

Abstract
The isocyclic ring of bacteriochlorophyll (BChl) is formed by the conversion of Mg-protoporphyrin monomethyl ester (MPE) to protochlorophyllide (PChlide). Similarities revealed by blast searches with the putative anaerobic MPE-cyclase BchE suggested to us that this protein also uses a cobalamin cofactor. We found that vitamin B12 (B12)-requiring mutants of the bluE and bluB genes of Rhodobacter capsulatus, grown without B12, accumulated Mg-porphyrins. Laser desorption/ionization time-of-flight (LDI-TOF) MS and NMR spectroscopy identified them as MPE and its 3-vinyl-8-ethyl (mvMPE) derivative. An in vivo assay was devised for the cyclase converting MPE to PChlide. Cyclase activity in the B12-dependent mutants required B12 but not protein synthesis. The following reaction mechanism is proposed for this MPE-cyclase reaction. Adenosylcobalamin forms the adenosyl radical, which leads to withdrawal of a hydrogen atom and formation of the benzylic-type 131-radical of MPE. Withdrawal of an electron gives the 131-cation of MPE. Hydroxyl ion attack on the cation gives 131-hydroxy-MPE. Withdrawal of three hydrogen atoms leads successively to 131-keto-MPE, its 132-radical, and cyclization to PChlide.
The Mg-protoporphyrin monomethyl ester (MPE)-cyclase is an enzyme activity converting MPE to protochlorophyllide (PChlide) in the Mg branch of the tetrapyrrole biosynthetic pathway leading to chlorophyll and bacteriochlorophyll (BChl) (Fig. 1). Cobalamin is formed from uroporphyrinogen, the first tetrapyrrole intermediate. During evolution cobalt tetrapyrrole may thus have preceded the formation of chlorophyll and an oxygen atmosphere. The cobalt tetrapyrroles still play an essential role in anaerobic reactions involved in energy production. Many photosynthetic bacteria form their BChl only under anaerobic conditions. Consequently, as shown in ref. 1, the 131-oxo group of the isocyclic ring in Rhodobacter sphaeroides is derived from water and not molecular oxygen, as it is in higher plants (2). Aerobic cyclase activity has been demonstrated in higher plants (3) and Synechocystis (4). In both cases a soluble and a membrane fraction were shown to be required, but purification and identification of the proteins involved were not reported. In vitro anaerobic cyclase activity has not been reported.
Figure 1.

Some intermediates of the BChl and cobalamin biosynthetic pathway. The role of the MPE-cyclase is shown.
MPE accumulates in transposon mutants of the Rhodobacter capsulatus bchE gene, suggesting that the encoded protein is required for MPE-cyclase activity (5). Sequence similarities of BchE to P-methylase from Streptomyces hygroscopicus suggested to us a cobalamin requirement for the MPE-cyclase. Disruption of the vitamin B12 (B12) biosynthetic genes bluE and bluB in R. capsulatus strongly inhibited formation of BChl and proteins of the photosynthetic apparatus (6, 7).
We show here that MPE, an intermediate of the BChl biosynthetic pathway, accumulates in B12-requiring mutants carrying bluE and bluB genes. Furthermore, MPE can be efficiently converted to PChlide by these mutants, but only when a form of cobalamin is provided.
Materials and Methods
Chemicals.
Mg-protoporphyrin (MP) and its dimethyl ester (MDE) were obtained from Porphyrin Products (Logan, UT). MPE and 3-vinyl-8-ethyl-MPE (mvMPE) were prepared from extracts of R. capsulatus kanamycin transposon mutant bchE and described below. PChlide was prepared by treatment of 7-day dark-grown barley shoots with 10 mM δ-aminolevulinate overnight.
Bacterial Strains and Cell Growth.
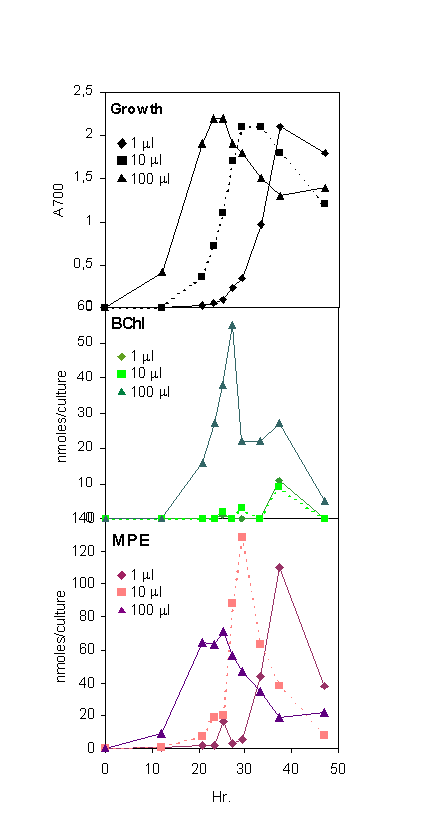
Cell growth of capsule-less wild-type R. capsulatus 37b4, and derived transposon Tn7 and Ω interposon mutants AH2 and BB1, in the bluE gene and the bluB gene, respectively (6, 7), was measured at 700 nm. R. capsulatus DB575, a kanamycin transposon mutant of the bchE gene (5, 8) was grown with shaking at 140 rpm, at 30°C in the light (fluorescent white light, Philips TLD 18-W) or in the dark in 100-ml conical flasks containing 50 ml of medium. For bluE and bluB mutant strains, a minimal malate medium was used (9), with or without the addition of 10 μg⋅ml−1 B12 (this amount of B12 is considered to be of critical importance). Cells were prepared for growth without B12 as follows. Cells (bluE or bluB) were grown for 2 days with B12, during which time the cells attained stationary phase and a purple reddish color. The cells were centrifuged for 20 min at 2,200 × g and resuspended in 1/10 vol of medium without B12. Inocula sizes of 10–50 μl (bluE) or 10–40 μl (bluB) were used to observe optimal Mg-porphyrin accumulation with a convenient culture time of 25–30 h. Larger inocula apparently contained enough residual B12 to allow more BChl formation (see Figs. 7 and 8 in the supplementary data at www.pnas.org). For large-scale pigment production the bchE mutant strain was grown in a medium with 0.1% peptone and 0.1% yeast extract (10).
