

Summary
The ability to reflect on one’s own mental processes, termed metacognition, is a defining feature of human existence [1, 2]. Consequently, a fundamental question in comparative cognition is whether non-human animals have knowledge of their own cognitive states [3]. Recent evidence suggests that people and non-human primates [4–8], but not less “cognitively sophisticated” species [3, 9, 10], are capable of metacognition. Here we demonstrate for the first time that rats are capable of metacognition – i.e., they know when they do not know the answer in a duration-discrimination test. Before taking the duration test, rats were given the opportunity to decline the test. On other trials, they were not given the option to decline the test. Accurate performance on the duration test yielded a large reward whereas inaccurate performance resulted in no reward. Declining a test yielded a small, but guaranteed, reward. If rats possess knowledge about whether they know or do not know the answer to the test, they would be expected to decline most frequently on difficult tests and show lowest accuracy on difficult tests that cannot be declined [4]. Our data provide evidence for both predictions and suggest that a non-primate has knowledge of its own cognitive state.
Results and Discussion
People are sometimes aware of their own cognitive processes. For example, a college student entering a classroom to take a test will often have some knowledge about how she will perform on the test. It is noteworthy that this knowledge (whether accurate or inaccurate) is available before the student actually responds to the test questions and obtains feedback about performance. This familiar experience (i.e., knowing that we know or do not know the answer) is easily assessed in humans by requesting verbal reports about our experiences. Of course, this option is not available with nonverbal species. Consequently, researchers in comparative cognition have sought to identify experimental conditions in which a human or non-human subject could demonstrate through its behavior knowledge of a cognitive state. Therefore, studies in metacognition test the hypothesis that animals behave functionally the same as an organism that is aware of its own cognitive state.
One approach used to study metacognition in non-humans [4] is to give the animal an option to decline to take a test. Presumably, an animal that knows that it does not know the answer to a test question will decline to take the test. Moreover, being forced to take a test is likely to degrade performance because forced tests include trials that would have been declined had that option been available. Although considerable evidence supports the existence of metacognition in primates, a paucity of research has been conducted with other mammalian species. Developing a rodent model of metacognition may allow for new opportunities to explore its underlying neural mechanisms. To this end, we adapted Hampton’s [4] experimental design with monkeys for an experiment with rats.
Each trial consisted of three phases: study, choice and test phases (Figure 1). In the study phase, a brief noise was presented for the subject to classify as short (2–3.62 s) or long (4.42–8 s). Stimuli with intermediate durations (e.g., 3.62 and 4.42 s) are most difficult to classify as short or long [11, 12]. By contrast, more widely spaced intervals (e.g., 2 and 8 s) are easiest to classify. In the choice phase, the rat was sometimes presented with two response options, signaled by the illumination of two nose-poke apertures. On these choice-test trials, a response in one of these apertures (referred to as a take-the-test response) led to the insertion of two response levers in the subsequent test phase; one lever was designated as the correct response after a short noise, and the other lever was designated correct after a long noise. The other aperture (referred to as the decline-the-test response) led to the omission of the duration test. On other trials in the choice phase, the rat was presented with only one response option; on these forced-test trials, the rat was required to select the aperture that led to the duration test (i.e., the option to decline the test was not available), which was followed by the duration test. In the test phase, a correct lever press with respect to the duration discrimination produced a large reward of 6 pellets; an incorrect lever press produced no reward. A decline response (provided that this option was, indeed, available) led to a guaranteed, but smaller, reward of 3 pellets.
Figure 1. Schematic Representation of Experimental Design Showing Trial Sequence in Study, Choice, and Test Phases.
