

SUMMARY
Normal patterning of the developing limb requires a tight restriction of Sonic hedgehog (Shh) mRNA to the posterior margin of the limb bud. While several positive and negative regulatory factors have been identified which serve to position the Shh expression domain in the distal posterior limb, these factors cannot in themselves explain the tight restriction of Shh to the posterior margin, nor can they explain the similarly tight restriction of Shh to the anterior margin when the regulatory factors are disrupted or misexpressed. We suggest that the transcription factors Tbx2 and Tbx3 are excellent candidates for positively-acting factors responsible for limiting Shh expression to the margins of the limb bud. These closely related factors are indeed expressed at the anterior and posterior limb margins over a wide range of limb bud stages. Moreover, previous reports indicate that in addition, misexpression of Tbx2 beyond the limb margin is sufficient to anteriorly expand Shh, and conversely, antagonizing Tbx2 function leads to loss of Shh. In contrast to this idea, previous models have placed Tbx2 expression downstream of Shh and Bone Morphogenetic Protein (BMP) signaling. We find, however, that Tbx2 expression is neither affected by blocking Shh signaling with cyclopamine nor by genetic removal of several BMP activities in the limb bud. To understand the true source of the positional information responsible for limiting Tbx2, Tbx3 and Shh expression to the marginal mesenchyme of the limb bud, we undertook a series of grafting and extirpation experiments, which led to the identification of the dorsal-ventral (DV) border ectoderm exclusive of the apical ectodermal ridge (AER) as a new signaling center in the limbbud. We find that maintenance of Tbx2 expression in the limb mesoderm requires proximity to the non-AER D-V border. Using chick-quail graft chimeras, we find that a graft of the non-AER D-V border ectoderm to a location on the surface of the middle of the limb bud is sufficient to induce ectopic expression of Tbx2 in underlying mesoderm. These data demonstrate that the non-AER D-V border ectoderm is necessary and sufficient for Tbx2 expression at the anterior and posterior limb margins. Similarly, we find that a graft of the non-AER D-V border can expand the domain of Shh anteriorly when grafted just anterior to the ZPA. It is notable that Tbx2 expression does not extend distally to the mesoderm underlying the AER. Moreover, we find that grafts of the AER to more proximal locations result in downregulation of Tbx2 expression, suggesting that the AER produces a negatively-acting signal opposing the activity of the non-AER DV border ectoderm. Indeed, implantation of beads soaked in fibroblast growth factor 8 (Fgf8), expressed in the AER, downregulates Tbx2 expression. The data presented here identify the non-AER border of dorsal-ventral ectoderm as a new signaling center in limb development that localizes the ZPA to the limb margin. This finding explains the tight restriction of Shh expression to the posterior margin throughout limb outgrowth as well as the tight restriction of Shh expression to the anterior margin in many mutants exhibiting preaxial polydactyly.
INTRODUCTION
Developmental fields are often organized by inductive cues emanating from distinct signaling centers. One important signaling center in limb development is the Zone of Polarizing Activity (ZPA). In a classical experiment by Saunders and Gasseling (1968), the ZPA was identified as a population of cells in the distal posterior margin of the chick limb bud capable of inducing mirror-image digit duplications when grafted into the anterior distal margin of another limb bud. We now know that limb bud cells with ZPA activity are congruent with cells that express the secreted factor Sonic hedgehog (Shh) (Riddle et al., 1993). Grafting cells that express Shh or a bead soaked in Shh protein into the anterior margin mimics the mirror-image digit duplications that ensue from ZPA grafts (Lopez-Martinez et al., 1995; Riddle et al., 1993; Yang et al., 1997).
These grafting experiments dramatically demonstrate the necessity for limiting Shh expression to the distal posterior margin of the limb bud if normal patterning is to emerge. Yet the localization of Shh expression at the distal posterior margin is not simply a passive result of the intrinsic properties of the ZPA cells themselves, but rather the Shh expression domain is continually reestablished along both the proximal distal and anterior-posterior axes. As the limb bud grows out, Shh expression is only maintained in the distal-most cells of the posterior margin, and is lost in more proximal cells even though they are descended from the ZPA (Vargesson et al., 1997). This distal shift in Shh expression is well understood. There is a second signaling center in the ectoderm at the distal tip of the limb bud called the Apical Ectodermal Ridge (AER). The AER, which forms at the border between the dorsal and ventral planes of ectoderm at the tip of the limb bud (hence forming a ridge running along the anterior-posterior axis of the future hand plate) is known to express members of the Fgf family which are required for maintenance of Shh expression (Laufer, 1994; Niswander, 1994). Thus, as the limb bud grows out only the ZPA cells towards the distal tip, within range of the Fgf signaling, are capable of continued Shh expression.
A similar phenomenon, albeit much less well understood, appears to continuously restrict Shh expression to the extreme posterior margin. Fate mapping studies have revealed that as the limb bud grows, descendants of Shh-expressing cells expand to populate most of the posterior third of the limb bud, a domain much larger than the region actively expressing Shh (Harfe et al., 2004). Thus, as this cell population proliferates, more anterior members of the population shut off Shh expression such that the Shh expression domain always remains tightly restricted to the posterior margin. While the factor(s) responsible for this tight posterior restriction are unknown, several factors (both negatively and positively acting) have been identified which together are responsible for broadly localizing Shh activity to the posterior and not anterior portions of the limb bud. Gli3 and Alx4 are negative regulators of Shh, expressed in the anterior of the limb bud and required to repress Shh in that domain.
Mice carrying mutations in these genes exhibit ectopic Shh in the anterior limb bud and subsequent preaxial polydactyly (Hui and Joyner, 1993; Masuya et al., 1995; Qu et al., 1998; Takahashi et al., 1998). Conversely, the transcription factors Hoxb-8 (in the forelimb bud) and Hand2 (previously called dHand) are broadly expressed in the posterior limb bud and positively regulate Shh expression. When these transcription factors are ectopically expressed in the anterior limb mesenchyme, they result in ectopic expression of Shh in the anterior (Charite et al., 1994; Charite et al., 2000; Fernandez-Teran et al., 2000).
While these factors localize the ZPA roughly to the posterior limb, their broad expression domains imply that an additional regulatory mechanism restricts Shh to the extreme posterior margin. Intriguingly, either disruption of the negative-acting regulatory factors, Gli3 or Alx4 (Hui and Joyner, 1993). (Qu et al., 1998; Masuya et al., 1995) or global misexpression of the positive-acting regulating factors Hoxb8 in the forelimb or Hand2 (in either the fore or hindlimb) (Charite, 1994; Charite, 2000, Fernandez-Teran, 2000), result in ectopic anterior expression domains of Shh which are tightly restricted to the anterior margin. Together, these findings provide compelling evidence that an additional mechanism is responsible for the tight restriction of Shh to the posterior margin, and moreover, that the same mechanism is likely responsible for restricting Shh to both anterior and posterior margins.
Strikingly, the T-box transcription factors Tbx2 and Tbx3 are expressed similarly in tight stripes along the anterior and posterior margins of the developing limb bud (Gibson-Brown et al., 1998; Isaac et al., 1998; Logan et al., 1998). These transcriptional repressors are related to the Drosophila gene optomotor blind (omb) and have been implicated in morphogenesis and organogenesis of a variety of tissues (Papaioannou, 2001; Papaioannou and Silver, 1998). Based on previous experiments in the chick limb bud, it has been postulated that Tbx3 is regulated downstream of signals which pattern the anterior-posterior axis such as Shh (Tumpel et al., 2002). However, reexamination of the literature suggests that the epistasis may actually be the reverse of this: in the human ulnar-mammary syndrome, haploinsufficiency of TBX3 leads to a reduced or absent ulna and loss of digit V in patients, suggestive of a loss-of-function of SHH (Bamshad et al., 1997). Indeed, this phenotype is phenocopied in mice lacking Tbx3 which exhibit a reduced or absent ulna/fibula, a reduced or absent digit V, and occasionally a reduced or absent digit IV, and these defects correspond with reduced or absent expression of Shh (Davenport et al., 2003). Similar data have been reported in the chick. Tbx2 and Tbx3 are believed to function exclusively as transcriptional repressors, hence fusing the DNA-binding domains of those proteins to the VP16 transcriptional activation domain, is predicted to yield a dominant-negative variant. As in the loss of function in mice, viral misexpression of VP16ΔTbx2 and VP16ΔTbx3 mutant genes, results in reduction or loss of posterior digits in the chick hindlimb (Suzuki et al., 2004). While these phenotypes were interpreted in terms of a direct role for Tbx genes in digit specification, the viral misexpression of VP16ΔTbx2 was also noted to reduce levels of Shh (Suzuki et al., 2004). Thus, a more parsimonious interpretation is that there is simply a loss of polarizing activity, and that Tbx2 and Tbx3 function are necessary for Shh expression. Conversely, viral misexpression of Tbx2 anteriorly expands the endogenous domain of Shh in the chick limb (Suzuki et al., 2004), indicating that Tbx2 activity is also sufficient to induce or maintain Shh expression within the context of the posterior limb bud.
Given the restriction of Tbx2 and Tbx3 to the anterior and posterior margins and the apparent requirement of these genes for initiation and/or maintenance of Shh, it is likely that these genes are involved in the restriction of Shh to the limb margin. From the standpoint of understanding the spatial localization of Shh expression, however, this only begs the question, how are the Tbx2 and Tbx3 genes themselves restricted to the anterior and posterior limb margins? In this study, we identify the dorsal-ventral ectoderm border proximal to the AER as a new signaling center in limb development and show that it serves to regulate the restricted expression of Tbx2 and Shh. We demonstrate that Tbx2 is not directly dependent on signals which pattern the anterior-posterior axis as previously postulated. We find that, instead, maintenance of Tbx2 requires proximity to the dorsal-ventral (D-V) border ectoderm at the limb margins. Even more, the D-V border ectoderm when grafted to the mid-limb is sufficient to induce ectopicTbx2. As would be predicted from expansion of Tbx2, the D-V border ectoderm when grafted in the posterior limb is capable of expanding the domain of Shh anteriorly. Finally, we investigate the exclusion of Tbx2 from subapical mesoderm and show that the AER signal Fgf8 negatively regulates mesodermal expression of Tbx2. Our data provide evidence that the non-AER D-V border ectoderm is an important signaling center, responsible for maintaining Shh expression specifically at the posterior margin of the limb.
MATERIALS and METHODS
Embryos
Experiments on wild-type chick embryos were performed on standard specific pathogen-free white Leghorn chick embryos provided by SPAFAS (Norwich, Connecticut). Fertile Japanese quail eggs were obtained from Truslow Farms. Eggs were incubated, windowed, and staged as described previously (Hamburger, 1951). Mouse Bmp2 and Bmp4 double mutants were generated as detailed elsewhere (Bandyopadhyay et al., submitted). Briefly, excision of floxed alleles of Bmp2 and Bmp4 was driven by a Prx1::Cre gene, resulting in removal of Bmp2 prior to any endogenous expression and removal of Bmp4 after only a few hours of very low expression, as judged by whole mount in situ hybridization.
