

Abstract
The induction of fibroblast apoptosis and their clearance by phagocytes is essential for normal wound healing and prevention of scarring. However, little is known about the clearance of apoptotic fibroblasts and whether apoptotic cells are active participants in the recruitment and activation of phagocytes. In this study, we provide the first evidence that apoptotic fibroblasts actively release increased amounts of thrombospondin (TSP1) to actively recruit macrophages. Expression of TSP1 and its receptor CD36 was increased on the surface of apoptotic fibroblasts. By chemical cross-linking and immunoprecipitation we show that TSP1 and CD36 were directly associated. This was confirmed by confocal microscopy. Blockade of either CD36 or TSP1 on apoptotic fibroblasts inhibited phagocytosis. Blockade of αvβ3 integrins as well as CD36 and TSP1 on macrophages inhibited phagocytosis. In contrast, phosphatidylserine or lectins were not involved. These findings suggest that apoptotic fibroblasts release TSP1 as a signal to recruit macrophages while the up-regulated expression of the CD36/TSP1 complex on their cell surface may form a ligand bridging the fibroblast to a complex consisting of αvβ3/CD36/TSP1 on macrophages. These results establish fundamental mechanisms for the clearance of apoptotic fibroblasts and may provide insights into the processes involved in normal wound repair.
In the lung and other organs, fibroblasts limit the extent of inflammation through the formation of fibrotic tissue. During the subsequent restoration phase, there is a regression of fibrosis by the induction of apoptosis in fibroblasts and their subsequent clearance by macrophages. 1 There is increasing evidence to suggest that impaired clearance of apoptotic cells may contribute to the pathogenesis of disease. 2 In this regard, it is possible that the impaired clearance of apoptotic fibroblasts may perpetuate the fibrotic response. However the processes involved in fibroblast apoptosis are poorly understood and despite the recent focus on the phagocytosis of apoptotic cells, no study to date has addressed the mechanisms by which apoptotic fibroblasts are cleared by phagocytes.
Macrophages have been shown to use a wide repertoire of receptors to phagocytose apoptotic cells. These include lectins; 3 asialoglycoprotein; 4 CD14, 5 a specific phosphatidylserine receptor; 6 and integrins. 7 The precise receptor involved seems to be dependent on the species and activation state of the macrophage as well as the type of apoptotic cell being phagocytosed. 8 In contrast, the ligands expressed on the apoptotic cell that signals phagocyte recognition are poorly understood. Furthermore little is known about factors that are released by apoptotic cells that may induce macrophage chemotaxis and activation.
Thrombospondin (TSP) 1 is a matricellular glycoprotein released by a number of cells including fibroblasts and mediates a variety of processes related to wound repair including the induction of apoptosis 9 and recruitment of macrophages. 10 Indeed, a complex consisting of TSP1, its receptor CD36, and the integrin αvβ3 has been shown to be instrumental in macrophage phagocytosis of apoptotic neutrophils. 11 However, Savill and colleagues 11 postulated that TSP1 was bound to an unknown ligand on the apoptotic neutrophil, acting as a bridge between the apoptotic cell and the CD36/αvβ3 complex on the macrophages.
In this study we demonstrate that TSP1 is released from apoptotic fibroblasts and is a potent chemoattractant for macrophages. We also show that TSP1 is also bound to CD36 on the cell surface of the apoptotic fibroblast and forms a complex recognized by the CD36/TSP1/αvβ3 complex on the macrophage that initiates phagocytosis. In this system, phosphatidyl serine did not seem to be involved in the recognition of phagocytosis of apoptotic fibroblasts.
Materials and Methods
Reagents and Antibodies
Dulbecco’s modified Eagle’s medium (DMEM), RPMI 1640, containing penicillin, gentamicin and amphotericin, Ficoll-Paque, mannose, fucose, galactosamine, glucose, galactose, glucosamine, N-acetylglucosamine, propidium iodide, and Hoescht 33342 were purchased from Sigma Chemical Co. (St. Louis, MO). Annexin V-fluorescein isothiocyanate and terminal dUTP nick-end labeling (TUNEL) were purchased from Roche Diagnostics (Sydney, Australia). DiI(282) a long-chain dialkylaminostyryl dye, was purchased from Molecular Probes (Eugene, OR). FasL was purchased from Calbiochem (Sydney, Australia). Antibodies against TSP1 (clone A4.1) and CD36 (clone 1A7) and 185-1G20 (polyclonal) were purchased from Neomarkers (Freemont, CA). Antibodies against αvβ3, α5β1, and β1 were purchased from Chemicon (Melbourne, Australia). Disuccinimidyl suberate was purchased from Pierce (Rockford, IL). Coverslip chamber wells were obtained from Labtec (NUNC, Roskilder, Denmark). fMLP was purchased from Biomol (Plymouth, PA). Liposomes were prepared by sonication of pure phospholipids as described previously (Sigma). 11 Phospholipids were dissolved in chloroform/methanol and the solvent was evaporated under N2 gas and the lipids were resuspended in phosphate-buffered saline (PBS) by vortexing. The mixtures were then sonicated for 5 minutes (Branson Sonifier 450). The liposomes contained phosphatidylcholine, phosphatidylcholine and phosphatidylserine (molar ratio, 70:30), or phosphatidylcholine and phosphatidylethanolamine (molar ratio, 70:30).
Macrophages
Heparinized peripheral blood was obtained from normal donors after informed written consent. Monocytes were isolated on Ficoll-Paque and macrophages were derived by culture of adherent monocytes were cultured for 5 to 7 days in RPMI 1640 supplemented with 10% fetal calf serum (FCS) and M-CSF as previously described. 12 Positive staining for CD68 staining was used to confirm the presence of macrophages. In this cell-culture system, 96 ± 3% of the cells were macrophages.
