

Abstract
Coupling of spine and hip joints during full body reaching tasks was investigated in sixteen participants (8 male & 8 female) who performed reaching tasks at comfortable and fast-paced movement speeds to three targets located in a para-sagittal plane. The participants paused at target contact for 500 ms and then returned to an upright posture. Three dimensional joint motions of the spine and hip were recorded using an electromagnetic tracking device. We found an effect of movement phase (i.e. reach and return) on the onset timing of the spine and hip joints. For most target locations and movement speeds, spine motion onset preceded hip motion onset during the reaching phase of the movement task. In the reach phase, when averaged across all movement conditions, spine joint motion preceded hip joint motion by an average of 48.9 ms. In contrast, in the return phase, hip joint motion preceded spine joint motion by an average of 63.0 ms. Additionally, when subjects were instructed to use either a knee flexion or knee extension strategy to perform the reaching tasks there was no effect of movement strategy on timing of the spine and hip. There was also no effect of target height on the spine hip ratio, but as movement speed increased, the spine/hip ratio decreased for all target locations due primarily to an increase in hip joint excursion. The findings indicate clear differences in onset timing of the spine and hip joints during reaching tasks that necessitate some forward bending of the trunk and that onset timing is reversed for the return to an upright posture.
Keywords: Kinematics Spine Hip Timing Coordination
1. Introduction
Coupling of the spine and hip joint, or spine-hip ratio, has been described as an important aspect of clinical examination of the spine for years (Cailliet, 1981). In a clinical examination the relative excursions of the spine and hip, as well as the timing of each joint is assessed while the patient bends forward as far as possible with the knees extended and then returns to an upright posture (i.e. forward bend test). Cailliet (1981) proposed that in healthy individuals the lumbar spine and hip joints work in unison, with the hips primarily responsible for forward flexion after 45° of trunk flexion. To return to an upright posture, the process is reversed with the motion initiated by extension of the hips and then followed by simultaneous rotation of the spine and hip joints. With respect to patient populations, Sahramann (2002) claims that postural impairments of the lumbar spine result in a change in the timing and relative contribution of the spine and hips during the forward bend test. However, while there is some evidence of a change in the relative contribution of the spine and hip in patients with low back pain (Esola et al., 1996; Paquet et al., 1994; Porter & Wilkinson, 1997), the findings are not consistent. There is also little empirical evidence to support the notion that low back pain influences timing of the spine and hip for forward bending (Wong & Lee, 2004). Furthermore, it is not known if timing and coupling of the spine and hip observed during a very constrained task such as the forward bend test occurs in more unconstrained movement tasks. Empirical data on the timing and coupling of the spine and hip in a healthy population performing less constrained movement tasks will provide important information regarding coordination of the trunk.
Timing of the spine and hip joint motions in a cyclic flexion extension task was assessed in healthy participants (Lee and Wong, 2002) and in participants with chronic low back pain (Wong and Lee, 2004). In both these studies, the authors used cross-correlation analyses to evaluate onset timing of the spine and hip joints and reported no timing differences in either healthy subjects or those with chronic low back pain. However, there are two potential limitations to these studies. First, cross-correlation analyses may not be particularly sensitive to timing differences in co-varying waveforms, and second, joint coupling is effected by whether a movement task is discrete or cyclic (Meulenbroek et al., 1993; Schillings et al., 1996). Therefore, the timing of the spine and hip needs to be assessed explicitly in subjects performing discrete movement tasks.
While the timing of the onset of spine and hip motion was not explicitly measured, Granata and Sanford (2000) reported that spine and hip motion occurs simultaneously in loaded flexion and extension tasks. In contrast, Nelson et al (1995) reported that spine and hip motion occur simultaneously in the forward bend portion of a trunk flexion task, but that hip motion precedes spine motion in the return to an upright posture. Unfortunately, these authors also did not explicitly measure onset timing and based their conclusions on qualitative assessment of the angle-angle plots. In an early investigation of spine hip coordination, Davis et al. (1965) reported differences in onset timing in the spine and hip for lifting tasks and determined that timing differences were influenced by the lifting strategy used to complete the task (i.e. bent knee versus straight knee). This suggests that joint onset timing is not tightly controlled in coordinated movement of the trunk. Thus, while the issue of onset timing of the spine and hip joints is considered by some to be important clinically (Sahrmann, 2002), only a few investigations have explicitly measured these differences and the evidence from these investigations is conflicting.
Most investigations of the coupling of the spine and hip have focused on the relative excursions of the spine and hip using the highly constrained forward bend test (Esola et al., 1996; Lariviere et al., 2000; Lee & Wong, 2002; McClure et al., 1997; Paquet et al., 1994; Porter & Wilkinson, 1997; Wong & Lee, 2004). While others have examined the spine hip ratio with weighted and un-weighted extension tasks performed from a flexed posture(Granata & Sanford, 2000; McClure et al., 1997; Nelson et al., 1995). In those studies that examined the coupling of the spine and hip during un-weighted forward bending, there is a consistent finding of a greater contribution from the spine than from the hips in the early stages of movement, while the hips contributed more than the spine toward the end of the movement task (Esola et al., 1996; Lee & Wong, 2002; Paquet et al., 1994; Porter & Wilkinson, 1997; Wong & Lee, 2004). The data from these studies indicate clear differences in how motion is apportioned to the spine and hips during the movement task and suggest that there are likely differences in the onset timing of these joints. However, the movement tasks used in these studies assessed end range motion, and it is not known if coupling of the spine and hip in these tasks reflects an underlying coordination strategy or simply indicates relative flexibility of these two joints.