D-V Border Ectoderm Grafts
The ectoderm overlying the anterior or posterior limb margins was removed using an established technique for limb ectoderm removal (Yang and Niswander, 1995). Briefly, 1 μL of 1.0% Nile in PBS was pipetted over the limb margin. After 2 minutes, slight “blistering” of the ectoderm peeling off the mesoderm could be observed. The rectangle of ectoderm overlying the margin (both dorsal and ventral ectoderm as well as the dorsal-ventral ectoderm border) was cut using a tungsten needle. The rectangle of ectoderm was transferred to a host limb using a mouth pipet. The host limb was prepared by removing an equivalent rectangle of ectoderm from the dorsal surface of the mid-limb, and the dorsal-ventral border ectoderm was set in its place. Each ectoderm graft was held in place for 5–15 minutes to allow adequate sticking to the host mesoderm surface and hence to prevent loss of the ectoderm graft. A slight amount of bleeding from the host mesoderm surface greatly enhanced adhesion of the graft. Grafts from quail to chick limbs were performed in the same way.
Cyclopamine Treatment
Cyclopamine (Toronto Research Chemicals) was added directly over the forelimb in ovo as previously described (Incardona et al., 1998). Briefly, 5 microliters of 1.0 mg/mL cyclopamine in 45% HBC (Sigma) in PBS was added over the limb.
Mesoderm Swapping Experiment
A square of mesoderm under the anterior margin dorsal-ventral ectoderm border was cut cleanly using tungsten needles. Another square of mesoderm of equivalent size was cut in the mid-limb. These squares of tissue were swapped and grafted firmly into the square hole. To repeat this experiment with quail donor tissue, a square of anterior margin tissue was cut from a quail limb and transferred to a chick host limb using a mouth pipet. A square of chick tissue was removed from the mid-limb to allow grafting of the chick square tissue.
Whole Mount In Situ Hybridization and Quail Antibody Detection
Whole mount in situ hybridization was performed as previously described (Dietrich et al., 1997) with minor modifications. Briefly, embryos were fixed overnight in 4% paraformaldehyde, washed in PBS, dehydrated into methanol, and bleached for 1 hour in 6% hydrogen peroxide in methanol. Embryos were then rehydrated into PBT and permeabilized with 10 μg/mL proteinase K in PBT for 20–30 minutes depending on embryonic stage. Embryos were then washed in PBT, post-fixed in 4% paraformaldehye/0.2% glutaraldehyde for 20 minutes, washed in PBT, and prehybridized for 1 hour at 70°C in hybridization buffer (50% formamide, 5X SSC pH 4.5, 2% SDS, 2% blocking reagent (Roche), 250μg/mL tRNA, 100μg/mL heparin). The embryos were then hybridized overnight at 70°C in hybridization buffer with probes. After hybridization, the embryos were washed four times for 30 minutes each in 50% formamide/2X SSC ph 4.5/1% SDS and then washed in MABT (100mM maleic acid, 150 mM NaCl, ph 7.5, 0.1% Tween-20). Embryos were blocked for 1 hour in 2% blocking reagent/MABT followed by 1 hour in 2% blocking reagent/20% heat inactivated goat serum/MABT, and then incubated overnight at 4°C with secondary antibody (1:2500 anti-DIG AP, Roche). Embryo were then washed multiple times in MABT at room temperature and washed overnight at 4°C. Embryos were then equilibrated in NTM (100mM NaCl, 100mM Tries pH 9.5, 50 mM MgCl2) and color detection was performed with NBT/BCIP (Sigma). DIG-labeled probes were generated for chick Fgf8, Tbx2 (Suzuki et al., 2004), and Shh (Riddle et al., 1993).
For whole mount quail antibody detection, embryos fixed in 4% paraformalydehyde and dehydrated into Dent’s fix (20% DMSO in methanol) and bleached. Embryos were then rehydrated into PBS + 0.1% Triton-X (PBS-Tx) and incubated overnight at 4°C with 1:10 dilution of QCPN antibody (Developmental Studies Hybridoma Bank) in 10% sheep serum/PBS-Tx. Embryos were washed several hours with PBS-Tx and then incubated overnight at 4°C with 1:100 secondary antibody (goat-anti-mouse HRP-conjugated, Molecular Probes) in 1% goat serum/PBS-Tx. Embryos were washed several hours with PBS-TX and then signal was detected by DAB color reaction (Sigma).
Section In Situ Hybridization and Quail Antibody Detection
Embryos were embedded in paraffin and 12μm sections were collected. Section in situ hybridization was performed as previously described (Murtaugh et al., 1999) with minor modifications. Paraffin sections were dewaxed in xylenes, washing ethanol, and rehydrated into PBS, fixed for 10 minutes in 4% paraformaldehyde, washed in PBT, and permeabilized with 1.0 μg/mL proteinase K for 10 minutes. Sections were then washed in PBS, fixed in 4% paraformaldehyde for 5 minutes, and acetylated in 0.25% acetic anhydride in 0.1 M triethanolamine for 10 minutes, washed in PBS, rinsed in water, and air dried for 30 minutes. RNA probe was then added in 100 μL of hybridization buffer (10 mM Tris pH 7.5, 600 mM NaCl, 1mM EDTA, 0.25% SDS, 10% Dextran Sulfate, 1X Denhardt’s, 200 μg/mL yeast tRNA, 50% formamide). Slides were covered with coverslips cut from polypropylene bags, placed in chambers humidified with 1X SSC/50% formamide, and incubated overnight at 65°C. The next day, coverslips were removed in 5X SSC and slides were washed for 30 minutes in 1X SSC/50% formamide at 65°C. Slides were then transferred to TNE (10mM Tris pH 7.5, 500 mM NaCl, 1mM EDTA) at 37°C for 10 minutes, incubated in RNase A (20 μg/mL, Roche) in TNE for 30 minutes at 37°C, and then washed in TNE for 10 minutes. Sections were then washed in 2X SSC for 20 minutes at 65°C, then washed twice for 20 minutes each in 0.2X SSC, and then transferred into MABT. Slides were blocked in 2% blocking reagent (Roche)/20% heat inactivated goat serum/MABT for 1 hour. Secondary antibody was added (anti-DIG AP antibody, 1:2500, Roche) in 2% blocking reagent/20% heat inactivated goat serum/MABT, and slides were incubated overnight at 4°C. The next day, slides were washed in MABT and equilibrated in NTM for 10 minutes. Color detection was performed using Fast Red (Sigma). DIG-labeled probes were used for chick Tbx2 (Suzuki et al., 2004).
Quail antibody was detected by rinsing slides in PBT, then incubating in 1:20 QCPN antibody in 5% heat inactivated goat serum/PBS, overnight at 4°C. The next day, slides were washed in PBT and then incubated with secondary antibody (Cy2-conjugated anti-mouse antibody, Jackson Immunoresearch) overnight at 4°C. The next day, slides were rinsed in PBT and visualized for immunofluorescence.
Bead Implants
Affi-Gel Blue beads (BioRad) were used to administer Fgf8 protein to limbs. Beads were washed in PBS and incubated at room temperature for 1 hour in 1.0 mg/mL human Fgf8 (PeproTech). Beads were then implanted into a small slit made in the anterior or posterior margin of the limb.
RESULTS
Expression of Tbx2 at Anterior and Posterior Margins of the Limb
Tbx2 and Tbx3 genes have been described as having very similar expression patterns during limb development, both restricted to the anterior and posterior margins of wing and leg buds, though Tbx3 in slightly wider domains (Gibson-Brown et al., 1998; Tumpel et al., 2002). For the purposes of this study, we chose to focus on Tbx2. To better understand how expression of Tbx2 relates to limb bud formation and outgrowth, we first characterized mesodermal Tbx2 expression at greater resolution than has been previously published, examining the wing bud from HH stages 15 through 27 (Fig. 1). Prior to wing bud formation at HH stage 15–16, Tbx2 is expressed in a strip running the length of the lateral plate mesoderm encompassing the presumptive wing region and flank (Fig. 1A,B). Upon emergence of the wing bud at HH stage 17–18, Tbx2 continues to be expressed in the flank and limb, but begins to disappear from the distal central limb (Fig. 1C,D). From HH stage 19–21, exclusion of Tbx2 from the central limb becomes more pronounced while expression continues at the anterior and posterior margins of the limb as well as in the flank (Fig. 1E–G). By HH stage 22, the stripes of Tbx2 expression at the anterior and posterior margins of the limb are clearly established (Fig. 1H). Shortly after this time, Tbx2 expression is down-regulated in the flank but maintained at high levels in the limb (Fig. 1K). This restriction to the anterior and posterior limb margins and flank continues through HH stage 27 and is apparent in both forelimbs and hindlimbs (Fig. 1I–L). At HH stage 27, Tbx2 begins to also be expressed in the posterior interdigital mesoderm (arrowhead, Fig. 1J).
Fig. 1.