Apoptotic Fibroblasts
Primary cultures of fibroblasts were derived from normal human lung as previously described. 13 Fibroblasts were cultured in DMEM supplemented with 10% FCS, penicillin, gentamicin, and amphotericin. For experiments, fibroblasts were quiesced by the addition of serum-free DMEM for 16 hours, then rendered apoptotic by exposure to FasL (50 μg/ml) for 24 hours. Binding of FasL to Fas (CD95) on the cell-surface has been shown to induce apoptosis in multiple cell types. Fibroblasts were defined as apoptotic on the basis of morphological changes such as cell shrinkage and chromatin condensation as well as positive staining with TUNEL and annexin V-fluorescein isothiocyanate. Positive staining with propidium iodide identified necrotic cells. After exposure to FasL for 24 hours, 48 ± 3% of fibroblasts were found to be apoptotic and less than 1% were necrotic. In contrast 1.5 ± 0.3% of fibroblasts were apoptotic and less than 1% were necrotic when cultured in DMEM supplemented with 10% FCS.
Phagocytosis Assay
The phagocytic assay was performed as previously described. 14 Briefly, macrophages were plated on to coverslip chamber wells (105 cells per well) in RPMI 1640. After adherence they were stained with DiI for 20 minutes. Macrophage viability was 98 ± 1% after this process. Apoptotic fibroblasts were suspended in RPMI and placed on the macrophage monolayer at a concentration of 106 cells/well. This interaction was allowed to take place at 37°C for 1 hour. Loosely adherent fibroblasts were removed by washing with cold PBS five times. Fibroblasts were either untreated or incubated with antibodies to CD36, TSP1, and αvβ3 for 1 hour. In separate experiments, macrophages were incubated with antibodies to CD36, TSP1, αvβ3, or cyclic RGD peptides for 1 hour or alternatively incubated with liposomes and amino sugars. For all experiments, 200 cells/high-power field were counted and three high-power fields per section were analyzed (for a total of 600 cells). Fluorescent images were captured on a BioRad MRC 1000 confocal laser-scanning microscope. The phagocytic index was determined as the proportion of macrophages that had ingested TUNEL-positive fibroblasts expressed as a percentage of the total number of macrophages. Only one TUNEL-positive nucleus was observed in each macrophage.
Video Time-Lapse Microscopy
Fibroblasts (104) were plated on a standard microscope slide and incubated with FasL for 24 hours. Two populations of macrophages were used to investigate the dynamics of chemotaxis and phagocytosis. One population was incubated with monoclonal antibodies to CD36 and the other population was stained with DiI. In a separate experiment one population of macrophages was incubated with monoclonal antibodies against TSP1 and the other population was stained with DiI. Macrophages (105) were added to the fibroblasts after 24 hours of incubation with FasL. Phagocytosis of apoptotic cells was visualized by a Nikon Eclipse TE300 microscope (Lab Supply, Australia) (×40) throughout a 12-hour time period at a capture rate of 40 frames/minute. Fluorescent images were simultaneously captured at the beginning of the video time-lapse microscopy and at 6-hour intervals for 24 hours. The images were analyzed by Adobe Premier 5.1 software.
Flow Cytometry
Apoptotic fibroblasts and healthy fibroblasts grown in DMEM supplemented with 10% FCS were suspended in PBS and were dual stained with tetramethyl-rhodamine isothiocyanate-labeled antibodies to TSP1 or CD36 as well as with annexin V-fluorescein isothiocyanate according to the manufacturer’s specifications. For detection of intracellular CD36 staining, fibroblasts were permeabilized by treatment with 0.1% Triton X-100 for 3 minutes before antibody incubation. Labeled cells were analyzed on a FACScalibur (Becton Dickinson, San Jose, CA) and quantified by CellQuest software. Ten to thirty thousand events were collected for analysis.
Chemotaxis Assay
Chemotaxis assays were conducted in modified Boyden chambers (Neuroprobe, Cabin John, MD) as previously described. 15 Briefly, polycarbonate membrane filters (5 μm, Neuroprobe) were used to separate the upper wells from the lower wells. Apoptotic fibroblasts (103/well) were seeded in the lower chambers and macrophages (104/well) were added to the upper wells. Macrophages and apoptotic fibroblasts were incubated with antibodies and RGD peptides as described and the chambers were placed in a humidified incubator at 37°C for 90 minutes. Formyl-methionyl-leucyl-phenylalanine (fMLP) was used as positive control for chemotaxis. Filters were fixed, stained, and washed, and the number of macrophages that had migrated was evaluated by microscopy and direct cell counting.
Chemical Cross-Linking and Immunoprecipitation
Chemical cross-linking of surface proteins using disuccinimidyl suberate was performed on fibroblasts maintained in media containing 10% FCS (healthy) as well as fibroblasts serum-starved for 16 hours and exposed to FasL for a further 1 hour or for 24 hours. Immunoprecipitation of cell-surface proteins were performed as described previously. 16 In brief, cells were lysed in RIPA buffer (Roche Diagnostics) and triturated intermittently for 30 minutes. Insoluble material was removed by centrifugation at 14,000 rpm for 20 minutes at 4°C. The supernatant was precleared by incubation with protein A-agarose (Santa Cruz, San Diego, CA) for 1 hour at 4°C with end-over-end mixing. After centrifugation, supernatants were incubated with a monoclonal antibody to CD36 for 1 hour at 4°C. Immune complexes were precipitated by addition of protein A-agarose for 2 hours. The immune complexes that bound to agarose beads were then washed three times in RIPA buffer, boiled for 5 minutes, fractionated on a 4% stacking/12.5% resolving gel, and transferred to a polyvinylidene difluoride membrane. The membrane was blocked overnight in 5% skim milk powder in TTBS (Tris-buffered saline/0.25% Tween-20) and then incubated with anti-TSP1 antibody (1:500) for 1 hour. After washing, horseradish peroxidase-conjugated anti-mouse IgG was added (1:2000, 1 hour). After washing in TTBS, blots were developed by using enhanced chemiluminescence (ECL, Amersham) and exposed to ECL-Hyperfilm.