Extraction and Quantitative Determination of MPE, mvMPE, PChlide, and BChl.
One milliliter of cell culture, or the in vitro assay mixture, was centrifuged briefly in orange Eppendorf tubes (to protect light-sensitive Mg-porphyrins). The pelleted cells were vigorously resuspended in 50 μl of the supernatant. Pigments were extracted from the cells by addition of 0.75 ml of acetone/0.35 M ammonia (4:1, vol/vol) and 1 ml of hexane. BChl in the hexane layer was calculated by using an ɛ of 91.1 mM−1⋅cm−1 at 770 nm (11). The ammoniacal acetone phase was diluted 20-fold and pigments were estimated with a Perkin–Elmer LS50B luminescence spectrometer. For MPE and mvMPE determination, excitation was at 418 nm and for PChlide at 440 nm. Emission intensities for MPE and mvMPE, at 595 nm, were standardized by using MP whose concentration was determined (12) with an 8452A Hewlett Packard diode array spectrophotometer. PChlide emission at 633 nm is presented directly.
Purification of Large Amounts of MPE from bchE and Mg-porphyrins from bluE or bluB Strains Grown Without B12.
A modification of the method of Granick (13) was used. Centrifuged cells, from 5 liters of medium, were resuspended in a small volume of the supernatant and extracted with 100 ml of ammoniacal acetone and hexane. The aqueous phase was adjusted to pH 7.2 with saturated KH2PO4, and 50 ml of diethyl ether and saturated KCl (30 μl per ml of aqueous phase) were added. The ether phase was separated and the ether extraction was repeated after addition of acetone. The combined ether extracts were then washed successively with M/15 potassium phosphate, pH 7.4, pH 7.7, and pH 8.0 and with water. The ether was evaporated to 50 μl in a stream of nitrogen. The residue was dissolved in 1 ml of ammoniacal acetone and the deep red solution was extracted three times with an equal volume of petroleum ether (bp 60–80°C) to remove most of the remaining carotenoid. The aqueous acetone phase was then evaporated to a small volume in a stream of nitrogen, and remaining water was removed under reduced pressure. The pigment, about 1 mg, was stored at −20°C until used.
Reversed-Phase C18 TLC.
HPTLC-Fertigplatten RP-18, WF254S TLC plates (Merck Art 13124) were prerun in acetone/1 M NH3 (9:1, vol/vol). After 20- to 30- min air-drying of the plates, samples were applied and the Mg-porphyrin was separated by development in acetone/0.1 M NH3 (4.5:5.5, vol/vol) for 45 min. The Mg-porphyrins were visualized under UV light. The separated Mg-porphyrins were extracted from the reversed-phase material by repeated extractions with small volumes of neutralized dimethyl sulfoxide.
Laser Desorption/Ionization Time-of-Flight (LDI-TOF) MS.
LDI-TOF MS was performed with a Bruker Reflex III MALDI-MS. Matrix, when used, was α-cyanohydroxycinnamic acid. Samples were purified by solvent extraction and, in some cases, by chromatography. They were dissolved in 70% (vol/vol) acetonitrile/30% water (containing 0.1% trichloroacetic acid when matrix was used) and applied to the sample holder.
NMR Spectroscopy.
NMR spectroscopy was carried out with a Varian UNITY INOVA 500 operating at 500.1 MHz for 1H. Samples were prepared in 600 μl of deuterated methanol at 25°C. Chemical shifts are given relative to methanol 3.35 ppm for 1H and 47.1 ppm for 13C. The double quantum filtered phase-sensitive correlated spectroscopy (COSY) spectrum was acquired by using the Varian standard pulse program TNDQCOSY (14). The nuclear Overhauser spectra were acquired by using the Varian standard pulse program TNNOESY (15), with mixing times of 200 and 400 ms. All spectra were assigned by using the computer program pronto (16), which allows the simultaneous display of several two-dimensional spectra.
