

Abstract
Tissue factor (TF) instigates the extrinsic pathway of blood coagulation. Plaque disruption and exposure of circulating factor VII/VIIa to subendothelial procoagulants such as TF leads to intravascular thrombosis, a frequent cause of acute atherosclerotic events. Although the expression of TF in the intima of human atherosclerotic lesions is well established, little is known about the mechanisms of TF regulation in vascular smooth muscle cells (SMC). We demonstrate here that TF colocalizes with the receptor CD40 on lesional SMC within atherosclerotic lesions in situ. In cultured vascular SMC, ligation of CD40 with native CD40 ligand (CD40L) derived from activated T lymphocytes or recombinant human CD40L (rCD40L) induced the transient expression of TF on the cell surface (as determined by FACS analysis) in a concentration- and time-dependent manner and enhanced total cell-associated TF (as determined by ELISA). CD40L-induced TF on vascular SMC is functional and activates coagulation. In accordance with the increased TF activity, stimulation of vascular SMC with rCD40L did not affect either protein expression or activity of tissue factor pathway inhibitors. In summary, these findings demonstrate the potential of the CD40/CD40L signaling pathway to augment the procoagulant activity in human vascular SMC. Because TF and CD40 colocalize on lesional SMC in human atheroma, CD40/CD40L signaling may contribute to the TF expression and hence to increased thrombogenicity of plaques during the inflammatory responses of atherogenesis and arterial injury.
Thrombosis, resulting in arterial occlusion, frequently causes myocardial infarction, unstable angina, stroke, or sudden death. 1,2 Furthermore, episodes of non-occlusive mural thrombosis in arteries may not produce clinical manifestations but may promote plaque growth and evolution. Plaque disruption precipitates many episodes of acute arterial thrombosis by allowing contact of the bloodstream with procoagulants within atheroma. 2,3 Tissue factor (TF) is one critical initiator of blood coagulation. 4,5 This membrane-bound 47-kd glycoprotein consists of three domains: the short cytoplasmic domain (19 amino acids), the single transmembrane domain (23 amino acids), and the extracellular domain (219 amino acids). Recent studies support the involvement of the intracellular domain in apoptosis, production of growth factors, smooth muscle cell (SMC) migration, and intracellular signaling. 6-9 TF’s well established biological function, however, depends on the large extracellular domain, which triggers the coagulation cascade. TF binds to factors VII and X, resulting in accelerated conversion of factors IX and X to the active factors IXa and Xa, respectively. TF thus promotes generation of thrombin, which converts fibrinogen to fibrin, a major component of the thrombus.
Many studies have explored the role of TF in the pathogenesis of atherosclerosis, as well reviewed elsewhere. 10 Active TF localizes in early atherosclerotic lesions and persists into the advanced stages. 1,11,12 In addition to primary arteriosclerosis, acute thrombosis triggered by procoagulant TF activity may complicate arterial interventions such as balloon angioplasty, atherectomy, or coronary artery stenting. 13-16 Such interventions expose vascular SMC to blood coagulation factors, highlighting the potential clinical relevance of the thrombogenicity of this cell type. 17
Little is known, however, regarding the regulation of this potent coagulant protein in vascular SMC. Mediators described to induce TF expression in human vascular SMC are restricted to soluble molecules, including proinflammatory cytokines, such as interleukin-1 (IL-1) and tumor necrosis factor (TNF), or growth factors such as platelet-derived growth factor. 17 Cell contact with platelets can induce TF expression on vascular SMC; however, the underlying mechanisms are unknown. 18
Recently, we localized the components of a novel signaling dyad, CD40 and CD40L, within human atherosclerotic lesions. 19 CD40, a 48-kd type I membrane protein, is a member of the TNF receptor family that includes the TNF receptors and Fas (CD95). 20-22 These receptors participate in the regulation of cell proliferation, differentiation, or apoptosis. 23-25 CD40, originally considered restricted to B lymphocytes, is expressed on vascular SMC within the atherosclerotic lesion. 19 CD40’s activator CD40 ligand (CD40L, CD154), a 40-kd cell surface molecule, originally considered restricted to activated CD4+ T lymphocytes, 26,27 is also expressed on vascular SMC 19 and belongs to the TNF family which includes TNFα, Fas ligand, and others. 24,28,29 Both CD40 and CD40L on SMC function in vitro. 30
This study tested the hypothesis that CD40 engagement on human vascular SMC via CD40L, expressed by activated T lymphocytes among other cells, induces the expression and/or activity of TF and its inhibitors by vascular SMC.
Materials and Methods
Materials
Phorbol-12 myristate 13-acetate (PMA) and Polymyxin B were purchased from Sigma (St. Louis, MO). Human recombinant CD40L (rCD40L) was generated as described previously. 31,32 To limit possible effects of endotoxin in the rCD40L preparation, polymyxin B (500 ng/ml) was added during the stimulation of the SMC. Polymyxin B alone did not affect SMC (data not shown). Anti-human CD40L and CD40 as well as fluorescein isothiocyanate (FITC)-conjugated control IgG1 mAb used for immunohistochemistry were obtained from Santa Cruz Biotechnology (Santa Cruz, CA) and PharMingen (La Jolla, CA), respectively.