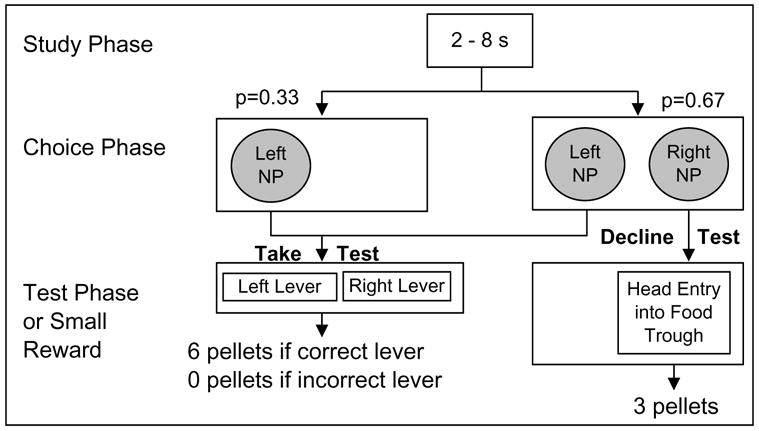
After presenting a white noise (2–8 s; study phase), a choice phase provided an opportunity to take or decline a duration test; declining the test produced a guaranteed, but smaller, reward than was earned if the test was selected and answered correctly (test phase). To the extent that a rat is uncertain about how to classify the interval, it will (1) decline to take the test and (2) show lower accuracy when forced to take the test relative to trials on which it chose to take the test. Grey shading indicates an illuminated nose-poke (NP) aperture.
The rate of declining to take the test increased as the difficulty of the discrimination increased (Figure 2A–D; see also Figure S1 in the Supplemental Data available with this article online). This observation was confirmed for the mean of the rats (F(3,6)=17.6, p<.01, Figure 2D) and for each rat separately (p< .01, .05, and .01 for panels 2A–C, respectively). The horizontal axis in Figure 2 represents an index of stimulus difficulty (see Data Analysis section below for details). For example, the leftmost point in each panel corresponds to performance after the study item was 2 or 8 s, which are the easiest stimuli to discriminate. By contrast, the rightmost point in each panel corresponds to the most difficult durations to discriminate (3.62 and 4.42 s).
Figure 2. Rats Decline Difficult Tests and Show Degraded Accuracy When They Cannot Decline the Difficult Test.
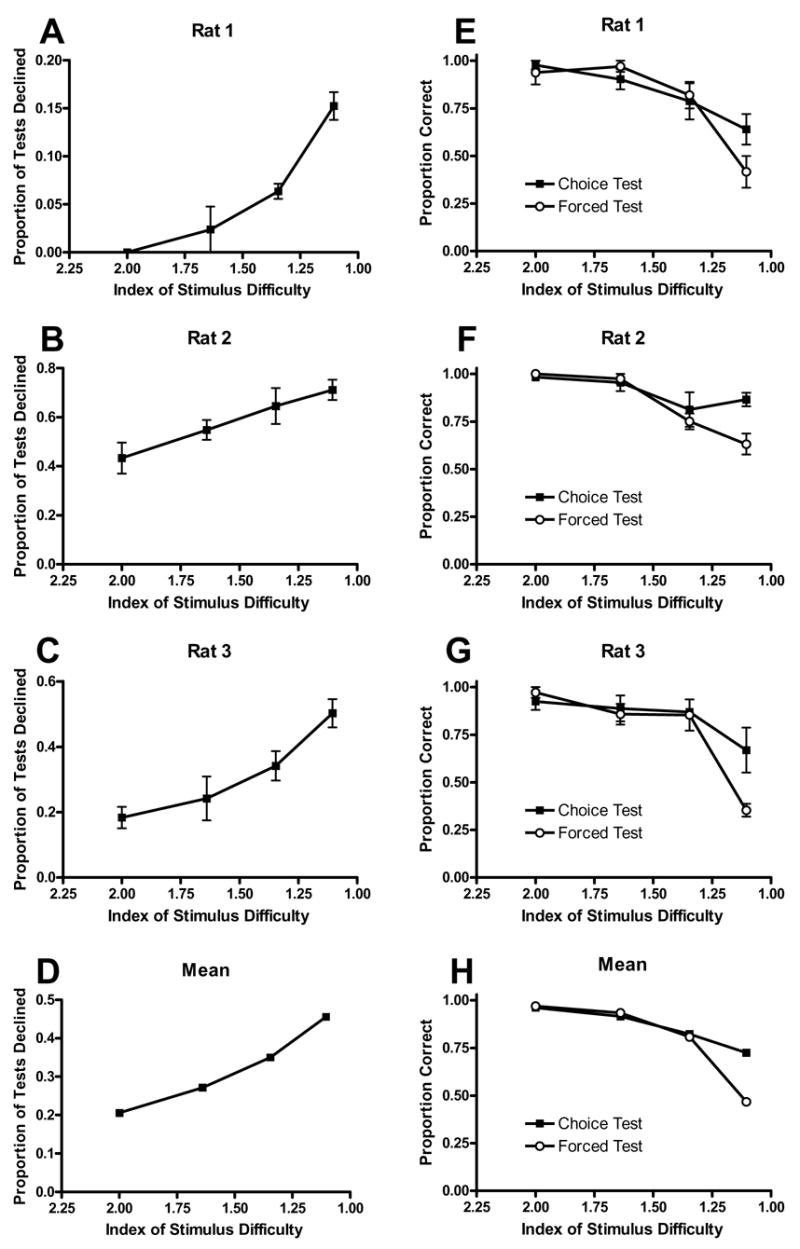
(A–D) Difficult tests (defined as stimulus duration near the subjective middle of the shortest and longest durations) were declined more frequently than easy tests (defined as stimulus durations near the shortest and longest durations).