Less constrained movement tasks that can be performed using different movement strategies may provide greater insight into how the central nervous systems plans and executes complex multi-joint tasks (Thomas et al., 1998; Thomas et al., 2005). Thomas et al 1998 examined the relationship between the spine and hip in unconstrained full body reaching tasks and found that the spine-hip ratio was invariant to target location, but was significantly different for the women (spine hip ratio of .20) and men (spine hip ratio of 1.20). While invariance of the spine hip ratio for reaches to two target locations suggest a relatively tight coupling of these joints, Thomas et al (1998) did not explicitly address timing of spine and hip joint motions, and the movement task was restricted to movement to the target. Other studies from this group indicate that in full body reaching tasks, joint excursions are influenced by movement speed (Thomas et al., 2003; Thomas et al., 2005) which will obviously influence the spine hip ratios. Thus, spine hip coordination should be assessed during both comfortable and fast paced movements as well as assessing the return to an upright posture.
In sum, while clinicians have made assumptions about normal and impaired timing of the spine and hip for forward bending and the return from flexion, there has been little empirical data provided to support these assumptions. Furthermore it is not known if joint onset timing and coupling of the spine and hip is simply a function of passive compliance of the joints or if it is influenced by changes in task constraints. Thus, the purpose of this investigation was to examine 1) onset timing of the spine joints in both the movement to the target and the return tasks, 2) determine if movement strategy influences onset timing of the spine and hip (i.e. knee flexion-ankle dorsiflexion or knee extension-ankle plantarflexion), 3) determine the effect of movement speed and target location on the apportionment of motion to the spine and hip joints across the movement task, and 4) determine if gender influences coordination of the spine and hip joints in these movement tasks.
2. Methods
Sixteen normal healthy individuals, eight men (mean age = 25.3 years, range = 21-33 years) and eight women (mean age = 23.6 years, range = 21-32 years) participated in this experiment. All participants signed informed consent and the institutional review board of Ohio University approved this study.
2.1. Procedure
The following anthropometric measures were obtained from the subjects: height, weight, trunk length (greater trochanter to acromion process), pelvic length (greater trochanter to umbilicus), hip height (greater trochanter to floor), thigh length (greater trochanter to lateral joint line of knee), shank length (lateral joint line of knee to lateral maleolus), foot length (medial maleolus to great toe), humerus length (acromion process to lateral epichondyle), and forearm length (lateral epichondyle to radial styloid).
2.2. Reaching Tasks
Subjects performed reaching tasks to three targets located in a para-sagittal plane. The target locations were determined for each subject based on their trunk and pelvic length, arm length (humerus + forearm), and hip height. Specifically, the subject could, in theory, reach the high target by flexing the hips 40° with the shoulder flexed 90° and the elbow fully extended (Fig 1). The middle and low targets could be reached by flexing the hips 60° and 80° respectively.
Figure 1.
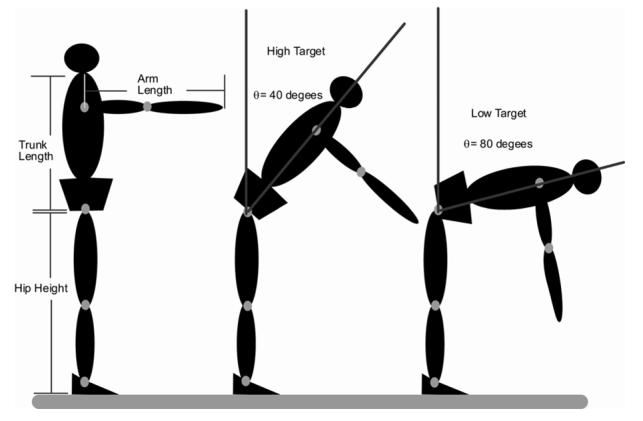
A diagrammatic representation of how target locations were normalized to each subject’s anthropomorphic characteristics. Target locations were determined for each subject based on their hip height, trunk length, and arm length. The high target was located such that the subject could, in theory, reach the target by flexing the hips 40° with shoulder flexed to 90° and the elbow extended. N.B. Target locations were determined mathematically and subjects were not actually placed in the positions illustrated.
The “go” signal was a green light emitting diode LED embedded in the target (7 × 5 cm metal plate) and the return signal was an orange LED embedded in the target. Subjects were instructed to wait for the “go” signal touch the target with the right index finger, and wait until the return signal (initiated 500 ms after contact) to return to an upright posture.
Prior to the start of a reaching task, subjects stood in an upright posture with their right hand on a flexible, carbon electrode strapped to their anterior proximal thigh, which was connected to a contact-detector circuit. When subjects received the “go” signal, and began the reaching task, contact with the thigh electrode was broken and a timer started. When the subject made contact with the target the timer stopped. Movement times determined by the timer were used to train the subjects for the fast paced movement trials.
