of cyanobacterial cells. Others have shown that the metabolic state of the organism affects the circadian clock in mammals, in which the phasing of circadian rhythms in the liver is reset by feeding (29). A possible molecular mechanism of feeding entrainment is that the redox state of nicotinamide adenine dinucleotide cofactors affects the DNA-binding capacity of CLOCK/BMAL1 and NPAS2/BMAL1 heterodimers, which are oscillator components in the mammalian clock (30). In plants, there is no evidence yet for metabolic control of the circadian clock; however, redox control of gene expression and some posttranscriptional processes is well established (31, 32).
Here, we report that quinones directly affect the structure of CikA (Fig. 5). It is known that ubiquinone Q0 affects kinase activities of the purified soluble histidine kinases BvgS (regulator of virulence in Bordetella pertussis) and EvgS (transcriptional regulator in E. coli) (27). Another kinase from E. coli, ArcB, which regulates gene expression during the switch between aerobic and anaerobic growth conditions, is sensitive to quinones such as ubiquinone (Q0) and menadione (MK-3) (33). These three kinases contain the PAS domain, a fold that is responsible for the quinone binding; in other proteins, PAS binds a variety of small ligands. However, the ArcB ortholog in Haemophilus influenzae lacks the PAS domain and nevertheless is able to mediate signal transduction in response to redox conditions, suggesting that domains other than PAS can be involved in quinone redox sensing (34).
In CikA, the PsR domain binds a PQ analog (Figs. 4 and 5). The founding members of the receiver family act as recipients of phosphoryl groups from histidine kinases by a reaction at a conserved active-site aspartyl residue (35). The PsRs of CikA and KaiA in the S. elongatus clock lack the aspartyl residue, and they are incapable of phosphoryl transfer based on structural analyses (8, 14). PsRs are present in other bacteria and in plants, and the biochemical functions of many are unknown. The current work suggests that diverse ligand-binding functions may be played by PsRs, as is true for PAS and GAF domains (36–38).
The PsR of CikA is responsible for localization of the protein at the cell pole (21). We show here that part of the CikA population copurifies with the membrane fraction, indicating that CikA is localized within close proximity of quinones, which are lipid-soluble molecules. Previously, we proposed that the redox status of the PQ pool affects the stability of CikA (18) because DBMIB causes reduction of the PQ pool, and it is widely used for altering the redox state of the cell (24). The current data show that DBMIB affects CikA directly, regardless of the effect that it has on the redox state of native PQ by blocking PET. Indeed, the quinone analog DCBQ, which accepts electrons directly from photosystem II and should compete with native PQ for reduction (such that the PQ pool is oxidized) (39), behaves in a manner similar to DBMIB, which binds to the cytochrome b6f complex and prevents PQH2 reoxidation (such that the PQ pool is reduced). We conclude that the exogenously added quinone itself binds to the PsR domain of CikA; if native oxidized and reduced PQ have different binding affinities for PsR, this distinction is overcome at the concentrations of DBMIB and DCBQ that are commonly used. This technical difference could explain why there is such a dramatic effect on CikA stability in the presence of DBMIB, whereas during the dark/light transition in vivo the CikA depletion is more modest. After DBMIB treatment the CikA antiserum sometimes detects a band of reduced mobility (Fig. 8C) that comigrates with a band detected by antiserum raised against ClpP (data not shown), a protease whose depletion affects circadian period (40); we propose that binding of DBMIB by CikA targets the protein for immediate degradation, accounting for its rapid disappearance.
TOC1, a light-sensitive component of the circadian oscillator in plants (41), also contains a PsR domain. It remains to be seen whether components of the clock in plants and other kingdoms are able to sense the light environment in a manner similar to that used by CikA.
Materials and Methods
Bacterial Strains, Culture Conditions, and DNA Manipulations.
All S. elongatus PCC 7942 strains were grown as described previously (17). Cyanobacterial and E. coli strains used in this study are listed in Table 1, which is published as supporting information on the PNAS web site. Null strains for cikA (AMC1426) or kaiC (AMC1511) were constructed by insertion of an inactivation cassette (42) or a Mu transposon (40). Cyanobacterial transformations were performed as described earlier (42). E. coli strains DH10B and BL21DE3 hosted plasmids, and they were grown as described previously (43). Basic DNA manipulation was performed with standard procedures (18, 43).
