

Abstract
We report six unsuccessful attempts to replicate the “work ethic” phenomenon reported by Clement, Feltus, Kaiser, and Zentall (2000). In Experiments 1–5, pigeons learned two simultaneous discriminations in which the S+ and S− stimuli were obtained by pecking an initial stimulus once or multiple (20 or 40) times. Subsequent preference tests between the S+ stimuli and between the S− stimuli mostly revealed indifference, on average, between the S+ from the multiple-peck (high-effort) trials and the S+ from the one-peck (low-effort) trials, and likewise between the two respective S− stimuli. Using a slightly different procedure that permitted assessment of the relative aversiveness of low versus high effort, Experiment 6 again revealed a pattern of indifference despite showing that pigeons took considerably longer to begin pecking on high- than on low-effort trials. Our findings call into question the reliability of the original findings and the sufficiency of the hypothesized within-trial contrast mechanism to produce them.
Keywords: work ethic; stimulus value; simultaneous discrimination; within-trial contrast; delay-reduction hypothesis; key peck, pigeons
This paper reports a series of experiments conducted over the course of a few years to investigate the interesting and provocative “work ethic” effect reported by Clement, Feltus, Kaiser, and Zentall (2000). Our initial follow-up experiments were designed to test a particular hypothesis about the origins of the effect. Unfortunately, we were unable to conduct an adequate test of that hypothesis because we were unable to replicate the findings of Clement et al. This failure led to additional replication attempts involving a variety of procedural variations, but all returned the same pattern of null findings. We also conducted an experiment resembling one described by Clement and Zentall (2002) to see if we could obtain results similar to theirs and evidence for the mechanism proposed by Zentall and his associates to explain their findings. Once again, however, we did not obtain the preference effects they reported. Considering what we view as an extensive and thorough effort at replication, we are reporting our null findings to underscore the elusiveness of the work ethic effect and the possibility that the original findings may have been a Type I error (Davison & Nevin, 2005).
Clement et al. (2000) trained pigeons on two simultaneous discriminations in which the discriminative stimuli (S+FR1 and S−FR1) for one task were obtained by pecking once to a center-key stimulus and those for the other task (S+FR20 and S−FR20) were obtained by pecking 20 times to that same initial stimulus. These low-effort and high-effort trials, respectively, were randomly intermixed in each training session. After acquiring both discriminations to high and stable levels of accuracy, pigeons then received periodic, nondifferentially reinforced probe trials on which they chose between the two S+ stimuli and, on separate probe trials, between the two S− stimuli. Clement et al. found that pigeons preferred the high-effort S+ over the low-effort S+, and the high-effort S− over the low-effort S−, even though the two S+ stimuli had virtually identical reinforcement histories (as did the two S− stimuli). Averaged across test sessions and three probe-presentation formats, pigeons selected the high-effort S+ 69% of the time and the high-effort S− 84% of the time. Thus, pigeons clearly preferred the discriminative stimuli that they had to work relatively hard to obtain in training.
Clement et al.'s (2000) explanation for this finding was that stimuli following high effort have greater conditioned reinforcing value. This greater value purportedly arises because high effort is aversive (Eisenberger, 1992) which, in turn, produces a particularly large within-trial contrast between this aversive state and the positive state generated when choosing S+ is reinforced (Clement et al., 2000; Clement & Zentall, 2002; Friedrich & Zentall, 2004). We hypothesized, however, that differences in the conditioned reinforcing values of the high- versus low-effort stimuli were explicable in terms of the delay-reduction hypothesis of conditioned reinforcement (Fantino, 1977). Our reasoning was as follows. It takes longer to complete 20 pecks than one peck, so the overall wait (or delay) to reinforcement timed from the onset of the initial stimulus is longer on high- than on low-effort trials, TH > TL. Consequently, even though the delay to reinforcement timed from the appearance of the discriminative stimuli is the same for both sets of stimuli (tH = tL), the reduction in the overall delay to reinforcement associated with the appearance of the high-effort stimuli is greater than it is with the low-effort stimuli: (TH − tH) > (TL − tL). This should make the high-effort discriminative stimuli more potent conditioned reinforcers.
To distinguish between a purely effort-based account and a time-based account, we implemented a design that involved a low- versus high-effort difference (e.g., 1 vs. 20 pecks to the initial stimulus) but an equal overall delay to reinforcement. In other words, our procedure insured that (TH − tH) = (TL − tL). By our hypothesis, this should eliminate the preference for the high-effort stimuli in the Clement et al. (2000) procedure.
Unfortunately, there was no preference to eliminate: Pigeons trained under conditions identical to those of Clement et al. (2000) were indifferent, on average, to the two S+ stimuli and, for the most part, to the two S− stimuli on the probe-trial preference tests. The first five experiments detail the null findings we obtained using the Clement et al. methodology. Experiment 6 was modeled after Clement and Zentall (2002, Experiment 1) in order to see if a different procedure might yield a stimulus preference and, at the same time, if prechoice patterns of performance were consistent with greater aversiveness of high effort. Once again, preferences failed to emerge despite independent evidence that high effort is indeed aversive relative to low effort.
Experiments 1–5
Experiments 1–5 were structurally very similar. They differed in terms of the stimuli used for simultaneous discrimination training, the size of the large fixed-ratio (FR) component, whether or not the initial-link FR requirement was signaled, and whether or not they included a separate group for which trial durations on small- and large-FR trials were equated.
Method
Subjects
White Carneau retired breeders obtained from the Palmetto Pigeon Plant (Sumter, SC), and maintained at 80% of their free-feeding body weights by restricted feeding, were used. Each pigeon was housed individually in a stainless steel, wire-mesh cage in a colony room on a 14 hr:10 hr day–night cycle with lights on at approximately 07:00. Grit and water were always available in the home cage. Daily food rations were obtained in the experimental apparatus except on the 1 day per week pigeons were not run. Four experimentally naïve pigeons were used in Experiment 4. Eight pigeons, all with prior experience in two-alternative delayed matching and randomly divided into two groups of four, participated in Experiments 1, 2, 3, and 5. Different pigeons were used in each experiment.
Apparatus
The experiments were run using either one or two pigeon chambers, consisting of BRS/LVE (Laurel, MD) Model PIP-016 panels inside BRS/LVE Model SEC-002 enclosures. The pigeon's compartment in each chamber (36.8 cm high × 30.5 cm wide × 34.3 cm deep) was equipped with an aluminum grid floor. Three clear plastic pecking keys, 2.5 cm in diameter and spaced 8.3 cm center-to-center, were about 25 cm above the floor on the panel. Each had a BRS/LVE Model IC-901-IDD in-line projector mounted behind it for displaying visual stimuli. The center key projector could display a solid white dot and three white vertical or horizontal lines on black backgrounds (BRS/LVE Pattern No. 692). The side-key projectors could display a white vertical or a white horizontal line on a black background, a white triangle and a white annulus also on black backgrounds (BRS/LVE Pattern No. 696) and various homogeneous colored fields (red, green, blue, and yellow) via filters provided with the projectors (Experiments 1 and 2) or via Wratten filters #26, #60, #38a, and #9, respectively (Experiments 3–5). A partially covered GE #1829 bulb 5.7 cm above the center key served as a houselight. Its light was directed toward the ceiling. Purina ProGrains was delivered in a magazine which, when raised, was accompanied by lighting a small miniature bulb (ESB-28) in a metal housing that covered it. A 5.8-cm-square opening centered approximately 13 cm below the center key allowed access to the back-mounted magazine. Ventilation and masking noise was provided by continuously running blower fans attached to the chambers. IBM-compatible 386 computers located in an adjacent room controlled and recorded all experimental events.