(E–H) The decline in accuracy as a function of stimulus difficulty was more pronounced when tests could not be declined (forced test) compared to tests that could have been declined (choice test).
The x-axes are plotted in reversed order. Error bars represent SEM.
Accuracy declined as the difficulty of the discrimination increased, but this decline was greater when the rats were forced to take the test compared to trials on which the rats chose to take the test (Figure 2E–H). This observation was confirmed by an interaction of trial type and stimulus difficulty for the mean of the rats (F(3,6)=20.3, p<.01, Figure 2H) and for each rat separately (p<.05, .01, and .05 for panels 2E–G, respectively). No difference between choice- and forced-test accuracy is expected for easy discriminations because there is little room for improvement. However, for the most difficult discrimination (right most data in Figure 2E–H; 3.62- and 4.42-s stimuli), each rat showed higher accuracy on trials in which it had chosen to take the test.
The present experiment suggests that rats know when they do not know the answer to a duration discrimination. The duration discrimination involved classifying brief noises as short or long. Consequently, durations near the middle of the range are more difficult to classify. The rats were more likely to decline these difficult tests, as would be expected if the rats knew that they did not know the correct duration-discrimination response. Moreover, when we compared accuracy on the duration test, the rats were more accurate when they had chosen to take the duration test compared to trials in which they were forced to take the test. This pattern is predicted by the metacognition hypothesis because forced-test performance includes trials that would have been declined had the option to do so been available. Knowledge about temporal information may play a central role in the organization of behavior [13], and consequently, detailed knowledge about when events occur [14–20] may have contributed to our experimental evidence for metacogntion.
An important feature of the design of this study is that we evaluated accuracy on trials in which the rats were forced to take the test. Moreover, the rats initiated these forced tests in the same manner as they initiated choice tests (i.e., with a nose-poke response). These factors limit the ability of alternative hypotheses to explain our data. For example, the forced duration tests are unlikely to have been unexpected, thereby disrupting accuracy. If forced choices had been surprising, we would expect longer latencies to respond in forced tests relative to choice tests. However, there was no evidence for a difference in the latency to respond with either a nose-poke (t(2)=1.1, p>.05; mean ± SEM: 0.61 ± 0.19 and 0.80 ± 0.12 s for choice- and forced-tests, respectively) or lever-press response (t(2)=0.4, p>.05; mean ± SEM: 2.70 ± 0.47 and 2.65 ± 0.28 s for choice- and forced-tests, respectively). Consequently, the accuracy difference on forced and choice tests was not likely due to performance disruption from unexpected forced tests.
Similarly, discrimination of a purely behavioral state (e.g., physical proximity to the response levers), in contrast to discrimination of a cognitive state, is unlikely to fully account for our data. Discrimination of a purely behavioral state predicts equivalent performance on forced- and choice-tests, which is not supported by our data (Figure 2E–H). Although our data do not preclude the possibility that the discrimination of behavioral states contribute to the use of the decline response, such a contribution could be empirically tested in future experiments by collecting video recordings of the rats’ behavior during stimulus presentation. It is also unlikely that learning a contingency between reinforcement rate and specific stimulus durations could account for our data. First, the observed difference in accuracy between choice- and forced-tests is not predicted by differential reinforcement associated with specific stimulus durations. Second, for a learned contingency to contribute to the observed increase in the decline rate, one must presuppose that rats would prefer guaranteed small rewards more than uncertain large rewards (i.e., that rats are risk averse). However, there is evidence that rats are risk prone in a situation similar to our own [21]. Ultimately, it is important to experimentally dissociate stimulus identity and difficulty by evaluating the ability of rats to generalize the use of the decline response to a different discrimination, which has recently been reported with monkeys [7, 8].