Thrombospondin Enzyme-Linked Immunosorbent Assay (ELISA)
Thrombospondin ELISA was performed using the sandwich method as previously described. 17 Each well of a 96-well ELISA plate (Corning, Cambridge, MA) was incubated with 100 μl of 1:1000 rabbit polyclonal anti-human TSP1 IgG overnight at room temperature and blocked with blocking solution (Block Ace; Dainihon, Osaka, Japan). The sample solution (100 μl) was added to each well and incubated for at 37°C for 1 hour, washed twice with washing solution (0.05% Nonidet P-40 in barbiturate-buffered saline). The wells were incubated with 1 μg/ml of mouse monoclonal antibody at 37°C for 1 hour, washed then incubated with 1 μg/ml of biotinylated horse polyclonal anti-mouse IgG (Vector, Burlingame, CA) and 1 μg/ml horseradish peroxidase-avidin complex (Vector). The bound antibody was detected by the addition of o-phenylene-diamine diluted in methanol containing 0.03% hydrogen peroxidase. The absorbance at 490 nm of each well was measured and the TSP1 concentration in each sample was determined from a standard curve prepared with purified human TSP1 at concentrations ranging from 0.39 to 3.5 ng/ml.
Confocal Microscopy
Cells were serum-starved for 16 hours and exposed to FasL (50 μg/ml) for a further 24 hours before being fixed in 1% paraformaldehyde for 30 minutes at room temperature and permeabilized for 3 minutes with 0.2% Triton X-100 in PBS containing 1% bovine serum albumin (BSA). Cells were rinsed in PBS and incubated with monoclonal antibody to TSP1 for 60 minutes at room temperature in PBS/1% BSA. From this stage onwards all incubations and washes were performed in the dark. After several washes in PBS, coverslips were incubated for 60 minutes in PBS containing rabbit anti-mouse IgGs conjugated to Alexa-488 and 1% BSA. Coverslips were rinsed in PBS and then incubated in with a polyclonal antibody to CD36 for 60 minutes at room temperature in PBS/1% BSA. After washes in PBS, cells were incubated with goat anti-rabbit IgGs conjugated to Alexa-546 and 1% BSA. Fibroblasts were then stained with Hoescht 333N2 nuclear stain (0.1 μg/ml) for 45 minutes at room temperature. After washing in PBS, coverslips were mounted with fade-resistant aqueous mounting medium. Co-localization of CD36 and TSP1 was examined on both healthy and apoptotic fibroblasts. Apoptotic fibroblasts were identified by their fragmented nuclei with Hoescht staining. Fluorescent images were obtained using a BioRad MRC1000 confocal laser-scanning microscope using COMOS software. Image processing was performed using Confocal Assistant software and Adobe Photoshop.
Statistical Analysis
Data are expressed as mean ± SE of at least four experiments. Statistical comparisons of mean data were performed using one-way analysis of variance and Student’s t-test with Bonferroni correction performed posthoc to correct for multiple comparisons. A P value <0.05 was regarded as statistically significant.
Results
Apoptotic Fibroblasts Release TSP1
Figure 1 ▶ shows that the induction of apoptosis in fibroblasts induces the release of readily quantifiable levels of TSP1. Fibroblast cultures that were rendered quiescent by serum deprivation for 16 hours produced TSP1. However the induction of apoptosis by exposure to FasL (50 μg/ml) for an additional 24 hours produced a further significant increase of TSP1 from 1200 ± 15 pg/ml to 2100 ± 30 pg/ml (P < 0.01 compared to cells in serum-free media alone).
Figure 1.

TSP1 is secreted by apoptotic fibroblasts. TSP1 was measured by ELISA in the supernatants obtained from fibroblasts cultured in serum-free DMEM and fibroblasts rendered apoptotic by exposure to serum-free media (sfm) for 16 hours and FasL for 24 hours. There was secretion of TSP1 from cells in serum-free media and cells exposed to FasL compared with the levels in serum-free media. There was a significant rise in TSP1 production by fibroblasts exposed to FasL compared to fibroblasts in serum-free media. Results represent mean ± SEM of five experiments. **, P < 0.01 compared to cells in sfm.
TSP1 Released from Apoptotic Cells Is a Chemotactic Agent for Macrophages
Having established that TSP1 was released by apoptotic fibroblasts, we sought to examine its role as a macrophage chemoattractant. Figure 2 ▶ shows that in a modified Boyden chamber assay, macrophage migration was significantly greater toward apoptotic fibroblasts than toward nonapoptotic cells (P < 0.05). Incubation of apoptotic fibroblasts with antibodies against TSP1 significantly inhibited macrophage chemotaxis (P < 0.01). Blockade of macrophage-derived CD36 also diminished their migration in response to signals released from the apoptotic cells, but not in response to fMLP. In contrast, blockade of αvβ3 integrins on the macrophage or CD36 on the apoptotic fibroblasts did not influence the chemotactic response. Similarly, incubation of either cell type with function-blocking antibodies recognizing αvβ5 and αvβ1 integrins was also without effect.
Figure 2.