Determination of in Vivo Cyclase Activity of Wild-Type and B12-Requiring Mutant R. capsulatus.
Wild-type R. capsulatus was grown in 500 ml of medium with shaking at 150 rpm to OD700 = 0.8, OD770 = 0.2 in the light at 30°C, or as given. bluE mutant cells were grown from an inoculum of 100 μl in 500 ml of medium without B12. Cells were harvested by centrifugation at 30,000 × g for 5 min and resuspended, with or without a wash, in 1/100 vol of 50 mM potassium phosphate buffer, pH 7.3. Cells (200 μl) were assayed in closed Eppendorf tubes with slow shaking at 30°C for 1 h in the dark. Exogenous mvMPE + MPE, purified by chromatography, from the bluB mutant, or endogenous mvMPE + MPE was used as substrate.
Database Searches and Pileups.
Searches were performed by using the blast and Ψ-blast (17) programs at the www.ncbi.nlm.nih.gov/BLAST site. fasta searches and Pileups were run on a local server, using the Wisconsin Package, Version 9.1, Genetics Computer Group (GCG; Madison, WI). Alignments were also adjusted manually. Preliminary sequence data for the bchE homologue of Chlorobium tepidum were obtained from The Institute for Genomic Research web site at http://www.tigr.org.
Results and Discussion
Sequence Similarity of bchE Gene Product to a Methylcobalamin-Using P-Methylase.
The involvement of a cobalamin cofactor in the MPE-cyclase reaction was first suggested to us by resemblance of the primary structure of Rhodobacter BchE to a methylcobalamin-requiring P-methylase from Streptomyces hygroscopicus (18). Position-iterated blast (Ψ-blast) of the BchE homologue from Chlorobium tepidum revealed a highly significant similarity to other bchE gene products and to the P-methylase (Fig. 2). Homologues of the Rhodobacter capsulatus cyclase gene, bchE, are thus additionally found in the photosynthetic bacteria Chlorobium tepidum, Synechocystis, and Heliobacillus mobilis.
Figure 2.

Pileup of BchE homologs and P-methylase. Organism, accession no., Swiss Prot or other database name, and Ψ-blast E values (exponential notation) with C. tepidum BchE as search profile (7 iterations) are shown at lower right. ANCOPROX, anaerobic coproporphyrinogen oxidase.
A role of B12 in anaerobic cyclase activity was also suggested to us by two observations from the literature. In Chlorobium liminicola strain 1230 (B12-requiring), BChl a and c decreased sharply under B12 limitation (19). Furthermore, treatment of Chlorobium cells with nitrous oxide and other anesthetics leads to MPE accumulation and inhibition of antennae-associated BChl d formation (20). Nitrous oxide is well known as an inhibitor of cobalamin-containing enzymes (21–24). Fig. 2 also shows that, except for the missing N-terminal domain, (which may bind B12), another similar protein is the anaerobic coproporphyrinogen oxidase (ANCOPROX).
Effect of B12 Depletion on Mg-Porphyrin Accumulation.
To test the hypothesis that the MPE-cyclase requires a cobalamin cofactor, we investigated the effect of B12 deprivation on pigment synthesis in R. capsulatus. Accordingly, the B12-requiring mutants of R. capsulatus bluE or bluB were investigated. Cells were grown without B12 as described. Cultures of R. capsulatus form photosynthetic pigments only under anaerobic growth conditions. The mutants can grow without B12 and form a little BChl (see Fig. 7 in the supplementary data at www.pnas.org) even after repeated subculture without B12. Possibly, some cells may lose the kanamycin transposon and consequently form B12. Minute amounts of B12 in the culture from these cells may be required for expression of gene products important for the observation of the results in this paper. Under anaerobic growth conditions without B12 the blu mutants have a pink color (blush) that is not caused by just carotenoid formation. The crude culture extracts showed characteristic emission spectral maxima (595 nm) for excitation at 418 nm, suggesting Mg-porphyrin accumulation. A priori it might have been expected that B12-deficient cells would lack methionine, if they use only a B12-dependent methionine synthase and therefore are unable to carry out the S-adenosylmethionine-requiring MPE transferase reaction. MP should therefore accumulate. However, addition of methionine did not restore BChl formation to the B12-requiring mutants at the bluE or bluB loci (G. Klug, personal communication).
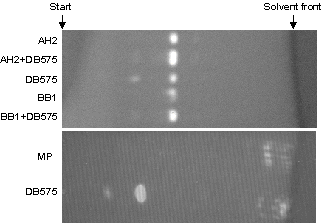
Preliminary Identification of Accumulated Metalloporphyrins by Reversed-Phase Chromatography.
