

Abstract
A fragment of the 16S RNA of Thermus thermophilus corresponding to the central domain (nucleotides 547–895) has been prepared by transcription in vitro. Incubation of this fragment with the total 30S ribosomal proteins has resulted in the formation of a compact 12S ribonucleoprotein particle. This particle contained five T. thermophilus proteins corresponding to Escherichia coli ribosomal proteins S6, S8, S11, S15, and possibly S18, all of which were previously shown to interact with the central domain of the 16S RNA and to be localized in the platform (side bulge) of the 30S ribosomal subunit. When examined by electron microscopy, isolated particles have an appearance that is similar in size and shape to the corresponding morphological features of the 30S subunit. We conclude that the central domain of the 16S RNA can independently and specifically assemble with a defined subset of ribosomal proteins into a compact ribonucleoprotein particle corresponding to the platform (side bulge) of the 30S subunit.
The secondary structure of the 16S RNA of the small 30S ribosomal subunit is divided into three major domains (1). Each domain and a defined set of ribosomal proteins are involved in the formation of different morphological parts of the subunit. The 5′ domain corresponds to the central body of the 30S subunit, the central domain is involved in the formation of the side bulge or platform, as well as part of the body, and the 3′ major domain is localized in the head region of the subunit (2–4).
It has been shown that fragments of the 16S RNA corresponding to the domains of the 16S RNA are capable of assembling in vitro with specific sets of ribosomal proteins into compact ribonucleoprotein particles (RNPs). Ofengand and coworkers (5) demonstrated formation of the RNP containing the 5′ domain of the 16S RNA and four ribosomal proteins. Noller and coworkers (6) showed that the fragment of the 16S RNA, corresponding to the 3′ domain of the 16S RNA, assembles with the group of eight ribosomal proteins. It is noteworthy that the proteins bound in the both complexes are the same as those found associated with these domains in the intact 30S subunit. In this work, we describe the assembly of an RNP containing the central domain of the 16S RNA and five ribosomal proteins of the Thermus thermophilus ribosome.
The sequence of the 16S RNA from T. thermophilus was determined, and a high homology of the sequences of the 16S RNA from T. thermophilus and Escherichia coli was shown (7). The homology is 75%, and the computer model of its secondary structure (Fig. 1) does not differ from the model of the E. coli 16S RNA (8). The ribosomal proteins of the 30S subunit of T. thermophilus were purified and identified both by two-dimensional gel electrophoresis and amino-terminal sequence analysis (9, 10). The homology of 17 of 19 30S ribosomal proteins from T. thermophilus with corresponding proteins of E. coli was established. Thus the results presented herein are interpretable within the framework of the well-known findings concerning the structure of the E. coli 30S ribosomal subunit.
Figure 1.

Secondary structure of the T. thermophilus 16S RNA (8). The outlined region corresponds to the central domain studied in this work.
Our results support the idea that the domain concept for the organization of the ribosome extends to its assembly (5, 6) and show that the central domain of the 16S RNA can also assemble independently of the rest of the 30S subunit into a specifically folded compact particle that resembles the corresponding morphological feature of the 30S subunit.
MATERIALS AND METHODS
Plasmid Construction and in Vitro Transcription.
We followed the routine PCR and cloning methods as described (11). All the enzymes used in this work were purified by us. E. coli SURE (Stratagene) was used for electrotransformation.