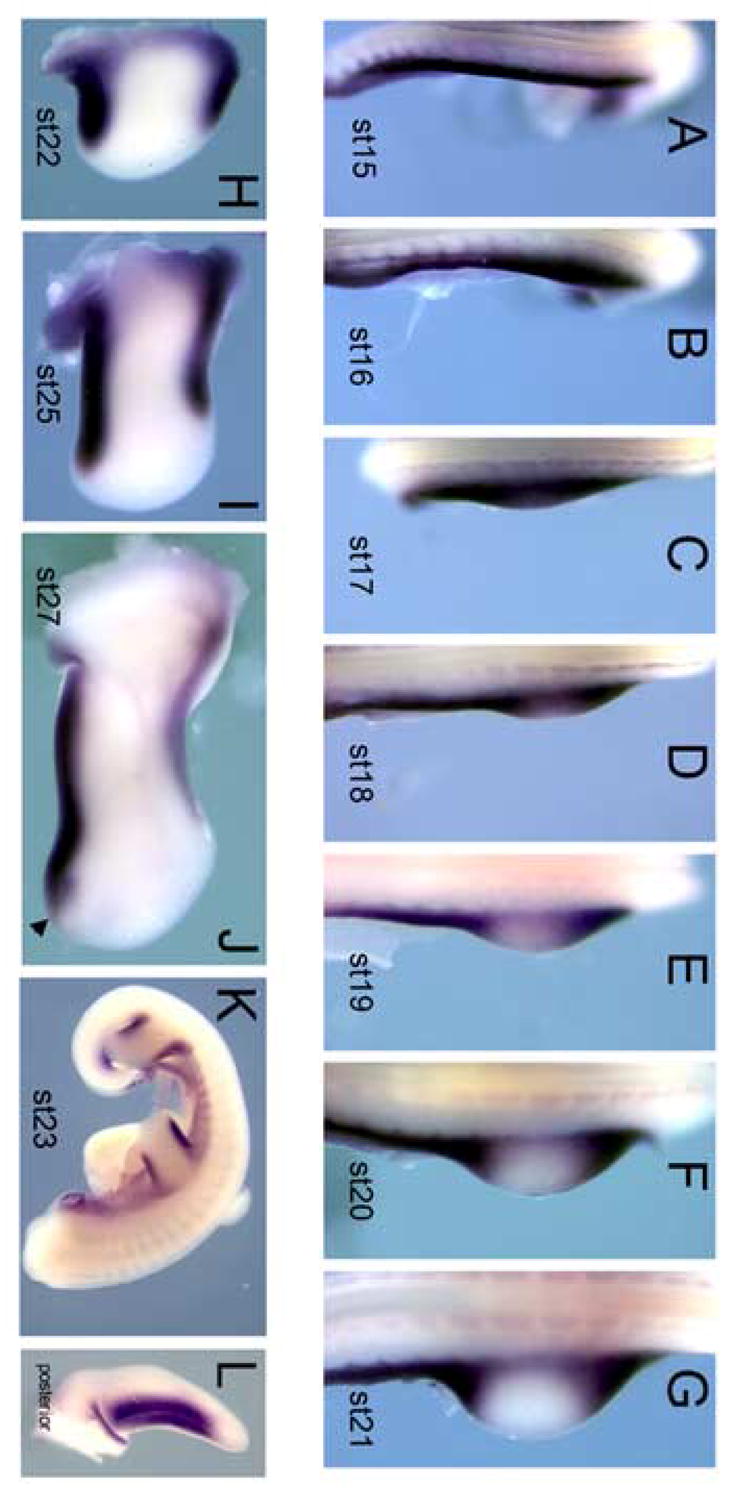
Expression of Tbx2 throughout limb bud development. (A–J) Dorsal view before and after limb bud formation from HH stages 15–27. (A–C) Prior to limb bud formation and just as the limb bud emerges, Tbx2 is expressed throughout the lateral plate mesoderm in the presumptive wing region and flank. (D–G) Upon emergence of the wing bud, Tbx2 begins to disappear from the mid-limb but continues to be expressed in the anterior and posterior of the limb as well as in the flank. (H–I, K) By HH stage 22, Tbx2 expression in the limb is restricted to the anterior and posterior margins. (K) In the HH stage 23 embryo, Tbx2 expression is restricted to the flank and the anterior and posterior margins of both forelimb and hindlimb. This restriction to the limb margins continues through HH stage 27 (J). At HH stage 27, Tbx2 is also expressed in the posterior interdigital mesoderm (arrowhead). (L) Posterior view of a HH stage 24 limb. The domain of Tbx2 tightly parallels a faint line that demarcates the non-AER D-V ectoderm border.
Relative Epistasis of Shh and Tbx Genes
Our reading of the literature led us to postulate that Tbx gene activity acts upstream to regulate Shh expression. There is evidence that Tbx3 is necessary to initiate and/or maintain proper expression of Shh (Davenport et al., 2003; Suzuki et al., 2004) and ectopic misexpression of Tbx2 is sufficient to expand the Shh expression domain (Suzuki et al., 2004). However, the reverse epistatic relationship has also been proposed. In particular, it has been suggested that the posterior stripe of Tbx3 expression is positively regulated by Shh and by BMP2 which is itself induced by Shh. In contrast, the anterior stripe of Tbx3 was proposed to be negatively regulated by Shh but positively regulated by BMP4. One difficulty in this model lies in the relative expression domains of these genes. While at HH stage 22, the limb expression of Tbx2 encompasses expression of Shh in the posterior margin, extending more proximally and slightly more anteriorly than the Shh domain (Fig. 2A), by HH stage 25, the posterior Tbx2 domain can be seen to extend much further proximally than Shh (Fig. 2A). It is not immediately clear why Shh and/or BMP2 signaling would be able to maintain Tbx3 expression at such a long distance proximally, yet be unable to do so to an equivalent distance anteriorly. To directly examine whether Tbx2 is regulated by Shh activity, we tested for changes in Tbx2 expression after blocking the activity of Shh with cyclopamine in a HH stage 22 limb. Cyclopamine is a small steroidal alkaloid that blocks the cellular response to Shh (Cogler, 1998; Incordena, 1998). Whereas genes upregulated directly in response to Shh such as Ptc1, Ptc2, and Gli1 are downregulated as quickly as 4 hours after cyclopamine is added to the limb (Scherz et al., submitted), Tbx2 expression was maintained at the anterior and posterior margins 8 hours after addition of cyclopamine (n=6) (Fig. 2B,C). To test the necessity for BMP signaling to attain proper Tbx gene regulation, we examined Tbx2 expression in mouse limbs in which both BMP2 and BMP4 had been genetically removed. Tbx2 expression appears unchanged in these limbs compared to expression in wild-type limbs (Fig. 2D–E). Because BMP7 is, like BMP2, a target of Shh signaling in the posterior limb bud, it was possible that they might play redundant roles in regulating Tbx gene expression. We therefore, also analyzed limb buds devoid of both BMP2 and BMP7 activity, and again saw no alterations in Tbx2 expression (data not shown).
Fig. 2.
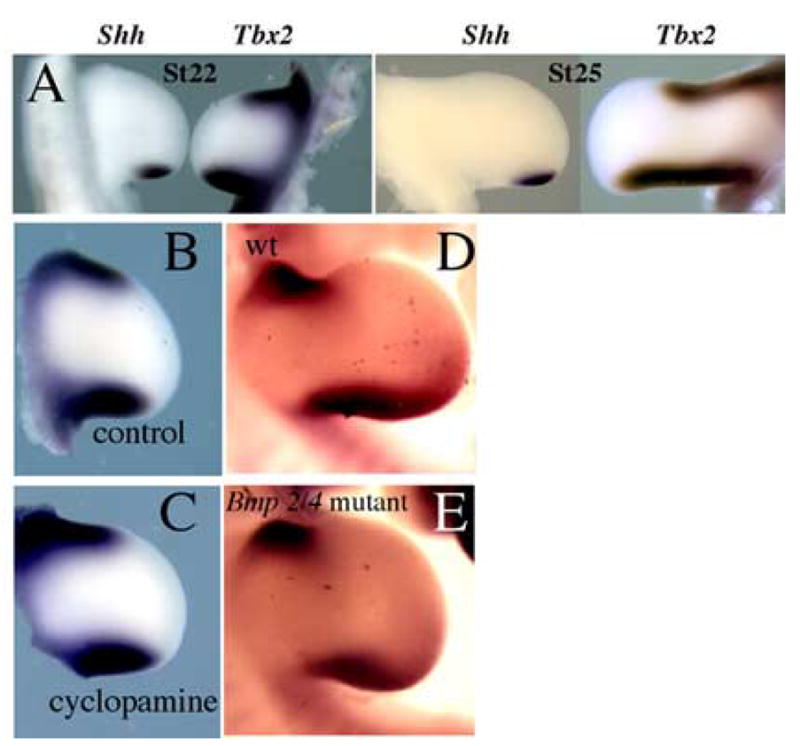
Expression of Tbx2 is not directly dependent on signals which pattern the anterior-posterior axis. (A) Comparison of Shh and Tbx2 expression at HH stage 22 and HH stage 25. (B,C) Control limb and limb treated with cyclopamine and examined after 8 hours for changes in Tbx2 expression. (D,E) Comparison of Tbx2 expression in wildtype mouse limb vs. limb lacking Bmp2 and Bmp4 activity.
Regulation of Tbx2 by D-V Border Ectoderm
Signaling between the epithelial and mesenchymal tissues of the limb play many key roles in orchestrating limb development (for example, the signaling between the AER and ZPA discussed above). Thus, an attractive alternative hypothesis for explaining the regulation of Tbx2 gene expression would be signals emanating from the overlying ectoderm rather than from the mesenchyme itself. The anterior and posterior margins of the limb mesenchyme lie underneath the location where the dorsal and ventral ectodermal surfaces meet (see posterior view of the edge of a HH stage 22 wing bud in Fig. 1L). The dorsal and ventral ectoderm have distinct properties. For example, the dorsal ectoderm expresses the secreted protein Wnt7a (Dealy et al., 1993; Parr et al., 1993), while the ventral ectoderm expresses members of the Bmp family (Lyons et al., 1990; Pizette et al., 2001). Moreover, at the distal tip of the limb bud, the border between the dorsal and ventral ectoderm forms a unique morphological structure with distinct signaling properties, the AER. These considerations suggested that either a combination of signals from the dorsal and ventral ectoderm only found in proximity to each other at the dorsal-ventral border or alternatively, a unique signal produced by the dorsal-ventral border ectoderm itself might be responsible for regulating expression of Tbx genes in the subjacent mesenchyme.
To test if signals present at the border between dorsal and ventral ectoderm regulate Tbx2 expression in the anterior and posterior margins, we swapped a block of mesoderm at the anterior margin with a block of mesoderm in the mid-limb at HH stage 22/23 (Fig. 3A). This procedure effectively moves tissue that expresses Tbx2 away from the D-V border and thereby tests the requirement of the D-V border to maintain Tbx2 expression. At the same time, tissue that does not express Tbx2 is moved under the D-V border, testing the sufficiency of the D-V border to upregulate Tbx2 in underlying mesoderm. Importantly, the surgery was designed such that the D-V border ectoderm is kept intact during the operation (arrowhead). This swap can be appreciated by analyzing Tbx2 expression 2 hours after the operation (Fig. 3B). A block of limb tissue expressing Tbx2 has been grafted into the mid-limb away from the D-V border, and a block of limb tissue not expressing Tbx2 has been grafted under the D-V border. After 24 hours, the block of tissue moved away from the D-V border into the mid-limb loses its Tbx2 expression (n=9/9) (Fig. 3C). The same was observed when a block of tissue expressing Tbx2 in the posterior limb was moved away from the posterior D-V border into the mid-limb (n=2/3) (Fig. 3F); we noted that Tbx2 may be maintained in the posterior mesoderm grafts longer than in the anterior mesoderm grafts, as weak expression of Tbx2 in a block of posterior limb tissue persisted at 24 hours after grafting to the mid-limb (n=1/3), a finding consistent with similar persistence observed with Tbx3 expression (Tümpel et al, 2002). The loss of Tbx2 in mesoderm moved away from the D-V border suggests that the D-V border is necessary to maintain Tbx2 expression. To verify that loss of Tbx2 expression after 24 hours is not simply due to loss of the graft, we performed a similar procedure grafting a piece of quail anterior margin tissue into the chick mid-limb. The quail tissue can be unambiguously identified using a specific antibody. As expected, 24 hours (n=2/2) and even 48 hours (n=2/2) after the operation, the quail graft is viable and appears to have integrated and grown with the host chick limb (Fig. 3D,E). Quail grafts remain viable in other regions of the limb as well, including the anterior and posterior margins (data not shown). These data suggest that the D-V border is necessary to maintain Tbx2 expression in underlying mesoderm. To corroborate this conclusion, we examined the mesodermal expression of Tbx2 at 0, 2, 6, and 10 hours after removal of the D-V border ectoderm. While we expect the D-V border ectoderm to reconstitute after removal, we did observe a transitory downregulation of Tbx2 occurring maximally at 6 hours after removal of the overlying D-V border ectoderm (n=5) (Fig. 3G). By TUNEL analysis, we confirmed that this downregulation of Tbx2 was not a result of apoptosis. The transitory downregulation of Tbx2 following removal of the overlying D-V ectoderm border is consistent with the specific requirement of the D-V border ectoderm to maintain mesodermal Tbx2 expression.