Procedure
Each experiment involved preliminary training, choice training on two concurrent simultaneous discriminations, and 5 days of preference testing. The general aspects of these experiments are described first, followed by the particulars for each experiment.
Preliminary training
Experimentally naïve pigeons (Experiment 4) initially were taught to eat quickly and reliably from the lit food magazine, after which their key peck response to the center-key dot was shaped via the method of successive approximations. Next, they learned to peck at red and green, or blue and yellow, stimuli on the left and right keys. A single peck to the color appearing on each trial turned it off and produced 3-s access to food. Successive trials in these two 60-trial sessions were separated by a 10-s intertrial interval (ITI). Each color appeared equally often on both keys, and the houselight remained on throughout each session.
Next, the pigeons in Experiments 1–4 learned to obtain food by pecking the center-key dot multiple times. There were 60 stimulus presentations per session, and the number of pecks required for reinforcement was increased from 2 to 20 across seven to eight sessions. Completion of the FR requirement immediately turned off the dot and produced food. The houselight was off during each trial and was on throughout the ensuing 10-s ITI. After two sessions with the FR 20, pigeons received two additional sessions in which pecking the dot was reinforced on a mixed FR 1 FR 20 schedule. On half of the 60 trials in these sessions, food was contingent on a single peck to the dot; on the other half of the trials, it was contingent on completing an FR 20. Pigeons in Experiment 5 received either mixed- or multiple-schedule training from the outset (see below), with the large FR gradually raised from 3 to 40 over the course of 15 to 16 sessions. The duration of food reinforcement was constant within a session but varied from 2 to 6 s across sessions so as to maintain body weights as close to the 80% free-feeding values as possible.
Choice training
During choice training, pigeons learned two simultaneous discriminations, one preceded by a single peck to the center-key stimulus (FR 1 trials) and the other by pecking the center-key stimulus multiple times (FR 20 in Experiments 1–4 or FR 40 in Experiment 5). Completion of the scheduled FR on each trial turned off the center-key stimulus and immediately produced (or eventually produced—see Experiment 1 procedure below) an S+ and an S− stimulus on the two side keys. One set of discriminative stimuli always appeared on FR 1 trials, and the other set always appeared on FR 20 (or FR 40) trials, with each stimulus in a set appearing equally often on the left and right keys. The first key peck to either discriminative stimulus turned off the other side-key stimulus and initiated a 6-s interval that ended response-independently with the offset of the remaining stimulus and presentation of food (if the S+ had been chosen) or a timeout of equivalent duration (if the S− had been chosen).
The two simultaneous discriminations occurred in one of two random orders with the constraints that each occur 48 times per 96-trial session and that none of the four possible trial types—FR 1 versus FR 20 (or FR 40) initial link × the two left–right positions of the terminal-link discriminative stimuli—occur more than three times in a row. The houselight was off during a trial and was on throughout the 10-s ITI. The duration of food reinforcement was again varied across sessions as needed to maintain each pigeon's 80% body weight.
Pigeons were trained to a criterion of one session in which they chose the S+ stimulus 90% or more of the time on both the FR 1 and FR 20 (or FR 40) simultaneous discriminations. Twenty overtraining sessions were then given.
Preference testing
Choice training was followed by five preference-test sessions. Each contained 60 baseline trials, 30 of each simultaneous discrimination, and 36 test trials. Half of the test trials involved a choice between the S+ from the FR 1 simultaneous discrimination and the S+ from the FR 20 (or FR 40) discrimination. The other half involved a choice between the two respective S− stimuli. A single peck to either of the two S+ or the two S− stimuli immediately turned off the other stimulus and initiated a 6-s interval that ended with the remaining stimulus turned off and with food with a .5 probability. In other words, choices on the test trials were partially and nondifferentially reinforced.
Twelve test trials (six with the two S+ stimuli and six with the two S− stimuli) were preceded by a single peck (FR 1) to the center-key stimulus. Another twelve were preceded by completing an FR 20 (or 40) to the center-key stimulus. On the remaining twelve test trials, the two S+ or the two S− stimuli simply appeared on the side keys at the end of the ITI (viz., they were not preceded by a center-key stimulus). The 36 test trials were randomly intermixed among the 60 training trials with the constraints that each of the four possible trial types in training occur at least once before the first test trial and that no more than two test trials occur in a row.
Experiment 1
Two groups of 4 pigeons were run, one under conditions identical to those described by Clement et al. (2000) and the other for which FR 1 and FR 20 trial durations (timed from the onset of a center-key dot to the onset of the choice stimuli) were equated. This was accomplished by recording FR 20 trial durations on a trial-by-trial basis and, on an FR 1 trial, inserting a blank interval between the offset of the center-key dot and onset of the FR 1 choice stimuli that equaled a previous FR 20 duration. The FR 20 durations were used sequentially. A “seed” duration was used if the first trial of a session was an FR 1 trial.1
For half of the pigeons in each group, red and yellow were the choice stimuli for the FR 1 trials, and blue and green were the choice stimuli for the FR 20 trials. For the other half, these assignments were reversed. Also, red and blue served as the S+ stimuli, and green and yellow as the S− stimuli, for half of the pigeons in each group and vice versa for the other half. During the preference test, then, pigeons chose between red versus blue and between green versus yellow.
Experiment 2
Two groups like those described above were run, except that red and green served as the S+ stimuli (and yellow and blue as the S− stimuli) for half of the pigeons, and vice versa for the other half. Test trials, then, involved choices between red versus green and blue versus yellow. This change kept the color pairings identical to those used by Clement et al. (2000).
Experiment 3
This was identical to Experiment 2 except that Wratten filters #26, #60, #38a, and #9, respectively, were used to produce the red, green, blue, and yellow discriminative stimuli. These were the same filters used by Clement et al. (2000) and were provided to us by the Zentall lab.
Experiment 4
Experimentally naïve pigeons were used, and all were run under conditions like those described by Clement et al. (2000). The Wratten filters again were used to produce the color stimuli. After the five color preference tests between red and green and between blue and yellow, pigeons were retrained using vertical and horizontal lines, and the triangle and annulus forms, as the choice stimuli. These were counterbalanced with respect to the FR initial-link requirements and their roles as S+ and S−. Test trials involved choices between vertical and triangle, and between horizontal and annulus.
Experiment 5
Two groups of pigeons were trained with FR 40 as the large requirement and with the same choice stimuli as in Experiment 3. For Group Dot, a mixed FR 1 FR 40 schedule was in effect for pecking the center-key dot. For Group Line, vertical and horizontal center-key lines signaled the two response requirements (a multiple FR 1 FR 40 schedule), with the line-FR relations counterbalanced across pigeons.
Results
Choice training
In all five experiments, pigeons rapidly acquired the two simultaneous discriminations in training. With hues as the discriminative stimuli, the average number of sessions to criterion (including the criterion session) across all five experiments was 2.7, with most pigeons needing only 2 or 3 sessions to reach 90% or better accuracy on both discriminations. With the line orientations and forms as stimuli (Experiment 4), pigeons required an average of 4.8 sessions to meet criterion (range: 4 to 6 sessions). Terminal levels of accuracy were virtually indistinguishable for the two discriminations. Over the last five overtraining sessions, the mean across-experiment accuracy on the discrimination that followed the FR 1 initial link was 99.5% (range: 98.3%–99.9%) versus 99.8% on the discrimination that followed the FR 20 (or FR 40) initial link (range: 98.8%–100.0%). There were no significant between-discrimination differences within any experiment, largest F(1, 3) = 2.01.