Cell Isolation and Culture
Human vascular SMC were isolated from human saphenous veins by explant outgrowth and subcultured in DMEM (BioWhittaker, Walkersville, MD) supplemented with 1% L-glutamine (BioWhittaker), 1% penicillin/streptomycin, and 10% FBS, as described previously. 33 SMC were characterized by immunostaining with anti-smooth muscle cell α-actin antibody (Dako, Carpinteria, CA). The cells were cultured 24 hours before the experiment in insulin/transferrin (IT) medium, as described previously. 34 For preparation of cell lysates, as used for enzyme-linked immunosorbent assay (ELISA), SMC stimulated for the respective times were chilled on ice (10 minutes) and harvested via cell scraper techniques. Pelleted cells were resuspended in sterile phosphate-buffered saline (PBS) and lysed via three freeze/thaw cycles. Finally, total protein amount was determined.
T lymphocytes were isolated from freshly prepared human peripheral blood mononuclear cells obtained from leukopacs of healthy donors by CD4+ selection employing magnetic beads coated with anti-CD4 mAb (Dynabeads M450 CD4; Dynal Inc., Oslo, Norway) and were kindly provided by Dr. Andrew Lichtmann (Brigham and Women’s Hospital, Boston, MA). The purity of the T lymphocyte preparations was ≥98%, as determined by fluorescence-activated cell sorter (FACS) analysis (anti-human CD4+ mAb, FITC-conjugated; PharMingen, San Diego, CA). The cells were cultured 12 hours in RPMI1640 (BioWhittaker) in absence or presence of 50 ng/ml PMA and CD40L cell surface expression confirmed by FACS analysis (anti-CD40L mAb, FITC-conjugated; Calbiochem).
For preparation of cell membranes, SMC (1 × 10 6 cells/ml) or T lymphocytes (3 × 10 7 cells/ml) were resuspended in 50 mmol/L Tris-HCl, pH 7.4, 250 mmol/L NaCl, 500 mmol/L MgCl2, 0.1 mmol/L EDTA (final concentrations) and sonicated (Heat Systems Ultrasonics Inc., Plainview, NY). Cell membranes were separated from whole lysates using two-layer (0.32/2.14 mmol/L) sucrose gradient centrifugation (25,000 × g, 90 minutes, 4°C). The interface band was harvested, washed twice in 0.32 mmol/L sucrose, 100 mmol/L HEPES, 0.5 mmol/L EDTA (1,500g, 15 minutes, 4°C), and loaded again on a two-layer (0.32/1.96 mmol/L) sucrose-gradient-centrifugation (40,000 × g, 60 minutes, 4°C). Finally, the interphase band was harvested, centrifuged (10,000 × g, 15 minutes, 4°C) and the membrane preparation resuspended in phosphate buffered saline. Cultures of human vascular SMC were incubated with CD4+ T lymphocyte membrane preparations equivalent to a ratio of 1 SMC:10 T cells.
Immunohistochemistry
Surgical specimens of human carotid atheroma and aorta were obtained by protocols approved by the Human Investigation Review Committee at Brigham and Women’s Hospital. Serial cryostat sections (5 μm) were cut, air-dried onto microscope slides (Fisher Scientific, Pittsburgh, PA), and fixed in acetone at −20°C for 5 minutes. Sections were pre-incubated with PBS containing 0.3% hydrogen peroxide to inhibit endogenous peroxidase activity. The sections were then incubated (90 minutes) with primary or control (mouse myeloma protein MOPC-21, Sigma, St. Louis, MO) antibody diluted in PBS supplemented with 5% appropriate serum. After washing three times in PBS, sections were incubated with the respective biotinylated secondary antibody (45 minutes, Vector, Burlingame, CA) followed by avidin-biotin-peroxidase complex (Vectastain ABC kit, Vector), and antibody binding was visualized with 3-amino-9-ethyl carbazole (Vector) according to the recommendations provided by the supplier. Colocalization of TF with CD40 or the respective cell type used double-immunofluorescence staining. The mouse anti-human TF antibody (1:200; no. 4901, American Diagnostica, Greenwich, CT) was applied for 60 minutes followed by biotinylated anti-mouse secondary antibody for 45 minutes and Texas red-conjugated streptavidin (Amersham, Arlington Heights, IL). Subsequent to application of the avidin/biotin blocking kit (Vector), rabbit-anti-human CD40 antibody (1:250, Santa Cruz Biotechnology) or anti-smooth muscle actin mAb for SMC (Enzo Diagnostics, New York, NY) was added and sections incubated overnight at 4°C. Subsequently, the appropriate secondary antibodies were applied for 30 minutes followed by streptavidin-FITC (Amersham).