Reversed-phase chromatography of the ammoniacal acetone layer from crude extracts of bluE and bluB mutant strains indicated that the accumulated metalloporphyrins had a low Rf, similar to MPE prepared from the R. capsulatus bchE mutant, compared with the dicarboxylic MP, which ran near the solvent front. Cochromatography of metalloporphyrins from bluE and bluB mutants with the MPE extracted from the R. capsulatus mutant bchE showed only one main fluorescent spot (see Fig. 9 in the supplementary data at www.pnas.org). These experiments showed that the main accumulated metalloporphyrins in the B12-requiring mutants were not like MP but were very similar or identical to the MPE from the bchE mutant. Spectroscopic characterization of the metalloporphyrins was similar to that of MPE from the bchE mutant. After chromatographic purification, absorption spectra of the main porphyrins from the B12-requiring mutants were very similar to the MPE from the bchE mutant. (Table 1 and Fig. 10 in the supplementary data at www.pnas.org). The blue-shifted Soret maximum relative to MP suggests the presence of the less-conjugated monovinyl-Mg-protoporphyrin entity in all of the mutants.
Table 1.
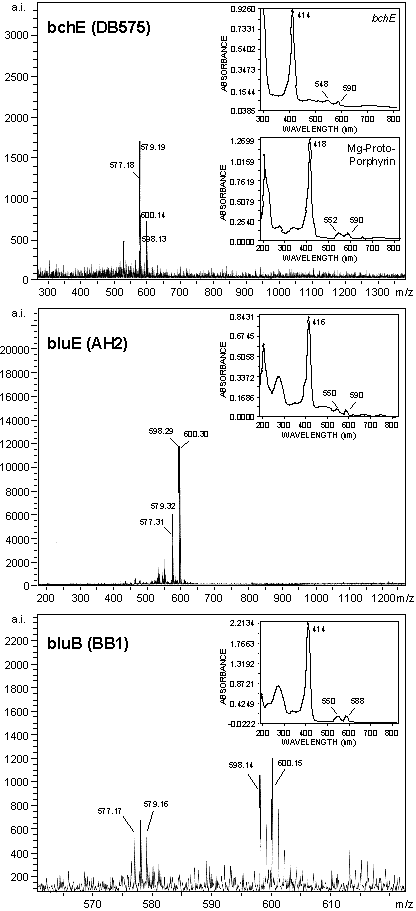
Absorption maxima and mass spectral properties of isolated metalloporphyrins of bluE, bluB, and bchE mutants
| Source of compound | Spectral maxima, nm | LDI-TOF MS mass ions
|
Molecular weight | ||
|---|---|---|---|---|---|
| (M)+ | (M+H)+ | Substance | |||
| bluE | 416 (S) | 598.3 | 598.6 | MPE | |
| 551, 590 | 600.3 | 600.6 | mvMPE | ||
| 577.3 | 576.2 | PME | |||
| 579.3 | 578.2 | mvPME | |||
| bluB | 415 (S) | 598.1 | 598.6 | MPE | |
| 550, 588 | 600.1 | 600.6 | mvMPE | ||
| 577.2 | 576.2 | PME | |||
| 579.2 | 578.2 | mvPME | |||
| bchE | 414 (S) | 598.1 | 598.6 | MPE | |
| 549, 590 | 600.1 | 600.6 | mvMPE | ||
| 577.2 | 576.2 | PME | |||
| 579.2 | 578.2 | mvPME | |||
| MP | 419 (S) | ND | 584.6 | MP | |
| 551, 589 | |||||
(M)+ is for metalloporphyrins; (M+H)+ is for porphyrins. S, Soret band; PME, protoporphyrin monomethyl ester; ND, not determined.
Identification of the Metalloporphyrins in the bchE, bluE, and bluB Mutants as Mg-Porphyrins.
Mass peaks given in Table 1 and shown in Fig. 10 of the supplementary data at www.pnas.org) for MPE from the bchE mutant, used as a standard, have identical mass to the molecular ions, (M)+, of MPE and mvMPE. Mass peaks values of the respective metal-free porphyrins were also observed as the (M+H)+ ions. For all mutants the difference in mass between each pair of metallated (M)+, and metal-free (M+H)+ porphyrin species is 21 units. Three extra mass units should be added. Two extra protons attach to two pyrrole nitrogens after loss of the metal and a third is added to produce the (M+H)+ species. This gives the mass of the metal as 24 units, identifying it as magnesium (atomic weight 24.3). It should be noted that the “MPE” accumulated by the bchE mutant (5) is shown by this analysis to consist of a mixture of MPE and its monovinyl derivative. As indicated, identical results were obtained with the porphyrins isolated from the bluE and bluB mutants. When matrix was used with samples dissolved in acid acetonitrile, only the two (M+H)+ ions reported were observed for all mutants (data not shown). These results establish that Mg-porphyrins with the molecular weights of MPE and mvMPE accumulate in the bluE and bluB mutants, when B12 is lacking. Although the samples for mass spectra were not purified by chromatography, no other peaks attributable to other metalloporphyrins or porphyrins were found. However, peaks attributable to fatty acids or carotenoids were present. LDI-TOF MS spectra of porphyrins without the use of classical matrices have previously been reported (25).