The central fragment of the 16S RNA gene of T. thermophilus HB8 was obtained by PCR amplification from the plasmid pBR322-16S Tth (8) containing the whole 16S RNA gene. The 5′ end oligonucleotide for PCR amplification (5′-ATATGCATCTGGGCGTAAA-3′) contained a NsiI site (underlined), and the 3′ oligonucleotide (5′-TATCTAGAAGACATTCCTTTGAGTTT-3′) contained XbaI and BbvII sites (underlined). The sites NsiI and XbaI were used for cloning the PCR product in the vector plasmid pGEM-9Zf(−) (Promega) under control of the SP6 promoter. The site of IIS type restriction enzyme BbvII (12) was used for run-off transcription in vitro with the SP6 RNA polymerase. In this case, no additional nucleotides from the vector molecule appeared at the 3′ end of the transcript. The fragment of the 16S RNA from 547 to 895 nucleotides was obtained as a result of the in vitro transcription. It had the sequence of 10 additional nucleotides GAAUAUGCAU at its 5′ end that originated from the vector; it is supposed not to interfere with the folding of the domain and the assembly of RNP particles from the obtained RNA and ribosomal proteins. The transcription in vitro was carried out according to ref. 13. The plasmid pSP64-lys with the lysozyme gene under control of the SP6 promoter was obtained from Amersham.
In Vitro Reconstitution.
After transcription with SP6 RNA polymerase, the mixture was treated with pancreatic DNase I at a final concentration of 0.005 mg/ml for 30 min at 37°C and then dialyzed overnight at 4°C vs. the Rec20 buffer (20 mM Tris⋅HCl, pH 7.6/330 mM KCl/20 mM MgCl2/1 mM DTT). Prior to reconstitution, the RNA was heated for 10 min at 40°C.
Total 30S ribosomal proteins (TP30) were extracted by treating the T. thermophilus 30S subunits with 6 M LiCl in the presence of 100 mM MgCl2 for 20 h at 4°C (10). Isolation of the T. thermophilus 30S ribosomal subunits was made as described (14). Prior to reconstitution, TP30 was dialyzed overnight at 4°C vs. Rec20 buffer. Reconstitution was performed at a final RNA concentration of 0.3 mg/ml by using a 2-fold molar excess of TP30 in Rec20 buffer for 40 min at 42°C. The reconstituted RNPs were purified by sucrose gradient centrifugation.
The protein composition of the RNPs was controlled by two-dimensional gel electrophoresis (15). Analytical centrifugation was carried out in the analytical scanning ultracentrifuge UCA-10 (SKB BFA, Moscow) with ultraviolet optics.
Electron Microscopy.
The samples of 30S subunits and the RNPs in the Rec20 buffer were negatively stained with 1% uranyl acetate by the single-layer carbon technique (16). Specimens were examined with a JEM-100C electron microscope at 80 kV and ×40,000 magnification.
RESULTS
Characterization of the Transcript.
The plasmid pGEM9-c16STth containing the DNA fragment corresponding to the central domain of the 16S ribosomal RNA of T. thermophilus was constructed as described above. Electrophoresis of the product of SP6 transcription in vitro of the plasmid linearized with BbvII in a denaturing 5% polyacrylamide gel shows a single band corresponding to RNA of the appropriate length (Fig. 2).
Figure 2.
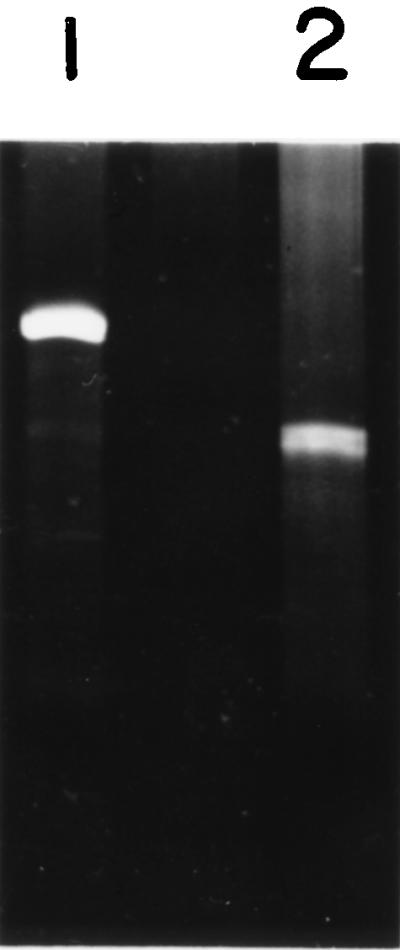
The 7 M urea/5% polyacrylamide gel electrophoresis analysis of in vitro transcripts. Lanes: 1, transcript of lysozyme gene of 527 nucleotides for linearized plasmid pSp64lys; 2, transcript of the DNA fragment of 359 nucleotides corresponding to the central domain of the 16S RNA.