Preference testing
The data from the three “standard” groups (cf. Clement et al., 2000) in Experiments 1–3 were combined, as were the data from the three equal-duration groups, because preliminary analyses showed no significant across-experiment differences in overall S+ preferences or in overall S− preferences for either training condition, largest F(2, 9) = 2.35, and no significant interactions between training condition and the response requirement (none, FR 1, or FR 20) preceding the S+ or S− preference test, largest F(4, 18) = 1.16. The combination was also justified on the grounds that all pigeons were experienced, hues were used as discriminative stimuli in each experiment, and the same contingencies and procedures were in effect across experiments.
Figure 1 shows mean preferences averaged over the five test sessions for choosing the S+ stimulus (solid bars) and the S− stimulus (hatched bars) that followed the FR 20 peck requirement in training. The results from the standard and equal-duration groups appear in the top and bottom panels, respectively, and are broken down according to the initial-link work requirement that preceded the two S+ stimuli or the two S− stimuli on test trials. (“None” indicates that there was no initial link prior to the appearance of the test stimuli—i.e., those stimuli were presented immediately after the preceding intertrial interval). Appendix A shows the preference data for individual subjects.
Fig 1. Average percentage choices (+ SEM) of the FR 20 stimuli from training on the probe-trial tests in Experiments 1, 2, and 3 combined as a function of initial-link work requirement.

Solid bars show the results from the S+ probe trials; hatched bars show the results from the S− probe trials. Top panel: Data from groups run in the same fashion as that reported in Clement et al. (2000). Bottom panel: Data from groups for which trial durations timed from the onset of the initial stimulus to onset of the discriminative stimuli were the same on low- versus high-effort trials.
None of the mean preferences in the standard group deviated significantly from chance, all ts (11) < 1.60, although there was a clear tendency toward choosing the low-effort (FR 1) S+ and S− on test trials preceded by an FR 20 initial link. For example, 7 of the 12 pigeons chose the FR 1 S− from training 83% of the time or more when given a choice between that stimulus and the FR 20 S− training stimulus. In the equal-duration group, there was a significant preference for the high-effort (FR 20) S− on the S− test trials preceded by an FR 1 initial link, t(11) = 2.41. No other preferences were significant, ts(11) < 1.80, although again there was a tendency toward choosing the low-effort S− on FR 20 initial-link test trials.
Figure 2 depicts the corresponding mean preferences for the experimentally naïve pigeons in Experiment 4 averaged across their five hue and five form test sessions. There were no significant overall preferences for the high-effort S+ (solid bars) on either test, ts(3) < 2.06, although there was a tendency in that direction on the form S+ test following an FR 20 initial link. On the S− test trials (hatched bars), the preference for the high-effort S− approached significance on the form test trials preceded by an FR 1 initial link, t(3) = 2.86, p = .06, two-tailed. All other S− preferences were not significant despite appearances, ts(3) < 1.61, due to considerable pigeon-to-pigeon variability. For example, on the hue S− test trials preceded by an FR 1 initial link, 2 pigeons chose the high-effort S− more than 90% of the time but the other 2 pigeons chose the high-effort S− less than 50% of the time (see Appendix A).
Fig 2. Average percentage choices (+ SEM) of the FR 20 stimuli from training on the probe-trial tests in Experiment 4 as a function of initial-link work requirement.

Solid bars show the results from the S+ probe trials; hatched bars show the results from the S− probe trials. Top panel: Preference data following simultaneous discrimination training with hue stimuli. Bottom panel: Preference data following simultaneous discrimination training with form stimuli.
Figure 3 shows the high-effort stimulus preferences in Experiment 5 where the large FR was 40 (rather than 20). Data from the group trained on the mixed FR 1 FR 40 schedule with the center-key dot (Group Dot) are shown in the top panel; data from the group trained on the mult FR 1 FR 40 schedule with different center-key line orientations (Group Line) are shown in the bottom panel. There were no significant preferences on the S+ test trials (solid bars) in either group. Likewise, there were no significant preferences in Group Line on the S− test trials (hatched bars), although there was a tendency toward choosing the low-effort S− following an FR 40 initial link, t(3) = −2.17, p < .12, two-tailed. Despite appearances, there were no significant overall preferences on the S− test trials in Group Dot, ts(3) < 1.21, because 1 pigeon in this group showed a very strong preference for the low-effort (FR 1) S− even though the other 3 pigeons showed strong preferences for the high-effort (FR 40) S− (see Appendix A).
Fig 3. Average percentage choices (+ SEM) of the FR 40 stimuli from training on the probe-trial tests in Experiment 5 as a function of initial-link work requirement.

Solid bars show the results from the S+ probe trials; hatched bars show the results from the S− probe trials. Top panel: Preference data from pigeons trained with a dot initial stimulus. Bottom panel: Preference data from pigeons trained with vertical and horizontal lines as initial stimuli signaling low- versus high-effort requirements.
Discussion
None of our five experiments replicated the work-ethic effect reported by Clement et al. (2000) despite procedures virtually identical to theirs and, in three experiments, using hue stimuli generated from the same Wratten filters. One reasonable concern is that our groups contained fewer subjects than in Clement et al., so we had less power of detecting a true preference effect. But this concern is muted by the fact that pooling the data from Experiments 1 to 3, thus creating larger overall Ns than used by Clement et al., did not change the outcome, except for a significant preference for the high-effort S− on one type of test trial in the equal-duration group. Also, if the subject-to-subject variability observed with our small sample sizes is indicative of the variability we would have seen had we run another 4 subjects per condition or group, the results would likely have been the same. The fact that this variability was frequently reproduced across experiments gives us confidence in this assessment.
But what about the obvious trends toward a preference for the high-effort S− in Experiments 4 and 5 and the significant preference for this stimulus on the FR 1 test trials for the equal-duration groups of Experiments 1–3? Assuming there are true preferences here, we are left with the problem of explaining how they could arise despite no preference for the high-effort S+. After all, the latter would seem to be necessary for the former (Clement et al., 2000) because reinforcing correct choice of the high-effort S+ during training purportedly endows that S+ with a higher overall value than the value imparted to the low-effort S+ when it is correctly chosen. This value difference is thought to arise from a larger within-trial contrast between the positive attributes of reinforcement and the substantially negative attributes of a large work requirement vis-à-vis the less negative attributes of a small work requirement (e.g., Friedrich & Zentall, 2004). The larger positive value of the high-effort S+ in turn creates a preference for the high-effort S− through the process of value transfer (e.g., Urcuioli, 2006; Zentall & Sherburne, 1994): A high-effort S− has more positive value available to be transferred to it from the high-effort S+ than does the low-effort S− from the low-effort S+. But this analysis must certainly predict that the hypothesized greater positive value of the high-effort S+ would be reflected in a preference for that stimulus over the low-effort S+. None of our experiments, however, showed such a preference.