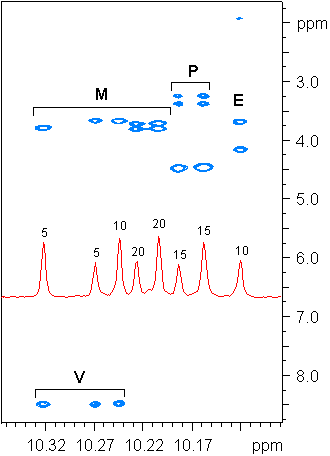
MPE and mvMPE from the bluE and bluB mutants were confirmed by NMR spectroscopy. The assignments of 1H data were carried out as for heme systems (26). This assignment procedure relies on the nuclear Overhauser effect (NOE) between adjacent substituents of the ring system—e.g., between 2-CH3 and the 20-meso proton and likewise between 2-CH3 and the 31 proton of the 3-vinyl group (Fig. 3). These NOEs form a self-consistent network around the ring system. The difference in the structure of MPE and mvMPE is clearly seen by the differences in chemical shifts. That is, the 8-ethyl group of mvMPE gives clearly distinct shifts from the corresponding vinyl group at position 3 (Table 2), and the position is established by the NOE correlations (see Fig. 11 in the supplementary data at www.pnas.org). The methyl ester positions of MPE and mvMPE were assigned with the help of commercial MP and MDE, whose assignments agree well with other data (27, 28) when differences of solvent and complex form are considered. These results clearly show that only the propionic acid group at position 13 on the ring is methylated. The 132-CH2 of methyl propionate in MPE and mvMPE shows distinct chemical shifts for 1H (3.39 ppm) and 13C (36.2 ppm). These shifts are in good agreement with those for the corresponding atoms of the methyl propionate at the 13 position in the MDE, 3.37 ppm and 36.1 ppm (Table 2). In contrast, the 172-CH2 chemical shifts (3.24 ppm and 39.2 ppm) are in good agreement with the corresponding shifts in MP (3.19 ppm and 40.5 ppm). Based on the analysis above, the MS and NMR data lead to the structures for MPE and mvMPE shown in Fig. 6.
Figure 3.

The down-field region of the 1H NMR spectra of the MPE (major component) and mvMPE (minor component) from the bluE mutant and of Mg-protoporphyrin and its diester (MDE). The meso protons are indicated for the peaks.
Table 2.
Assigned 1H and 13C chemical shift data for the isolated MPE and mvMPE and for comparison the data for MP and MDE
| Group | δ, ppm
|
|||||||
|---|---|---|---|---|---|---|---|---|
| mvMPE
|
MPE
|
MDE
|
MP
|
|||||
| 1H | 13C | 1H | 13C | 1H | 13C | 1H | 13C | |
| 2-CH3 | 3.814 | 10.8 | 3.808 | 10.7 | 3.795 | 10.7 | 3.815 | 10.5 |
| 31-CH | 8.495 | 130.5 | 8.489 | 130.5 | 8.471 | 130.3 | 8.502 | 130.5 |
| 32-CH2z | 6.417 | 117.2 | 6.414 | 117.2 | 6.410 | 117.2 | 6.414 | 117.7 |
| 32-CH2e | 6.152 | 6.152 | 6.165 | 6.153 | ||||
| 5-Meso | 10.270 | 97.2 | 10.323 | 97.9 | 10.313 | 97.8 | 10.323 | 98.0 |
| 7-CH3 | 3.668 | 10.8 | 3.793 | 10.6 | 3.780 | 10.8 | 3.803 | 10.4 |
| 81-CH* | 4.166 | 22.2 | 8.478 | 130.5 | 8.463 | 130.3 | 8.483 | 130.5 |
| 81-CH2z† | 1.938 | 16.3 | 6.409 | 117.2 | 6.408 | 117.2 | 6.414 | 117.7 |
| 82-CH2e | 6.161 | 6.162 | 6.153 | |||||
| 10-Meso | 10.119 | 96.1 | 10.244 | 97.4 | 10.236 | 97.1 | 10.242 | 97.4 |
| 12-CH3 | 3.693 | 9.6 | 3.674 | 9.7 | 3.668 | 9.5 | 3.702 | 9.4 |
| 131-CH2 | 4.500 | 20.8 | 4.485 | 21.2 | 4.457 | 20.8 | 4.466 | 22.4 |
| 132-CH2 | 3.393 | 36.2 | 3.388 | 36.2 | 3.372 | 36.1 | 3.194 | 40.5 |
| 13-OCH3 | 3.674 | 49.9 | 3.674 | 49.9 | 3.656 | 49.9 | ||
| 15-Meso | 10.184 | 96.6 | 10.157 | 96.5 | 10.074 | 96.1 | 10.231 | 97.3 |
| 171-CH2 | 4.454 | 22.1 | 4.446 | 22.1 | 4.457 | 20.8 | 4.466 | 22.4 |
| 172-CH2 | 3.246 | 39.2 | 3.243 | 39.2 | 3.372 | 36.1 | 3.194 | 40.5 |
| 17-OCH3 | 3.656 | 49.9 | ||||||
| 18-CH3 | 3.725 | 9.7 | 3.719 | 9.7 | 3.688 | 9.5 | 3.725 | 9.4 |
| 20-Meso | 10.227 | 97.2 | 10.204 | 97.1 | 10.190 | 97.0 | 10.201 | 97.1 |
Only chemical shifts for 13C bearing protons are included, and no attempt was made to assign quaternary carbon. e and z indicate the entgegen and zusammen hydrogens.