Subjects were asked to perform three reaches to each target at a “comfortable pace”. After performing 3 movement trials to each target location at a comfortable pace, subjects then performed 3 movement trials to each target location at twice their comfortable pace. Prior to recording the fast-paced reaching trials, subjects were given practice trials in which the movement time was determined from the contact detector circuit. Subjects were given the instruction to “go faster” or “slow down” until they were able to reach the target in movement times that were approximately half that of the comfortable-paced trials.
After completing the reaching tasks at the fast-paced movement speed, subjects then performed movement trials to the middle and low targets with explicit instructions in the body geometry to be used. Specifically, they were first instructed to bend their knees to reach the target locations. After three trials to each target using a knee flexion movement pattern, they were instructed to reach for the targets while keeping their knees extended. Finally, subjects performed two movement trials of a classic forward bend test. In this test, the subject is asked to bend forward as far as possible while keeping their knees extended.
2.3. Data Collection
Movements of the trunk and limb segments were recorded using a magnetic based kinematic system that can track the three-dimensional coordinates of sensors with a spatial resolution of .03 inch (Ascension™). Sensors were attached by Velcro® straps to the limb segments (at the midpoint between the joints) of the right and left shank, thigh, arm, and forearm, as well as the thoracic vertebra (T1) and sacrum. Kinematic data were collected at 100 Hz for a total of 5 seconds, and data collection began 500 ms prior to the “go” signal.
2.4. Data Analysis
Motion Monitor software (Innovation Sports, Chicago, IL) was used to derive time series joint angle data of the lumbar spine and hip joints using an Euler angle sequence. Lumbar spine motion was defined by the change in orientation of the sensor on T-1 relative to the sensor on the sacrum. Hip motion was defined as the change in orientation of the sensor on the sacrum relative to the sensor on the right thigh. The Euler angle sequence extracted the flexion angles, followed by rotation angles, and then lateral flexion angles. Since the motions used to perform these movements were primarily in the sagittal plane, we restricted our analyses to spine and hip flexion angles. The orientation of the reference frames were such that the joint angles were defined as positive in the counterclockwise direction as seen from the subject’s right side, with the vertical direction corresponding to zero. The time series joint angle data were first filtered using a 4th order zero lag butterworth filter with cutoff frequency of 6 Hz. Joint angular velocity of the spine and hip joints was then calculated from the time series joint angle data using custom algorithms developed in Matlab™. Based on these velocity data, the movement task was broken into two phases: 1) movement from an upright posture to target contact, and 2) movement from target contact to an upright posture.
Calculating Spine Hip Latencies
For each phase of the movement task, we determined initiation of joint motion, time of peak velocity, and termination of joint motion. Initiation of joint motion was defined as the time at which joint angular velocity exceeded 5% of peak velocity, and termination was defined as the time at which joint angular velocity dropped below 5% of peak velocity. Custom algorithms developed in Matlab™ used a backward search from peak velocity to determine joint onsets. This method has been reported to be a robust method for determining movement onsets from time series data (Teasdale et al., 1993). For each movement trial, we defined the onset latency as the difference between initiation of spine joint motion and initiation of hip joint motion. We also determined latencies for peak joint velocity using the same convention. The latencies were then normalized by the movement time to the target. The normalized latencies from the movement trials with no instruction on movement strategy (i.e. knee flexion or knee extension) were analyzed using mixed model ANOVAs in which the within subject variables were movement phase (reach, return), movement speed (comfortable, fast pace), and target location (high, middle, low). The between subjects variable was gender.
The spine hip latencies from movement trials with specific instructions on movement strategy were analyzed separately using mixed model ANOVAs in which movement phase (reach, return), movement strategy (knee flexion, knee extension), and target location (middle, low) were within subject variables, and gender was the between subjects variable.
Calculating Spine Hip Ratios
To better understand the coupling of the spine and hip during these reaching tasks, we chose to examine the excursions of the spine and hip, as well as the spine hip ratio, across reaching task conditions. The joint angles were averaged over a 100 ms epoch at the beginning of the trial before the go signal, and another 100 ms epoch beginning 100 ms after target contact. The difference of these two values defined the joint angle excursions (ΔΦ). The spine hip ratio was defined as ΔΦspine/ΔΦhip.
To determine the effects of target location and movement speed on the excursions of the spine, hip and the spine hip ratio, we used mixed model ANOVAs in which target location (3) and movement speed (2), were within subjects factors, and gender was a between subjects factor.
3. Results
A typical movement trial in this study consisted of the subject reaching toward the target, pausing for 500 ms and then returning to an upright position. Fig. 2A depicts the joint angle time series data for a typical subject reaching toward the low target at a comfortable pace. Fig. 2A reveals distinct differences in the onset of spine motion relative to hip motion in both movement towards the target and in the return from target. Joint angular velocity (Fig. 2B) was derived from the time series angular data and was used to determine the timing of spine and hip joint motions. Examination of the joint angular velocity time series of this subject clearly reveals 1) the differences in onset timing and peak velocity between the spine and hip for both phases of the reaching task, and 2) the reversal in timing of the spine and hip motion for movement toward the target compared to the return to an upright posture (Fig. 2B).
Figure 2.

A) The time series data from the spine and hip joint of a typical subject performing a movement trial to the low target. B) The time series angular velocity data derived from the data in A. The differences in onset timing of the spine and hip are obvious from visual inspection of joint velocity data.