Fig. 3.
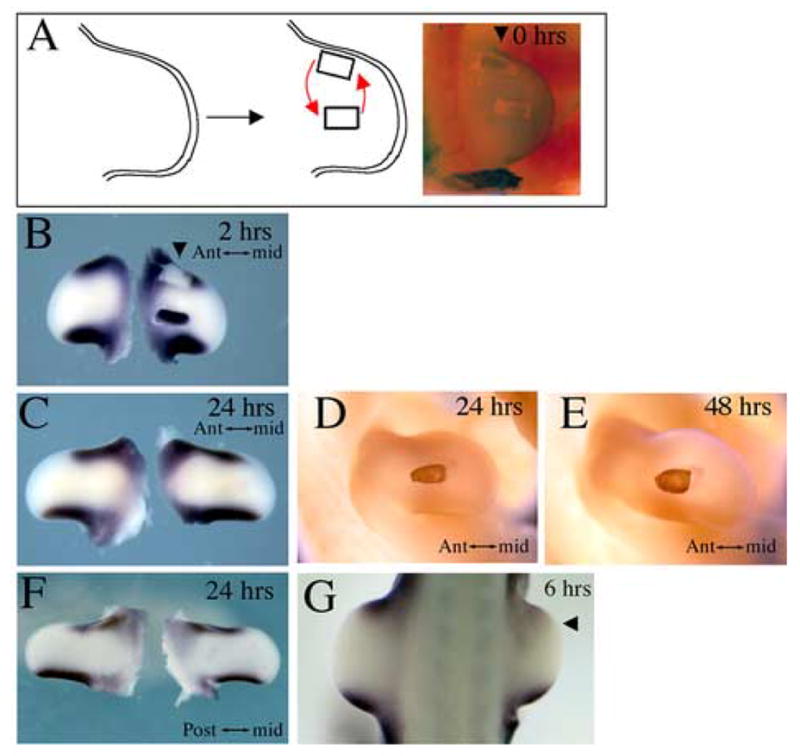
Maintenance of Tbx2 expression in the limb margin mesoderm requires proximity to the D-V ectoderm border. (A) Schematic representation and photograph of operated limb immediately after mesoderm swap in which mesoderm expressing Tbx2 is swapped with mesoderm from mid-limb not expressing Tbx2. (B) Expression of Tbx2 2 hours later confirms mesoderm swap. Note that the anterior D-V ectoderm border remains intact (arrowhead). (C) After 24 hours, mesoderm moved to mid-limb has lost its Tbx2 expression and mesoderm moved to anterior margin has gained Tbx2 expression. (D,E) A graft of quail anterior margin mesoderm to the chick mid-limb after 24 or 48 hours demonstrates that disappearance of Tbx2 in such a graft (C) is not due to loss of the graft. (F) After 24 hours, mesoderm moved from the posterior to the mid-limb has lost its Tbx2 expression and mesoderm moved to the posterior margin has gained Tbx2 expression. (G) Removal of the anterior D-V border ectoderm at stage 19 results in transitory downregulation of Tbx2, maximally observed at 6 hours after removal of the DV border ectoderm.
In our experiments, the mesodermal grafts also included a reciprocal translocation of a block of tissue that does not express Tbx2 from the mid-limb to a position under the D-V border ectoderm (Fig. 3A,B). After 24 hours, this tissue appears to upregulate Tbx2 expression, suggesting that the D-V border ectoderm is sufficient to induce Tbx2 in underlying mesoderm (n=4/9) (Fig. 3C). We also observed upregulation of Tbx2 when mid-limb tissue was placed in proximity of the posterior D-V border ectoderm (n=3/3) (Fig. 3F). In principle, the upregulation of Tbx2 in mesoderm transplanted to the anterior margin could be explained by a combination of signals from the dorsal and ventral ectoderm or by unique signals emanating from the ectoderm at the dorsal-ventral border. To test the former possibility, we transplanted a patch of ventral ectoderm onto the dorsal surface, creating new regions of dorsal-ventral ectodermal juxtaposition over the central limb bud. Although the ectoderm healed and the transplants remained in place, we never saw upregulation of Tbx2 in the underlying mesoderm (data not shown) (n=0/10). In control experiments, we also never saw ectopic AER induction following dorsal-ventral ectodermal juxtaposition at HH stage 22 (data not shown), consistent with previous reports that the ability to induce ectopic AERs in this manner is lost after HH stage 17 (Tanaka et al., 1997).
To test the alternative possibility that a unique Tbx2-inductive signal is produced by D-V border ectoderm, we grafted D-V ectoderm border from the anterior margin of one limb into an ectodermal gap cut in the dorsal surface of another limb at the level of the mid-limb where Tbx2 is not normally expressed (Fig. 4A). We then examined whether this ectodermal border graft could induce Tbx2 expression in the mid-limb. Indeed, when analyzed after 24 or 48 hours, a clear ectopic band of Tbx2 expression could be detected in the mid-limb (n=6/22) (Fig. 4B and data not shown). This ectopic band of Tbx2 was observed with equivalent grafts of the D-V ectoderm border from the posterior margin as well (n=3/9, data not shown). The placement of the D-V ectoderm border on the host limb bud was somewhat variable, and indicated that virtually the entire limb bud mesenchyme is competent to respond to a D-V ectoderm border graft by inducing Tbx2 expression. To rigorously demonstrate that a graft of D-V ectoderm border can non-autonomously induce Tbx2 in underlying mesoderm, and that no marginal mesoderm was transplanted along with the ectodermal graft. We repeated the procedure grafting the anterior D-V ectoderm border from quail onto a chick mid-limb and visualized both grafted quail tissue and Tbx2 expression (Fig. 4C–F). Indeed, no contaminating quail mesoderm is observed and the quail ectoderm graft can be seen overlying Tbx2 induced in chick mesoderm. These data demonstrate that the D-V ectoderm border is sufficient to induce Tbx2 in underlying mesoderm and establish the non-AER dorsal-ventral ectoderm border as a new signaling center in the limb.
Fig. 4.
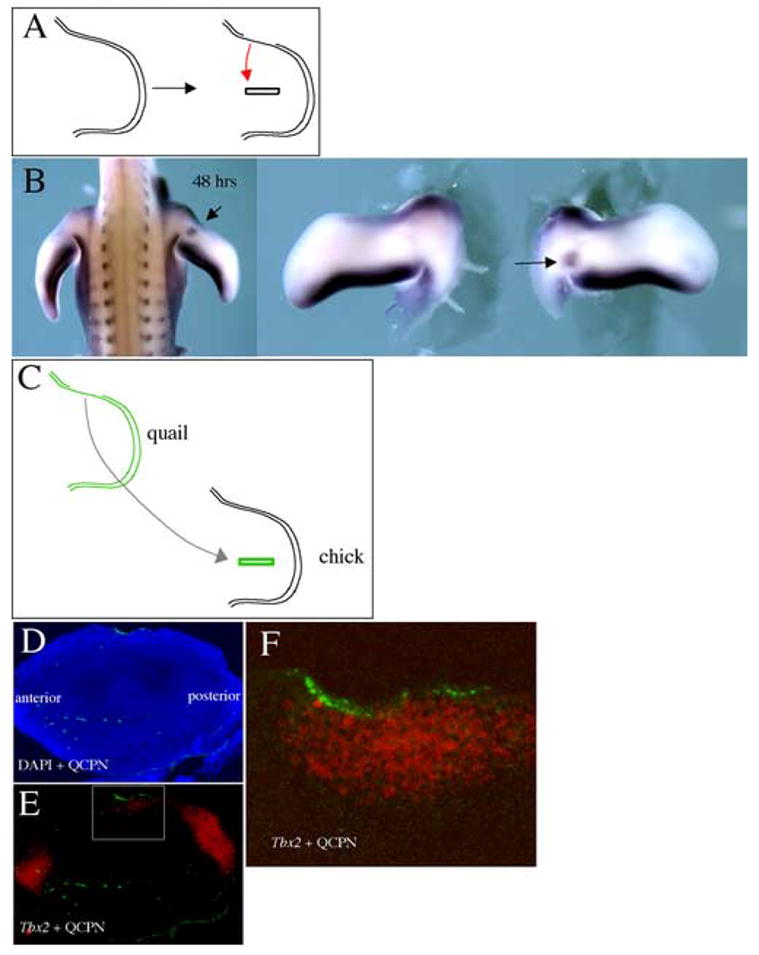
The D-V ectoderm border grafted to the mid-limb is sufficient to induce ectopic Tbx2 expression. (A) Schematic representation of the graft. (B) Tbx2 expression 48 hours after the graft. An ectopic domain of Tbx2 can be seen in the mid-limb. (C) Schematic representation of quail anterior margin ectoderm grafted to a hole cut in the ectoderm of the chick mid-limb. (D,E) After 24 hrs, serial sections showing quail tissue (green) + DAPI nuclear stain (blue) (D) and quail tissue (green) + Tbx2 expression (red) (E). (F) Higher magnification of inset in (E) showing quail ectoderm graft (green) + Tbx2 expression (red) in underlying mesoderm.
In the flank, Tbx2 is also expressed along the D-V ectoderm border before and after limb outgrowth (see Fig. 1). This expression is consistent with a role for the D-V ectoderm border in regulating Tbx2 expression in the flank as well as in the limb. Because no morphologic landmarks visibly demarcate the D-V ectoderm border in the flank as in the limb bud, we did not test this possibility directly.