An alternative explanation for the trend toward preference for the high-effort S− is suggested by the fact that this preference was apparent only (or mostly) when choices were preceded by an FR 1 initial link or by no initial link. The bias toward pecking the high-effort S− in testing could have occurred if instead of learning to peck the S+ on FR 1 trials during choice training, the pigeons learned not to peck the S− on these trials (Clement et al., 2000). Avoidance of the low-effort S− on the FR 1 test trials would result in the selection of the alternative, high-effort S−. Also, if not pecking is more similar to one peck than it is to 20 (or 40 pecks), generalization would produce the same bias on the S− test trials conducted without an initial link. This same argument, of course, should apply to test trials in which the initial link was an FR 20 (or FR 40): Pigeons should be more likely to select the low-effort S− if they had learned to avoid the high-effort S− during training. This overall trend can also be seen in most of the data.
Experiment 6
Clement and Zentall (2002) proposed that the within-trial contrast purportedly responsible for the work-ethic effect can also occur from the anticipation of differential effort. In their Experiment 1, trials began with the presentation of either vertical or horizontal lines. Pecking the vertical lines produced a simultaneous discrimination on half of the trials and a low-effort (FR 1 reinforced) stimulus on the other trials. Pecking the horizontal lines produced a different simultaneous discrimination on half of the trials and a high-effort (FR 30 reinforced) stimulus on the other trials. During subsequent testing, pigeons periodically chose between the S+ stimuli from the simultaneous discriminations and between the corresponding S− stimuli. Consistent with their hypothesis, Clement and Zentall found a preference for the high-effort S+ and for the high-effort S−. Because the high- versus low-FR stimuli and the simultaneous discriminations were presented on separate trials (i.e., differential effort did not directly precede the discriminative stimuli), the authors suggested that their pigeons judged the value of each S+ and S− relative to their expectation of differential effort as signaled by the initial line stimuli, a second-order contrast effect.
To follow up on these more recent findings and to evaluate the sufficiency of the mechanism(s) proposed by Clement and Zentall (2002; see also Friedrich & Zentall, 2004), we arranged that both the anticipation of different effort and differences in actual effort would precede presentation of different sets of discriminative stimuli for one group (Group Signaled). This was accomplished by using different initial stimuli to signal low versus high forthcoming effort. For a second group (Group Unsignaled), the initial stimulus was always the same and, hence, uninformative regarding forthcoming effort. In short, the discriminative stimuli for Group Signaled were preceded by the anticipation of effort and actual effort, whereas for Group Unsignaled they were preceded only by actual effort.
For both groups, the high-effort FR requirement was set to 80 in an attempt to increase the chances of seeing a work ethic effect which, to this point, had failed to materialize. According to Clement et al. (2000; see also Clement & Zentall, 2002; DiGian, Friedrich, & Zentall, 2004; Friedrich & Zentall, 2004), preference for the discriminative stimuli preceded by this especially large effort should be apparent in both groups. Furthermore, if the anticipation of effort combines with actual effort, those preferences should be more pronounced in Group Signaled than in Group Unsignaled or, at a minimum, should appear in the former group even if they remain absent in the latter group.
Method
Subjects and Apparatus
Sixteen White Carneau retired breeders participated in Experiment 6. All were experienced with a variety of procedures orthogonal to those of the present experiment. They were housed and maintained as described previously. Two operant chambers similar to those used in Experiments 1 to 5 were used.
Procedure
Pigeons were randomly assigned to Group Signaled or Group Unsignaled and to each experimental chamber, with 8 pigeons per group and chamber.
Preliminary training
Subjects initially were trained to obtain food by pecking a dot on the center key, with the response requirement progressively increased to 80 pecks (FR 80) over 19 to 27 sessions. Next, pigeons were trained to peck vertical and horizontal lines on the center key, with these trials randomly intermixed within each session. The FR requirement was progressively increased to FR 5 over three sessions. Completion of the FR requirements produced 2 to 6-s access to food, adjusted individually for each pigeon in each session so as to maintain its 80% body weight. Each preliminary training session had 60 trials with successive trials separated by a 10-s ITI. The houselight remained on during the ITI and off during the presentation of the stimuli and food reinforcement.
Choice training
As shown in Figure 4, each discrimination training trial for Group Signaled began with the offset of the houselight and the illumination of either vertical or horizontal lines on the center key. Completing an FR 5 requirement turned off the center-key line stimulus and turned on a center-key dot, to which 1 or 80 pecks (FR 1 or FR 80) were required, depending on which initial line stimulus had been presented. For 4 pigeons, one peck to the dot was required when the initial stimulus was vertical and 80 pecks to the dot were required when the initial stimulus was horizontal. For the remaining 4 pigeons, these contingencies were reversed. Completion of the FR 1 or FR 80 requirement terminated the dot and produced two different hues on the side keys. On trials with the FR 80 work requirement, one set of hues (high-effort S+ and high-effort S−) was presented, whereas on trials with the FR 1 work requirement, a different set of hues (low-effort S+ and low-effort S−) was presented. Once the discriminative stimuli appeared, a single peck to either hue turned off the other side-key stimulus and initiated a 6-s period that ended in food reinforcement if the S+ was pecked first or in nonreinforcement if the S− was pecked first. A 10-s ITI with the houselight on then commenced.
Fig 4. High- and low-effort training trials for Groups Signaled and Unsignaled in Experiment 6.
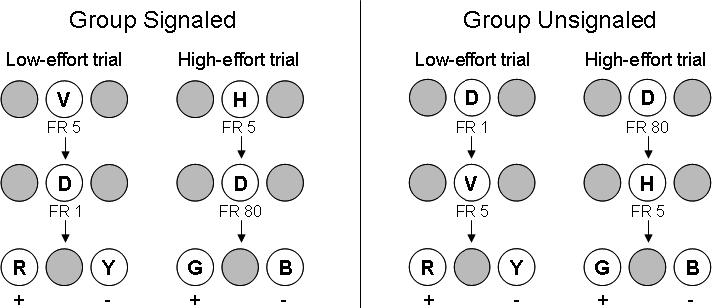
Location of the S+ and S− (left or right) was counterbalanced over trials, and the S+ and S− hues were counterbalanced within each group. V = vertical lines; H = horizontal lines; D = dot ; R = red; Y = yellow; G = green; B = blue; + = reinforced choice; − = nonreinforced choice.
For Group Unsignaled, the order of the line and dot stimuli was reversed. The initial center-key stimulus was the dot to which an FR 1 or FR 80 was required, and the second stimulus was either vertical or horizontal to which an FR 5 was then required. For 4 pigeons, the vertical lines appeared after completing an FR 1 to the dot and the horizontal lines appeared after completing an FR 80 requirement. For the remaining pigeons, these contingencies were reversed. After subsequently completing the FR 5 requirement to the lines, the discrimination trial proceeded as described for the signaled group (i.e., one set of hues appeared on FR 1 trials and a different set appeared on FR 80 trials).
For both groups, red and yellow, and green and blue always appeared together as discriminative stimuli. The colors within a set appeared equally often as high-effort S+ and high-effort S−, or as low-effort S+ and low-effort S−, across the pigeons in each group. Each training session contained 96 trials, 48 involving high effort and 48 involving low effort, with left-right locations of the S+ and S− stimuli counterbalanced across trials. Trials were presented in a pseudorandom order, with the constraint that no trial type (high- vs. low-effort) occur more than three times in succession. During choice training (and subsequent preference testing), the shortest possible reinforcement duration was 1.0 s rather than 2.0 s.