TSP1 released by apoptotic fibroblasts is a chemoattractant for macrophages. Apoptotic fibroblasts were either untreated or incubated with antibodies against CD36 or TSP1. Macrophages were incubated with antibodies against CD36 or TSP1 or treated with various inhibitors. Incubation of apoptotic fibroblasts with antibodies against TSP1 and incubation of macrophages with antibodies against CD36 inhibited chemotaxis. Data represent mean ± SEM of five experiments. **, P < 0.01 compared to chemotaxis toward apoptotic fibroblasts.
We confirmed the role of TSP1/CD36 in macrophage recognition and phagocytosis of apoptotic fibroblasts using video time-lapse microscopy. In these experiments, healthy macrophages appeared to migrate toward and phagocytose an apoptotic fibroblast, whereas macrophages treated with function-blocking antibodies against CD36 did not appear to actively phagocytose these cells (data not shown).
Surface Expression of CD36 and TSP1 Are Up-Regulated on Apoptotic Fibroblasts
Nonapoptotic fibroblasts appeared to contain an intracellular pool of CD36 as evidenced by the rightward shift in mean fluorescence intensity (Figure 3A) ▶ . Exposure of fibroblasts to FasL (50 μg/ml) for 24 hours induced a significant increase in TSP1 (Figure 3, B and C) ▶ and CD36 (Figure 3, D and E) ▶ mean fluorescence intensity compared to healthy untreated cells. Cell counts of 300 fibroblasts by confocal microscopy or 30,000 fibroblasts by flow cytometry demonstrated that the vast majority of apoptotic fibroblasts were also CD36+ve and TSP1+ve compared to healthy cells (Table 1) ▶ .
Figure 3.

The expression of CD36 and TSP1 is up-regulated on the surface of apoptotic fibroblasts. Histograms of FACS analysis illustrating the presence of an intracellular pool of CD36 in healthy fibroblasts (A) and the up-regulation of TSP1 (B) and CD36 (D) on apoptotic fibroblasts as compared to healthy fibroblasts. Four-quadrant analysis demonstrates a significant number of annexin+ve cells are also TSP1+ve (top right) (C) and CD36+ve (E). Nonapoptotic fibroblasts were cultured in DMEM supplemented with 10% FCS. Apoptotic fibroblasts were quiesced in serum-free DMEM for 12 hours and treated with FasL for 24 hours. Fibroblasts were suspended in 1% BSA/PBS and stained with tetramethyl-rhodamine isothiocyanate-labeled monoclonal antibodies to CD36 or TSP1. Similar results were obtained in each of four experiments.
Table 1.
The Effect of FasL on Expression of CD36 and TSP1 on the Surface of Human Lung Fibroblasts
| Annexin V−ve | Annexin V+ve | |
|---|---|---|
| CD36+ve | 4 ± 0.5 | 78 ± 9* |
| TSP+ve | 6 ± 1.5 | 68 ± 5* |
Cells were exposed to FasL (50 μg/ml) for 24 hours before FACS analysis as described in Materials and Methods. Values represent mean ± SE of at least four independent experiments.
*Denotes statistically different to Annexin V−ve cells, P < 0.01.
TSP1 Associates with CD36 on the Surface of Apoptotic Fibroblasts
Having demonstrated that expression of both TSP1 and CD36 were up-regulated on apoptotic fibroblasts, we sought to determine whether TSP1 was directly associated with CD36. Figure 4A ▶ shows a single band corresponding to the molecular weight of monomeric TSP1 (450 kd) in cell lysates immunoprecipitated with antibodies against CD36. After exposure of cells to FasL for 24 hours, an extra band corresponding to the expected size of TSP1 and CD36 (88 kd) is observed. In contrast, no extra band was detectable in cell lysates of healthy cells at 24 hours or in fibroblasts exposed to FasL for 1 hour. Visual confirmation of this association was provided by confocal microscopy (Figure 4; B, C, and D) ▶ . Apoptotic fibroblasts (nuclear fragmentation revealed by Hoechst 33342) that were double stained with antibodies to CD36 (green) and TSP1 (red), show distinct areas of co-localization (yellow). In contrast, healthy fibroblasts (intact nuclei with Hoechst staining) showed no co-localization.
Figure 4.

TSP1 associates with CD36 on apoptotic fibroblasts. A: Surface receptors on healthy fibroblasts and apoptotic fibroblasts were chemically cross-linked and protein was immunoprecipitated with antibodies against CD36, lysed, and the resultant protein complexes were then probed with antibodies against TSP1. CD36 associated with TSP1 on the apoptotic fibroblasts but not on healthy fibroblasts or on fibroblasts exposed to FasL for 1 hour. Similar results were obtained in each of three experiments. Double staining with CD36 (B; Alexa 546, red) or TSP1 (C; Alexa 488, green) reveals significant areas of co-localization (yellow staining in D) (↑). Fibroblasts that positively stained for CD36 and TSP1 were confirmed to be apoptotic by nuclear fragmentation on Hoescht 33342 (blue) staining. Healthy fibroblasts with intact nuclei do not demonstrate co-localization of CD36 and TSP1 ( ↑) Scale bar, 50 μm.
Macrophages Use the αvβ3 Integrin, CD36, and TSP1 for the Recognition and Phagocytosis of Apoptotic Fibroblasts
Incubation of macrophages with function-blocking antibodies against αvβ3 reduced the percentage of macrophages that phagocytosed apoptotic fibroblasts from 90 ± 3% to 11 ± 1% (P < 0.01, Figure 5A ▶ ). This appeared specific for αvβ3 because the specific cyclic RGD peptide (GPenGRGDSPCA) also dose dependently inhibited phagocytosis, whereas the control peptide RGES (Figure 5A) ▶ and isotypically matched antibodies against αvβ5 and αvβ1 integrins did not influence phagocytosis. Incubation of macrophages with antibodies against CD36 and TSP1 also abrogated phagocytosis of apoptotic fibroblasts to the same level as inhibition by αvβ3 antibodies (Figure 5A) ▶ . Incubation of apoptotic fibroblasts with function-blocking antibodies to CD36 and TSP1 also inhibited phagocytosis (Figure 5B) ▶ .