Regulation of Shh by D-V Ectoderm Border
Previous studies have demonstrated that the Shh expression domain is regulated by Tbx2. When Tbx2 is virally misexpressed in the limb, the domain of Shh expression is expanded anteriorly (Suzuki et al., 2004). This ectopic expansion of Shh did not extend across the entire limb bud, perhaps limited by other posteriorly restricted factors required for Shh expression such as Hand2. Our data shows that the D-V ectoderm border regulates Tbx2 at the limb margins, and a graft of the D-V ectoderm border is sufficient to induce ectopic Tbx2. Put together, these findings predict that a graft of D-V ectoderm border in the posterior limb bud should induce Shh expression in response to the ectopic Tbx2 within the posterior domain where Hand2 and other posteriorly acting factors are expressed.
To test this prediction, we grafted the D-V border ectoderm from the anterior margin of one limb to the dorsal surface of another limb at a position just anterior to the ZPA and monitored Shh expression (Fig. 5A). After 24 hours, we found that an ectopic domain of Shh expression was indeed induced (n=4/13) (Fig. 5B,C). This result supports a model that a signal from the D-V ectoderm border regulates the expression of Shh at the posterior margin. Furthermore, the previous observation that misexpression of Tbx2 expands expression of Shh (Suzuki et al., 2004) is consistent with the possibility that the effect of the D-V ectoderm border on Shh is mediated through expansion of the Tbx2 domain, although a direct response of Shh to the non-AER dorsal-ventral ectodermal border signal is also possible.
Fig. 5.
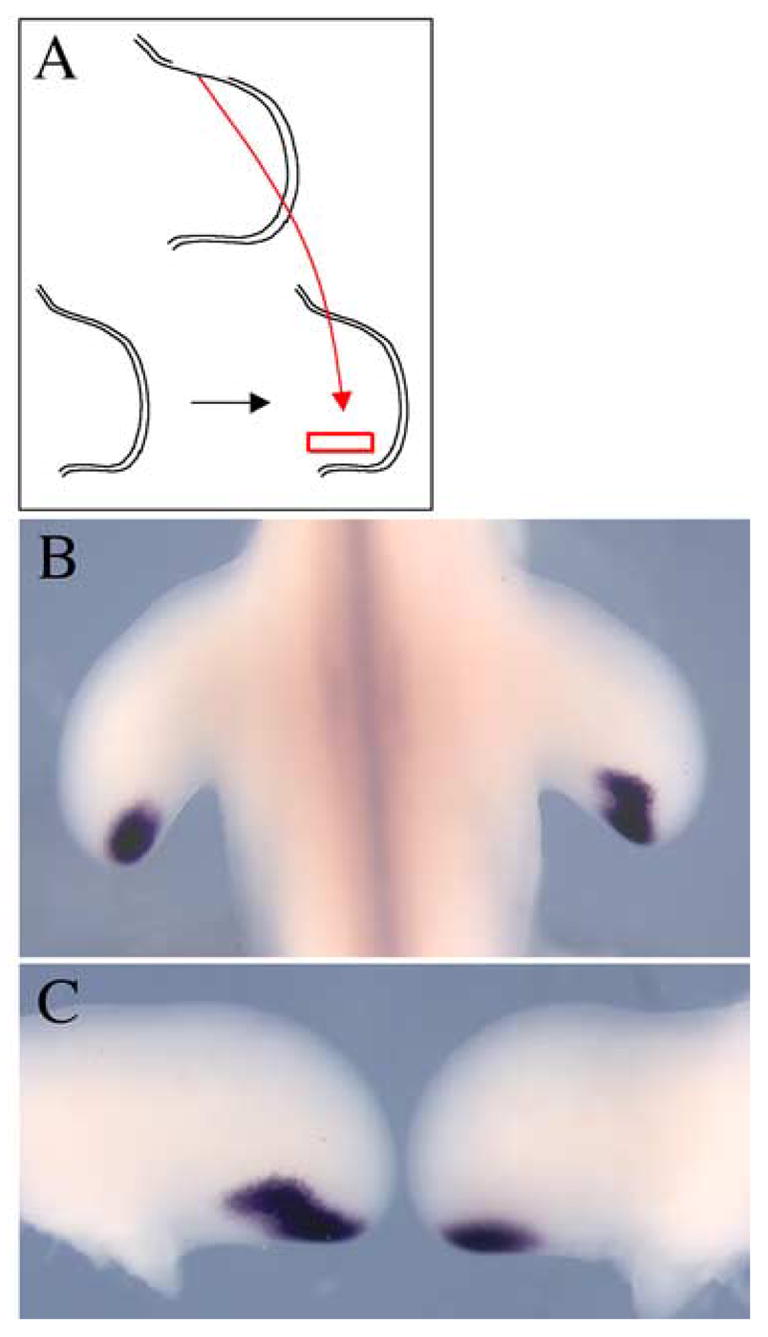
Anterior margin D-V ectoderm border is sufficient to expand Shh expression domain anteriorly. (A) Schematic representation of graft in which anterior margin ectoderm is grafted slightly anterior to the ZPA. (B,C) Dorsal view of contralateral limbs showing anterior expansion of Shh expression in right operated limb.
The AER and Non-AER D-V Ectoderm Border
We have identified the ectoderm at the dorsal-ventral border along the anterior and posterior margins of the limb bud as a new signaling center regulating Tbx gene expression. It is striking, however, that while Tbx2 is expressed along the entire anterior and posterior edges of the limb bud, it is completely absent from the distal mesenchyme, which also abuts a dorsal ventral ectodermal border, the AER. Indeed, direct comparison of mesodermal expression of Tbx2 to expression of Fgf8, a marker of the AER shows that the domains are mutually exclusive in HH21 and HH23 limbs (Fig. 6A–D). The distal extent of Tbx2 approaches the proximal extent of Fgf8 in the AER, but is never found in mesenchyme under the AER. We also previously noted that mesodermal expression of Tbx2 begins to be excluded from the distal tip as the limb bud emerges and the AER forms (see Fig. 1).
Fig. 6.
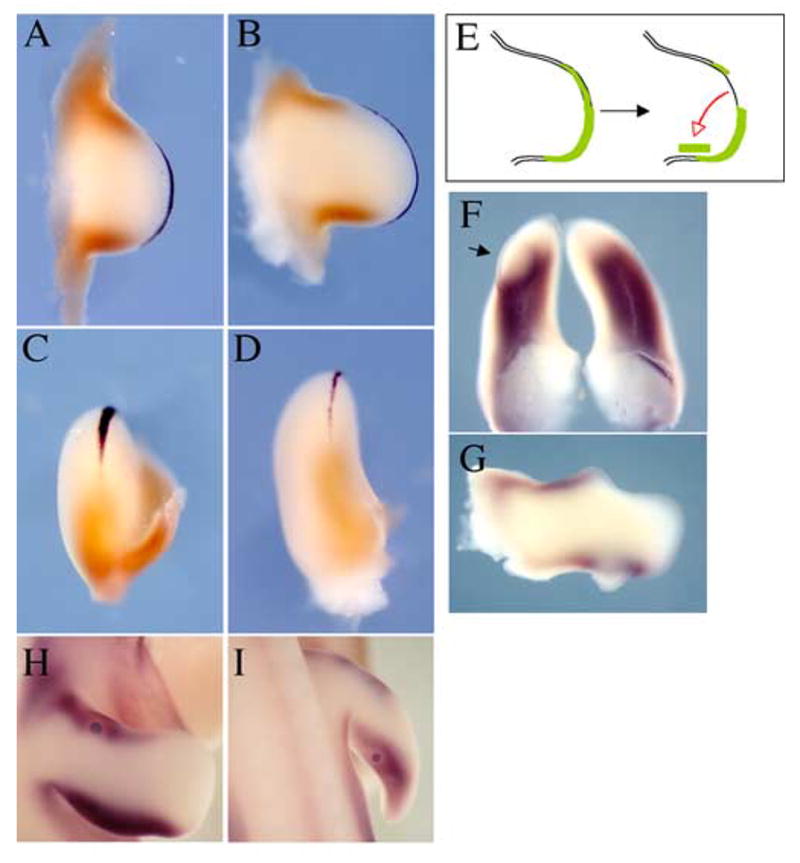
Tbx2 mesodermal expression domain in relation to AER. (A–D) Tbx2 expression (brown) and Fgf8 expression (black) at HH stage 21 (A,C) and stage 23 (B,D). Dorsal views (A,B) and posterior views (C,D) show that Tbx2 expression abuts but does not extend under AER. (E–G) A graft of AER to the dorsal posterior surface causes ectopic proliferation and downregulation of Tbx2 in underlying mesenchyme. A notch in the growth of the limb is apparent in the anterior limb where the AER was removed (G). A bead soaked in Fgf8 downregulates expression of Tbx2 around the bead when implanted in the anterior (H) or posterior (I) margin of the limb.
These observations are consistent with the dorsal-ventral border ectodermal signal, which we have shown is both able to induce Tbx2 and is required for its maintenance, being turned off as the distinct AER signaling center is formed. It remains plausible, however, that mesodermal expression of Tbx2 is also actively excluded from subapical mesoderm by a signal from the AER. To test this latter possibility, we grafted portions of the AER onto the dorsal-posterior surface of the limb at HH stage 22 and examined if this graft of AER changed the underlying Tbx2 expression domain (Fig. 6E). As expected, the graft of AER induced ectopic outgrowth of the underlying mesoderm at the posterior edge of the limb (arrow, Fig. 6F, G), while a notch in the growth of the limb bud was evident anteriorly where the AER graft had been removed (Fig. 6G). Expression of Tbx2 in the posterior margin within and around the ectopic outgrowth was downregulated (n = 5). These data support the possibility that a signal from the AER actively inhibits expression of Tbx2 in underlying mesenchyme. Members of the Fgf family are obvious candidates for such a signal emanating from the AER as several members of the Fgf family are specifically expressed in the AER and can substitute for the AER if it is surgically extirpated (Niswander, 1998; Fallon, 1999). To test this possibility, we implanted beads soaked in Fgf8, normally expressed throughout the AER, into the anterior or posterior margin mesoderm of HH stage 22 limbs and examined for changes in the expression of Tbx2 after 24 hours. We observed that Tbx2 expression was downregulated immediately around the Fgf8 bead (Fig. 6H, I; n = 7/7 anterior, n=7/7 posterior). This suggests that Fgf8 secreted from the AER excludes Tbx2 expression from subapical mesoderm.
DISCUSSION
D-V Ectodermal Border as Signaling Center
Classical studies and more recent molecular analyses have identified the AER as a signaling center necessary for proper outgrowth of the limb. The AER coincides with the dorsal-ventral ectodermal border at the tip of the limb. The data presented in this study indicate that the dorsal-ventral border ectoderm proximal to the AER also serves as a signaling center, regulating expression of genes restricted to the anterior and posterior margins of the limb.