For mvMPE, 81-CH2.
† For mvMPE, 82-CH3.
Figure 6.

Proposed radical reaction mechanism for an AdCob-dependent anaerobic MPE-cyclase.
Effects of B12 on bluE and bluB Mutant Cells.
The accumulation of MPE and mvMPE at the expense of BChl, by the B12-depleted mutant bluE or bluB cells could be prevented by B12 (Fig. 4), suggesting that a cobalamin cofactor is required to convert MPE to PChlide in R. capsulatus.
Figure 4.

Percent growth, BChl formation, and MPE content as a function of B12 concentration in cultures of B12-deficient mutants bluE and bluB. Growth (shown as percent of maximal growth observed, OD700 = 2.4) was for 26 h in the dark from inocula sizes predetermined to cause MPE accumulation.
Nicotinamide causes PChlide accumulation in R. sphaeroides (29) resulting from an inhibition of PChlide reductase. In agreement therewith, vitamin B12-deficient bluE cells incubated with vitamin B12, 10 mM nicotinamide, and chloramphenicol accumulated PChlide. Fig. 5A shows the characteristic absorption maxima at 436 and 630 nm, of PChlide in the B12-treated cells relative to control. Chromatography of the pigments from the B12-treated cells showed a green band, running slightly ahead of MPE, with the same Rf as PChlide. The eluted green band had a Soret absorption at 434 nm. Further experiments showed that after about 4–6 h, chloramphenicol begins increasingly to inhibit PChlide formation.
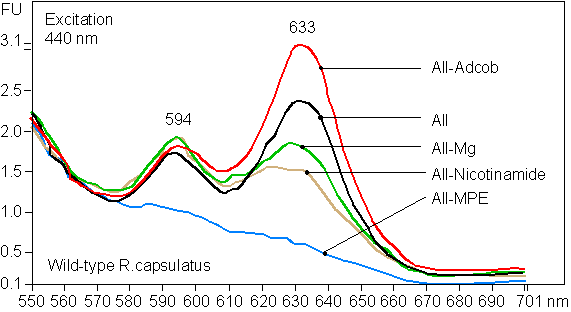
Figure 5.

(A) PChlide formation induced by B12 in B12-deficient bluE cells. Absorption spectra of extracts of bluE cells (inoculum size 10 μl), grown to OD700 = 1.7, in 50 ml. Nicotinamide (10 mM) and 34 μg⋅ml−1 chloramphenicol (CAP) were added, followed by incubation for 6 h, with or without 5 μM B12. (B) Fluorescence emission spectra of extracts of cells from in vivo cyclase assays of wild-type R. capsulatus. FU, fluorescence units. The assays were performed with and without 10 mM nicotinamide with 8 μM MPE in 2.8 μl of DMSO. (C) Fluorescence emission spectra of cells from in vivo cyclase assays of bluE mutant. Chloramphenicol, 34 μg⋅ml−1; adenosylcobalamin (AdCob), 15 μM; B12, 15 μM; nicotinamide, 10 mM.
In Vivo Cyclase Activity in the bluE Mutant Requires a Form of Cobalamin.
It has not yet been possible to measure cyclase activity in cell-free preparations of R. capsulatus, so an in vivo assay for the cyclase was developed for wild-type cells (Fig. 5B). Cells supplied with MPE and nicotinamide exhibited high MPE-cyclase activity. Sonication destroyed the activity. The pH optimum of this activity is remarkably narrow at pH 7.3. To demonstrate that MPE-cyclase is directly dependent on a cobalamin cofactor, we repeated this experiment with the B12-requiring bluE mutant under conditions where protein synthesis was inhibited (Fig. 5C). The MPE that accumulated in the bluE mutant could be used as internal substrate in cyclase assays. Fig. 5C shows that mutant MPE cyclase activity has an absolute demand for exogenous B12 or AdCob. This experiment shows that a cobalamin cofactor acts directly on the cyclase reaction. Since methylation is not involved, the cofactor is presumably AdCob and not methylcobalamin. This result is supported by experiments with wild-type cells permeabilized by two freeze-thaw cycles (see Fig. 12 in supplementary data at www.pnas.org). Substantial MPE-cyclase activity is observed with these cells supplied with 10 mM nicotinamide, 30 mM MgCl2, and 8 μM MPE (purified from bluB mutant) in 2.8 μl of DMSO. The system has an absolute requirement for MPE and partial requirements for Mg2+ and nicotinamide. Addition of 15 μM AdCob partially inhibited formation of PChlide.
Role of Iron in BChl Biosynthesis. An Iron–Sulfur Center in the BchE Gene Product?
