

Abstract
In mouse mutants incapable of expressing μ chains, VκJκ joints are detected in the CD43+ B cell progenitors. In agreement with these earlier results, we show by a molecular single cell analysis that 4–7% of CD43+ B cell progenitors in wild-type mice rearrange immunoglobulin (Ig)κ genes before the assembly of a productive VHDHJH joint. Thus, μ chain expression is not a prerequisite to Igκ light chain gene rearrangements in normal development. Overall, ∼15% of the total CD43+ B cell progenitor population carry Igκ gene rearrangements in wild-type mice. Together with the results obtained in the mouse mutants, these data fit a model in which CD43+ progenitors rearrange IgH and Igκ loci independently, with a seven times higher frequency in the former. In addition, we show that in B cell progenitors VκJκ joining rapidly initiates κ chain expression, irrespective of the presence of a μ chain.
Keywords: B cell development, bone marrow, immunoglobulin gene rearrangement
During B cell development, genes encoding immunoglobulin V regions are generated by recombination of individual gene segments. Genes encoding Ig heavy chains (IgH genes) are formed by first rearranging a DH to a JH segment, followed by a VH to DHJH rearrangement. In the light chain (L) loci, a VL to JL recombination event generates an Ig light chain (IgL) gene. If the resulting joints are in a contiguous open reading frame, the rearrangements are referred to as “productive”.
In regard to the relative order of VHDHJH and VL JL recombination events, two models have been proposed. According to the “ordered” model, expression of a μ heavy chain from a productively rearranged IgH gene induces light chain gene rearrangement. Evidence that formation of VHDHJH complexes usually precedes light chain gene rearrangement comes from the analysis of Abelson murine leukemia virus–transformed pre-B cells in culture (1, 2) and from studies of Ig gene rearrangements in B cell precursor populations isolated ex vivo (3). Furthermore, the expression of a transfected membrane-bound μ chain as well as cross-linking of pre-B cell receptor complexes (consisting of membrane-bound μ chains and the products encoded by the λ5 and VpreB genes; reference 4) stimulated the rearrangement of endogenous κ light chain genes in transformed pre-B cell lines (5–7). In addition, an increased number of VκJκ rearrangements was observed in fetal livers of heavy chain transgenic mice as compared with nontransgenic mice (8).
In contrast, the “stochastic” model of IgH and IgL gene recombination states that μ chain expression and pre-B receptor signaling are not required for IgL gene rearrangement and suggests that IgH and IgL loci rearrange independently of each other (9, 10). This hypothesis is supported by the analysis of Abelson murine leukemia virus–transformed murine pre-B cell lines derived from normal (11) and scid mice (12). In both cases, some cells were shown to rearrange Igκ loci in the absence of a membrane-bound μ heavy chain. In vitro differentiation experiments using normal murine pre-B cell lines have also demonstrated that κ protein could be expressed in the absence of a μ chain (13). Moreover, κ chain expression was detected in the absence of productive VHDHJH rearrangements in immortalized B cell precursors of human fetal bone marrow (14). Examination of transformed embryonic bursal cells showed that during chicken B cell development, IgL genes can also be rearranged before IgH gene rearrangement has been completed (15).
Although in vivo most Igκ rearrangements occur in the pre-B cell compartment into which progenitor cells are driven upon pre-B cell receptor (i.e., μ chain) expression (5, 16), evidence indicates that initially, when gene rearrangements in IgH are set in motion in CD43+ progenitors, VκJκ rearrangements also occur, albeit at low frequency (17, 18). At this early stage of development, κ rearrangements appear to be independent of μ chain expression and, indeed, any rearrangement in the IgH locus, as they are also seen in mutant mice unable to either express membrane-bound μ chains (μMT mice; 19) or generate VHDHJH joints due to a targeted deletion of the JH elements (20, 18). These data suggest that gene rearrangements in the Igκ locus occur at two stages of development: in early CD43+ progenitors at low frequency and independent of μ chain expression, and later on, at high frequency, in pre-B cells upon pre-B cell receptor expression. However, one might argue that the analysis of the mutant mice could be misleading because in these animals the progenitors do not develop beyond the CD43+ stage and therefore persist in this cellular compartment for a prolonged time, accumulating gene rearrangements that normally would not have occurred. On the other hand, Igκ gene rearrangements seen in CD43+ B cell progenitors of wild-type mice (17) could be derived from cells already expressing μ chains.
We therefore decided to verify the results obtained in the mouse mutants by the analysis of IgH and Igκ rearrangements in individual CD43+ B cell progenitors in wild-type animals. This approach allows us to investigate whether recombination of IgL loci can indeed precede the generation of productive IgH gene rearrangements in the course of B cell development under physiological conditions, and, if so, to evaluate the frequency of these events.
Materials and Methods
Cell Sorting
Single cell suspensions were prepared from bone marrow by flushing femurs with DMEM (containing 5% FCS, 0.1% NaN3) or from splenic tissues of BALB/c mice (8–12 wk old; Bomholtgaard, Denmark). Cells were treated with Tris-buffered 0.165 M NH4Cl to eliminate erythrocytes and washed by centrifugation through FCS.