Several lines of evidence support this model. The spatial expression pattern of Tbx2 very closely relates to the non-AER D-V border ectoderm over a wide range of limb stages. When we moved mesoderm expressing Tbx2 away from the D-V border ectoderm Tbx2 expression was lost after 24 hours, suggesting that proximity to the D-V border ectoderm is necessary to maintain mesodermal expression of Tbx2. Furthermore, grafts of the D-V border ectoderm from both the anterior and posterior margins were capable of inducing ectopic Tbx2 in underlying mesoderm. This non-autonomous effect suggests that a signal from the non-AER D-V border ectoderm is sufficient to upregulate Tbx2 in underlying mesoderm. The closely related transcription factor Tbx3 is also expressed along the anterior and posterior limb margins, though in slightly larger domains. This suggests that Tbx3 may also serve as a readout of signaling from the D-V border, but may require a lower level of this signal than Tbx2.
While the AER signaling center is characterized by distinct stratified (in the mouse) or pseudostratified (in the chick) epithelium, the non-AER border ectoderm is morphologically indistinguishable from adjacent dorsal and ventral ectoderm. This may partially explain why this signaling center escaped notice until now.
Regulation of Tbx2 Gene Expression
Our data suggest that Tbx2 is regulated by the adjacent dorsal-ventral ectoderm border. A previous study has found that a bead soaked in Shh and placed in the mid-limb can expand the posterior domain of Tbx3, whereas a Shh bead placed in the anterior can cause downregulation of Tbx3 (Tumpel et al., 2002). These observations were interpreted as evidence that the anterior and posterior domains of Tbx3 are established by signals patterned along the anterior-posterior axis (Tumpel et al., 2002). This interpretation seems incomplete, given that Tbx2 and Tbx3 are maintained at the anterior and posterior margins all the way to the flank at considerable distances from the ZPA, even as late as HH stage 27 when Shh expression is quite distal. These anterior and posterior limb expression domains are continuous with flank expression, where regulation by Shh is very unlikely. Moreover, we have shown that Tbx2 expression is not lost from the margins 8 hours after blocking Shh activity nor in the context of limbs deficient in mesodermal expression of BMP2 and BMP7, suggesting that normal Tbx2 expression is not directly regulated by signals which pattern the anterior-posterior axis. Conversely, there is considerable evidence that Tbx2/3 act upstream of Shh in the posterior margin of the limb. A mouse carrying a targeted deletion of Tbx3 exhibits loss or absence of Shh (Davenport et al., 2003), and viral misexpression studies demonstrate that Tbx2 is both necessary for normal Shh expression and sufficient to expand Shh expression in the posterior chick limb (Suzuki et al., 2004). Whereas loss-of-function of Tbx2 or Tbx3 in chick limbs and loss-of-function of Tbx3 in humans leads to loss of posterior elements, as would be expected by downregulation of Shh, we note that Tbx2 null mutant mice do not exhibit loss of posterior elements (Harrelson et al., 2004). Instead, these Tbx2 mutant mice have a hindlimb-specific duplication of the most distal phalanx of digit IV at E14.5, and this likely reflects the interphalangeal expression of Tbx2 where it may be involved in apoptosis. The lack of any effect on posterior patterning in Tbx2 mutant mice may reflect redundancy with Tbx3.
The previous finding that a bead soaked in Shh and placed in the mid-limb can expand the posterior domain of Tbx3 may be explained by a positive feedback interaction between Shh and Tbx2/3. It is possible that while Tbx2/3 act upstream of Shh, Shh in turn can positively regulate Tbx2/3. Indeed, a pellet of cells expressing Shh, when implanted in the anterior limb from which the anterior AER has been removed, has been shown to upregulate Tbx2 around the pellet (Gibson-Brown et al., 1998). A similar positive feedback loop has been demonstrated between Shh and Hand2, another transcription factor that is believed to regulate Shh expression in the posterior. The previous observation that ectopic Shh can down-regulate Tbx3 in the anterior requires a different explanation. It has previously been demonstrated that ectopic placement of Shh in the anterior margin causes the AER to extend anteriorly (Laufer et al., 1994). In the current study, we have shown that Tbx2 appears to be excluded from subapical mesoderm over a wide range of limb development (HH stage 19 to 27), and this pattern appears to be true for Tbx3 as well (Tumpel et al., 2002), suggesting that the AER may negatively regulate Tbx2/3 in subapical mesoderm and/or that the D-V border ectoderm signals required to maintain Tbx2/3 are excluded from the AER (discussed below). We therefore speculate that the downregulation of Tbx3 observed when a Shh bead is placed in the anterior margin may reflect expansion of the AER over the Shh bead and consequent downregulation of Tbx3.
The signal from the D-V ectoderm border that regulates mesodermal Tbx2 expression is currently unknown. While not a homologous structure, an interesting parallel exists in the regulation of the orthologous Drosophila optomotor blind (omb) gene in the wing disc. In the early wing disc, omb is expressed throughout the entire presumptive wing domain (del Alamo Rodriguez et al., 2004; Grimm and Pflugfelder, 1996). However, as the wing disc develops, omb disappears from the lateral-most regions of the disc and is most highly expressed in the central region of the disc (del Alamo Rodriguez et al., 2004). Strikingly, in this central region, omb becomes restricted to 3 domains: a stripe at the wing margin demarcating, as in the vertebrate limb ectoderm, the dorsal-ventral border of the wing disc and along 2 rings that surround the presumptive wing hinge and delimit the wing region (del Alamo Rodriguez et al., 2004). These 3 domains are marked by expression of Wingless, the Drosophila ortholog of the Wnt family. While Wnt3a is specifically expressed in the AER, no Wnt genes have yet been identified specifically in the non-AER D-V border ectoderm (Kengaku et al., 1998). How the signal from the D-V ectoderm border acts to regulate Tbx2 expression also remains to be determined.
AER vs. non-AER D-V Border Ectoderm
From the time the limb bud emerges until the time digit rays begin to form, we observed that Tbx2 is excluded from mesoderm under the AER. Interestingly, the appearance of Tbx2 and Tbx3 in the interdigital mesoderm at HH stage 27 correlates with degeneration of the AER overlying the interdigital mesoderm (Ganan et al., 1998). Furthermore, by using Tbx2 expression as a readout of signaling from the D-V border ectoderm and Fgf8 as a marker of the AER, we found that the domain of Tbx2 approaches the AER but does not extend underneath it. This data suggests a compartmentalization of the D-V border ectoderm in which the AER specifically excludes Tbx2 in underlying mesoderm while the non-AER D-V ectoderm border specifically upregulates Tbx2 in underlying mesoderm. This model was supported by the finding that grafting an AER next to the non-AER border ectoderm resulted in a downregulation of underlying Tbx2. Indeed, we show that Fgf8, normally secreted from the AER, is capable of downregulating mesodermal Tbx2. It is also known that Fgfs from the AER are required to maintain Shh expression in the ZPA. Put together, these data suggest that high levels of Fgf8 immediately under the AER (and immediately around an Fgf8 bead) repress Tbx2 expression, while lower levels of Fgfs are sufficient to maintain Shh and do not act to downregulate the distal-most portion of the Tbx2 domain along the posterior limb margin.
Fate mapping studies of the AER reveal that cells in the posterior 2/3 of the AER remain within the AER after 2 days but cells in the anterior 1/3 AER move out to the anterior margin ectoderm just proximal to the AER (Vargesson et al., 1997). Thus, it will be interesting to elucidate how distinct signaling from the AER vs. non-AER D-V border ectoderm is maintained as the distal border ectoderm first converts to an AER during limb bud initiation and then converts back to non-AER D-V border in the anterior.
The AER D-V ectodermal border manifests as a conspicuous ridge at the distal tip of the limb. The AER D-V ectodermal border is also characterized by gene expression boundaries and cell lineage boundaries which have functional significance in proper AER initiation and maintenance. For example, cells expressing Engrailed1 (En1) contribute to the ventral ectoderm and ventral half of the AER (Kimmel et al., 2000), confirming a lineage compartment boundary in the mid-AER (Altabef et al., 1997). When this lineage boundary is disrupted by misexpressing En1 in the dorsal AER half, the AER fails to form properly (Kimmel et al., 2000). It will therefore be interesting to characterize whether the non-AER D-V ectodermal border also has gene expression and cell lineage boundaries that are critical to its function.
Regulation of Shh at the Limb Margin
Placed in the context of prior work on various factors regulating Shh expression in the limb, our results allow us to formulate a model explaining the spatial control of Shh expression, and hence the localization of polarizing activity during limb development (Fig. 7). Prior to limb outgrowth or the expression of Shh, opposing gradients of negatively acting factors such as Gli3 and positively acting factors such as Hand2 (te Welscher et al., 2002) result in a posterior quadrant of the limb bud competent to express Shh. Of course, Shh regulation is complex and also requires the activity of Hox genes (Kmita et al., 2005). Signals from the non-AER D-V border ectoderm induce expression of Tbx2, required for Shh expression, along both the anterior and posterior margins of the limb mesoderm. However, Shh is only induced within the posterior margin due to the competence of that mesoderm conferred by Hand2 expression. Indeed, the timing of ectopic Shh expression in response to a graft of the D-V border ectoderm (Fig. 5) is consistent with a requirement for Tbx2 expression 24 hours after placement of the graft, and the location of ectopic Shh expression is presumably limited to the posterior by the domain of Hand2 expression. In addition, Shh expression requires Fgf signaling, provided from the distal tip as the distal dorsal-ventral ectoderm border is converted to the AER turning on Fgf8 and other Fgf family members at the expense of the non-AER border signal. As the limb bud grows out, the AER, and hence the source of Fgf signaling, remains at the distal tip, thus assuring that only the most distal Tbx2-expressing domain continues to express Shh.
Fig. 7.