Figure 5.

Macrophages use the αvβ3/CD36/TSP1 complex to phagocytose apoptotic fibroblasts. A: Phagocytosis of apoptotic fibroblasts by human monocyte-derived macrophages was inhibited by incubation of macrophages with CD36, αvβ3, and TSP1 antibodies. Incubation of macrophages with the cyclic RGD peptide GPenGRGDSPCA also dose dependently inhibited phagocytosis. B: Incubation of apoptotic fibroblasts with antibodies against CD36 and TSP1 inhibited phagocytosis. Incubation with a function-blocking antibody against αvβ5 was without effect. *, P < 0.05; **, P < 0.01, compared to phagocytosis of apoptotic fibroblasts alone.
Blockade of Phosphatidylserine or Amino Sugars Does Not Influence Phagocytosis of Apoptotic Fibroblasts
In contrast to inhibition of αvβ3, CD36, and TSP1, incubation of macrophages with liposomes containing phosphatidylserine or control liposomes containing phosphatidylcholine or a mixture of the two lipids had no effect on phagocytosis of apoptotic fibroblasts (Figure 6A) ▶ . Similarly, incubation of macrophages with a variety of specific amino sugars did not influence the ability of macrophages to phagocytose apoptotic fibroblasts (Figure 6B) ▶ .
Figure 6.
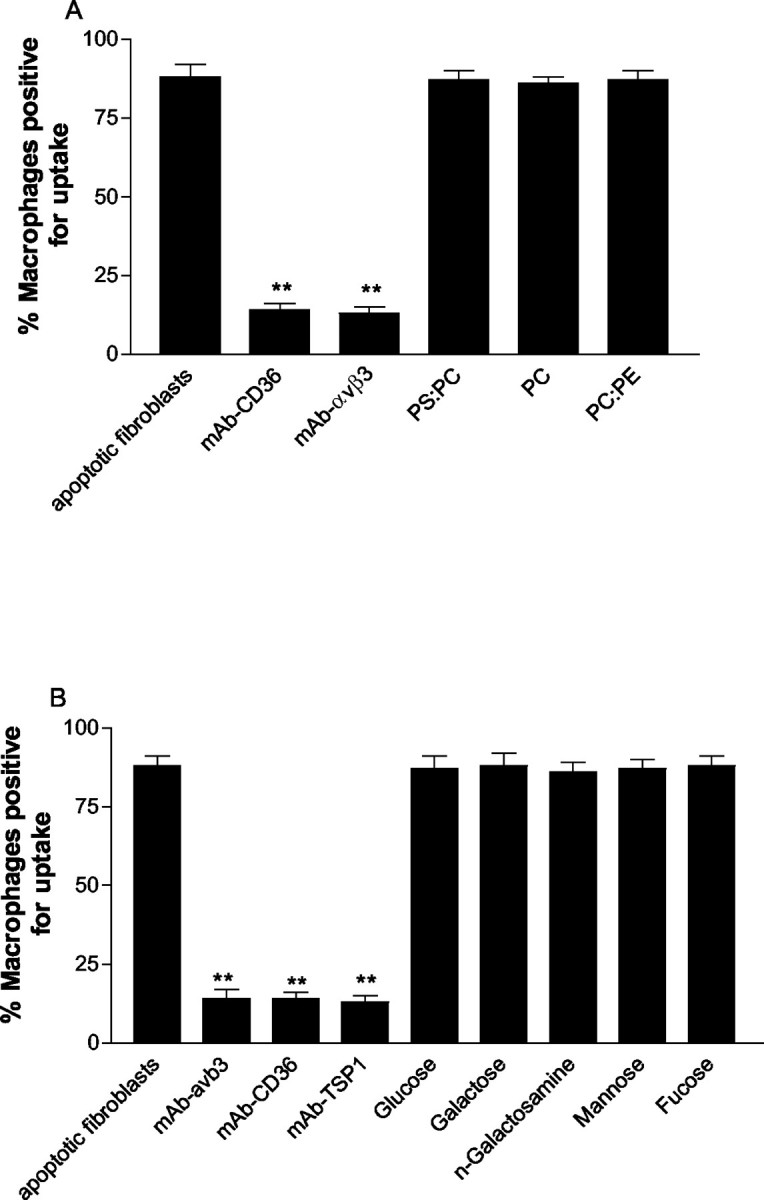
A: Incubation of macrophages with liposomes containing phosphatidylserine (PS) (PC:PS, 70:30), phosphatidylcholine (PC), or phosphatidylethanolamine (PE) did not influence their ability to phagocytose apoptotic fibroblasts. B: Incubation with amino sugars also had no effect on the ability of macrophages to phagocytose apoptotic fibroblasts. Data represent mean ± SEM of five experiments. **, P < 0.01 compared to phagocytosis of apoptotic fibroblasts alone.
Discussion
This study establishes fundamental principles in the clearance of apoptotic fibroblasts by macrophages. Firstly, apoptotic fibroblasts secrete TSP1, which acts as a chemoattractant for macrophages. Secondly we demonstrate for the first time that the CD36 and TSP1 directly associate on the surface of apoptotic fibroblasts and this complex is a ligand for the αvβ3/CD36/TSP1-receptor complex on macrophages. This is particularly significant because the αvβ3/CD36/TSP1 receptor complex on macrophages plays a crucial role in the clearance of apoptotic cells but no ligand on an apoptotic cell has been previously described for this receptor. Finally, we show that receptors for phosphatidylserine or amino sugars are not involved in macrophage phagocytosis of apoptotic fibroblasts.