In Vitro Reconstitution and Physical Characteristics of RNP.
The RNA fragment corresponding to the central domain of the 16S RNA (Fig. 1) was reconstituted with TP30 and purified by sucrose gradient centrifugation (Fig. 3). The fractions indicated by larger symbols were pooled for analysis by analytical centrifugation and two-dimensional gel electrophoresis. The yield of these RNPs was about 30%. Analytical centrifugation analysis revealed the homogeneity of the RNP complex (Fig. 4). The sedimentation coefficient measured in the Rec20 buffer was 12S. The isolated RNA fragment sedimented in the same buffer at about 8.5S (data not shown). The shift of the sedimentation coefficient value of the RNA after incubation with TP30 means that complex formation with ribosomal proteins did occur.
Figure 3.

Isolation of the reconstituted central domain RNPs. The reconstitution mixture was loaded on 5–20% linear sucrose gradient in Rec20 buffer and centrifuged for 17 h at 33,000 rpm at 4°C in a Spinco SW41 rotor. The fractions (0.5 ml) indicated by the larger symbols (fractions 5–8) were pooled for subsequent analysis.
Figure 4.

Sedimentation analysis of the central domain RNPs. Centrifugation was done in Rec20 buffer at 40,000 rpm in the analytical scanning ultracentrifuge UCA-10 with ultraviolet optics. Recording at 4-min intervals.
Protein Composition of the RNP.
The ribosomal proteins in the 12S RNP were extracted and analyzed by two-dimensional gel electrophoresis. Fig. 5 shows the two-dimensional gel electrophoresis pattern of the proteins extracted from original 30S subunits (Fig. 5B) and from the reconstituted RNP particles (Fig. 5A). All of the T. thermophilus small subunit proteins are present in original 30S subunit, whereas the RNP contains only proteins TS7 (homologous to S8 from E. coli), TS9 (S6), TS11 (S11), TS16 (S15), and TS20 (homology with the known proteins from E. coli is not revealed), as well as TS13 (S13) in clearly reduced amounts. Thus, the fragment of the 16S RNA corresponding to the central domain selects out a subset of the total 30S proteins. S6, S8, S11, and S15 are proteins that have binding sites in the central domain as determined by the footprinting experiments (19, 20). Among the proteins exclusively localized by the footprinting experiments to the central domain (19, 20), only S18 is absent from the 12S RNP. At the same time, protein TS20 whose homolog in E. coli is not known is present in the complex.
Figure 5.

Ribosomal protein composition of the 12S RNP particle (A) and of the original 30S subunits (B). Proteins were extracted as described (15) and analyzed by two-dimensional gel electrophoresis as described (17) with minor modifications. Nomenclature of ribosomal proteins is according to ref. 18.
Electron Microscopy.
Micrographs of the reconstituted central domain RNPs are presented in Fig. 6 in comparison with micrographs of the original T. thermophilus 30S subunits. The field of the RNP preparation contains particles very homogeneous in size and shape. The particles appear as compact elongated and slightly curved bodies. Their profiles have dimensions of approximately 100 × 60 Å. Such a shape and dimensions of the RNPs correspond well to the overall shape of the platform region in the recently published three-dimensional reconstruction of the 30S subunit (21), where the platform is found to be spade-shaped and extended, with the total length of about 100 Å.
Figure 6.

Electron micrographs of the T. thermophilus 30S subunits (a) and the reconstituted 12S RNP particles (b).