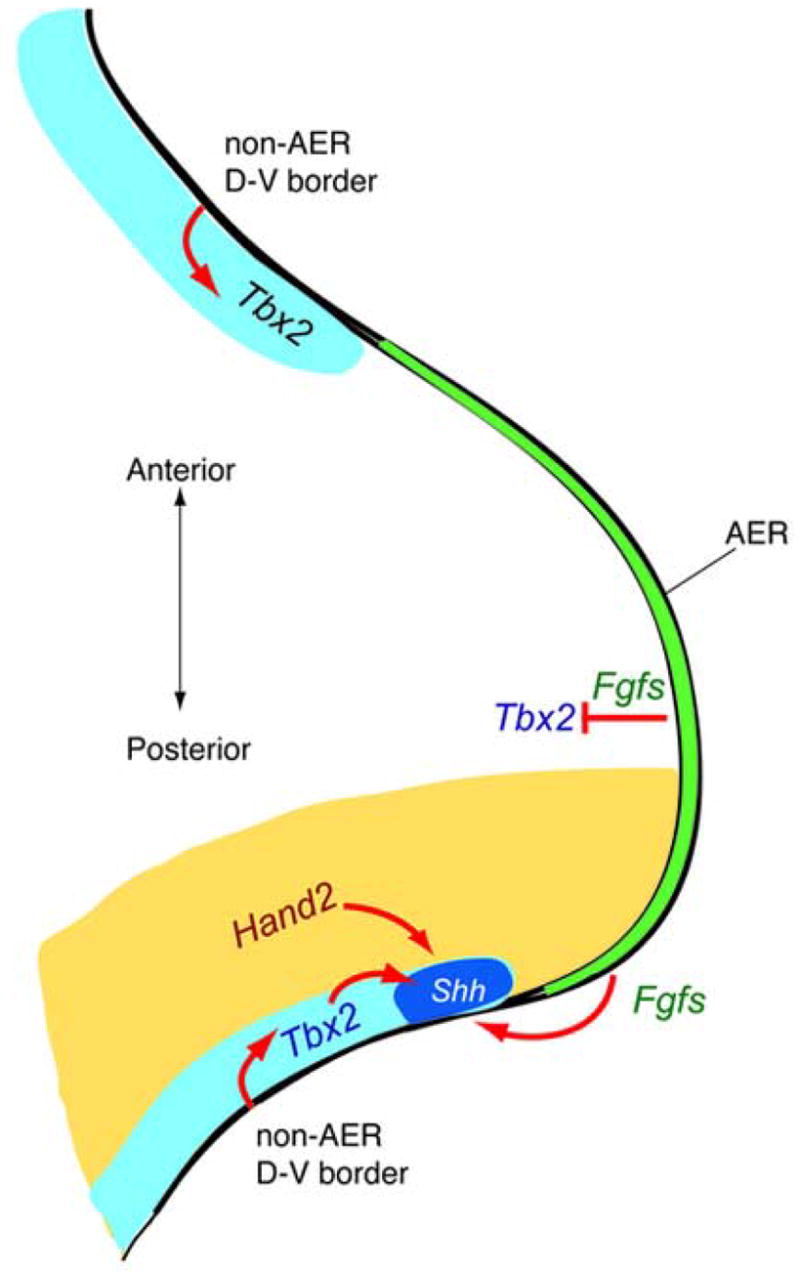
Model for non-AER D-V ectoderm border function as a signaling center that localizes the ZPA to the limb margin. Expression of Tbx2 serves as a readout for signaling from non-AER D-V border at the anterior and posterior limb margins. Signaling from non-AER D-V border restricts Shh expression to posterior margin, possibly via requirement of Tbx2 for proper Shh expression. Other regulatory factors collaborate with the non-AER D-V border signal in positioning the ZPA. For example, Hand2 restricts Shh to the posterior and not anterior margin, and Fgfs from the AER restrict Shh to the distal end of the posterior margin.
In support of this model, we find that the non-AER dorsal-ventral border ectoderm is both necessary and sufficient for the induction of Tbx2 expression and is also capable of inducing Shh expression in the posterior limb mesenchyme. Moreover, previous studies have shown that viral misexpression of Tbx2 expands Shh expression anteriorly (Suzuki et al., 2004). We noted, however, that the normal Tbx2 expression domain extends slightly more anteriorly than the Shh domain. This could reflect a threshold requirement for levels of Tbx2 gene expression not achieved at the edge of their expression domain. Alternatively, it could reflect an independent, direct requirement for the non-AER dorsal-ventral border ectoderm signal. In that case, Shh expression would be predicted to require a higher threshold level of that signal than Tbx2.
In addition to explaining the localization of Shh expression in the posterior-most distal mesenchyme, this model also provides an explanation for the limited ectopic expression of Shh at the extreme anterior limb margin seen in many mutants leading to preaxial polydactyly. When negative-acting factors are lost, the transcription factors providing competence for Shh expression, such as Hand2, can expand their expression across the entire limb bud mesenchyme (te Welscher et al., 2002). In this situation, the cells subjacent to the anteriornon-AER dorsal-ventral ectoderm border are able to respond to the signal within the domain marked by anterior Tbx2 expression, and turn on an ectopic domain of Shh expression. In addition to the AER and the non-AER dorsal-ventral border ectoderm, there is a third ectodermal signaling center in the limb bud; the entire dorsal ectodermal surface. The dorsal ectoderm plays an essential role in establishing the dorsal-ventral polarity of the limb bud (MacCabe et al., 1974; Pautou, 1977; Geduspan and MacCabe, 1987, 1989). This activity is mediated by Wnt7a (Parr and McMahon, 1995); and interestingly Wnt7a also plays a role in supporting high levels of Shh expression in the ZPA (Parr and McMahon, 1995; Yang and Niswander, 1995). As a result, there is a dorsal bias in the expression domain of Shh within the ZPA, although the importance of this for limb patterning remains unclear.
In contrast, the restriction of Shh mRNA to the posterior margin is critical in establishing a proper spatial and temporal gradient of Shh activity. The cells initially expressing Shh proliferate and expand disproportionately, such that they end up encompassing most of the posterior half of the distal limb bud. As they do so, those cells that are forced further away from the posterior margin shut off Shh such that the domain of cells that continue to express Shh remains relatively quite small, although all these cells presumably are competent to express Shh (Harfe et al., 2004). Our current results explain this, as only the cells remaining along the extreme posterior margin during this expansion will still be within range of the non-AER dorsal-ventral border signal and hence only in those cells will Tbx2, Tbx3 and Shh expression be maintained.
Acknowledgments
We thank Paul Scherz for extremely helpful discussions of the models presented in this study. This work was supported by a grant, 2 R37 HD032443, from the NIH to CJT.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Bamshad M, Lin RC, Law DJ, Watkins WC, Krakowiak PA, Moore ME, Franceschini P, Lala R, Holmes LB, Gebuhr TC, et al. Mutations in human TBX3 alter limb, apocrine and genital development in ulnar-mammary syndrome. Nat Genet. 1997;16:311–5. doi: 10.1038/ng0797-311. [DOI] [PubMed] [Google Scholar]
- Charite J, de Graaff W, Shen S, Deschamps J. Ectopic expression of Hoxb-8 causes duplication of the ZPA in the forelimb and homeotic transformation of axial structures. Cell. 1994;78:589–601. doi: 10.1016/0092-8674(94)90524-x. [DOI] [PubMed] [Google Scholar]
- Charite J, McFadden DG, Olson EN. The bHLH transcription factor dHAND controls Sonic hedgehog expression and establishment of the zone of polarizing activity during limb development. Development. 2000;127:2461–70. doi: 10.1242/dev.127.11.2461. [DOI] [PubMed] [Google Scholar]
- Chen JK, Taipale J, Cooper MK, Beachy PA. Inhibition of Hedgehog signaling by direct binding of cyclopamine to Smoothened. Genes Dev. 2002;16:2743–8. doi: 10.1101/gad.1025302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davenport TG, Jerome-Majewska LA, Papaioannou VE. Mammary gland, limb and yolk sac defects in mice lacking Tbx3, the gene mutated in human ulnar mammary syndrome. Development. 2003;130:2263–73. doi: 10.1242/dev.00431. [DOI] [PubMed] [Google Scholar]
- Dealy CN, Roth A, Ferrari D, Brown AM, Kosher RA. Wnt-5a and Wnt-7a are expressed in the developing chick limb bud in a manner suggesting roles in pattern formation along the proximodistal and dorsoventral axes. Mech Dev. 1993;43:175–86. doi: 10.1016/0925-4773(93)90034-u. [DOI] [PubMed] [Google Scholar]
- del Alamo Rodriguez D, Terriente Felix J, Diaz-Benjumea FJ. The role of the T-box gene optomotor-blind in patterning the Drosophila wing. Dev Biol. 2004;268:481–92. doi: 10.1016/j.ydbio.2004.01.005. [DOI] [PubMed] [Google Scholar]
- Dietrich S, Schubert FR, Lumsden A. Control of dorsoventral pattern in the chick paraxial mesoderm. Development. 1997;124:3895–908. doi: 10.1242/dev.124.19.3895. [DOI] [PubMed] [Google Scholar]
- Fernandez-Teran M, Piedra ME, Kathiriya IS, Srivastava D, Rodriguez-Rey JC, Ros MA. Role of dHAND in the anterior-posterior polarization of the limb bud: implications for the Sonic hedgehog pathway. Development. 2000;127:2133–42. doi: 10.1242/dev.127.10.2133. [DOI] [PubMed] [Google Scholar]
- Ganan Y, Macias D, Basco RD, Merino R, Hurle JM. Morphological diversity of the avian foot is related with the pattern of msx gene expression in the developing autopod. Dev Biol. 1998;196:33–41. doi: 10.1006/dbio.1997.8843. [DOI] [PubMed] [Google Scholar]
- Geduspan JS, MacCabe JA. The ectodermal control of mesodermal patterns of differentiation in the developing chick wing. Dev Biol. 1987;124:398–408. doi: 10.1016/0012-1606(87)90492-1. [DOI] [PubMed] [Google Scholar]
- Geduspan JS, MacCabe JA. Transfer of dorsoventral information from mesoderm to ectoderm at the onset of limb development. Anat Rec. 1989;224:79–87. doi: 10.1002/ar.1092240110. [DOI] [PubMed] [Google Scholar]
- Gibson-Brown JJ, S IA, Silver LM, Papaioannou VE. Expression of T-box genes Tbx2-Tbx5 during chick organogenesis. Mech Dev. 1998;74:165–9. doi: 10.1016/s0925-4773(98)00056-2. [DOI] [PubMed] [Google Scholar]
- Grimm S, Pflugfelder GO. Control of the gene optomotor-blind in Drosophila wing development by decapentaplegic and wingless. Science. 1996;271:1601–4. doi: 10.1126/science.271.5255.1601. [DOI] [PubMed] [Google Scholar]
- Hamburger V, Hamilton HL. A series of normal stages in the development of the chick embryo. J of Morphol. 1951;88:49–92. [PubMed] [Google Scholar]
- Harrelson Z, Kelly RG, Goldin SN, Gibson-Brown JJ, Bollag RJ, Silver LM, Papaioannou VE. Tbx2 is essential for patterning the atrioventricular canal and for morphogenesis of the outflow tract during heart development. Development. 2004;131:5041–52. doi: 10.1242/dev.01378. [DOI] [PubMed] [Google Scholar]
- Harfe BD, Scherz PJ, Nissim S, Tian H, McMahon AP, Tabin CJ. Evidence for an expansion-based temporal Shh gradient in specifying vertebrate digit identities. Cell. 2004;118:517–28. doi: 10.1016/j.cell.2004.07.024. [DOI] [PubMed] [Google Scholar]
- Hui CC, Joyner AL. A mouse model of greig cephalopolysyndactyly syndrome: the extra-toesJ mutation contains an intragenic deletion of the Gli3 gene. Nat Genet. 1993;3:241–6. doi: 10.1038/ng0393-241. [DOI] [PubMed] [Google Scholar]
- Incardona JP, Gaffield W, Kapur RP, Roelink H. The teratogenic Veratrum alkaloid cyclopamine inhibits sonic hedgehog signal transduction. Development. 1998;125:3553–62. doi: 10.1242/dev.125.18.3553. [DOI] [PubMed] [Google Scholar]