The findings of this study have implications for the understanding of normal wound healing. During inflammation, growth factors and cytokines stimulate fibroblasts to form scar tissue. 18 The restoration of normal tissue homeostasis by the replacement of scar tissue with healthy tissue requires the induction of apoptosis and the subsequent clearance of apoptotic fibroblasts. Despite the obvious importance of this process to normal tissue homeostasis and the prevention of fibrosis, the mechanisms involved in the recruitment of macrophages and their recognition of apoptotic fibroblasts has not been previously addressed.
We examined TSP1 for several reasons: TSP1 is present at an early time point at sites of inflammation 19 and plays many roles during wound repair such as inhibition of angiogenesis, 20 activation of transforming growth factor-β, 21 as well as several proapoptotic functions. 8,12 TSP1 is also a chemotactic agent for a variety of cells including macrophages. 9 Previous studies have also shown that fibroblasts release small amounts of TSP1. 22,23 We confirmed and extended these findings by demonstrating that TSP1 is released by fibroblasts cultured in serum-free media but release substantially more TSP1 after exposure to FasL. Using a blind-well chemotaxis assay we have shown TSP1 derived from apoptotic fibroblasts is a potent chemoattractant for macrophages. Furthermore, our data show that CD36 is the macrophage receptor responsible for chemotaxis, in response to TSP1. Other candidate receptors for TSP1 include αvβ1, αvβ3, and αvβ5 integrins. 24 Using function-blocking antibodies, we have shown that TSP1-stimulated chemotaxis of macrophages does not involve these particular integrins. The role of CD36 in this context is at variance with Mansfied and Suchard 10 who suggested that that this receptor was not involved in macrophage chemotaxis. The reason for this discrepancy is unclear, but the use of different antibodies may have contributed. In addition we evaluated macrophage chemotaxis toward apoptotic cells whereas in the study of Mansfield and Suchard 10 macrophage chemotaxis was toward intact and fragmented TSP1. This would exclude other cellular components or accessory molecules present on apoptotic fibroblasts that may serve to enhance the chemoattractant effect of TSP1.
Although a growing number of distinct macrophage receptors involved in the recognition and phagocytosis of apoptotic cells has been identified, the corresponding ligand(s) on apoptotic cells remain poorly characterized. We provide several lines of evidence to show that CD36 and TSP1 associate on the surface of apoptotic fibroblasts and act as a recognition signal for macrophages. Firstly, we used flow cytometry to show that surface expression of both CD36 and TSP1 are up-regulated on apoptotic fibroblasts when compared to healthy cells. CD36 expression has been described on a variety of cell types including macrophages, 25 endothelial cells, 26 and platelets. 27 In contrast, little is known about the regulation of CD36 expression on fibroblasts, although Stomski and colleagues 28 described an intracellular pool of CD36 in embryonic human fibroblasts that became expressed on the cell surface after exposure to TSP1. Our data showing that nonapoptotic fibroblasts appear to express intracellular CD36 support this earlier finding. Alternatively, up-regulation of surface CD36 expression may also be a consequence of changes in cell membrane phospholipid symmetry that occurs during apoptosis. 29
Secondly, we used chemical cross-linking and immunoprecipitation as well as confocal microscopy to demonstrate that CD36 and TSP1 co-localize and directly associate on the surface of apoptotic, but not healthy, fibroblasts. Thirdly, we showed that blockade of either CD36 or TSP1 on the apoptotic fibroblast inhibits phagocytosis. Taken together, these findings suggest that the CD36/TSP1 complex on apoptotic fibroblasts may constitute a recognition ligand and docking site for macrophages. We speculate that TSP1 forms a bridge between CD36 on the apoptotic fibroblast and αvβ3/CD36 on the macrophage. Indeed, TSP1 has been implicated in adhesion between platelets and monocyte by forming molecular bridges with CD36 and other integrin receptors. 18 Lawler and colleagues 30 demonstrated patchy fibroblastic foci in the lungs of TSP1-deficient mice and suggested that might in part be because of the impaired phagocytosis of apoptotic cells because the TSP1-bridging molecule between the apoptotic cell and phagocyte is absent. Thus it is feasible that the absence of TSP1 may adversely affect phagocyte recognition and subsequent engulfment of the apoptotic cell.
The interaction between the receptor on the macrophage and the ligand on the apoptotic cell during phagocytosis is poorly understood, but is likely to be cell type- and species-dependent. We have demonstrated that unstimulated human monocyte-derived macrophages use a complex consisting of CD36/TSP1/αvβ3 to phagocytose apoptotic fibroblasts. This is in agreement with previous studies that have shown that macrophages also use this complex to phagocytose apoptotic neutrophils 31 and eosinophils. 32 To determine whether alternative pathways or receptors participated in the phagocytosis of apoptotic fibroblasts, a battery of inhibitors and blocking antibodies were used in phagocytic assays. Blockade of lectins using amino sugars or phosphatidylserine with liposomes did not influence macrophage phagocytosis of apoptotic fibroblasts. These findings contrast with previous studies that have demonstrated that the phosphatidylserine receptor on peritoneal-derived mouse macrophages 33 and activated human bone marrow-derived macrophages 34 is instrumental in the phagocytosis of apoptotic cells. Similarly, although the mannose/fucose lectin on fibroblasts has been implicated in phagocytosis of apoptotic neutrophils, 35 blockade of this lectin on macrophages did not influence engulfment of apoptotic fibroblasts. The reasons for these differences are unclear, although the source of macrophages may have contributed.