3.1 Movement Time
The movement time to target was significantly longer for comfortable pace movement trials compared to fast pace trials (F(1,14)=108.7, p<.05, η2p = .89). In the comfortable pace movement trials, subjects, on average, performed the movement task in 930 ms (SE ± 14.7 ms) and in 655 ms (SE ± 8.0 ms) for the fast-pace movement trials. There was no effect of target location or gender on the movement time to target. Although not explicitly controlled for, there was also a main effect of movement speed on the movement time for return to upright posture (F(1,14)=7.66, p<.05, η2p = .43). On average, subjects returned to upright posture in 800 ms (SE ± 19 ms) in comfortable pace trials and 675 ms (SE ± 9.3 ms) in fast pace trials. There was no effect of target location or gender on movement time for the return phase of the movement task.
3.2 Spine Hip Latencies
The mixed model ANOVA’s revealed a significant effect of movement phase on spine hip onset latency normalized to movement time (F(1,14)=12.4, p<.05, η2p = .55) and peak velocity latency normalized to movement time (F(1,14)=32.7, p<.05, η2p = .76). Fig. 3A and 3B illustrate clear differences in timing of the spine and hip joint motion for the reach phase compared to the return phase of the movement task (negative latencies indicate spine motion preceded hip motion, while positive latencies indicate hip motion preceded spine motion). However, there were no main effects of movement speed, target location, or gender on either onset or peak velocity latency. While the ANOVAs revealed significant differences in timing of the spine and hip for two phases of the movement, it did not test if the latencies (for each phase) were significantly different from zero. Thus, for each target location, separate T-Tests were performed on the spine hip onset latencies normalized to movement time, and peak velocity latencies normalized to movement time for both the reach and return phase of the movement. These analyses revealed that for the reaching phase of the movement tasks, onset of spine motion preceded hip motion (i.e. spine hip latencies were less than zero) for the high (t(15)= -2.73, p<.05), middle (t(15)= -5.22, p<.05), and low (t(15)= -7.21, p<.05) targets. For the return phase of the movement trials, onset of hip motion preceded spine motion (i.e. spine hip latencies were greater than zero) only for the middle (t(15)= 2.32, p<.05), and low targets (t(15)= 3.51, p<.05) (Fig 3A).
Figure 3.

The mean spine hip joint latencies normalized to movement time are plotted for both phases of the movement trials to each target location. A) Joint onset latencies, and B) Peak velocity latencies. The data were averaged across gender and movement speed and the error bars represent standard error of the mean. Negative values indicate that spine motion preceded hip motion and positive values indicate that hip motion preceded spine motion.
Peak spine velocity preceded peak hip velocity in the reaching phase of the movement tasks for high (t(15)= -3.59, p<.05), middle (t(15)= -3.59, p<.05), and low (t(15)= -9.73, p<.05) targets. However, for the return phase, peak hip velocity preceded peak spine velocity only for the middle (t(15)= 4.90, p<.05), and low targets (t(15)= 5.04, p<.05) (Fig 3B).
The differences in timing of the spine and hip joints will have an obvious effect on the spine hip ratios across both phases of the movement task (Fig 4.). In this figure, each phase of the movement task (i.e. reach and return) is broken up into quartiles and the spine hip ratios for each quartile of movement are plotted. It is clear that for the movement towards the target, spine motion is more prevalent in the first two quartiles (i.e. spine hip ratio >1) and then spine and hip motions are approximately equal as the spine hip ratio approaches a value near one for the last two quartiles. In contrast, for the return to upright posture, hip motion is prevalent for the first two quartiles of the movement (i.e. spine hip ratio <1) and then spine and hip motion are approximately equal as the spine hip ratio again approaches a value near one for the last two quartiles. This is consistent with the latencies reported and suggests that different movement strategies are used to initiate movement towards the target and the return to upright posture.
Figure 4.

The mean spine hip ratio (± SEM) for each quartile of movement for both phases of the movement task is plotted. The spine hip ratios for each quartile were averaged over target height, movement speed and gender. For the movement towards the target, the spine motion is greater than hip motion for the first two quartiles of motion. The opposite is true for the return to upright posture.
3.3. Spine Hip Latencies for Reaching Trials with Instructions on Knee Flexion
To determine if the movement pattern used to reach the targets influenced the timing of joint motion onset or peak velocity, we had participants reach for the middle and low targets with instructions to either bend their knees when they reach for the target or to keep the knees extended. We then compared the onset latencies and the peak velocity latencies from the two movement patterns using mixed model ANOVAs. There was no effect of movement strategy or gender on either onset or peak velocity latencies. Thus, while movement strategy used to perform this task will obviously influence the spine hip ratio (i.e. knee flexion will increase hip flexion and reduce the spine hip ratio), it had no influence on the onset timing of spine and hip joints.