Expression and Purification of Tagged Proteins.
Expression and purification of tagged proteins from S. elongatus under low-stringency conditions were performed as described previously (18) with minor modifications. KaiC-His (Fig. 1A) was induced with 7 μM isopropyl β-d-thiogalactoside (IPTG) for 24 h. This level of expression complemented a kaiC mutant, as judged by restoration of rhythmicity in a bioluminescent reporter gene assay (44). Expression of CikA-His to the WT level was induced with 10 μM IPTG for 24 h (Fig. 1B). For the experiment shown in Fig. 6, cultures were incubated for 24 h in the presence of 100 μM IPTG. Eluted fractions were analyzed directly by 12.5% SDS/PAGE (43), with the exception of the CikA-His fraction used to detect KaiA (Fig. 1B), which was concentrated 25-fold with trichloroacetic acid precipitation. Each experiment was repeated at least three times, yielding essentially the same results. The PsR domain was purified from E. coli AM3677, as described earlier (38).
Gel-Filtration Chromatography.
Chromatographic analysis was performed as described previously (19) with some modifications. WT S. elongatus (400 ml at OD750 = 0.7) was collected at 1,500 × g for 10 min and washed once in 2 ml of BB5 buffer. The pellet was resuspended in 0.25 ml of BB5 buffer containing 1 mM ATP, 5 μg/ml DNase I, 10 μg/ml RNase, and a protease inhibitor mixture for use with bacterial cell extracts (Sigma, St. Louis, MO) according to the manufacturer's recommendations. The soluble fraction was prepared as described (45), brought to a total volume of 3 ml, and loaded on a HiPrep 26/60 Sephacryl S-300 HR (GE Healthcare, Piscataway, NJ) equilibrated with BB5 buffer with 1 mM ATP by using an ÄKTA Explorer chromatography system (Amersham Biosciences, Piscataway, NJ). Protein fractions were eluted by using BB5 buffer with 1 mM ATP. To detect CikA and KaiA proteins, the fractionated protein samples were analyzed directly by SDS/PAGE (12.5% gel) (43). To detect KaiC, 1 ml of each fractionated protein sample was precipitated with trichloroacetic acid to concentrate the sample ≈25-fold before SDS/PAGE separation. Protein standards for calibration of the column included thyroglobulin (669 kDa), ferritin (440 kDa), and lactose dehydrogenase (140 kDa) (GE Healthcare). At least three independent experiments were performed for each time point, yielding essentially identical results.
Immunoblot Analysis.
Immunoblot analysis was performed as described earlier (18, 46). Proteins were transferred to nitrocellulose membranes by using either capillary (43) or electrophoretic transfer with a Transblot SD semidry transfer cell according to the manufacturer's recommendations (Bio-Rad, Hercules, CA). Phosphorylated and unphosphorylated bands of KaiC (47) were confirmed by treatment of samples with lambda protein phosphatase (New England Biolabs, Beverly, MA).
Light Treatment.
Cyanobacterial cultures (100 ml) were synchronized by two LD cycles. One set of cultures was kept in LL, and the second set was subjected either to dark-pulse treatment or to LD. For Fig. 2 samples (1 ml, OD750 = 0.7) were collected at ZT8 (8 h after coming out of LD entrainment), at 1 h, 3 h, and 5 h from the start of the dark pulse, at 1 h after the dark pulse ended (+1), and at corresponding times of LL. For Fig. 3A, samples (1 ml) were collected every 4 h from cells in LD or LL. Cells were collected for 2 min at 16,000 × g, and pellets were frozen at −80°C before processing. The total protein fraction was prepared as described (18) with minor modifications. Pellets were resuspended in 70 μl of ice-cold IA lysis buffer (18) and kept on ice for the remainder of the procedure. Cells were broken with glass beads (100 μg) for 1.5 min in a MiniBeadbeater-8 (BioSpec Products, Bartlesville, OK). Ice-cold IA lysis buffer was added (60 μl). Samples were mixed by vortexing and spun at 1,000 × g to remove beads, and the supernatant fractions were collected, yielding whole-cell extract. A sample (8 μg of protein) was loaded for each lane of SDS/PAGE.
Treatment with Inhibitors.