DISCUSSION
This study shows that the fragment of the 16S RNA corresponding to the central domain can complex with ribosomal proteins and form a specific RNP. Four ribosomal proteins, S6, S8, S15, and S11, were previously shown to bind exclusively to sites within the central domain of the 16S RNA (19, 20, 22). Proteins S8 and S15 directly and independently bind to the 16S RNA at the early stage of ribosome assembly (23), whereas protein S6, according to the assembly map of the 30S subunit (23), binds only after S15 is bound. The binding of protein S11 also depends on the presence of proteins S15, S6, and S8. This suggests specificity of the RNP formation and indicates that intradomain protein–protein interactions may take place in this complex.
Proteins S6 and S18 bind cooperatively and show very low capability for binding in the absence of each other (22, 23). The homology between protein S18 of E. coli and any of the 30S ribosomal proteins of T. thermophilus is not revealed (9, 10). On the other hand, protein TS20, present in the 12S RNP, has no homolog among the 30S proteins of E. coli. Thus, it is possible to speculate that the T. thermophilus protein TS20 may play the same structural role as the E. coli protein S18.
Quite unexpected was the presence of low amounts of protein S13 bound to the central domain RNP. According to three-dimensional model of the 30S subunit, this protein is localized in the head region (2, 3, 24). Moreover, S13 has been shown to assemble directly with the 3′ domain of the 16S RNA in stoichiometric amounts (6). The lack of evidence for direct interactions between protein S13 and the central domain of the 16S RNA (25) suggests that the appearance of protein S13 in the reconstituted RNP may result from some nonspecific interactions during reconstitution.
The reconstituted RNP has a sedimentation coefficient of 12S. The calculated molecular mass of the complex is 195 kDa and the partial specific volume is 0.61 cm3/g. A comparison of the molecular characteristics of our complex with those of the original 30S subunit (the molecular mass is 850 kDa, the partial specific volume is 0.61 cm3/g, and the sedimentation coefficient is 31 S; unpublished data) indicates that the compactness of the complex is close to that of the 30S subunit. This follows from the dependence S on M2/3, which is valid for compact globular particles with the same partial specific volume (26). Molecular characteristics of the 12S RNP and 30S subunit are in good agreement with this dependence. Thus our complex is highly compact and its overall size and shape are similar to those within the 30S subunit.
The studies of Ofengand and coworkers (5) demonstrated the independent assembly of the 5′ domain RNP, which seems to correspond to the main part of the body of the 30S subunit. Noller and coworkers (6) showed the formation of 3′ domain RNP corresponding to the head of the subunit. Our findings provide evidence for independent assembly of the platform of the 30S subunit. Thus each of three major parts (domains) of the 30S subunit are capable of self-assembly in vitro, relatively independent of each other.
Acknowledgments
We are very grateful to Prof. I. N. Serdyuk for constant interest, stimulating support, and fruitful discussion of the results reported.
ABBREVIATION
- RNP
ribonucleoprotein particle
References
- 1.Noller H F, Woese C R. Science. 1981;212:402–411. doi: 10.1126/science.6163215. [DOI] [PubMed] [Google Scholar]
- 2.Brimacombe R, Atmaja J, Stiege W, Shuler D. J Mol Biol. 1988;199:115–136. doi: 10.1016/0022-2836(88)90383-x. [DOI] [PubMed] [Google Scholar]