3-83κi mice (21) were used at 8–12 wk of age. Wild-type mice used in the staining shown in Fig. 1 were F1 at the age of 8–12 wk from a 129sv × BALB/c cross.
Figure 1.

Staining of bone marrow B cell precursors for intracellular Igκ expression. Fraction B cells from wild-type (A) and 3-83κi/+ mice (B). Fraction D cells from wild-type (C) and 3-83κi/+ mice (D). Anti-Igκ chain antibody is shown on the y-axis, and the forward scatter of the cells is shown on the x-axis. Fraction D cells were used to gate κ+ cells. Numbers indicate the percentage of cells in the window.
Cell sorting was performed using a dual laser flow cytometer (FACStar®). Single cells were directly deposited into 0.5-ml microtubes containing 20 μl 1× PCR buffer (GIBCO BRL, 2.5 mM MgCl2) supplemented with 1 μg/ml rRNA from Escherichia coli (Boehringer Mannheim), immediately frozen on dry ice, and stored at −80°C. Single cells from the E14 embryonic stem cell line (22) were isolated accordingly as negative controls for the PCR. Depending on the set of the cytometer, up to 20% of the tubes could be empty during a particular sorting procedure.
Isolation of Fraction B, C, and D Cells.
Fractions were classified according to Hardy et al. (23). Pooled bone marrow cells from two to six mice were depleted of MAC-1+ cells (and of IgM+ cells in the experiment with subsequent κ protein staining) by magnetic cell separation (24) using antibody M1/70.15.11/2 (anti–Mac-1; reference 25) or antibody CD11B (anti–Mac-1; Miltenyi Biotec), and in addition rat anti–mouse IgM (Miltenyi Biotec) antibodies for the experiment with subsequent κ protein staining, coupled to magnetic beads (Miltenyi Biotec). Cells passing through the column in the magnetic field were collected and further stained by a combination of FITC-S7 (anti-CD43; reference 26), PE-BP-1 (anti–BP-1; reference 23), biotin-30F1 (anti– heat-stable antigen; reference 23), and allophycocyanin-RA3-6B2 (anti-CD45R/B220; reference 23) in staining medium, washed, and counterstained by Texas red–avidin (Boehringer Mannheim).
To obtain fraction B cells that expressed κ chains intracellularly, sorted fraction B cells (∼105 cells) from pooled bone marrow of five mice were fixed in PBS containing 2% formaldehyde for 20 min at room temperature. After washing with PBS, the cells were resuspended in PBS containing 0.1% NaN3 and 1% BSA, bleached overnight, and then stained with FITC–R33-18 (anti-κ; reference 27) in PBS containing 1% saponin (Sigma Chemical Co.).
The extent of possible contamination of CD43+ by CD43− B cell precursors (pre-B cells) or by B cells (all bearing productive VHDHJH joints) can be estimated as not exceeding 10% from the staining data (not shown) for the sortings of fraction B cells. By selecting κ+ cells, one would expect to enrich for contaminating cells, so that the proportion of cells bearing productive VHDHJH rearrangements would be higher in the κ chain–expressing than in total fraction B cells. Since this is not the case (8 out of 15 κ+ cells compared with 7 out of 11 unselected fraction B cells; see Tables V and VI), a significant contamination of fraction B cells by pre-B or B cells seems excluded. In the sortings of fraction C cells, the staining data do not allow us to rule out the possibility of a contamination by pre-B or B cells that could be >10%. Note, however, that contaminating cells, if present, would appear only among the cells with productive VHDHJH joints, and would thus lead to an underestimation of the true proportion of cells that form VκJκ joints while lacking productive VHDHJH rearrangements in early B cell development.