As Hampton [4] has argued, although observing putative subjective experiences that may accompany metacognition is not possible, progress in the study of evolution of the human mind and in evaluating the validity of animal models of human cognition will come from documenting objective functional features of human conscious cognition. To this end, we have documented the ability of rats to make adaptive decisions about future behavior contingent on the current availability of knowledge. Developing a rodent model of metacognition may promote new opportunities to explore the neuroanatomical, neurochemical, neurophysiological, and molecular mechanisms of metacognition.
Experimental Procedures
Animals
Eight male Sprague-Dawley rats (Harlan, Madison, WI; 249 g, 85 days old) were individually housed in a room with a reversed light cycle (light offset at 0700, onset at 1900). The rats consumed 45-mg pellets (PJAI-0045, Research Diets Inc., New Brunswick, NJ) during testing sessions and a ration of 5001-Rodent-Diet (Lab Diet, Brentwood, MO), which was adjusted so that total daily intake was 15–20 g. Water was continuously available. All procedures were approved by the institutional animal care and use committee and followed guidelines of the National Research Council Guide for the Care and Use of Laboratory Animals. Five rats rarely declined to take the duration test (M = 97.8%, SEM = ± .01%). The performance for these five rats was likely due to response bias as it appears that these rats failed to learn the experimental contingency of the nose-poke apertures. As a result, these five rats did not provide evidence for or against metacognition. The rats had participated in a pilot study, in which they received a similar training regimen to that described below, during which the reward sizes and intertrial interval were adjusted. All subjects (n=3) that learned the experimental contingency are included in the data analysis.
Apparatus
Identical operant chambers (described in [22]), each located in a ventilated sound attenuation cubicle, contained a recessed food trough (equipped with a photobeam used to detect head entries) centered between two levers on one wall of the chamber. A 45-mg pellet dispenser was positioned outside the chamber and attached to the food trough. A water bottle was placed outside of the chamber with the tube inserted across from the food trough. This wall also contained nose-poke apertures on the left and right sides of the sipper tube. Small white lights were recessed at the back of each nose-poke aperture. Each nose-poke contained a photobeam that detected the entry of the rat’s snout. The chamber floor was constructed from nineteen stainless steel rods. Other equipment in each chamber included a speaker, clicker, lights, and additional photobeams. Lever-press and nose-poke responses were recorded (10-msec resolution) using MED-PC software (version 4.1) on a computer in a nearby room.
Procedure
Pretraining
Rats were given one pellet per minute accompanied by a click for 30 min per day on the first pretraining session. On subsequent pretraining sessions, the left lever was inserted, and each food pellet was delivered contingent on a single lever press until 10 pellets had been earned. Next, the left lever was retracted and the right lever was inserted until 10 additional pellets had been earned. The lever training session continued, in this manner, until 60 min had passed or 60 pellets had been earned.
Duration Discrimination Training
Rats were initially trained to discriminate short and long durations. A trial consisted of presentation of a white noise (70 dB) for a randomly and independently selected duration (2.00, 2.44, 2.97, 3.62, 4.42, 5.38, 6.56, and 8.00 s; equally spaced on a logarithmic scale), insertion of two response levers, the rat’s lever press, feedback, and retraction of the levers, which was followed by an intertrial interval of 45 s. The first four durations (i.e., values below 4 s, designated as short durations) were rewarded for pressing one lever (e.g., left) and the other four durations (values above 4 s, designated as long) were rewarded for pressing the other lever (e.g., right) as described elsewhere [23, 24]; the assignment of left and right levers was counterbalanced across rats prior to the start of the experiment. Each session lasted 2 hours, 5 days per week, for 35 sessions.
Nose-Poke Pretraining
Rats were trained for three daily sessions to break the photobeam in the nose-poke apertures. At the beginning of each session, the left nose-poke light was illuminated and the rat was required to break the photobeam in the left nose-poke aperture to earn a reward. After the rat had broken the left nose-poke photobeam, the light in the left nose poke aperture was turned off and a pellet and a click were delivered. This process continued for 10 pellets on the left nose-poke aperture followed by a switch to the right nose-poke aperture for the next 10 pellets. The nose-poke trials continued, in this manner, until 60 min had elapsed or 60 pellets had been earned.