- 3.Stern S, Weiser B, Noller H F. J Mol Biol. 1988;204:447–481. doi: 10.1016/0022-2836(88)90588-8. [DOI] [PubMed] [Google Scholar]
- 4.Stern R, Powers T, Cnangchien L M, Noller H F. Science. 1989;44:783–790. doi: 10.1126/science.2658053. [DOI] [PubMed] [Google Scholar]
- 5.Weitzmann C J, Cunningham P R, Nurse K, Ofengand J. FASEB J. 1993;7:177–180. doi: 10.1096/fasebj.7.1.7916699. [DOI] [PubMed] [Google Scholar]
- 6.Samaha R R, O’Brien B, O’Brien T W, Noller F. Proc Natl Acad Sci USA. 1994;91:7884–7888. doi: 10.1073/pnas.91.17.7884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Murzina N V, Vorozheikina D P, Matvienko N I. Nucleic Acids Res. 1988;16:8172. doi: 10.1093/nar/16.16.8172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Murzina N V, Vorozheikina D P, Matvienko N I. Structure and Biosynthesis of Proteins. USSR: Institute of Protein Research; 1988. 2, 25–35. [Google Scholar]
- 9.Tsiboli P, Herfurth E, Choli T. Eur J Biochem. 1994;226:169–177. doi: 10.1111/j.1432-1033.1994.tb20038.x. [DOI] [PubMed] [Google Scholar]
- 10.Eliseikina I A, Agalarov S C, Muranova T L, Markova L F, Kashparov I A, Avliyakulov N K, Garber M B. Biokhimiya. 1995;60:1720–1730. [PubMed] [Google Scholar]
- 11.Sambrook J, Fritsch E F, Maniatis T. Molecular Cloning. A Laboratory Manual. 2nd Ed. Plainview, NY: Cold Spring Harbor Lab. Press; 1989. [Google Scholar]
- 12.Matvienko N I, Pachkunov D M, Kramarov V M. FEBS Lett. 1984;177:23–26. [Google Scholar]
- 13.Melton D A, Krieg P A, Rebagliat M R, Maniatis T, Zinn K, Green M R. Nucleic Acids Res. 1984;12:7035–7056. doi: 10.1093/nar/12.18.7035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Gogia Z V, Yusupov M M, Spirina T N. Mol Biol. 1986;20:519–526. [Google Scholar]
- 15.Kanny J W, Lambert J M, Traut R R. Methods Enzymol. 1979;59:539–550. doi: 10.1016/0076-6879(79)59112-5. [DOI] [PubMed] [Google Scholar]
- 16.Valentine R S, Shapiro B M, Stadtman E R. Biochemistry. 1968;7:2143–2152. doi: 10.1021/bi00846a017. [DOI] [PubMed] [Google Scholar]
- 17.Hardy S J S, Kurland C G, Voynov P, Mora G. Biochemistry. 1969;8:2897–2905. doi: 10.1021/bi00835a031. [DOI] [PubMed] [Google Scholar]
- 18.Sedelnikova S E, Agalarov S C, Garber M B, Yusupov M M. FEBS Lett. 1987;220:227–230. [Google Scholar]
- 19.Svensson P, Changchien L-M, Craven G R, Noller H F. J Mol Biol. 1988;200:301–308. doi: 10.1016/0022-2836(88)90242-2. [DOI] [PubMed] [Google Scholar]
- 20.Stern S, Powers T, Changchien L-M, Noller H F. J Mol Biol. 1988;201:683–695. doi: 10.1016/0022-2836(88)90467-6. [DOI] [PubMed] [Google Scholar]
- 21.Lata K R, Agrawal R K, Penczek P, Grassucci R, Zhu Y, Frank J. J Mol Biol. 1996;262:43–52. doi: 10.1006/jmbi.1996.0497. [DOI] [PubMed] [Google Scholar]
- 22.Gregory R S, Zeller M L, Thurlow D L, Gourse R G, Stark M J R, Dalhberg A E, Zimmermann R A. J Mol Biol. 1984;178:287–302. doi: 10.1016/0022-2836(84)90145-1. [DOI] [PubMed] [Google Scholar]
- 23.Held W A, Ballou B, Mizushima S, Nomura M. J Biol Chem. 1974;249:3103–3111. [PubMed] [Google Scholar]
- 24.Spirin A S, Serdyuk I N, Shpungin J L, Vasiliev V D. Proc Natl Acad Sci USA. 1979;76:4867–4871. doi: 10.1073/pnas.76.10.4867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Powers T, Stern S, Changchien L-M, Noller H F. J Mol Biol. 1988;201:697–716. doi: 10.1016/0022-2836(88)90468-8. [DOI] [PubMed] [Google Scholar]
- 26.Serdyuk I N, Shpungin J L, Zaccai G. J Mol Biol. 1980;137:109–121. doi: 10.1016/0022-2836(80)90160-6. [DOI] [PubMed] [Google Scholar]