3.4. Spine and Hip Joint Excursions
Given that these reaching tasks could be performed with a variety of movement strategies (e.g. knee flexion, knee extension), we sought to determine if there was a reasonably fixed coupling of the spine and hip across the different task conditions. If the relative excursions of the spine and hip are fixed across task conditions, then it would suggest an underlying strategy that simplifies control of these two joints. However, it is clear from Fig. 5A, that the spine hip ratio is not invariant across task conditions. Specifically, there was an interaction of movement speed and target location on the spine hip ratio (F(2,13)=4.14, p<.05, η2p = .41). Testing of simple effects revealed significant effects of movement speed for the high (F(1,14)=9.51, p<.05, η2p = .43), middle (F(1,14)=18.19, p<.05, η2p = .28), and low target (F(1,14)=11.3, p<.05, η2p = .47) locations. For each target location, the spine hip ratio decreased as movement speed increased. In contrast, testing of simple effects of target location revealed that there was no main effect of target location on the spine hip ratio for comfortable pace (F(2,13)=3.27, p<.05, η2p = .35) or fast pace (F(2,13)=3.53, p<.05, η2p = .37) movement trials. While the spine hip ratio was not significantly different across the three target locations, it appears from Fig. 5A that the spine hip ratio was greater for reaches to the high target compared to the middle and low targets. We next examined how excursions of the spine and hip were affected by task conditions.
Figure 5.

The effect of target location and movement speed on A) spine hip ratio, B) spine joint excursion, and C) hip joint excursion is depicted. The data were averaged across gender and standard error bars plotted.
While there was no effect of gender on the excursions of the spine and hip, as expected, there was a main effect of target location on the excursions of the spine (F(2,13)=109.1, p<.05, η2p = .94) and the hip (F(2,13)=76.2, p<.05, η2p = .92). On average, subjects used 18.8°, 32.2° and 44.2° of spine excursion for the high, middle and low targets respectively. Hip excursions for the three target locations were, on average, 21.5°, 39.8°, and 53.5°. There was also an interaction of movement speed and target location on spine excursion (F(2,13)=13.2, p<.05, η2p = .68). Testing of simple effects revealed significant effects of movement speed on the spine excursion for the middle (F(1,14)=10.1, p<.05, η2 = .44) and low target (F(1,14)=8.77, p<.05, ηp = .40) only (Fig 5B). For reaches to the middle target, on average, spine excursion was reduced from 34.6° to 29.8° for comfortable pace reaches compared to fast pace reaches. For reaches to the low target, spine excursion was reduced from 46.1° to 42.4° for the two movement speeds (Fig 5B). While spine excursion decreased as movement speed increased, the opposite was true for hip joint excursion (Fig 5C). Hip joint excursion in fast pace movement trials compared to comfortable pace trials, on average, increased from 17.2° to 35.4° for the high target, 35.3° to 44.2° for the middle target, and from 49.9° to 57.1°for the low target (F(1,14)=53.4, p<.05, η2p = .80).
3.5. Spine Hip Ratio in Forward Bend Test
Since most studies on the sine hip ratio have used the forward bend test (FBT) as the movement task, we included two trials of the FBT after all movement trials to the three target locations were complete. For the FBT, the spine hip ratio was, on average, 3.61 for the first quartile of movement, 3.39 for the second quartile, 1.67 for the third quartile, and 1.19 for the fourth quartile (Fig. 6). The spine hip ratio as defined in the methods (i.e. ΔΦspine/ΔΦhip) was, on average, 1.09 for the FBT which is consistent with the fourth quartile data presented in Fig (6).
Figure 6.

The mean spine hip ratio (± SEM) for each quartile of movement for the forward bend test is plotted.
4. Discussion
In the present study, we found significant differences in the timing of the spine and hip joints in healthy subjects performing full body reaching tasks. Furthermore, the differences in timing of the spine and hip joints were reversed for movements toward the target compared to the return movements to an upright posture. Importantly, we found that the timing of the spine and hip joint motions were not influenced by task manipulations such as target location and movement speed or by the choice of knee flexion strategy (i.e. knee flexion or knee extension) used to reach the target locations. The relative invariance in joint timing suggests an underlying control strategy that may optimize the mechanics of the spine hip coupling for flexion versus extension movement tasks. The timing differences in joint motion result in increased spine motion early in the movement, an increase in hip motion as the subject approached the target, and are consistent with previous findings on the coupling of the spine and hip (Esola et al., 1996; Lee & Wong, 2002; Paquet et al., 1994; Porter & Wilkinson, 1997; Wong & Lee, 2004).
In contrast to onset timing of the spine and hip joints, the spine hip ratio was not invariant to task manipulations of movement speed. As movement speed increased, the spine hip ratio decreased at all three target locations. The changes in spine hip ratio were driven by a decrease in spine motion along with an increase in hip motion as movement speed increased. With respect to target location, spine hip ratio was not significantly different for reaches to the three target locations.
4.1. Timing of Spine and Hip Joints
We found that, when averaged over all target locations and movement speeds, spine motion preceded hip motion by 48.9 ms for the reach phase of the movement task, and hip motion preceded spine motion by 63.0 ms for the return phase of the movement task. The differences in onset timing of the spine hip ratio during the movement are clearly illustrated in Figs. 3A & 3B. Our findings generally agree with other investigators who reported a delay in the movement of the spine when compared to the hip in subjects performing lifting tasks (Davis et al., 1965; Nelson et al., 1995). Nelson et al. (1995) reported that the lumbar pelvic motions were more sequential in the extension phase of a lifting task and more simultaneous in the flexion phase of the lifting tasks. Davis et al. (1965) inferred spine hip coordination by measuring the height of the lifted weight during the lifting task and reported that hip extension preceded lumbar spine extension in lifting trials using a knee flexion strategy as well as trials in which a knee extension strategy was used. This is consistent with our current findings on the effect of knee strategy on timing of the spine and hip joints during the return phase. However Davis et al. (1965) found that the load lifted influenced onset timing only in lifting trials in which subjects used a bent knee strategy. As the load increased there was a significant increase in the delay of lumbar extension. If differences in onset timing serve to reduce peak loading, then perhaps using a bent knee movement strategy allows for greater reduction of peak load while maintaining requirements for postural stability. Since our movement tasks did not require lifting any loads, there was no requirement for timing differences with the two movement strategies in order to further reduce loading of the lumbar spine.