Pigeons were trained to a criterion of at least 90% correct on both discriminations within the same session. After meeting this criterion, each pigeon received an additional 20 overtraining sessions. During overtraining, 3 pigeons from Group Signaled and 2 from Group Unsignaled stopped pecking during high-effort trials. For 3 of these pigeons, the FR 80 requirement was reduced to FR 60; for the remaining 2 pigeons, the FR requirement was further reduced to FR 50 and FR 40. One pigeon from Group Signaled was eventually dropped from the experiment due to an injury.
Preference testing
Five test sessions immediately followed discrimination training. Each test session included 36 test trials randomly intermixed among 60 training trials. Half of the test trials involved a choice between the two S+ stimuli (high-effort S+ vs. low-effort S+) and the other half a choice between the two S− stimuli (high-effort S− vs. low-effort S−). All test trials commenced with the presentation of the two S+ or the two S− stimuli, with left–right locations counterbalanced across test trials. As on training trials, a single peck to either side key turned off the other side key and initiated a 6-s period. After the 6-s interval elapsed, however, food reinforcement randomly occurred 50% of the time independently of which stimulus was initially pecked.
Results
Choice training
Group Signaled and Group Unsignaled learned the two hue discriminations in an average of 2.7 and 3.0 sessions, respectively. A two-way, mixed ANOVA with group and trial type as factors yielded no significant effect of group, trial type, or their interaction, all Fs(1,13) < 2.67. During overtraining, both groups continued to perform accurately on each trial type: 99.8% and 99.7% correct on high- and low-effort trials, respectively, for Group Signaled; 99.6% and 99.8% correct, respectively, for Group Unsignaled. There were no significant effects of group, trial type, or their interaction, all Fs(1,13) < 1.00.
The top panel of Figure 5 shows the average latency to peck the initial stimulus (the dot for Group Unsignaled, and vertical or horizontal lines for Group Signaled) over the last five training sessions. On the small FR (low-effort) trials, the groups did not differ in their latencies: 1.4 s for both groups. On the large FR (high-effort) trials, however, Group Signaled showed much longer latencies than Group Unsignaled (52.5 s vs. 2.1 s, respectively). A two-way, mixed ANOVA performed on these data yielded a significant effect of group [F(1,13) = 10.60], trial type [F(1,13) = 11.33], and their interaction [F(1,13) = 10.73]. The significant interaction confirms that Group Signaled could anticipate low versus high effort upon seeing the initial stimulus whereas Group Unsignaled could not.
Fig 5. Top panel: Average latency (+ SEM) to peck the initial stimulus on low-effort (Small FR) and high-effort (Large FR) trials over the last five training sessions for each group in Experiment 6.

Bottom panel: Trial duration (+ SEM) measured from the onset of the initial center-key stimulus to the onset of the choice stimuli averaged over the last five training sessions for each group in Experiment 6 on low-effort (Small FR) and high-effort (Large FR) trials.
The bottom panel of Figure 5 plots the duration of each trial type measured from the onset of the initial center-key stimulus to the appearance of the hue choice stimuli averaged over the last five training sessions. As expected, the duration of the large-FR (high-effort) trials was significantly greater than the duration of the small-FR (low-effort) trials for both groups. ANOVA showed an overall effect of trial type, F(1,13) = 32.92, but no effect of group [F(1,13) < 1.00] or Group × Trial-type interaction [F(1,13) < 1.00].
The absence of a Group × Trial type interaction seems, at first glance, at odds with the significant interaction found for latencies. This apparent inconsistency is explained by the behavior of the pigeons in Group Unsignaled on large-FR trials. If the first peck to the initial, center-key dot did not remove the dot, then these pigeons typically would pause before continuing to peck. Thus, the high-effort trials for Group Unsignaled were characterized by an initial short latency followed by a long run time whereas, for Group Signaled, these trials were characterized by a long latency followed by a somewhat shorter run time. Stated otherwise, the long run time of Group Unsignaled compensated for the long latency of Group Signaled.
Preference testing
During testing, the two groups continued to perform accurately on the baseline discriminations. Accuracy on high- and low-effort trials averaged 99.9% and 99.0%, respectively, in Group Signaled and 99.7% and 99.9%, respectively, in Group Unsignaled. ANOVA revealed no significant effects, all Fs(1,13) < 4.38.
As shown in the top panel of Figure 6, on the first test session the pigeons in Group Signaled chose the high-effort S+ over the low-effort S+ 38.1% of the time (filled bars) and chose the high-effort S− over the low-effort S− 61.9% of the time (hatched bars). The corresponding percentages for Group Unsignaled were 39.6% and 54.2%, respectively. t-tests showed that there were no significant deviations from chance in Group Signaled [t(6) = 1.99 for the high-effort S+ and t(6) = −1.26 for the high-effort S−] or in Group Unsignaled [t(7) = −1.36 for the high-effort S+ and t(7) = 1.0 for the high-effort S−].
Fig 6. Average percentage choices (+ SEM) of the large-FR S+ stimulus (solid bars) and large-FR S− stimulus (hatched bars) stimuli from training on the probe-trial tests for each group in Experiment 6.

Top panel: Preference data from the first test session. Bottom panel: Preference data averaged across the five test sessions.
ANOVA using test-trial type and group as factors showed a significant overall effect of test-trial type, F(1,13) = 7.45, but no significant group effect or Group × Trial type interaction, Fs(1,13) < 1.00. In short, the preference for the high-effort S− averaged across groups was higher than the overall preference for the high-effort S+.
The bottom panel of Figure 6 shows the preferences averaged over all five test sessions. The pattern of results is similar to the first test session. Group Signaled chose the high-effort S+ 43.3% of the time (filled bars) and the high-effort S− 62.4% of the time (hatched bars). The corresponding percentages for Group Unsignaled were 42.9% and 50.8%, respectively. t-tests again showed no significant deviation from chance either in Group Signaled [t(6) = −0.89 and t(6) = 1.64 for the high-effort S+ and the high-effort S−, respectively] or in Group Unsignaled [t(7 ) = −1.18 for the high-effort S+ and t(7) = 0.26 for the high-effort S−]. Individual-subject data averaged over the five test sessions are shown in Appendix A.
As for the first test session, ANOVA yielded a significant overall effect of test-trial type, F(1,13) = 4.73, but no significant group effect or Group × Trial-type interaction, Fs(1,13) < 1.00. Thus, across the five test sessions, the overall preference for the high-effort S− was significantly higher than the overall preference for the high-effort S+.
Discussion
According to Clement and Zentall (2002), within-trial contrast can operate on the basis of actual differential effort or the anticipation of differential effort. In this experiment, both groups experienced differential effort and, for Group Signaled, conditions were manipulated to produce the anticipation of differential effort as well. Additionally, the high-effort FR requirement was increased to FR 80 in order to enhance any within-trial contrast that might operate.
Despite our efforts, neither group showed a preference for the high-effort S+ or for the high-effort S−. Overall, results from Group Unsignaled followed the pattern of the previous five experiments in the sense that no work-ethic effect was observed. Likewise, Group Signaled revealed no significant deviations from chance during testing, although the latency data strongly suggest that these pigeons were able to anticipate the FR requirement involved in each trial. Thus, the work-ethic effect did not materialize even when actual differential effort was combined with the anticipation of effort.