Flow Cytometry
Human vascular SMC were washed with ice-cold PBS (Ca2+/Mg2+ free), harvested by scraping and subsequently washed once with PBS/1% FBS before being incubated with the FITC-conjugated TF antibody (30 minutes, 4°C). After immunofluorescence labeling, cells were washed twice with PBS/1% FBS and analyzed in a Becton Dickinson FACScan flow cytometer. Data were analyzed using CellQuest software (Becton Dickinson). At least 20,000 viable cells per condition were analyzed for the determination of percentage of positive cells.
Analysis of TF and TF Pathway Inhibitor (TFPI) Protein and Activity
Detection of TF and TFPI protein used the Immunobind TF and total TFPI sandwich ELISA kit provided by American Diagnostica, respectively. The anti-TFPI antibodies recognized full length as well as truncated forms of TFPI-1. The subsequent binding of the streptavidin-conjugated horseradish peroxidase is visualized by addition of TMB substrate as blue color, and TF/TFPI levels determined by measuring absorbance at 405 nm.
Detection of TF and TFPI activity used the Actichrome TF and TFPI Activity Assay provided by American Diagnostica. TF activity was determined as the peptidyl activity for complex formation with recombinant Factor VII and the conversion of a chromogenic substrate. Absorbance read at 405 nm was compared to those values obtained with recombinant TF standard. TFPI activity was determined as the ability to inhibit the catalytic activity of the TF/VIIa to convert recombinant factor X to Xa. Remaining TF/VIIa activity was quantitated by cleavage of a chromogenic factor Xa substrate. Absorbance read at 405 nm was compared to those values obtained with recombinant TFPI standard.
Results
Vascular SMC Coexpress TF and CD40 within Human Atherosclerotic Lesions
In accordance with previous studies, 1,11,12,35,36 human carotid atherosclerotic lesions (n = 6) consistently showed strong immunoreactivity for TF (Figure 1) ▶ . Because we recently localized the CD40/CD40L receptor/ligand pair in human atherosclerotic plaques and demonstrated that CD40 ligation induces atheroma-associated functions in human vascular SMC, 19,30,37 we investigated the possible colocalization of TF with CD40. Indeed, cells expressing TF also bear CD40 (Figure 1) ▶ . Cell morphology as well as the colocalization of immunofluorescence double-labeling of CD40 and TF with α-actin-positive cells on adjacent sections, supported the localization of TF on CD40-positive vascular SMC within atherosclerotic lesions. No immunoreactivity was observed in tissues stained with the respective control IgG.
Figure 1.
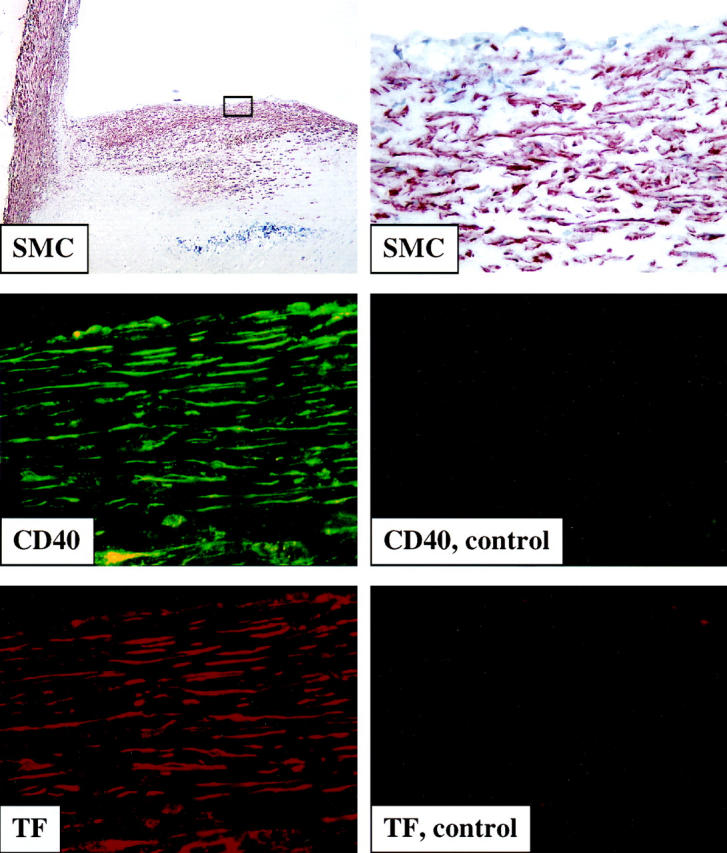
Tissue factor colocalizes with CD40-positive smooth muscle cells in human atherosclerotic lesions. Frozen sections of human carotid lesions were analyzed for α-actin (top panels: left, ×100; right, ×400), CD40 (left middle panel, green, ×400) and TF (lower left panel, red, ×400) staining within human atherosclerotic lesions, employing immunhistochemistry (for SMC) and immunofluorescence double labeling (for CD40 and TF). The lumen of the artery is at the top of each photomicrograph. Staining employing control IgG is shown for CD40 (right middle panel, ×400) and TF (right lower panel, ×400). Analysis of atheroma from six different donors showed similar results.