Both tissue localization and functional studies suggest that TSP1 is an important mediator in wound healing and inflammation. Our results establish that apoptotic fibroblasts actively secrete TSP1 that attracts macrophages and use surface-bound CD36/TSP1 as a recognition signal for macrophages to dock via the counter-receptor complex consisting of αvβ3/CD36/TSP1 and commence phagocytosis. In contrast, lectins or phosphatidylserine on the surface of apoptotic fibroblasts were not involved. Taken together our results provide greater insight into the mechanisms involved in the resolution of normal wound healing.
Acknowledgments
We thank Mr. Guy Ben-Ary for his technical assistance with the video time-lapse microscopy and Dr. Delia Nelson and Dr. Andrew McWilliam for their scientific input and comments. The authors would like to acknowledge the Lotteries Commission of Western Australia for their support of the Biomedical Confocal Research Centre.
Footnotes
Address reprint requests to Dr. Darryl Knight, Asthma and Allergy Research Institute, Ground Floor, “E” Block, Sir Charles GairdnerHospital, Verdun St., Nedlands, Western Australia, 6009. E-mail: dknight@cyllene.uwa.edu.au.
Supported by grant 110206 from the National Health and Medical Research Council (NHMRC) and a South African Pulmonology Society (SAPS) 3M Respiratory Fellowship (to Y. M.).
References
- 1.Lorena D, Uchio K, Costa AM, Desmouliere A: Normal scarring: importance of myofibroblasts. Wound Repair Regen 2002, 10:86-92 [DOI] [PubMed] [Google Scholar]
- 2.Ogasawara JR, Watanabe-Fukunaga M, Adachi A, Matsuzawa T, Kasugai Y, Kitamura N, Itoh T, Suda S, Nagata S: Lethal effect of the anti-Fas antibody in mice. Nature 1993, 364:806-809 [DOI] [PubMed] [Google Scholar]
- 3.Pradhan D, Krahling S, Williamson P, Schlegel R: Multiple systems for recognition of apoptotic lymphocytes by macrophages. Mol Biol Cell 1997, 8:767-778 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Dini L, Autuori F, Lentini A, Oliverio S, Piacentini M: The clearance of apoptotic cells in the liver is mediated by the asialoglycoprotein receptor. FEBS Lett 1992, 296:174-178 [DOI] [PubMed] [Google Scholar]
- 5.Schlegel R, Krahling S, Callahan MK, Williamson P: CD14 is a component of multiple recognition systems used by macrophages to phagocytose apoptotic lymphocytes. Cell Death Differ 1999, 6:583-592 [DOI] [PubMed] [Google Scholar]
- 6.Fadok VA, Bratton DL, Rose DM, Pearson A, Ezekewitz RA, Henson PM: A receptor for phosphatidylserine-specific clearance of apoptotic cells. Nature 2000, 405:85-90 [DOI] [PubMed] [Google Scholar]
- 7.Erwig LP, Gordon S, Walsh GM, Rees AJ: Previous uptake of apoptotic neutrophils or ligation of integrin receptors downmodulates the ability of macrophages to ingest apoptotic neutrophils. Blood 1999, 93:1406-1412 [PubMed] [Google Scholar]
- 8.Reed JC: Mechanisms of apoptosis. Am J Pathol 2000, 157:1415-1430 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Jimenez B, Volpert OV, Crawford SE, Febbraio ME, Silverstein RL, Bouck N: Signals leading to apoptosis-dependent inhibition of neovascularization by thrombospondin-1. Nat Med 2000, 6:41-48 [DOI] [PubMed] [Google Scholar]
- 10.Mansfield PJ, Suchard SJ: Thrombospondin promotes chemotaxis and haptotaxis of human peripheral blood monocytes. J Immunol 1994, 153:4219-4229 [PubMed] [Google Scholar]
- 11.Savill J, Hogg N, Ren Y, Haslett C: Thrombospondin cooperates with CD36 and the vitronectin receptor in macrophage recognition of neutrophils undergoing apoptosis. J Clin Invest 1992, 90:1513-1522 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Gantner F, Kupferschmidt R, Schudt C, Wendel A, Hatzelmann A: In vitro differentiation of human monocytes to macrophages: change of PDE profile and its relationship to suppression of tumour necrosis factor-alpha release by PDE inhibitors. Br J Pharmacol 1997, 121:221-231 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Keerthisingam CB, Jenkins RG, Harrison NK, Hernandez-Rodriguez NA, Booth H, Laurent GJ, Hart SL, Foster ML, McAnulty RJ: Cyclooxygenase-2 deficiency results in a loss of the anti-proliferative response to transforming growth factor-beta in human fibrotic lung fibroblasts and promotes bleomycin-induced pulmonary fibrosis in mice. Am J Pathol 2001, 158:1411-1422 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Fadok VA, Savill JA, Haslett C, Bratton DL, Doherty DE, Campbell PA, Henson PM: Different populations of macrophages use either the vitronectin receptor or the phosphatidylserine receptor to recognize and remove apoptotic cells. J Immunol 1992, 149:4029-4035 [PubMed] [Google Scholar]