Another interesting feature of the data is that during training, the two simultaneous discriminations were preceded by different delays (i.e., different times required to complete all response requirements prior to the appearance of the S+ and S−; Figure 5, bottom panel). In principle, this fact should, by itself, produce within-trial contrast given that a long delay is considered relatively more aversive than a short one (DiGian et al., 2004). In DiGian et al., completion of an FR 20 requirement to an initial stimulus immediately produced one set of discriminative stimuli on some trials (i.e., after a 0-s delay) and a different set of discriminative stimuli on other trials after a 6-s delay. For some pigeons, delay was unsignaled (the initial stimulus was white for both delays) whereas for others, it was signaled (the initial stimulus was either vertical or horizontal depending on the delay it would follow). In testing, DiGian et al. found a significant preference for the long-delay S+ in the signaled group but not in the unsignaled group. Although it is not entirely clear why different delays must be signaled to produce the effect, one would nonetheless predict given their findings that a preference for the high-effort S+ should be found in Group Signaled in the present experiment. Clearly, that was not the case.
A possible objection is that effort and time were confounded in the present experiment. After all, we observed different delays only because different work requirements were imposed. But it seems implausible that the combination of long delay to reinforcement and high work would detract from the hypothesized influence of within-trial contrast. Besides, in the original work-ethic demonstration (Clement et al., 2000), work and delay were confounded but preferences nonetheless materialized. In fact, Experiments 1–3 reported here were run precisely to separate the effect, if any, of each confounded variable. Theoretically, contrast should be enhanced by any factor capable of producing differentially preferred events (for details, see DiGian et al., 2004).
In summary, despite several ingredients that should reasonably be expected to increase the probability of observing a work-ethic effect—differential effort, an especially large, high-effort FR, and (in one case) the anticipation of high effort—no significant preferences were observed. These findings are, however, consistent with the results of Experiments 1 to 5.
General Discussion
Studies of the effect of effort on the reinforcing value of discriminative stimuli previously associated with, or produced by that effort date back at least to the 1960s. For example, Lewis (1965) trained rats to obtain food by pulling an 80-g weight across a chamber compartment painted with stripes in one orientation (vertical or horizontal) and by pulling a 5-g weight across another compartment painted with stripes in the opposite orientation. Later, rats were tested for their preference for vertical versus horizontal stripes in a T maze that had differently striped side arms. There was no significant choice preference over the first 10 test trials, but preference for the high-effort stripes became apparent during 10 subsequent test trials. The reason for the difference in results across test blocks is unclear given that lighting conditions varied between the first and second block as did the spacing of the trials within blocks. In any event, Lewis concluded that the choice data provided only “…limited support…” for the influence of effort on the secondary reinforcing value of the striped stimuli (see also Lewis, 1964a, b).
Subsequent instrumental conditioning studies by Armus (1999, 2001) and Jellison (2003) demonstrate how limited this support is. Armus (1999), for example, trained rats to run on a 26° incline to a baited black (or white) goal box and on a horizontal runway to a baited goal box of the opposite brightness. Preference for black versus white was later carried out in a Y maze with stems angled upward 13°. No significant preference was found for the goal-box brightness that had followed greater running effort (i.e., the 26° incline) during training. Similar null results were reported by Armus (2001) and by Jellison (2003) when differently flavored food pellets followed high- and low-effort lever pressing during training. In those studies, the force required to operate one lever was low (5 or 8 g) relative to the force required to operate the other (50 or 75 g). When later given a choice between the differently flavored pellets in a T or Y maze, rats were indifferent.
On the other side of the ledger are the effects of effort and other apparently aversive conditions on choice preferences in avians. Pigeons have been shown to prefer discriminative stimuli that follow high effort (Clement et al., 2000), signaled long delays to reinforcement (DiGian et al., 2004), or the absence of food (Friedrich, Clement, & Zentall, 2005). This preference also extends to discriminative stimuli that do not directly follow high effort or the absence of food but, instead, follow stimuli which signal high effort or no food on other trials (Clement & Zentall, 2002). Kacelnik and his associates (Kacelnik & Marsh, 2002; Marsh, Schuck-Palm, & Kacelnik, 2004; Pompilio & Kacelnik, 2005) have reported similar findings with European starlings. For example, in Kacelnik and Marsh, starlings learned to obtain food during training by pecking a key lit by one color after 16 1-m flights (high-effort trials) and by pecking another key lit by a different color after 4 1-m flights (low-effort trials). When later given a choice between these two colors during testing, 10 out of 12 starlings showed a significant preference for the color associated with high effort.
But the reported manipulations do not always yield comparable effects. For instance, Kacelnik and Marsh (2002) did not require explicit choices during training (i.e., only an S+ stimulus appeared following low effort and following high effort) but they nonetheless obtained the work-ethic effect in their starlings during a subsequent choice test. By contrast, Clement et al. (2000) reported that in a preliminary experiment, pigeons did not exhibit preference for a high-effort stimulus after similar training during which just a single (S+) stimulus followed low-effort (FR 1) and high-effort (FR 20) responding.
Likewise, Marsh et al. (2004) showed that a starling's energetic state at the time it encounters certain stimuli influences the value of those stimuli. In that study, starlings were either food deprived or prefed before each training session during which they learned to respond to different discriminative stimuli. Later, they preferred the stimulus to which they responded when food-deprived over the one to which they responded when prefed (see also Pompilio, Kacelnik, & Behmer, 2006). But these results contrast with previous findings obtained with rats (although see Revusky, 1967). For example, in a series of experiments, Capaldi, Myers, Campbell, and Sheffer (1983) found that rats were either indifferent to tastes consumed under high versus low deprivation or preferred the taste consumed under low deprivation. Although these sorts of comparisons are confounded by across-species differences and all of the methodological differences they typically entail, the differences should nonetheless be a relatively minor concern if preferences are primarily driven by a common mechanism (e.g., within-trial contrast).
More important, methodological differences between our training and testing protocols in Experiments 1 to 5 and those of Clement et al. (2000) were essentially absent; in some cases, we used exactly the same choice stimuli. Nevertheless, preference for high-effort discriminative stimuli obtained by Clement et al. did not materialize here.
Why, then, were we unable to obtain a work-ethic effect in pigeons whereas Clement et al. (2000) obtained a clear effect? Could inconspicuous albeit functionally important details be responsible for the discrepancy? For instance, our stimuli appeared on three 2.5-cm-diameter circular pecking keys spaced 8.3 cm apart center to center. Clement et al. reported using three rectangular response keys (3.0 cm wide, 2.5 cm high) that were closer together (0.8 cm apart or 3.8 cm apart center-to-center). Comparing pretest performances (e.g., number of sessions to criterion, terminal levels of accuracy) in our experiments versus theirs gives no indication that these differences were important.
Our experiments and Clement et al. (2000) also differed in the reinforcement durations used throughout choice training and testing. Clement et al. used a fixed 1.5-s duration which usually required postsession feedings in order to maintain pigeons at 80–85% body weights (T. Zentall, personal communication, November 27, 2006). Our reinforcement durations were adjusted individually and daily so that pigeons would obtain sufficient food during their experimental sessions to maintain 80% body weights. But these adjustments were uncommon and unnecessary after pigeons reached asymptotic performance on the choice training task. Furthermore, 73% of the 275 preference tests across our six experiments were run using a 2.0-s reinforcement duration. Still, our 2.0-s duration versus the 1.5-s duration used by Clement et al. was large enough that their pigeons needed postsession feedings whereas our pigeons did not. Might the “open economy” conditions of Clement et al. versus our “closed economy” conditions (e.g., Hursh, 1980) be important? Perhaps, but it seems to us that if pigeons do not receive any extra food outside of the experimental sessions, there should be a larger positive effect of food within a session (i.e., larger within-trial contrast effects). Our conditions, then, ought to enhance rather than diminish the chances of observing the work-ethic effect.