Ligation of CD40 on Human Vascular SMC Induces the Transient Surface Expression of TF in Vitro
The regulation of TF in vascular SMC by CD40 ligation was further studied in vitro. Ligation of CD40 on human vascular SMC induced the transient cell surface expression of TF in vitro, whereas unstimulated cells did not express this procoagulant molecule, as demonstrated by FACS analysis (Figure 2A) ▶ . Smooth muscle cells exposed to either CD40L-positive cell membrane preparations of PMA-activated (50 ng/ml, 12 hours) T lymphocytes or recombinant human CD40L (rCD40L) showed the time-dependent induction of cell surface TF. Tissue factor expression peaked after 6 to 12 hours of coculture with the respective CD40L source, and decreased during more prolonged incubation. Addition of an anti-CD40L mAb blocked induction of TF in response to CD40 ligation. Maximal production of TF protein required the coculture of SMC with T cell membrane preparations at an equivalent of 10 T cells:1 SMC or with 10 μg/ml rCD40L (Figure 2) ▶ .
Figure 2.

CD40L induces tissue factor in human vascular smooth muscle cells in vitro. A: FACS analysis for human TF on vascular SMC, cultured 24 hours before the experiment in IT medium and subsequently for the indicated times with cell-membrane preparations of PMA-activated (50 ng/ml, 12 hours) T lymphocytes (left panels) or recombinant CD40L (10 μg/ml rCD40L; right panels) in the absence or presence of the α-CD40L mAb (α-CD40L, 10 μg/ml). Staining performed with control IgG is shown as lines. B: Lysates of human vascular SMC, previously cultured 24 hours in IT medium, then incubated for 24 hours with the respective concentrations of recombinant human CD40 ligand (rCD40L) in the absence or presence of the α-CD40L mAb (α-CD40L, 10 μg/ml), were analyzed for TF protein expression by ELISA (upper panel). TF activity was analyzed in membrane-preparations of human vascular SMC (lower panel), as described in Materials and Methods. Data shown are representative of three experiments performed with cells of different origins.
Ligation of CD40 on Human Vascular SMC Induces the Transient Expression of TF Protein and Activity
The induction of TF protein by CD40 engagement was further evaluated in lysates of cultured vascular SMC. In accordance with the findings of the FACS analysis, stimulation of vascular SMC with rCD40L induced the cell-associated expression of TF as determined by ELISA in a concentration-dependent manner. Lysates of unstimulated, quiescent SMC contained little or no detectable TF. Human rCD40L, however, induced the expression of TF protein in a concentration-dependent manner, requiring at least 1 to 3 μg/ml and achieving an 11.2 ± 1.1-fold increase employing 10 μg/ml rCD40L (Figure 2B ▶ , upper panel). Also in accordance with the FACS analysis, maximal induction of TF occurred after approximately 6 to 12 hours of stimulation with CD40L and prolonged incubation decreased TF protein (data not shown). Presence of the anti-CD40L antibody markedly reduced this induction of TF, indicating the specificity of the CD40L-mediated mechanism (Figure 2B ▶ , upper panel). Furthermore, addition of a TF blocking antibody during the ELISA abolished the detection of TF, demonstrating the specificity of the assay used.
Because the presence of TF protein does not necessarily establish its biological function, we further determined the procoagulant activity of SMC. In accordance with the findings at the protein level, CD40L induced the expression of TF activity in a similar concentration- and time-dependent manner. Because the use of total cell lysates yielded high background signals in the activity assay, membrane preparations of human vascular SMC were used. Maximal activity levels were obtained after 6 to 12 hours of stimulation with the ligand (data not shown). Membrane preparations of human vascular SMC stimulated with 10 μg/ml rCD40L contained 4.8 ± 1.2-fold more TF activity than unstimulated cells, as determined by comparison with recombinant TF (Figure 2B ▶ , lower panel).
Ligation of CD40 on Human Vascular SMC Does Not Affect the Expression of TFPI
Because the activity of TF in vivo depends on its balance with endogenous inhibitors, we further analyzed whether CD40 engagement alters the expression of TFPI. Stimulation of human vascular SMC with rCD40L did not affect the constitutive expression of TFPI-1, independent of the concentrations (0.01 to 10 μg/ml rCD40L; Figure 3A ▶ ) or time of application (0.5 to 48 hours, data not shown) of CD40L. In accordance with the findings on protein expression, ligation of CD40L did not affect TFPI activity in lysates (data not shown) or membrane preparations of human vascular smooth muscle cells (Figure 3B) ▶ .
Figure 3.

CD40L does not affect expression of TFPI in human vascular SMC. Human vascular SMC, cultured 24 hours before the experiment in IT medium, were incubated for 24 hours with the respective concentrations of rCD40L in the absence or presence of the anti-CD40L mAb (α-CD40L, 10 μg/ml) and analyzed for TFPI expression by ELISA (A) as well as TFPI-activity assay (B). Recombinant TFPI-1 was applied for control purposes. Data shown are representative of three and seven experiments, respectively, performed with cells of different origins.