- Isaac A, Rodriguez-Esteban C, Ryan A, Altabef M, Tsukui T, Patel K, Tickle C, Izpisua-Belmonte JC. Tbx genes and limb identity in chick embryo development. Development. 1998;125:1867–75. doi: 10.1242/dev.125.10.1867. [DOI] [PubMed] [Google Scholar]
- Kengaku M, Capdevila J, Rodriguez-Esteban C, De La Pena J, Johnson RL, Belmonte JC, Tabin CJ. Distinct WNT pathways regulating AER formation and dorsoventral polarity in the chick limb bud. Science. 1998;280:1274–7. doi: 10.1126/science.280.5367.1274. [DOI] [PubMed] [Google Scholar]
- Kimmel RA, Turnbull DH, Blanquet V, Wurst W, Loomis CA, Joyner AL. Two lineage boundaries coordinate vertebrate apical ectodermal ridge formation. Genes Dev. 2000;14:1377–89. [PMC free article] [PubMed] [Google Scholar]
- Kmita M, Tarchini B, Zakany J, Logan M, Tabin CJ, Duboule D. Early developmental arrest of mammalian limbs lacking HoxA/HoxD gene function. Nature. 2005;435:1113–6. doi: 10.1038/nature03648. [DOI] [PubMed] [Google Scholar]
- Laufer E, Nelson CE, Johnson RL, Morgan BA, Tabin C. Sonic hedgehog and Fgf-4 act through a signaling cascade and feedback loop to integrate growth and patterning of the developing limb bud. Cell. 1994;79:993–1003. doi: 10.1016/0092-8674(94)90030-2. [DOI] [PubMed] [Google Scholar]
- Lewis PM, Dunn MP, McMahon JA, Logan M, Martin JF, St-Jacques B, McMahon AP. Cholesterol modification of sonic hedgehog is required for long-range signaling activity and effective modulation of signaling by Ptc1. Cell. 2001;105:599–612. doi: 10.1016/s0092-8674(01)00369-5. [DOI] [PubMed] [Google Scholar]
- Logan M, Simon HG, Tabin C. Differential regulation of T-box and homeobox transcription factors suggests roles in controlling chick limb-type identity. Development. 1998;125:2825–35. doi: 10.1242/dev.125.15.2825. [DOI] [PubMed] [Google Scholar]
- Lopez-Martinez A, Chang DT, Chiang C, Porter JA, Ros MA, Simandl BK, Beachy PA, Fallon JF. Limb-patterning activity and restricted posterior localization of the amino-terminal product of Sonic hedgehog cleavage. Curr Biol. 1995;5:791–6. doi: 10.1016/s0960-9822(95)00156-4. [DOI] [PubMed] [Google Scholar]
- Lyons KM, Pelton RW, Hogan BL. Organogenesis and pattern formation in the mouse: RNA distribution patterns suggest a role for bone morphogenetic protein-2A (BMP-2A) Development. 1990;109:833–44. doi: 10.1242/dev.109.4.833. [DOI] [PubMed] [Google Scholar]
- Marigo V, Scott MP, Johnson RL, Goodrich LV, Tabin CJ. Conservation in hedgehog signaling: induction of a chicken patched homolog by Sonic hedgehog in the developing limb. Development. 1996;122:1225–33. doi: 10.1242/dev.122.4.1225. [DOI] [PubMed] [Google Scholar]
- Martin GR. The roles of FGFs in the early development of vertebrate limbs. Genes Dev. 1998;12:1571–86. doi: 10.1101/gad.12.11.1571. [DOI] [PubMed] [Google Scholar]
- Masuya H, Sagai T, Wakana S, Moriwaki K, Shiroishi T. A duplicated zone of polarizing activity in polydactylous mouse mutants. Genes Dev. 1995;9:1645–53. doi: 10.1101/gad.9.13.1645. [DOI] [PubMed] [Google Scholar]
- MacCabe JA, Errick J, Suanders JW., Jr Ectodermal control of the dorsoventral axis in the leg bud of the chick embryo. Dev Biol. 1974;39:69–82. doi: 10.1016/s0012-1606(74)80009-6. [DOI] [PubMed] [Google Scholar]
- Murtaugh LC, Chyung JH, Lassar AB. Sonic hedgehog promotes somitic chondrogenesis by altering the cellular response to BMP signaling. Genes Dev. 1999;13:225–37. doi: 10.1101/gad.13.2.225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niswander L, Jeffrey S, Martin GR, Tickle C. A positive feedback loop coordinates growth and patterning in the vertebrate limb. Nature. 1994;371:609–12. doi: 10.1038/371609a0. [DOI] [PubMed] [Google Scholar]
- Niswander L, Tickle C, Vogel A, Booth I, Martin GR. FGF-4 replaces the apical ectodermal ridge and directs outgrowth and patterning of the limb. Cell. 1993;75:579–87. doi: 10.1016/0092-8674(93)90391-3. [DOI] [PubMed] [Google Scholar]
- Papaioannou VE. T-box genes in development: from hydra to humans. Int Rev Cytol. 2001;207:1–70. doi: 10.1016/s0074-7696(01)07002-4. [DOI] [PubMed] [Google Scholar]
- Papaioannou VE, Silver LM. The T-box gene family. Bioessays. 1998;20:9–19. doi: 10.1002/(SICI)1521-1878(199801)20:1<9::AID-BIES4>3.0.CO;2-Q. [DOI] [PubMed] [Google Scholar]
- Parr BA, McMahon AP. Dorsalizing signal Wnt-7a required for normal polarity of D-V and A-P axes in mouse limb. Nature. 1995;374:350–353. doi: 10.1038/374350a0. [DOI] [PubMed] [Google Scholar]
- Parr BA, Shea MJ, Vassileva G, McMahon AP. Mouse Wnt genes exhibit discrete domains of expression in the early embryonic CNS and limb buds. Development. 1993;119:247–61. doi: 10.1242/dev.119.1.247. [DOI] [PubMed] [Google Scholar]
- Pautou MP. Etablissement de l’axe dorso-ventral dans le pied de l’embryon de poulet. J Embryol Exp Morphol. 1977;42:177–194. [Google Scholar]
- Pizette S, Abate-Shen C, Niswander L. BMP controls proximodistal outgrowth, via induction of the apical ectodermal ridge, and dorsoventral patterning in the vertebrate limb. Development. 2001;128:4463–74. doi: 10.1242/dev.128.22.4463. [DOI] [PubMed] [Google Scholar]
- Qu S, Tucker SC, Ehrlich JS, Levorse JM, Flaherty LA, Wisdom R, Vogt TF. Mutations in mouse Aristaless-like4 cause Strong’s luxoid polydactyly. Development. 1998;125:2711–21. doi: 10.1242/dev.125.14.2711. [DOI] [PubMed] [Google Scholar]
- Riddle RD, Johnson RL, Laufer E, Tabin C. Sonic hedgehog mediates the polarizing activity of the ZPA. Cell. 1993;75:1401–16. doi: 10.1016/0092-8674(93)90626-2. [DOI] [PubMed] [Google Scholar]
- Suzuki T, Takeuchi J, Koshiba-Takeuchi K, Ogura T. Tbx Genes Specify Posterior Digit Identity through Shh and BMP Signaling. Dev Cell. 2004;6:43–53. doi: 10.1016/s1534-5807(03)00401-5. [DOI] [PubMed] [Google Scholar]
- Takahashi M, Tamura K, Buscher D, Masuya H, Yonei-Tamura S, Matsumoto K, Naitoh-Matsuo M, Takeuchi J, Ogura K, Shiroishi T, et al. The role of Alx-4 in the establishment of anteroposterior polarity during vertebrate limb development. Development. 1998;125:4417–25. doi: 10.1242/dev.125.22.4417. [DOI] [PubMed] [Google Scholar]
- Tanaka M, Tamura K, Noji S, Nohno T, Ide H. Induction of additional limb at the dorsal-ventral boundary of a chick embryo. Dev Biol. 1997;182:191–203. doi: 10.1006/dbio.1996.8476. [DOI] [PubMed] [Google Scholar]
- te Welscher P, Fernandez-Teran M, Ros MA, Zeller R. Mutual genetic antagonism involving GLI3 and dHAND prepatterns the vertebrate limb bud mesenchyme prior to SHH signaling. Genes Dev. 2002;16:421–6. doi: 10.1101/gad.219202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tickle C. The number of polarizing region cells required to specify additional digits in the developing chick wing. Nature. 1981;289:295–8. doi: 10.1038/289295a0. [DOI] [PubMed] [Google Scholar]
- Todt WL, Fallon JF. Posterior apical ectodermal ridge removal in the chick wing bud triggers a series of events resulting in defective anterior pattern formation. Development. 1987;101:501–15. doi: 10.1242/dev.101.3.501. [DOI] [PubMed] [Google Scholar]
- Tumpel S, Sanz-Ezquerro JJ, Isaac A, Eblaghie MC, Dobson J, Tickle C. Regulation of Tbx3 expression by anteroposterior signalling in vertebrate limb development. Dev Biol. 2002;250:251–62. [PubMed] [Google Scholar]
- Vargesson N, Clarke JD, Vincent K, Coles C, Wolpert L, Tickle C. Cell fate in the chick limb bud and relationship to gene expression. Development. 1997;124:1909–18. doi: 10.1242/dev.124.10.1909. [DOI] [PubMed] [Google Scholar]
- Vogel A, Tickle C. FGF-4 maintains polarizing activity of posterior limb bud cells in vivo and in vitro. Development. 1993;119:199–206. doi: 10.1242/dev.119.1.199. [DOI] [PubMed] [Google Scholar]
- Wolpert L. Positional information and the spatial pattern of cellular differentiation. J Theor Biol. 1969;25:1–47. doi: 10.1016/s0022-5193(69)80016-0. [DOI] [PubMed] [Google Scholar]
- Yang Y, Drossopoulou G, Chuang PT, Duprez D, Marti E, Bumcrot D, Vargesson N, Clarke J, Niswander L, McMahon A, et al. Relationship between dose, distance and time in Sonic Hedgehog-mediated regulation of anteroposterior polarity in the chick limb. Development. 1997;124:4393–404. doi: 10.1242/dev.124.21.4393. [DOI] [PubMed] [Google Scholar]
- Yang Y, Niswander L. Interaction between the signaling molecules WNT7a and SHH during vertebrate limb development: dorsal signals regulate anteroposterior patterning. Cell. 1995;80:939–47. doi: 10.1016/0092-8674(95)90297-x. [DOI] [PubMed] [Google Scholar]