Another way in which a work-ethic effect might be obscured is if at least some pigeons choose between the stimuli on the preference tests conditional upon the events or responses—of the sort suggested in the form test of Experiment 4 and the S+ preference test for Group Dot in Experiment 5—that immediately precede their appearance (see also Capaldi & Myers, 1982, Experiment 1 for a similar effect in rats). Although not an issue in Clement et al. (2000), other data reported by Zentall and his associates indicate that pigeons may indeed learn which choice to make and which choice not to make in training, conditional upon the particular events preceding them. DiGian et al. (2004), for example, found that pigeons were more likely to choose the S+ that was preceded by a delay during training when the test trials were initiated by a delay than when they were initiated by no delay. Although not statistically significant by their analyses, the trend seems very clear. Furthermore, an effect of the initiating event on choices made in testing was obtained in Friedrich et al. (2005): Pigeons were significantly more likely to choose the S+ that had been preceded by no food during training when the test trials began with no food than when they began with food (81.2% vs. 53.1%, respectively). The authors pointed out, however, that a significant overall preference for the S+ preceded by no food still remained when the results were pooled over the three possible initiating events for the test trials (food, no food, and no stimulus). However, it seems clear that if a high proportion of pigeons learn to make their choices in training as if those choices were conditional upon a prior event (even though they were not), the chances of detecting an overall preference will diminish.
This sort of unnecessary learning, however, does not appear to contribute in any substantive way to our null findings. Consequently, at least in terms of the specific paradigm used here, our systematic failed attempts to show preference for discriminative stimuli following high effort, over discriminative stimuli following low effort, lead us to conclude that the seminal demonstration of a work-ethic effect in pigeons (Clement et al., 2000) is not a reliable finding.
Although failed replications such as ours are often regarded (at least implicitly) as less newsworthy than demonstrations of novel effects, Davison and Nevin (2005) argue that this sort of bias, and the reinforcement contingencies from which they arise, can foster inaccurate representations of psychological processes and phenomena. At a minimum, then, our findings should leave open (or re-open) the question of whether a pigeon's prior effort enhances the conditioned reinforcing properties of stimuli that follow such effort.
Acknowledgments
Preparation of this article was made possible by National Institute of Mental Health Grant MH 66195 to Peter J. Urcuioli. The authors thank Andrea Friedrich, Christine Hoeing, Benny Kartono, Natasha Killen, Nicole Madou, Sarah Michalek, Maya Romero, John Schmeling, Samantha Steen, Steven Suska, Kristine Trent, Matt Watts, Dawn White and Holly Youngs for assistance in running subjects.
Appendix A
Individual subject preferences for the high-effort S+ and the high-effort S− averaged over the five test sessions in Experiments 1–6. Preference data are divided according to whether the choice stimuli were obtained by a single initial-link peck (FR 1), 20 or more initial-link pecks (FR 20+), or no initial-link peck (i.e., no initial-link stimulus; None).
| Experiment-Group/Condition | Subject | High-effort S+ Preference |
High-effort S− Preference |
||||
| None | FR1 | FR20+ | None | FR1 | FR20+ | ||
| Exp. 1 -Standard Group | ST1 | 46.67 | 36.67 | 3.33 | 6.67 | 13.33 | 0.00 |
| ST2 | 46.67 | 33.33 | 46.67 | 36.67 | 43.33 | 16.67 | |
| ST3 | 56.67 | 56.67 | 93.33 | 16.67 | 20.00 | 40.00 | |
| ST4 | 66.67 | 63.33 | 26.67 | 50.00 | 46.67 | 50.00 | |
| Exp. 1 - Equal-duration Group | EQ1 | 43.33 | 40.00 | 53.33 | 46.67 | 50.00 | 3.33 |
| EQ2 | 53.33 | 56.67 | 50.00 | 46.67 | 70.00 | 56.67 | |
| EQ3 | 60.00 | 60.00 | 60.00 | 80.00 | 80.00 | 76.67 | |
| EQ4 | 56.67 | 73.33 | 50.00 | 63.33 | 76.67 | 50.00 | |
| Exp. 2 - Standard Group | ST5 | 16.67 | 23.33 | 6.67 | 60.00 | 53.33 | 13.33 |
| ST6 | 53.33 | 86.67 | 56.67 | 6.67 | 0.00 | 0.00 | |
| ST7 | 60.00 | 46.67 | 20.00 | 23.33 | 16.67 | 10.00 | |
| ST8 | 43.33 | 63.33 | 36.67 | 56.67 | 56.67 | 66.67 | |
| Exp. 2 - Equal-duration Group | EQ5 | 36.67 | 23.33 | 36.67 | 36.67 | 36.67 | 3.33 |
| EQ6 | 66.67 | 63.33 | 76.67 | 73.33 | 96.67 | 46.67 | |
| EQ7 | 33.33 | 46.67 | 33.33 | 16.67 | 30.00 | 13.33 | |
| EQ8 | 56.67 | 46.67 | 40.00 | 63.33 | 66.67 | 13.33 | |
| Exp. 3 - Standard Group | ST9 | 40.00 | 16.67 | 43.33 | 96.67 | 90.00 | 93.33 |
| ST10 | 60.00 | 70.00 | 26.67 | 83.33 | 96.67 | 13.33 | |
| ST11 | 30.00 | 13.33 | 43.33 | 26.67 | 10.00 | 6.67 | |
| ST12 | 46.67 | 26.67 | 60.00 | 93.33 | 96.67 | 93.33 | |
| Exp. 3 - Equal-duration Group | EQ9 | 23.33 | 36.67 | 10.00 | 40.00 | 90.00 | 6.67 |
| EQ10 | 46.67 | 60.00 | 46.67 | 66.67 | 93.33 | 13.33 | |
| EQ11 | 60.00 | 86.67 | 80.00 | 90.00 | 93.33 | 86.67 | |
| EQ12 | 66.67 | 56.67 | 86.67 | 43.33 | 33.33 | 40.00 | |
| Exp. 4 - Hue Test | ST13 | 26.67 | 30.00 | 26.67 | 36.67 | 46.67 | 20.00 |
| ST14 | 66.67 | 70.00 | 80.00 | 93.33 | 96.67 | 86.67 | |
| ST15 | 53.33 | 30.00 | 40.00 | 36.67 | 43.33 | 0.00 | |
| ST16 | 53.33 | 53.33 | 43.33 | 76.67 | 93.33 | 56.67 | |
| Exp. 4 - Form Test | ST13 | 66.67 | 86.67 | 76.67 | 100.00 | 100.00 | 90.00 |
| ST14 | 40.00 | 26.67 | 63.33 | 43.33 | 73.33 | 46.67 | |
| ST15 | 66.67 | 50.00 | 56.67 | 60.00 | 53.33 | 26.67 | |
| ST16 | 50.00 | 53.33 | 50.00 | 66.67 | 90.00 | 10.00 | |
| Exp. 5 - Group Dot | D1 | 33.33 | 36.67 | 53.33 | 10.00 | 13.33 | 0.00 |
| D2 | 46.67 | 50.00 | 50.00 | 100.00 | 96.67 | 90.00 | |
| D3 | 20.00 | 16.67 | 33.33 | 93.33 | 96.67 | 93.33 | |
| D4 | 50.00 | 60.00 | 76.67 | 66.67 | 93.33 | 83.33 | |
| Exp. 5 - Group Line | L1 | 40.00 | 56.67 | 36.67 | 0.00 | 0.00 | 0.00 |
| L2 | 50.00 | 70.00 | 13.33 | 23.33 | 43.33 | 0.00 | |
| L3 | 53.33 | 40.00 | 76.67 | 86.67 | 93.33 | 60.00 | |
| L4 | 56.67 | 53.33 | 50.00 | 53.33 | 56.67 | 16.67 | |
| Subject | High-effort S+ Preference |
High-effort S− Preference |
|
| None | None | ||
| Exp 6 - Group Unsignaled | U1 | 46.67 | 50.00 |
| U2 | 50.00 | 33.33 | |
| U3 | 50.00 | 43.33 | |
| U4 | 70.00 | 56.67 | |
| U5 | 36.67 | 63.33 | |
| U6 | 10.00 | 50.00 | |
| U7 | 36.67 | 56.67 | |
| U8 | 43.33 | 53.33 | |
| Exp. 6 - Group Signaled | S1 | 13.33 | 43.33 |
| S2 | 43.33 | 63.33 | |
| S3 | 40.00 | 93.33 | |
| S4 | 30.00 | 46.67 | |
| S5 | 70.00 | 50.00 | |
| S6 | 66.67 | 53.33 | |
| S7 | 40.00 | 86.67 |
Footnotes
The seed duration for each pigeon in the equal duration condition was computed by taking the average difference in trial duration between FR 20 and FR 1 trials for the two sessions of mixed FR 1 FR 20 training preceding choice training and adding 6 to 7 s to the difference. The latter compensated for the 6-s interval of time between the pigeon's choice of one of the discriminative stimuli and food presentation (or timeout) during choice training. The seed duration for each pigeon was constant throughout its training, but differed across pigeons.