Discussion
The present study demonstrates the novel finding that the CD40/CD40L-signaling pathway regulates TF protein and activity in human vascular SMC. Ligation of the receptor CD40 by native as well as recombinant human CD40 ligand augments the expression of TF protein and activity, whereas engagement of CD40 did not affect the constitutive expression of TFPI.
Triggering blood coagulation precipitates the major acute manifestations of the chronic inflammatory disease atherosclerosis. Thrombotic complications of atheroma involve various cellular components. Indeed, SMC and macrophages furnish the major source of the potent procoagulant TF. 1,10,11,35,38 Several studies demonstrated that nearly every atherosclerotic lesion analyzed expressed TF mRNA, protein, and activity. 10 Although much information exists regarding regulation of TF produced by macrophages, the regulation of TF expression in human vascular SMC remains poorly understood. Previous reports focused on soluble mediators including proinflammatory cytokines and growth factors. 17,39 However, these molecules do not explain TF induction observed after cell contact, 18 for example with platelets, recently found to express functional CD40L. 40 We and others have recently presented evidence implicating the CD40 signaling pathway in atherogenesis, because it mediates functions such as cytokine, adhesion molecule, and matrix metalloproteinase expression on atheroma-associated cells in vitro, 19,30,33,37,41-47 and affects atherogenesis in vivo. 48 The present study demonstrates that CD40L can indeed mediate the expression of TF protein and activity in human vascular SMC, providing a potential mediator of cell contact-dependent induction of this procoagulant activity. The time-dependent induction of TF via CD40 signaling resembles that found previously with soluble mediators. 17
CD40 ligand may arise from multiple sources within atheromatous tissue. We and others recently demonstrated that CD40L is expressed on endothelial cells and SMC as well as macrophages. 19,49,50 Furthermore, T lymphocytes, cells found early in atherogenesis, also bear CD40L. 19 Finally, recent studies by others demonstrated that platelets can also mediate functions via CD40 ligand. 40 Indeed, platelets can induce TF expression on SMC as well as on endothelial cells and macrophages, as demonstrated by several groups. 18,51-54
In addition to its cellular location, TF also occurs in the extracellular pool of the lipid core within the atherosclerotic lesion. 10 SMC constitute a large portion of the cells in many progressing atheroma. These cells may undergo apoptosis during the evolution toward more vulnerable lesions. 55 SMC-derived TF thus might contribute to the acellular pool of plaques as a deposit by activated and/or dying SMC within the lipid core. On the other hand, TF can also promote cell proliferation by augmenting growth factor expression. 9 Thus, SMC-derived TF may participate in both the proliferative phase of atherogenesis as well as the later stage of lesion destabilization, associated with SMC drop-out.
Within the core of atheroma only a subpopulation of macrophages expresses TF. 1 In contrast, the present results show a homogenous expression of tissue factor by SMC in vitro and in situ. These results may have important implications for thrombosis caused by superficial erosion of atheroma during which blood may contact intimal SMC but not the central core of the lesion, as in rupture of the plaque’s fibrous cap. 56,57
TF-associated procoagulant activity depends on the expression of TFPI. TFPI-1 interacts, probably via its Kunitz domain 2, with the ternary procoagulant complex of TF, factor VIIa, and factor X, forming an inactive quaternary complex. Originally, TFPI-1 was described in plasma as a complex with lipoproteins. Recent 58 as well as our own unpublished studies, however, revealed its presence within human atherosclerotic lesions. Thus, imbalance of procoagulant and inhibitory activity might determine the thrombogenicity of lesions.
The present study demonstrates increased TF protein and activity combined with unaltered TFPI protein and activity, indicating that the balance of TF and TFPI in human vascular SMC is shifted toward increased procoagulant activity after CD40 activation.
The findings of the current study, combined with the observation that CD40 ligation also induces TF on macrophages and endothelial cells, 37,51-53 demonstrates the capability of this cell-associated activator to regulate the thrombotic potential of human atherosclerotic lesion.
Acknowledgments
We thank Maria Muszynski, Irina Chulsky, Eugenia Shvartz, and Elissa Simon-Morrissey (Brigham and Women’s Hospital) for skillful technical assistance, and Karen Williams for editorial assistance.
Footnotes
Address reprint requests to Peter Libby, Vascular Medicine and Atherosclerosis Unit, Cardiovascular Division, Department of Medicine, Brigham and Women’s Hospital, Harvard Medical School, 221 Longwood Avenue, LMRC 307, Boston, MA 02115. E-mail: plibby@rics.bwh.harvard.edu.
Supported in part by grants of the National Heart, Lung and Blood Institute (HL-56985 to P. L.) and performed during the tenure of U. S. as Paul Dudley White Fellow of the American Heart Association.