Iron-deficient R. capsulatus accumulates coproporphyrin and MPE (30), suggesting the presence of iron in both the ANCOPROX and the cyclase. There are four conserved cysteines in the MPE-cyclases and the methylases, shown in Fig. 2. Three cysteines are grouped between position 230 and 237. The fourth is at position 297. ANCOPROX has only the three grouped cysteines. This cysteine arrangement resembles somewhat an iron–sulfur center as found in biotin synthase (31). As proposed below, an iron–sulfur center could mediate one-electron transfer from the substrate. The iron–sulfur center of the biotin synthase and related enzymes is thought to be required for generation of the adenosyl radical from S-adenosylmethionine (32).
Proposal of an AdCob-Dependent Anaerobic Mechanism for Formation of the Isocyclic Ring of Chlorophyll.
The proposed reaction mechanism for an AdCob-requiring cyclase reaction is shown in Fig. 6. In the reaction six hydrogen atoms are formally removed from the substrates MPE and water. The electrons are removed from the enzyme by an iron–sulfur center. We propose that the enzyme catalyzes formation of the adenosyl radical from AdCob. This radical then transfers its free electron to the substrate, possibly via a protein radical (step 1, Fig. 6). Initially the radical abstracts a hydrogen atom, forming the 131-radical of the methyl propionate side chain of MPE. This radical, analogous to a benzylic radical, is stabilized by electron delocalization over the ring. If, during the further reactions, radical termination occurs, regenerated adenosyl radical initiates the process again. Electron withdrawal (step 2) leads to the 131-cation of MPE. This cation is attacked, in step 3, by OH− derived from water, forming the 131-hydroxy derivative of MPE. Withdrawal of a hydrogen atom (step 4) gives the 131-radical of 131-hydroxy-MPE. This will make the 131-hydroxyl group much more acid (33) so that, in step 5, it will dissociate and attack a metal ion such as Fe(III) in the iron–sulfur center. This would facilitate intermolecular electron transfer to give 131-keto-MPE. Removal of a hydrogen atom (step 6) then occurs to form the 132-radical of 131-keto-MPE. Finally, this radical attacks the adjacent methene carbon and a hydrogen atom is again removed to give the isocyclic ring of PChlide. A radical reaction by itself is not expected to provide the stereospecificity required for the isocyclic ring closure. Such stereospecificity may involve participation of auxiliary protein as exemplified by (+)-pinoresinol formation in lignan biosynthesis (34). Alternatively, a dehydrogenase may oxidize the hydroxy intermediate to the keto form and the ring closure may not require a radical intermediate, if an enol intermediate is generated after the keto group is introduced.
The in vitro system with permeabilized cells synthesizing PChlide from externally added Mg-protoporphyrin IX monomethyl ester and the cofactor vitamin B12 provides an opportunity to investigate the enzymology and the mechanism of the isocyclic ring formation catalyzed by the BchE protein.
Supplementary Material
Acknowledgments
We thank Drs. Gamini Kannangara, Ulf Berg, Morten Meldal, and Cecilia Hägerhäll for helpful discussions; Drs. Carl Bauer and Cornelia Wersig for providing the mutants; Ms. Pia Breddam for performing the LDI-TOF MS; Mss. Nina Rasmussen and Ann-Sofi Steinholz for artwork; and Dr. Mats Hansson for essential support and encouragement.
Abbreviations
- B12
vitamin B12
- AdCob
adenosylcobalamin
- MP
Mg-protoporphyrin
- MDE
Mg-protoporphyrin dimethyl ester
- MPE
Mg-protoporphyrin monomethyl ester
- mvMPE
3-vinyl-8-ethyl-MPE
- PChlide
protochlorophyllide
- BChl
bacteriochlorophyll
- LDI-TOF MS
laser desorption/ionization time-of-flight mass spectrometry
- ANCOPROX
anaerobic coproporphyrinogen oxidase
References
- 1.Porra R J, Schafer W, Gad'on N, Katheder I, Drews G, Scheer H. Eur J Biochem. 1996;239:85–92. doi: 10.1111/j.1432-1033.1996.0085u.x. [DOI] [PubMed] [Google Scholar]