Several investigators have reported that the spine hip ratio is greater than one during the initial portion of the flexion phase of the movement and less than one during the initial portion of the extension phase of the movement (Esola et al., 1996; Granata & Sanford, 2000; McClure et al., 1997; Paquet et al., 1994). Our findings on the changes in the spine hip ratio during the flexion and extension phases of the movement task are consistent with these previous investigations (Fig. 4). Although these previous investigators did not explicitly measure timing differences, we conjecture that their results were driven by differences in onset timing as we report here. However, other investigators suggest that motion of the spine and hip is simultaneous (Granata & Sanford, 2000; Lee & Wong, 2002). Granata and Sanford (2002) concluded that motion of the spine and pelvis was simultaneous, with hip motion dominant at extreme flexion angles. However, joint motion onset was never explicitly measured in this investigation and was inferred from angle-angle plots. Wong and Lee (2004) also found no time lag between the spine and hip in healthy subjects performing cyclic trunk flexion and extension tasks. Their conclusions were based on the high cross-correlations between the spine and hip joint time-series for these cyclic bending tasks. It could be that the method used to assess differences in timing of the joint motions was not particularly sensitive to timing differences, or perhaps the cyclic nature of the task may result in different coordination strategies that result in more simultaneous motions at the two joints. Previous research indicates that movement joint coupling is effected by whether a movement task is discrete or cyclic (Meulenbroek et al., 1993; Schillings et al., 1996).
While there have been very few investigations that explicitly measured joint onset timing of the spine and hip joints, there is evidence to suggest that joint movement onsets are influenced by the goals and constraints of the movement task (Kaminski et al., 1995; Lim et al., 2004; Putnam, 1993; Saling et al., 1996). Putnam (1993) described a proximal to distal organization of movement for both the upper and lower extremities for striking and throwing tasks. In order to attain the maximum speed at the distal segment, the movement must start at the proximal segment and progress distally. Thus, for striking and throwing it is clear that the goal requires a proximal to distal sequence of joint motion to maximize acceleration at the distal segment. In contrast, Lim et al (2004) found that in seated reaching movements made at a comfortable speed, the proximal joints lagged behind that of the hand as long as the target was within arm-length. If the target was further than an arm-length away, the differences in timing between the hand and the rest of the arm were very small. These two studies indicate that goals of the movement task can influence the timing and sequencing of the joint motions.
Consistent with the idea that task constraints can influence joint timing, we found reversals in the onset timing of the spine and hip for the two phases of this movement. With respect to the flexion phase of the reaching task, our findings were similar to those of Lim et al. (2004) in that the distal segment (the lumbar spine) moved first compared to the more proximal segment (the hips). However, this was reversed for the return to an upright posture and, therefore joint onset timing does not follow a strict distal to proximal order. In an earlier study of joint control strategies in multi-joint reaching, Kaminski and Gentile (1986) found a direct relationship between joint movement onset and joint displacement. Onset of joint motion occurred first in the joints with the largest displacement. If these findings generalized to full body reaching tasks, we would expect no differences in timing of joint motions and clearly would not expect a reversal in timing since hip and spine excursions are approximately equal for these tasks. Given that our findings do not fit cleanly with these other proposed rules (e.g. proximal to distal sequencing), it may be that the control strategy is biased towards movement efficiency and minimizing peak loading of the lumbar spine.
We speculate that the timing differences in spine and hip joints, and specifically, the reversal in timing between the two phases, serves to reduce loading on the spine. When initiating trunk flexion from an upright posture, the lumbar spine is very compliant (Dolan & Adams, 1993), and with the large mass of the trunk superior to the lumbar spine, it may be most efficient to allow gravity to assist in trunk flexion. That is, the trunk is positioned where gravity can be used to flex the lumbar spine and then initiate flexion of the hips. Once the trunk is moving into flexion, passive connective tissues such as the thoraco-lumbar fascia and iliolumbar ligaments that connect the lumbar spine segments and pelvis, may initiate the anterior rotation of the pelvis and hips. While these reaching tasks will obviously require active control by the para-spinal muscles as well as the hip extensors, our data suggest an optimization of the passive components of these tissues.
Conversely, during return movements to an upright posture, the mass of the trunk must be accelerated against gravity. It may be that the hip extensor muscles are better suited to overcome the large inertia of the forward flexed trunk (Farfan, 1975). Once the pelvis and hips have begun to rotate posteriorly, the same passive structures discussed previously would exert forces on the trunk, initiating trunk extension. Using the hips and pelvis to begin this acceleration may reduce the work load of the extensor muscles of the spine and reduce the peak compressive loading (Davis et al., 1965). The timing differences reported in this study were invariant to task manipulations that would necessarily affect peak loading of the spine. This finding suggests that there is an optimal joint timing that serves to minimize peak spinal loading. For example, fast paced reaches to a given target location will result in larger peak spinal loads than comfortable paced reaches to the same target but there was no effect on the onset timing of the spine and hip for these conditions. Thus, this suggests that timing of spine and hip joint motions are optimized to either reduce peak load or to capitalize on performance characteristics of the hip and spine extensors.