References
- 1.Wilcox JN, Smith KM, Schwartz SM, Gordon D: Localization of tissue factor in the normal vessel wall and in the atherosclerotic plaque. Proc Natl Acad Sci USA 1989, 86:2839-2843 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Fuster V, Badimon I, Badimon JJ, Chesebro JH: The pathogenesis of coronary artery disease and the acute coronary syndromes. N Engl J Med 1992, 326:242-250 [DOI] [PubMed] [Google Scholar]
- 3.Davies MJ, Thomas AC: Plaque fissuring: the cause of acute myocardial infarction, sudden ischemic death, and crescendo angina. Br Heart J 1985, 53:363-373 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Nemerson Y: Tissue factor and hemostasis. Blood 1988, 71:1-8 [PubMed] [Google Scholar]
- 5.Edgington TS, Mackman N, Brand K, Ruf W: The structutal biology of expression and function of tissue factor. Thromb Haemost 1991, 66:67-79 [PubMed] [Google Scholar]
- 6.Mueller BM, Ruf W: Requirement for binding of catalytically active factor VIIa in tissue factor-dependent experimental metastasis. J Clin Invest 1998, 101:1372-1378 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ott I, Fischer EG, Miyagi Y, Mueller BM, Ruf W: A role for tissue factor in cell adhesion and migration mediated by interaction with actin-binding protein 280. J Cell Biol 1998, 140:1241-1253 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sato Y, Asada Y, Marutsuka K, Hatakeyama K, Sumiyoshi A: Tissue factor induces migration of cultured aortic smooth muscle cells. Thromb Haemost 1996, 75:389-392 [PubMed] [Google Scholar]
- 9.Carmeliet P, Collen D: Tissue factor. Int J Biochem Cell Biol 1998, 30:661-667 [DOI] [PubMed] [Google Scholar]
- 10.Taubman MB, Fallon JT, Schecter AD, Giesen P, Mendlowitz M, Fyfe BS, Marmur JD, Nemerson Y: Tissue factor in the pathogenesis of atherosclerosis. Thromb Haemost 1997, 78:200-204 [PubMed] [Google Scholar]
- 11.Hatekeyama K, Asada Y, Marutsuka K, Sato Y, Kamikubo K, Sumiyoshi A: Localization and activity of tissue factor in human aortic atherosclerotic lesions. Atherosclerosis 1997, 133:213-219 [DOI] [PubMed] [Google Scholar]
- 12.Annex BH, Denning SM, Channon KM, Sketch MH, Jr, Stack RS, Morrissey JH, Peters KG: Differential expression of tissue factor protein in directional atherectomy specimens from patients with stable and unstable coronary syndromes. Circulation 1995, 91:619-622 [DOI] [PubMed] [Google Scholar]
- 13.Losordo DW, Rosenfield K, Pieczek A, Baker K, Harding M, Isner JM: How does angioplasty work? Serial analysis of human iliac arteries using intravascular ultrasound. Circulation 1992, 86:1845-1858 [DOI] [PubMed] [Google Scholar]
- 14.Nath FC, Muller DW, Ellis SG, Rosenschein U, Chapekis A, Quain L, Zimmerman C, Topol EJ: Thrombosis of a flexible coil coronary stent: frequency, predictors, and clinical outcome. J Am Col Cardiol 1993, 21:622-627 [DOI] [PubMed] [Google Scholar]
- 15.Carrozza JP, Baim DS: Complications of directional coronary atherectomy: incidence, causes, and management. Am J Cardiol 1993, 72:47E-54E [DOI] [PubMed] [Google Scholar]
- 16.Channon KM, Fulton GJ, Davies MG, Peters KG, Ezekowitz MD, Hagen PO, Annex BH: Modulation of tissue factor protein expression in experimental venous bypass grafts. Arterioscler Thromb Vasc Biol 1997, 17:1313-1319 [DOI] [PubMed] [Google Scholar]
- 17.Taubman MB: Tissue factor regulation in vascular smooth muscle: a summary of studies performed during in vivo and in vitro models. Am J Cardiol 1993, 72:55C-60C [DOI] [PubMed] [Google Scholar]
- 18.Smariga PE, Maynard JR: Platelet effects on tissue factor and fibrinolytic inhibition of cultured human fibroblasts and vascular cells. Blood 1982, 60:140-147 [PubMed] [Google Scholar]
- 19.Mach F, Schonbeck U, Sukhova GK, Bourcier T, Bonnefoy JY, Pober JS, Libby P: Functional CD40 ligand is expressed on human vascular endothelial cells, smooth muscle cells, and macrophages: implications for CD40-CD40 ligand signaling in atherosclerosis. Proc Natl Acad Sci USA 1997, 1936, 94:1931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Mallett S, Barclay AN: A new superfamily of cell surface proteins related to the nerve growth factor receptor. Immunol Today 1991, 12:220-223 [DOI] [PubMed] [Google Scholar]
- 21.Smith CA, Farrah T, Goodwin RG: The TNF receptor superfamily of cellular, and viral proteins: activation, costimulation, and death. Cell 1994, 76:959-962 [DOI] [PubMed] [Google Scholar]