For Fig. 3B, the following inhibitors were added to the cyanobacterial culture to a final concentration of 10 μM: DBMIB, DCMU, CCCP, methylamine hydrochloride, and hydroxylamine hydrochloride. All inhibitors were from Sigma except DCMU (Serva, Germany). Because inhibitors were dissolved in ethanol (except hydroxylamine, dissolved in water), an equal volume of ethanol was added to the control sample. Cells at OD750 = 0.7 (1 ml) were treated for 15 min and collected for 2 min at 16,000 × g. The total protein fraction was prepared as described above. For Fig. 3C, chloramphenicol was added to samples (250 μg/ml) 5 min before the addition of DBMIB. For Fig. 4, samples were treated as for Fig. 3B, except a gradient of DBMIB concentrations was used (3, 5, and 10 μM).
NMR Analysis.
NMR samples of 15N-enriched PsR domain were prepared as described earlier (38). Final protein concentrations were 0.2 mM. NMR spectra of the PsR domain alone and in the presence of DBMIB (0.2 mM) were acquired on an Inova spectrometer (Varian, Palo Alto, CA) operating at 600-MHz proton frequency. An equal volume of ethanol was added to the control sample to a final concentration of 0.1%. The sample temperature was maintained at 25°C during data acquisition. Two-dimensional 1H,15N correlation spectra were collected as described previously (48). Acquisition times were 64 ms in both the t2 and t1 dimensions. All chemical shifts were referenced to internal DSS (49).
Supplementary Material
Acknowledgments
We thank Dr. T.C. Hall for use of the chromatography system, Dr. K. Tanaka for RpoD1 antiserum, and members of the clocks journal club at Texas A&M for insightful suggestions. Our laboratory members, Y. Chen, G. Dong, Dr. K. Holtman, and Dr. S. Mackey, contributed strains and helpful advice. The Gene Technologies Laboratory (Texas A&M University) provided sequencing and oligonucleotide services. This work was supported in part by National Institutes of Health Grants GM62419 and NS39546 (to S.S.G.) and GM064576 (to A.C.L.). N.B.I. was supported by a National Science Foundation–North Atlantic Treaty Organization postdoctoral fellowship. The NMR instrumentation in the Biomolecular NMR Laboratory at Texas A&M University was supported by National Science Foundation Grant DBI-9970232.
Abbreviations
- CCCP
carbonylcyanide m-chlorophenylhydrazone
- DBMIB
2,5-dibromo-3-methyl-6-isopropyl-p-benzoquinone
- DCBQ
2,5-dichloro-1,4-benzoquinone
- DCMU
3-(3,4-dichlorophenyl)-1,1-dimethylurea
- LD
12-h light/12-h dark cycle
- LL
constant light
- PET
photosynthetic electron transport
- PQ
plastoquinone
- PsR domain
pseudo-receiver domain
- ZT
zeitgeber time.
Footnotes
The authors declare no conflict of interest.
This article is a PNAS direct submission.
Data deposition: The chemical shift reported in this paper has been deposited in the Biological Magnetic Resonance Data Bank, www.bmrb.wisc.edu (accession no. 6438).