- 15.Mansfield PJ, Boxer LA, Suchard SJ: Thrombospondin stimulates motility of human neutrophils. J Cell Biol 1990, 111:3077-3086 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Scaffidi AK, Moodley YP, Weichselbaum M, Thompson PJ, Knight DA: Regulation of human lung fibroblast phenotype and function by vitronectin and vitronectin integrins. J Cell Sci 2001, 114:3507-3516 [DOI] [PubMed] [Google Scholar]
- 17.Miyajima-Uchida H, Hayashi H, Beppu R, Kuroki M, Fukami M, Arakawa F, Tomita Y, Oshima K: Production and accumulation of thrombospondin-1 in human retinal pigment epithelial cells. Invest Ophthalmol Vis Sci 2000, 41:561-567 [PubMed] [Google Scholar]
- 18.Mutsaers SE, Bishop JE, McGrouther G, Laurent GJ: Mechanisms of tissue repair: from wound healing to fibrosis. Int J Biochem Cell Biol 1997, 29:5-17 [DOI] [PubMed] [Google Scholar]
- 19.DiPietro LA, Nissen NN, Gamelli RL, Koch AE, Pyle JM, Polverini PJ: Thrombospondin 1 synthesis and function in wound repair. Am J Pathol 1996, 148:1851-1860 [PMC free article] [PubMed] [Google Scholar]
- 20.Iruela-Arispe ML, Lombardo M, Krutzsch HC, Lawler J, Roberts DD: Inhibition of angiogenesis by thrombospondin-1 is mediated by 2 independent regions within the type 1 repeats. Circulation 1999, 100:1423-1431 [DOI] [PubMed] [Google Scholar]
- 21.Crawford SE, Stellmach V, Murphy-Ullrich JE, Ribeiro SM, Lawler J, Hynes RO, Boivin GP, Bouck N: Thrombospondin-1 is a major activator of TGF-beta1 in vivo. Cell 1998, 93:1159-1170 [DOI] [PubMed] [Google Scholar]
- 22.Jaffe EA, Ruggiero JT, Leung LK, Doyle MJ, McKeown-Longo PJ, Mosher DF: Cultured human fibroblasts synthesize and secrete thrombospondin and incorporate it into extracellular matrix. Proc Natl Acad Sci USA 1983, 80:998-1002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Mumby SM, Abbott-Brown D, Raugi GJ, Bornstein P: Regulation of thrombospondin secretion by cells in culture. J Cell Physiol 1984, 120:280-288 [DOI] [PubMed] [Google Scholar]
- 24.Chandrasekaran L, He CZ, Al-Barazi H, Krutzsch HC, Iruela-Arispe ML, Roberts DD: Cell contact-dependent activation of alpha3beta1 integrin modulates endothelial cell responses to thrombospondin-1. Mol Biol Cell 2000, 11:2885-2900 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Huh HY, Lo SK, Yesner LM, Silverstein RL: CD36 induction on human monocytes upon adhesion to tumor necrosis factor-activated endothelial cells. J Biol Chem 1995, 270:6267-6271 [DOI] [PubMed] [Google Scholar]
- 26.Dawson DW, Pearce SF, Zhong R, Silverstein RL, Frazier WA, Bouck NP: CD36 mediates the in vitro inhibitory effects of thrombospondin-1 on endothelial cells. J Cell Biol 1997, 138:707-717 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Swerlick RA, Brown EJ, Xu Y, Lee KH, Manos S, Lawley TJ: Expression and modulation of the vitronectin receptor on human dermal microvascular endothelial cells. J Invest Dermatol 1992, 99:715-722 [DOI] [PubMed] [Google Scholar]
- 28.Stomski FC, Gani JS, Bates RC, Burns RF: Adhesion to thrombospondin by human embryonic fibroblasts is mediated by multiple receptors and includes a role for glycoprotein 88 (CD36). Exp Cell Res 1992, 198:85-92 [DOI] [PubMed] [Google Scholar]
- 29.Fadok VA, de Cathelineau A, Daleke DL, Henson PM, Bratton DL: Loss of phospholipid asymmetry and surface exposure of phosphatidylserine is required for phagocytosis of apoptotic cells by macrophages and fibroblasts. J Biol Chem 2001, 276:1071-1077 [DOI] [PubMed] [Google Scholar]
- 30.Lawler J, Sunday M, Thibert V, Duquette M, George EL, Rayburn H, Hynes RO: Thrombospondin-1 is required for normal murine pulmonary homeostasis and its absence causes pneumonia. J Clin Invest 1998, 101:982-992 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Savill J, Fadok V, Henson P, Haslett C: Phagocyte recognition of cells undergoing apoptosis. Immunol Today 1993, 14:131-136 [DOI] [PubMed] [Google Scholar]
- 32.Stern M, Savill J, Haslett C: Human monocyte-derived macrophage phagocytosis of senescent eosinophils undergoing apoptosis. Mediation by alpha v beta 3/CD36/thrombospondin recognition mechanism and lack of phlogistic response. Am J Pathol 1996, 149:911-921 [PMC free article] [PubMed] [Google Scholar]
- 33.Fadok VA, Voelker DR, Campbell PA, Cohen JJ, Bratton DL, Henson PM: Exposure of phosphatidylserine on the surface of apoptotic lymphocytes triggers specific recognition and removal by macrophages. J Immunol 1992, 148:2207-2216 [PubMed] [Google Scholar]
- 34.Fadok VA, Warner ML, Bratton DL, Henson PM: CD36 is required for phagocytosis of apoptotic cells by human macrophages that use either a phosphatidylserine receptor or the vitronectin receptor (alpha v beta 3). J Immunol 1998, 161:6250-6257 [PubMed] [Google Scholar]
- 35.Hall SE, Savill JS, Henson PM, Haslett C: Apoptotic neutrophils are phagocytosed by fibroblasts with participation of the fibroblast vitronectin receptor and involvement of a mannose/fucose-specific lectin. J Immunol 1994, 153:3218-3227 [PubMed] [Google Scholar]