- 22.Armitage RJ: Tumor necrosis factor receptor superfamily members and their ligands. Curr Opin Immunol 1994, 6:407-413 [DOI] [PubMed] [Google Scholar]
- 23.Foy TM, Aruffo A, Bajorath J, Buhlmann JE, Noelle RJ: Immune regulation by CD40 and its ligand GP39. Annu Rev Immunol 1996, 14:591-617 [DOI] [PubMed] [Google Scholar]
- 24.Van Kooten C, Banchereau J: CD40-CD40 ligand: a multifunctional receptor-ligand pair. Adv Immunol 1996, 61:1-77 [DOI] [PubMed] [Google Scholar]
- 25.Grewal IS, Flavell RA: CD40, and CD154 in cell-mediated immunity. Annu Rev Immunol 1998, 16:111-135 [DOI] [PubMed] [Google Scholar]
- 26.Noelle RJ, Foy TM, Shepherd DM, Stamenkovic I, Ledbetter JA, Aruffo A: A 39-kD protein on activated helper T cells binds CD40, and transduces the signal for cognate activation of B cells. Proc Natl Acad Sci USA 1992, 98:6550-6554 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lederman S, Yellin MJ, Krichevsky A, Belko J, Lee JJ, Chess L: Identification of a novel surface protein on activated CD4+ T cells that induces contact-dependent B cell differentiation (help). J Exp Med 1992, 175:1091-1101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Laman JD, Claassen E, Noelle RJ: Functions of CD40 and its ligand, gp39 (CD40L). Crit Rev Immunol 1996, 16:59-108 [DOI] [PubMed] [Google Scholar]
- 29.Noelle RJ: CD40, and its ligand in cell-mediated immunity. Agents Actions Suppl 1998, 49:17-22 [DOI] [PubMed] [Google Scholar]
- 30.Schonbeck U, Mach F, Sukhova GK, Murphy C, Bonnefoy JY, Fabunmi RP, Libby P: Regulation of matrix metalloproteinase expression in human vascular smooth muscle cells by T lymphocytes: a role for CD40 signaling in plaque rupture? Circ Res 1997, 81:448-454 [DOI] [PubMed] [Google Scholar]
- 31.Mazzei GJ, Edgerton MD, Losberger C, Lecoanet-Henchoz S, Graber P, Durandy A, Gauchat J-F, Bernard A, Allet B, Bonnefoy J-Y: Recombinant soluble trimeric CD40 ligand is biologically active. J Biol Chem 1995, 270:7025-7028 [DOI] [PubMed] [Google Scholar]
- 32.Pullen SS, Labadia ME, Ingraham RH, McWhirter SM, Everdeen DS, Alber T, Crute JJ, Kehry MR: High-affinity interactions of tumor necrosis factor receptor-associated factors (TRAFs) and CD40 require TRAF trimerization and CD40 multimerization. Biochemistry 1999, 38:10168-10177 [DOI] [PubMed] [Google Scholar]
- 33.Schonbeck U, Mach F, Sukhova GK, Atkinson E, Levesque E, Herman M, Graber P, Basset P, Libby P: Expression of stromelysin-3 in atherosclerotic lesions: regulation via CD40-CD40 ligand signaling in vitro and in vivo. J Exp Med 1999, 189:843-853 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Libby P, O’Brien KV: Culture of quiescent arterial smooth muscle cells in a defined serum-free medium. J Cell Physiol 1983, 115:217-223 [DOI] [PubMed] [Google Scholar]
- 35.Marmur JD, Thiruvikraman SV, Fyfe BS, Guha A, Sharma SK, Ambrose JA, Fallon JT, Nemerson Y, Taubman MB: Identification of active tissue factor in human coronary atheroma. Circulation 1996, 94:1226-1232 [DOI] [PubMed] [Google Scholar]
- 36.Moreno PR, Bernardi VH, Lopez-Cuellar J, Murcia AM, Palacios IF, Gold HK, Mehran R, Sharma SK, Nemerson Y, Fuster V, Fallon J: Macrophages, smooth muscle cells, and tissue factor in unstable angina. Circulation 1996, 94:3090-3097 [DOI] [PubMed] [Google Scholar]
- 37.Mach F, Schonbeck U, Bonnefoy JY, Pober JS, Libby P: Activation of monocyte/macrophage functions related to acute atheroma complication by ligation of CD40: induction of collagenase, stromelysin, and tissue factor. Circulation 1997, 96:396-399 [DOI] [PubMed] [Google Scholar]
- 38.Moreno PR, Falk E, Palacios IF, Newell JB, Fuster V, Fallon JT: Macrophage infiltration in acute coronary syndromes: implications for plaque rupture. Circulation 1994, 90:775-778 [DOI] [PubMed] [Google Scholar]
- 39.Schecter AD, Giesen PL, Taby O, Rosenfield CL, Rossikhina M, Fyfe BS, Kohtz DS, Fallon JT, Nemerson Y, Taubman MB: Tissue factor expression in human arterial smooth muscle cells: TF is present in three cellular pools after growth factor stimulation. J Clin Invest 1997, 100:2276-2285 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Henn V, Slupsky JR, Grafe M, Anagnostopoulos I, Forster R, Muller-Berghaus G, Kroczek RA: CD40 ligand on activated platelets triggers an inflammatory reaction of endothelial cells. Nature 1998, 391:591-594 [DOI] [PubMed] [Google Scholar]