4.2. Spine/Hip Ratio
We found that there was no main effect of target location on spine hip ratio. However, on average, the spine hip ratio was 1.3 for the high target, .93 for the middle target and .88 for the low target. These data are in general agreement with data reported for male subjects in an earlier investigation (Thomas et al, 1998) that had subjects reaching to a 30 degree target and a 60 degree target. However, we did not find any main effects of gender on the spine hip ratio as reported by Thomas et al (1998). In that study female subjects tended to use a knee flexion strategy that resulted in an average spine hip ratio of .2. In the present study both male and female subjects used similar movement strategies to perform the reaching movements and thus had similar spine hip ratios.
The majority of the literature investigating spine hip ratios uses movement tasks that require subjects to maintain their knees in full extension (i.e. forward bend test). For these tasks, in general, it has been reported that the spine hip ratio decreases as a function of trunk flexion (Esola et al., 1996; Granata & Sanford, 2000; Lee & Wong, 2002; McClure et al., 1997; Porter & Wilkinson, 1997). In the present study, when subjects performed the FBT, the spine hip ratio decreased from the first quartile to the fourth quartile of movement (Fig. 6) in apparent agreement with previous investigations. However, the change in spine hip ratio is actually driven by timing differences rather than as a function of trunk angle. For example, while we found no significant main effect of target location on the spine hip ratio, it is obvious that the spine hip ratio decreased from the first quartile to the fourth quartile of movement in both the reaching tasks (Fig 4) and for the FBT (Fig 6). Furthermore, the excursions of the spine and hip from initial posture to the end of the movement task (i.e. target contact or end range flexion in the FBT) are approximately equal with a spine hip ratio around 1, even though the first and second quartile of the movement task were dominated by spine motion. This indicates that 1) there is a relatively fixed coupling of spine and hip excursions for movements to different areas of the workspace, and 2) early differences in the spine hip ratio are driven by differences of joint onset timing and not simply a function of trunk angle. This fairly close coupling of the spine and hip joint excursions for different portions of the reaching space indicates one way the central nervous system reduces complexity in this multi-joint task.
4.3. Movement Speed
With increases in movement speed, our subjects used less motion at the spine and more motion at the hips to complete reaches to the same target. This is consistent with Thomas et al. (2003) who found that increasing speeds of full body reaching tasks resulted in increased rotation of the pelvis and thigh segments, but not differences in the rotations of the trunk segment (Thomas et al., 2003). In healthy individuals, there may be a protective mechanism whereby the excursion of the lumbar spine is tightly regulated. That is, regardless of movement speed, subjects will only use a target specific amount of lumbar flexion. This may help prevent excessive loads and shearing forces on the lumbar spine. In fact preliminary evidence from our lab indicates that individuals with chronic low back pain use less lumbar flexion than healthy controls performing reaching tasks at fast paced conditions (Thomas, 2003).
5. Conclusion
Our findings indicate that there are clear differences in onset timing and peak velocity of the spine and hip joints during reaching tasks that necessitate some forward bending of the trunk. Furthermore, the timing is reversed for the return to an upright posture. This is in general agreement with clinical observations of healthy individuals performing forward bend tests (Cailliet, 1981; Sahrmann, 2002). However, it may be that patients with low back pain alter the timing of spine and hip joint motion to reduce extensor torque or loading of the lumbar spine and thus clinicians need to be aware of potential problems with interventions aimed at altering joint timing. Thus, future studies should 1) incorporate models to determine the effect of onset latencies on loading of the lumbar spine and 2) test for the influence of low back pain on timing differences in the spine and hip joints.