References
- 1.Bünning E. The Physiological Clock. New York: Springer; 1973. [Google Scholar]
- 2.Green RM, Tingay S, Wang ZY, Tobin EM. Plant Physiol. 2002;129:576–584. doi: 10.1104/pp.004374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ouyang Y, Andersson CR, Kondo T, Golden SS, Johnson CH. Proc Natl Acad Sci USA. 1998;95:8660–8664. doi: 10.1073/pnas.95.15.8660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bell-Pedersen D, Cassone VM, Earnest DJ, Golden SS, Hardin PE, Thomas TL, Zoran MJ. Nat Rev Genet. 2005;6:544–556. doi: 10.1038/nrg1633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kondo T, Tsinoremas NF, Golden SS, Johnson CH, Kutsuna S, Ishiura M. Science. 1994;266:1233–1236. doi: 10.1126/science.7973706. [DOI] [PubMed] [Google Scholar]
- 6.Ishiura M, Kutsuna S, Aoki S, Iwasaki H, Andersson CR, Tanabe A, Golden SS, Johnson CH, Kondo T. Science. 1998;281:1519–1523. doi: 10.1126/science.281.5382.1519. [DOI] [PubMed] [Google Scholar]
- 7.Nakajima M, Imai K, Ito H, Nishiwaki T, Murayama Y, Iwasaki H, Oyama T, Kondo T. Science. 2005;308:414–415. doi: 10.1126/science.1108451. [DOI] [PubMed] [Google Scholar]
- 8.Schmitz O, Katayama M, Williams SB, Kondo T, Golden SS. Science. 2000;289:765–768. doi: 10.1126/science.289.5480.765. [DOI] [PubMed] [Google Scholar]
- 9.Katayama M, Kondo T, Xiong J, Golden SS. J Bacteriol. 2003;185:1415–1422. doi: 10.1128/JB.185.4.1415-1422.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Iwasaki H, Williams SB, Kitayama Y, Ishiura M, Golden SS, Kondo T. Cell. 2000;101:223–233. doi: 10.1016/S0092-8674(00)80832-6. [DOI] [PubMed] [Google Scholar]
- 11.Smith RM, Williams SB. Proc Natl Acad Sci USA. 2006;103:8564–8569. doi: 10.1073/pnas.0508696103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ditty JL, Canales SR, Anderson BE, Williams SB, Golden SS. Microbiology. 2005;151:2605–2613. doi: 10.1099/mic.0.28030-0. [DOI] [PubMed] [Google Scholar]
- 13.Tomita J, Nakajima M, Kondo T, Iwasaki H. Science. 2005;307:251–254. doi: 10.1126/science.1102540. [DOI] [PubMed] [Google Scholar]
- 14.Williams SB, Vakonakis I, Golden SS, LiWang AC. Proc Natl Acad Sci USA. 2002;99:15357–15362. doi: 10.1073/pnas.232517099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Crosthwaite SK, Dunlap JC, Loros JJ. Science. 1997;276:763–769. doi: 10.1126/science.276.5313.763. [DOI] [PubMed] [Google Scholar]
- 16.Ceriani MF, Darlington TK, Staknis D, Mas P, Petti AA, Weitz CJ, Kay SA. Science. 1999;285:553–556. doi: 10.1126/science.285.5427.553. [DOI] [PubMed] [Google Scholar]
- 17.Mutsuda M, Michel KP, Zhang X, Montgomery BL, Golden SS. J Biol Chem. 2003;278:19102–19110. doi: 10.1074/jbc.M213255200. [DOI] [PubMed] [Google Scholar]
- 18.Ivleva NB, Bramlett MR, Lindahl PA, Golden SS. EMBO J. 2005;24:1202–1210. doi: 10.1038/sj.emboj.7600606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kageyama H, Kondo T, Iwasaki H. J Biol Chem. 2003;278:2388–2395. doi: 10.1074/jbc.M208899200. [DOI] [PubMed] [Google Scholar]
- 20.Golden SS. Proc Natl Acad Sci USA. 2004;101:13697–13698. doi: 10.1073/pnas.0405623101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Zhang X, Dong G, Golden SS. Mol Microbiol. 2006;60:658–668. doi: 10.1111/j.1365-2958.2006.05138.x. [DOI] [PubMed] [Google Scholar]
- 22.Nishiwaki T, Iwasaki H, Ishiura M, Kondo T. Proc Natl Acad Sci USA. 2000;97:495–499. doi: 10.1073/pnas.97.1.495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Escoubas JM, Lomas M, LaRoche J, Falkowski PG. Proc Natl Acad Sci USA. 1995;92:10237–10241. doi: 10.1073/pnas.92.22.10237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Trebst A. Methods Enzymol. 1980;69:675–715. [Google Scholar]