Testing
A noise was presented, as described above in Duration Discrimination Training. In the choice phase, two nose-poke apertures were illuminated in two-thirds of the trials (see Figure 1, right side). Breaking the photobeam inside one of the nose-poke apertures (designated as the take-the-test nose-poke response) caused the lights to be extinguished, the levers to be inserted, and a duration test to be initiated as described above. Breaking the photobeam inside the other nose-poke aperture (designated as the decline-the-test nose-poke response) caused the lights to be extinguished, but did not produce insertion of the levers. Instead, the rat was required to break a photobeam inside the food trough to obtain food and terminate the trial. The assignment of left and right nose-poke apertures to take and decline responses was counterbalanced across rats before the experiment began. In the remaining one-third of trials, only the take-the-test nose-poke aperture was illuminated (see Figure 1, left side); a response in the illuminated nose-poke aperture was required to insert the levers and initiate a duration test, as described above. By contrast, a response in the other (i.e., dark) nose-poke aperture did not advance the rat to the next phase of the trial. Choice- and forced-tests were randomly intermixed throughout the session. The reward size was 6 or 0 pellets after a correct or incorrect duration-discrimination lever press, respectively. If the rat declined the duration test, the reward size was 3 pellets. The inter-trial interval was 8 min, and each session lasted 9 hr, for 22–26 sessions (1546 trials, on average). The long inter-trial interval may have reduced confusion between individual trials and enhanced the salience of the auditory stimulus.
Data Analysis
Stimulus durations were equally spaced on a logarithmic scale to ensure that pairs of durations were equated for difficulty. When rats are trained to discriminate 2 and 8 s, the point of subjective equality (i.e., the stimulus duration at which the probability of judging the stimulus as short or long are equal) occurs at 4 s [11, 12], which is the geometric mean of 2 and 8 s (i.e., the midpoint on a logarithmic scale). Note that the absolute difference between 2 and 4 and between 4 and 8 are equal on a logarithmic scale; therefore, a point of subjective equality at 4 s means that 2 and 8 s are equated for stimulus difficulty [11, 12]. Similarly, the second and seventh stimuli (2.44 and 6.56 s) are equally spaced on a log scale with respect to 4 s, as are the third and sixth (2.97 and 5.38 s), and the fourth and fifth (3.62 and 4.42 s) stimuli. Consequently, we pooled the data within identical levels of stimulus difficulty to obtain 4 levels of stimulus difficulty. The index of stimulus difficulty (horizontal axes in Figure 2) was calculated as the absolute log distance of each stimulus duration from the geometric mean of 4 s, expressed in seconds. For example, the most difficult stimuli to discriminate are closely spaced (3.62 and 4.42 s) near the geometric mean (rightmost data in Figure 2; index of stimulus difficulty = 1.1), whereas the easiest stimuli to discriminate are widely spaced (2 and 8 s; leftmost data in Figure 2; index of stimulus difficulty = 2.0).
A median latency for each rat was calculated between duration-stimulus termination and the nose-poke response and between nose-poke and lever-press responses for choice- and forced-test trials.
Terminal performance (487 trials, on average) was analyzed with repeated measures analyses of variance. P < .05 was considered significant.
Supplementary Material
Acknowledgments
This work was supported by National Institute of Mental Health grants MH64799 to J.D.C. The raw data (time of occurrence of each stimulus and response) are available at www.uga.edu/animal-cognition-lab.