- 41.Wagner DH, Jr, Stout RD, Suttles J: Role of the CD40-CD40 ligand interaction in CD4+ T cell contact-dependent activation of monocyte interleukin-1 synthesis. Eur J Immunol 1994, 24:3148-3154 [DOI] [PubMed] [Google Scholar]
- 42.Karmann K, Hughes CC, Schechner J, Fanslow WC, Pober JS: CD40 on human endothelial cells: inducibility by cytokines and functional regulation of adhesion molecule expression. Proc Natl Acad Sci USA 1995, 92:4342-4346 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Hollenbaugh D, Mischel-Petty N, Edwards CP, Simon JC, Denfeld RW, Kiener PA, Aruffo A: Expression of functional CD40 by vascular endothelial cells. J Exp Med 1995, 182:33-40 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kiener PA, Moran-Davis P, Rankin BM, Wahl AF, Aruffo A, Hollenbaugh D: Stimulation of CD40 with purified soluble gp39 induces proinflammatory responses in human monocytes. J Immunol 1995, 155:4917-4925 [PubMed] [Google Scholar]
- 45.Kato T, Hakamada R, Yamane H, Nariuchi H: Induction of IL-12 p40 messenger RNA expression and IL-12 production of macrophages via CD40-CD40 ligand interaction. J Immunol 1996, 156:3932-3938 [PubMed] [Google Scholar]
- 46.Mach F, Schonbeck U, Fabunmi RP, Murphy C, Atkinson E, Bonnefoy JY, Graber P, Libby P: T lymphocytes induce endothelial cell matrix metalloproteinase expression by a CD40L-dependent mechanism: implications for tubule formation. Am J Pathol 1999, 154:229-238 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Schonbeck U, Mach F, Bonnefoy JY, Loppnow H, Flad HD, Libby P: Ligation of CD40 activates interleukin 1β-converting enzyme (caspase-1) activity in vascular smooth muscle and endothelial cells and promotes elaboration of active interleukin 1β. J Biol Chem 1997, 272:19569-19574 [DOI] [PubMed] [Google Scholar]
- 48.Mach F, Schonbeck U, Sukhova GK, Atkinson E, Libby P: Reduction of atherosclerosis in mice by inhibition of CD40 signalling. Nature 1998, 394:200-203 [DOI] [PubMed] [Google Scholar]
- 49.Briscoe DM, Alexander SI, Lichtman AH: Interactions between T lymphocytes and endothelial cells in allograft rejection. Curr Opin Immunol 1998, 10:525-531 [DOI] [PubMed] [Google Scholar]
- 50.Gaweco AS, Wiesner RH, Yong S, Krom R, Porayko M, Chejfec G, McClatchey KD, Van Thiel DH: CD40L (CD154) expression in human liver allografts during chronic ductopenic rejection. Liver Transpl Surg 1999, 5:1-7 [DOI] [PubMed] [Google Scholar]
- 51.Slupsky JR, Kalbas M, Willuweit A, Henn V, Kroczek RA, Muller-Berghaus G: Activated platelets induce tissue factor expression on human umbilical vein endothelial cells by ligation of CD40. Thromb Haemost 1998, 80:1008-1014 [PubMed] [Google Scholar]
- 52.Zhou L, Stordeur P, de Lavareille A, Thielemans K, Capel P, Goldman M, Pradier O: CD40 engagement on endothelial cells promotes tissue factor-dependent procoagulant activity. Thromb Haemost 1998, 79:1025-1028 [PubMed] [Google Scholar]
- 53.Miller DL, Yaron R, Yellin MJ: CD40L-CD40 interactions regulate endothelial cell surface tissue factor, and thrombomodulin expression. J Leukoc Biol 1998, 63:373-379 [DOI] [PubMed] [Google Scholar]
- 54.Gregory SA, Edgington TS: Tissue factor induction in human monocytes. Two distinct mechanisms displayed by different alloantigen-responsive T cell clones. J Clin Invest 1985, 76:2440-2445 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Geng YJ, Libby P: Evidence for apoptosis in advanced human atheroma: colocalization with interleukin-1β-converting enzyme. Am J Pathol 1995, 147:251-266 [PMC free article] [PubMed] [Google Scholar]
- 56.Farb A, Burke A, Tang A, Liang Y, Mannan P, Smialek J, Virmani R: Coronary plaque erosion without rupture into a lipid core: a frequent cause of coronary thrombosis in sudden coronary death. Circulation 1996, 93:1354-1363 [DOI] [PubMed] [Google Scholar]
- 57.van der Wal AC, Becker AE, van der Loos CM, Das PK: Site of intimal rupture or erosion of thrombosed coronary atherosclerotic plaques is characterized by an inflammatory process irrespective of the dominant plaque morphology. Circulation 1994, 89:36–44 [DOI] [PubMed]
- 58.Caplice NM, Mueske CS, Kleppe LS, Simari RD: Presence of tissue factor pathway inhibitor in human atherosclerotic plaques is associated with reduced tissue factor activity. Circulation 1998, 98:1051-1057 [DOI] [PubMed] [Google Scholar]