Acknowledgements
This work was supported by National Institutes of Health Grant grants RO1-HD045512.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Cailliet R. Low back pain syndrome. 3 ed. F. A. Davis Company; Philadelphia: 1981. [Google Scholar]
- Davis PR, Troup JD, Burnard JH. Movements of the thoracic and lumbar spine when lifting: A chrono-cyclophotographic study. Journal of Anatatomy. 1965;99:13–26. [PMC free article] [PubMed] [Google Scholar]
- Dolan P, Adams MA. Influence of lumbar and hip mobility on the bending stresses acting on the lumbar spine. Clinical Biomechanics. 1993;8(4):185–192. doi: 10.1016/0268-0033(93)90013-8. [DOI] [PubMed] [Google Scholar]
- Esola MA, McClure PW, Fitzgerald GK, Siegler S. Analysis of lumbar spine and hip motion during forward bending in subjects with and without a history of low back pain. Spine. 1996;21(1):71–78. doi: 10.1097/00007632-199601010-00017. [DOI] [PubMed] [Google Scholar]
- Farfan HF. Muscular mechanism of the lumbar spine and the position of power and efficiency. Orthopedic Clinics of North America. 1975;6(1):135–144. [PubMed] [Google Scholar]
- Granata KP, Sanford AH. Lumbar-pelvic coordination is influenced by lifting task parameters. Spine. 2000;25(11):1413–1418. doi: 10.1097/00007632-200006010-00014. [DOI] [PubMed] [Google Scholar]
- Kaminski TR, Bock C, Gentile AM. The coordination between trunk and arm motion during pointing movements. Experimental Brain Research. 1995;106(3):457–466. doi: 10.1007/BF00231068. [DOI] [PubMed] [Google Scholar]
- Lariviere C, Gagnon D, Loisel P. The effect of load on the coordination of the trunk for subjects with and without chronic low back pain during flexion-extension and lateral bending tasks. Clinical Biomechanics. 2000;15(6):407–416. doi: 10.1016/s0268-0033(00)00006-1. [DOI] [PubMed] [Google Scholar]
- Lee RY, Wong TK. Relationship between the movements of the lumbar spine and hip. Human Movement Science. 2002;21(4):481–494. doi: 10.1016/s0167-9457(02)00117-3. [DOI] [PubMed] [Google Scholar]
- Lim S, Martin BJ, Chung MK. The effects of target location on temporal coordination of the upper body during 3d seated reaches considering the range of motion. International Journal of Industrial Ergonomics. 2004;34:395–405. [Google Scholar]
- McClure PW, Esola M, Schreier R, Siegler S. Kinematic analysis of lumbar and hip motion while rising from a forward, flexed position in patients with and without a history of low back pain. Spine. 1997;22(5):552–558. doi: 10.1097/00007632-199703010-00019. [DOI] [PubMed] [Google Scholar]
- Meulenbroek RG, Rosenbaum DA, Thomassen AJ, Schomaker LR. Limb-segment selection in drawing behaviour. Quarterly Journal of Expimental Psychology [A] 1993;46(2):273–299. doi: 10.1080/14640749308401047. [DOI] [PubMed] [Google Scholar]
- Nelson JM, Walmsley RP, Stevenson JM. Relative lumbar and pelvic motion during loaded spinal flexion/extension. Spine. 1995;20(2):199–204. doi: 10.1097/00007632-199501150-00013. [DOI] [PubMed] [Google Scholar]
- Paquet N, Malouin F, Richards CL. Hip-spine movement interaction and muscle activation patterns during sagittal trunk movements in low back pain patients. Spine. 1994;19(5):596–603. doi: 10.1097/00007632-199403000-00016. [DOI] [PubMed] [Google Scholar]
- Porter JL, Wilkinson A. Lumbar-hip flexion motion. A comparative study between asymptomatic and chronic low back pain in 18- to 36-year-old men. Spine. 1997;22(13):1508–1513. doi: 10.1097/00007632-199707010-00017. discussion 1513-1504. [DOI] [PubMed] [Google Scholar]
- Putnam CA. Sequential motions of body segments in striking and throwing skills: Descriptions and explanations. Journal of Biomechics. 1993;26(Suppl 1):125–135. doi: 10.1016/0021-9290(93)90084-r. [DOI] [PubMed] [Google Scholar]
- Sahrmann SA. Diagnosis and treatment of movement imapirment syndromes. Mosby; Saint Louis: 2002. [Google Scholar]
- Saling M, Stelmach GE, Mescheriakov S, Berger M. Prehension with trunk assisted reaching. Behavioural Brain Research. 1996;80(12):153–160. doi: 10.1016/0166-4328(96)00030-7. [DOI] [PubMed] [Google Scholar]
- Schillings JJ, Meulenbroek RGJ, Thomassen AJWM. Limb segment recruitment as a function of movement direction, amplitude, and speed. Journal of Motor Behavior. 1996;28(3):241–254. doi: 10.1080/00222895.1996.9941749. [DOI] [PubMed] [Google Scholar]
- Teasdale N, Bard C, Fleury M, Young DE, Proteau L. Determing movement onsets from temporal series. Journal of Motor Behavior. 1993;25:97–106. doi: 10.1080/00222895.1993.9941644. [DOI] [PubMed] [Google Scholar]
- Thomas JS. The influence of chronic back pain on kinematic rules underlying multijoint reaching tasks. Society for Neuroscience. CD-ROM; Washington, DC: 2003. Program No. 767.13 Abstract Viewer/Itinerary Planner. [Google Scholar]
- Thomas JS, Corcos DM, Hasan Z. The influence of gender on spine, hip, knee, and ankle motions during a reaching task. Journal of Motor Behavior. 1998;30(2):98–103. doi: 10.1080/00222899809601327. [DOI] [PubMed] [Google Scholar]
- Thomas JS, Corcos DM, Hasan Z. Effect of movement speed on limb segment motions for reaching from a standing position. Experimental Brain Research. 2003;148:377–387. doi: 10.1007/s00221-002-1287-2. [DOI] [PubMed] [Google Scholar]
- Thomas JS, Corcos DM, Hasan Z. Kinematic and kinetic constraints on arm, trunk, and leg segments in target-reaching movements. Journal of Neurophysiology. 2005;93(1):352–364. doi: 10.1152/jn.00582.2004. [DOI] [PubMed] [Google Scholar]
- Wong TK, Lee RY. Effects of low back pain on the relationship between the movements of the lumbar spine and hip. Human Movement Science. 2004;23(1):21–34. doi: 10.1016/j.humov.2004.03.004. [DOI] [PubMed] [Google Scholar]