- 2.Walker C J, Mansfield K E, Smith K M, Castelfranco P A. Biochem J. 1989;257:599–602. doi: 10.1042/bj2570599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Wong Y-S, Castelfranco P A. Plant Physiol. 1984;75:658–661. doi: 10.1104/pp.75.3.658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bollivar D W, Beale S I. Plant Physiol. 1996;112:105–114. doi: 10.1104/pp.112.1.105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Yang Z M, Bauer C E. J Bacteriol. 1990;172:5001–5010. doi: 10.1128/jb.172.9.5001-5010.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Pollich M, Jock S, Klug G. Mol Microbiol. 1993;10:749–757. doi: 10.1111/j.1365-2958.1993.tb00945.x. [DOI] [PubMed] [Google Scholar]
- 7.Pollich M, Klug G. J Bacteriol. 1995;177:4481–4487. doi: 10.1128/jb.177.15.4481-4487.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bollivar D W, Suzuki J Y, Beatty J T, Dobrowolski J M, Bauer C E. J Mol Biol. 1994;237:622–640. doi: 10.1006/jmbi.1994.1260. [DOI] [PubMed] [Google Scholar]
- 9.Drews G. Mikrobiologisches Prakticum. Berlin: Springer; 1983. [Google Scholar]
- 10.Young D A, Bauer C E, Williams J, Marrs B L. Mol Gen Genet. 1989;218:1–12. doi: 10.1007/BF00330558. [DOI] [PubMed] [Google Scholar]
- 11.Smith J H C, Benitez A. In: Modern Methods of Plant Analysis. Paech K, Tracey M V, editors. Vol. 4. Berlin: Springer; 1955. p. 142. [Google Scholar]
- 12.Granick S. J Biol Chem. 1948;175:333–342. [PubMed] [Google Scholar]
- 13.Granick S. J Biol Chem. 1961;236:1168–1172. [PubMed] [Google Scholar]
- 14.Rance M, Sørensen O W, Bodenhausen G, Wagner G, Ernst R R, Wüthrich K. Biochem Biophys Res Commun. 1983;117:479–485. doi: 10.1016/0006-291x(83)91225-1. [DOI] [PubMed] [Google Scholar]
- 15.Jeener J, Meier B H, Bachmann P, Ernst R R. J Chem Phys. 1979;71:4546–4553. [Google Scholar]
- 16.Kjær M, Andersen K V, Poulsen F M. Methods Enzymol. 1994;239:288–307. doi: 10.1016/s0076-6879(94)39010-x. [DOI] [PubMed] [Google Scholar]
- 17.Altschul S F, Madden T L, Schäffer A A, Zhang J, Zhang Z, Miller W, Lipman D J. Nucleic Acids Res. 1997;25:3389–3402. doi: 10.1093/nar/25.17.3389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kuzuyama T, Seki T, Dairi T, Hidaka T, Seto H. J Antibiot (Tokyo) 1995;48:1191–1193. doi: 10.7164/antibiotics.48.1191. [DOI] [PubMed] [Google Scholar]
- 19.Fuhrman S, Overmann J, Pfennig N, Fischer U. Arch Microbiol. 1993;160:193–198. [Google Scholar]
- 20.Ormerod J G, Nesbakken T, Beale S I. J Bacteriol. 1990;172:1352–1360. doi: 10.1128/jb.172.3.1352-1360.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Alston T A. Life Sci. 1991;48:1591–1595. doi: 10.1016/0024-3205(91)90284-i. [DOI] [PubMed] [Google Scholar]
- 22.Drummond J T, Matthews R G. Biochemistry. 1994;33:3742–3750. doi: 10.1021/bi00178a034. [DOI] [PubMed] [Google Scholar]
- 23.Riedel B, Fiskerstrand T, Refsum H, Ueland P M. Biochem J. 1999;341:133–138. [PMC free article] [PubMed] [Google Scholar]
- 24.Muir M, Chanarin I. J Haematol. 1984;58:517–523. doi: 10.1111/j.1365-2141.1984.tb03999.x. [DOI] [PubMed] [Google Scholar]
- 25.Jones R M, Lamb J H, Lim C K. Rapid Commun Mass Spectrom. 1995;9:921–923. doi: 10.1002/rcm.1290091011. [DOI] [PubMed] [Google Scholar]
- 26.Low D L, Gray H B, Duus J Ø. J Am Chem Soc. 1997;119:1–5. [Google Scholar]
- 27.Abraham R J, Eivazi F, Nayyir-mazhir R, Pearson H, Smith K M. Org Magn Reson. 1978;11:52–54. [Google Scholar]
- 28.Shiau F-Y, Whyte B J, Castelfranco P A, Smith K M. J Chem Soc Perkin Trans. 1991;1:1781–1785. [Google Scholar]
- 29.Shioi Y, Doi M, Boddi B. Arch Biochem Biophys. 1988;267:69–74. doi: 10.1016/0003-9861(88)90009-4. [DOI] [PubMed] [Google Scholar]
- 30.Cooper R. Biochem J. 1963;89:100–108. doi: 10.1042/bj0890100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Johnson M K. In: Encyclopedia of Inorganic Chemistry. King R B, editor. New York: Wiley; 1994. pp. 1896–1913. [Google Scholar]
- 32.Duin E C, Lafferty M E, Crouse B R, Allen R M, Sanyal I, Flint D H, Johnson M K. Biochemistry. 1997;36:11811–11820. doi: 10.1021/bi9706430. [DOI] [PubMed] [Google Scholar]
- 33.Ochiai E-I. In: Metal Ions in Biological Systems. Sigel H, Sigel A, editors. Vol. 30. New York: Dekker; 1994. pp. 1–25. [Google Scholar]
- 34.Davin L B, Wang H-B, Crowell A L, Bedgar D L, Martin D M, Sarkanen S, Lewis N G. Science. 1997;35:362–366. doi: 10.1126/science.275.5298.362. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}