- 25.Schmitz O, Tsinoremas NF, Schaefer MR, Anandan S, Golden SS. Photosynth Res. 1999;62:261–271. [Google Scholar]
- 26.Fisher N, Rich PR. J Mol Biol. 2000;296:1153–1162. doi: 10.1006/jmbi.2000.3509. [DOI] [PubMed] [Google Scholar]
- 27.Bock A, Gross R. Eur J Biochem. 2002;269:3479–3484. doi: 10.1046/j.1432-1033.2002.03029.x. [DOI] [PubMed] [Google Scholar]
- 28.Kiyohara YB, Katayama M, Kondo T. J Bacteriol. 2005;187:2559–2564. doi: 10.1128/JB.187.8.2559-2564.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Stokkan KA, Yamazaki S, Tei H, Sakaki Y, Menaker M. Science. 2001;291:490–493. doi: 10.1126/science.291.5503.490. [DOI] [PubMed] [Google Scholar]
- 30.Rutter J, Reick M, Wu LC, McKnight SL. Science. 2001;293:510–514. doi: 10.1126/science.1060698. [DOI] [PubMed] [Google Scholar]
- 31.Pfannschmidt T, Liere K. Antioxid Redox Signal. 2005;7:607–618. doi: 10.1089/ars.2005.7.607. [DOI] [PubMed] [Google Scholar]
- 32.Barnes D, Mayfield SP. Antioxid Redox Signal. 2003;5:89–94. doi: 10.1089/152308603321223577. [DOI] [PubMed] [Google Scholar]
- 33.Georgellis D, Kwon O, Lin EC. Science. 2001;292:2314–2316. doi: 10.1126/science.1059361. [DOI] [PubMed] [Google Scholar]
- 34.Georgellis D, Kwon O, Lin EC, Wong SM, Akerley BJ. J Bacteriol. 2001;183:7206–7212. doi: 10.1128/JB.183.24.7206-7212.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Stock AM, Robinson VL, Goudreau PN. Annu Rev Biochem. 2000;69:183–215. doi: 10.1146/annurev.biochem.69.1.183. [DOI] [PubMed] [Google Scholar]
- 36.Taylor BL, Zhulin IB. Microbiol Mol Biol Rev. 1999;63:479–506. doi: 10.1128/mmbr.63.2.479-506.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Ho YS, Burden LM, Hurley JH. EMBO J. 2000;19:5288–5299. doi: 10.1093/emboj/19.20.5288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Gao T, Zhang X, Xia Y, Cho Y, Sacchettini JC, Golden SS, Liwang AC. J Biomol NMR. 2005;32:259. doi: 10.1007/s10858-005-7945-5. [DOI] [PubMed] [Google Scholar]
- 39.Hwang HJ, Burnap RL. Biochemistry. 2005;44:9766–9774. doi: 10.1021/bi050069p. [DOI] [PubMed] [Google Scholar]
- 40.Holtman CK, Chen Y, Sandoval P, Gonzales A, Nalty MS, Thomas TL, Youderian P, Golden SS. DNA Res. 2005;12:103–115. doi: 10.1093/dnares/12.2.103. [DOI] [PubMed] [Google Scholar]
- 41.Mas P, Kim WY, Somers DE, Kay SA. Nature. 2003;426:567–570. doi: 10.1038/nature02163. [DOI] [PubMed] [Google Scholar]
- 42.Clerico EM, Ditty JL, Golden SS. In: Methods in Molecural Biology. Rosato E, editor. Totowa, NJ: Humana; 2006. pp. 155–171. [Google Scholar]
- 43.Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning: A Laboratory Manual. 2nd Ed. Cold Spring Harbor, NY: Cold Spring Harbor Lab Press; 1989. [Google Scholar]
- 44.Mackey SR, Ditty JL, Clerico EM, Golden SS. In: Methods in Molecular Biology. Rosato E, editor. Totowa, NJ: Humana; 2006. pp. 115–129. [DOI] [PubMed] [Google Scholar]
- 45.Ivleva NB, Golden SS. In: Methods in Molecular Biology. Rosato E, editor. Totowa, NJ: Humana; 2006. pp. 365–373. [DOI] [PubMed] [Google Scholar]
- 46.Asayama M, Suzuki H, Sato A, Aida T, Tanaka K, Takahashi H, Shirai M. J Biochem (Tokyo) 1996;120:752–758. doi: 10.1093/oxfordjournals.jbchem.a021475. [DOI] [PubMed] [Google Scholar]
- 47.Iwasaki H, Nishiwaki T, Kitayama Y, Nakajima M, Kondo T. Proc Natl Acad Sci USA. 2002;99:15788–15793. doi: 10.1073/pnas.222467299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Kay L, Keifer EP, Saarinen T. J Am Chem Soc. 1992;114:10663–10665. [Google Scholar]
- 49.Markley JL, Bax A, Arata Y, Hilbers CW, Kaptein R, Sykes BD, Wright PE, Wüthrich K. Pure Appl Chem. 1998;70:117–142. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}
{kind=link}
{kind=link}