References
- Armus H.L. Effect of response effort on secondary reward value. Psychological Reports. 1999;84:323–328. [Google Scholar]
- Armus H.L. Effect of response effort on the reward value of distinctively flavored food pellets. Psychological Reports. 2001;88:1031–1034. doi: 10.2466/pr0.2001.88.3c.1031. [DOI] [PubMed] [Google Scholar]
- Capaldi E.D, Myers D.E. Taste preferences as a function of food deprivation during original taste exposure. Animal Learning & Behavior. 1982;10:211–219. [Google Scholar]
- Capaldi E.D, Myers D.E, Campbell D.H, Sheffer J.D. Conditioned flavor preferences based on hunger level during original flavor exposure. Animal Learning & Behavior. 1983;11:107–115. [Google Scholar]
- Clement T.S, Feltus J.R, Kaiser D.H, Zentall T.R. “Work ethic” in pigeons: Reward value is directly related to the effort or time required to obtain the reward. Psychonomic Bulletin & Review. 2000;7:100–106. doi: 10.3758/bf03210727. [DOI] [PubMed] [Google Scholar]
- Clement T.S, Zentall T.R. Second-order contrast based on the expectation of effort and reinforcement. Journal of Experimental Psychology: Animal Behavior Processes. 2002;28:64–74. [PubMed] [Google Scholar]
- Davison M, Nevin J.A. On science and the discriminative law of effect. Journal of the Experimental Analysis of Behavior. 2005;83:85–92. doi: 10.1901/jeab.2005.27-04. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DiGian K.A, Friedrich A.M, Zentall T.R. Discriminative stimuli that follow a delay have added value for pigeons. Psychonomic Bulletin & Review. 2004;11:889–895. doi: 10.3758/bf03196717. [DOI] [PubMed] [Google Scholar]
- Eisenberger R. Learned industriousness. Psychological Review. 1992;99:248–267. doi: 10.1037/0033-295x.99.2.248. [DOI] [PubMed] [Google Scholar]
- Fantino E. Conditioned reinforcement: Choice and information. In: Honig W.K, Staddon J.E.R, editors. Handbook of operant behavior. Englewood Cliffs, NJ: Prentice-Hall; 1977. pp. 313–339. [Google Scholar]
- Friedrich A.M, Clement T.S, Zentall T.R. Discriminative stimuli that follow the absence of reinforcement are preferred by pigeons over those that follow reinforcement. Learning & Behavior. 2005;33:337–342. doi: 10.3758/bf03192862. [DOI] [PubMed] [Google Scholar]
- Friedrich A.M, Zentall T.R. Pigeons shift their preference toward locations of food that take more effort to obtain. Behavioural Processes. 2004;67:405–415. doi: 10.1016/j.beproc.2004.07.001. [DOI] [PubMed] [Google Scholar]
- Hursh S.R. Economic concepts for the analysis of behavior. Journal of the Experimental Analysis of Behavior. 1980;34:219–238. doi: 10.1901/jeab.1980.34-219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jellison J.L. “Justification of effort” in rats: Effects of physical and discriminative effort on reward value. Psychological Reports. 2003;93:1095–1100. doi: 10.2466/pr0.2003.93.3f.1095. [DOI] [PubMed] [Google Scholar]
- Kacelnik A, Marsh B. Cost can increase preference in starlings. Animal Behaviour. 2002;63:245–250. [Google Scholar]
- Lewis M. Effect of effort on value: An exploratory study of children. Child Development. 1964a;35:1337–1342. doi: 10.1111/j.1467-8624.1964.tb05271.x. [DOI] [PubMed] [Google Scholar]
- Lewis M. Some nondecremental effects of effort. Journal of Comparative and Physiological Psychology. 1964b;57:367–372. doi: 10.1037/h0046172. [DOI] [PubMed] [Google Scholar]
- Lewis M. Effect of effort on choice: Value of a secondary reinforcer. Psychological Reports. 1965;16:557–560. doi: 10.2466/pr0.1965.16.2.557. [DOI] [PubMed] [Google Scholar]
- Marsh B, Schuck-Palm C, Kacelnik A. State-dependent learning affects foraging choices in starlings. Behavioral Ecology. 2004;15:396–399. [Google Scholar]
- Pompilio L, Kacelnik A. State-dependent learning and suboptimal choice: When starlings prefer long over short delays to food. Animal Behaviour. 2005;70:571–578. [Google Scholar]
- Pompilio L, Kacelnik A, Behmer S. State-dependent learned valuation drives choice in an invertebrate. Science. 2006 Mar 17;311:1613–1615. doi: 10.1126/science.1123924. [DOI] [PubMed] [Google Scholar]
- Revusky S.H. Hunger level during food consumption: Effects on subsequent preference. Psychonomic Science. 1967;7:109–110. [Google Scholar]
- Urcuioli P.J. When discrimination fails (or at least falters). Journal of Experimental Psychology: Animal Behavior Processes. 2006;32:359–370. doi: 10.1037/0097-7403.32.4.359. [DOI] [PubMed] [Google Scholar]
- Zentall T.R, Sherburne L.M. Transfer of value from S+ to S– in a simultaneous discrimination. Journal of Experimental Psychology: Animal Behavior Processes. 1994;20:176–183. doi: 10.1037//0097-7403.20.2.176. [DOI] [PubMed] [Google Scholar]