Footnotes
Supplemental Data include one figure and can be found with this article online at
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Descartes R. Discourse on method. 1637. [Google Scholar]
- 2.Metcalfe J, Kober H. Self-reflective consciousness and the projectable self. In: Terrace H, Metcalfe J, editors. The Missing Link in Cognition: Origions of Self-Reflective Consciousness. New York: Oxford University Press; 2005. pp. 57–83. [Google Scholar]
- 3.Smith JD, Shields WE, Washburn DA. The comparative psychology of uncertainty monitoring and metacognition. Beh Brain Sci. 2003;26:317–373. doi: 10.1017/s0140525x03000086. [DOI] [PubMed] [Google Scholar]
- 4.Hampton R. Rhesus monkeys know when they remember. Proc Natl Acad Sci USA. 2001;98:5359–5362. doi: 10.1073/pnas.071600998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Smith JD, Shields WE, Allendoerfer KR, Washburn DA. Memory monitoring by animals and humans. J Exp Psychol Gen. 1998;127:227–250. doi: 10.1037//0096-3445.127.3.227. [DOI] [PubMed] [Google Scholar]
- 6.Smith JD, Shields WE, Schull J, Washburn DA. The uncertain response in humans and animals. Cognition. 1997;62:75–97. doi: 10.1016/s0010-0277(96)00726-3. [DOI] [PubMed] [Google Scholar]
- 7.Washburn DA, Smith JD, Shields WE. Rhesus monkeys (Macaca mulatta) immediately generalize the uncertain response. J Exp Psychol: Anim Behav Process. 2006;32:185–189. doi: 10.1037/0097-7403.32.2.185. [DOI] [PubMed] [Google Scholar]
- 8.Kornell N, Son LK, Terrace HS. Metacognition in monkeys. Psychol Sci in press. [Google Scholar]
- 9.Inman A, Shettleworth SJ. Detecting metamemory in nonverbal subjects: A test with pigeons. J Exp Psychol: Anim Behav Process. 1999;25:389–395. [Google Scholar]
- 10.Sole LM, Shettleworth SJ, Bennett PJ. Uncertainty in pigeons. Psychonom Bull Rev. 2003;10:738–745. doi: 10.3758/bf03196540. [DOI] [PubMed] [Google Scholar]
- 11.Church RM, Deluty MZ. Bisection of temporal intervals. J Exp Psychol: Anim Behav Process. 1977;3:216–228. doi: 10.1037//0097-7403.3.3.216. [DOI] [PubMed] [Google Scholar]
- 12.Stubbs DA. Scaling of stimulus duration by pigeons. J Exp Anal Behav. 1976;26:15–25. doi: 10.1901/jeab.1976.26-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gallistel CR. The Organization of Learning. Cambridge, MA: MIT Press; 1990. [Google Scholar]
- 14.Clayton NS, Dickinson A. Episodic-like memory during cache recovery by scrub jays. Nature. 1998;395:272–274. doi: 10.1038/26216. [DOI] [PubMed] [Google Scholar]
- 15.Babb SJ, Crystal JD. Discrimination of what, when, and where: Implications for episodic-like memory in rats. Learn Motiv. 2005;36:177–189. [Google Scholar]
- 16.Babb SJ, Crystal JD. Discrimination of what, when, and where is not based on time of day. Learn Behav. 2006;34:124–130. doi: 10.3758/bf03193188. [DOI] [PubMed] [Google Scholar]
- 17.Babb SJ, Crystal JD. Episodic-like memory in the rat. Curr Biol. 2006;16:1317–1321. doi: 10.1016/j.cub.2006.05.025. [DOI] [PubMed] [Google Scholar]
- 18.Clayton NS, Bussey TJ, Dickinson A. Can animals recall the past and plan for the future? Nat Rev Neurosci. 2003;4:685–691. doi: 10.1038/nrn1180. [DOI] [PubMed] [Google Scholar]
- 19.Crystal JD. Time, place, and content. Comp Cogn Behav Rev. 2006;1:53–76. [Google Scholar]
- 20.Dally JM, Emery NJ, Clayton NS. Food-Caching Western Scrub-Jays Keep Track of Who Was Watching When. Science. 2006;312:1662–1665. doi: 10.1126/science.1126539. [DOI] [PubMed] [Google Scholar]
- 21.Mazur JE. Choice between small certain and large uncertain reinforcers. Anim Learn Behav. 1988;16:199–205. [Google Scholar]
- 22.Crystal JD. Long-interval timing is based on a self sustaining endogenous oscillator. Behav Process. 2006;72:149–160. doi: 10.1016/j.beproc.2006.01.010. [DOI] [PubMed] [Google Scholar]
- 23.Crystal JD. Systematic nonlinearities in the perception of temporal intervals. J Exp Psychol: Anim Behav Process. 1999;25:3–17. [PubMed] [Google Scholar]
- 24.Stubbs DA. Response bias and the discrimination of stimulus duration. 1976;25:243–250. doi: 10.1901/jeab.1976.25-243. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.