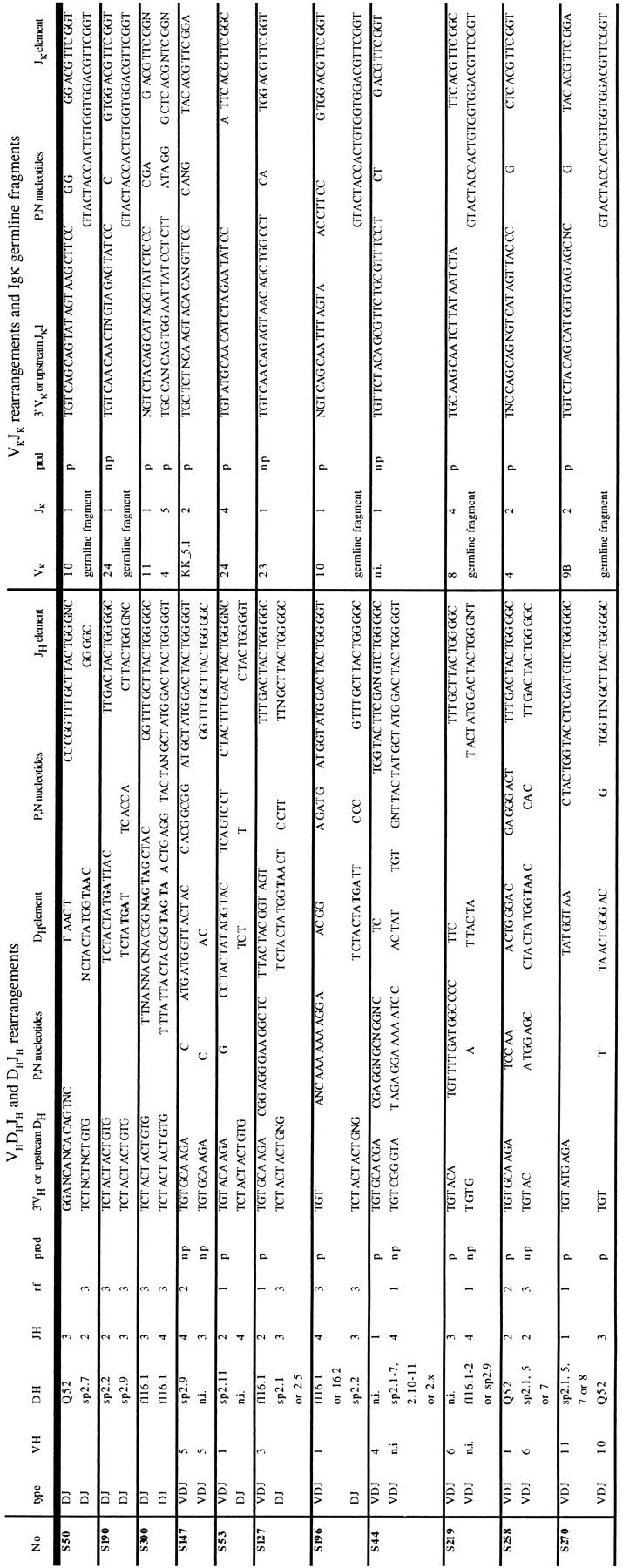
Table V.
Sequences of Dh Jh, VhDh Jh, and Vκ Jκ Junctional Regions Ig Gene Rearrangements in B Cell Progenitors from Fraction B
|
Designations are the same as in Table III.
Table VI.
Classification of B Cell Progenitors Carrying Vκ Jκ Rearrangements by the Configuration of Their IgH Loci
| Fraction | DJ/DJ | VDJ−/DJ | VDJ−/VDJ− | VDJ+/DJ | VDJ+/VDJ− | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| C | 1 | 4 | 2 | 6 | 1 | |||||
| (352) | (298, 321, 717, 718) | (78, 96) | (5, 80, 265, 294, 499, 530) | (538) | ||||||
| κ chain+ B | 5 | 2 | 0 | 6 | 2 | |||||
| (43, 52, 60, 64, 66) | (62, 110) | (40, 50, 57, 69, 70, 113) | (46, 87) | |||||||
| B | 3 | 0 | 1 | 3 | 3 | |||||
| (s50, s190, s300) | (s147) | (s53, s127, s196) | (s44, s219, s258) |
B cell progenitors of fractions B and C that carry VκJκ joints (Tables III IV V) are classified into five groups according to the rearrangements of the two IgH alleles. The number of cells in each group is indicated. Numbers in parentheses denote the designations of the cells as given in Tables III IV V. VDJ− and VDJ+ represent nonproductive and productive VHDHJH rearrangements, respectively. Cells with productive VκJκ rearrangements are shown in bold, and cells with a DHJH joint in reading frame 2 that can encode a Dμ protein are underlined.
Sorting of κ light chain expressing splenic B cells was done by staining splenocytes with PE-RA3-6B2 (23) and FITC-R33-18 (27).
PCR and Sequence Analysis of Ig Gene Rearrangements
To prepare DNA for amplification, 1 μl of an aqueous solution of proteinase K (10 mg/ml; Boehringer Mannheim) was added, samples were overlaid with paraffin oil, and were incubated for 30 min at 55°C. Subsequently, proteinase K was inactivated for 10 min at 95°C. PCR amplification was carried out in two rounds: the first reaction contained all 5′ primers, JH4E (29), and Jκ5E primers (2.5 pmol each; Table I). Amplification was done over 30 cycles (1 min at 95°C, 1 min at 60°C, and 2.5 min at 72°C). For the second PCR, 1.5-μl aliquots of the first round amplification product were transferred into separate reactions (set up in 96-well microtiter plates; Costar Corp.), each containing a single 5′ primer in combination with either the nested JH4A (amplification of IgH genes; reference 29) or the Jκ5A primer (amplification of Igκ genes) (7 pmol of each primer). 30 cycles were performed (1 min at 95°C, 1 min at 63°C, and 1.5 min at 72°C). All PCRs contained dATP, dCTP, dGTP, dTTP (Pharmacia Biotech) at 200 μM each, PCR buffer (GIBCO BRL), 2.5 mM MgCl2, 5 U of Taq DNA polymerase (GIBCO BRL) in the first round, and 3 U of Taq DNA polymerase in the second round. The final volume of each reaction was 50 μl. Each PCR was followed by a 5–10-min incubation at 72°C. 10 μl of the second-round PCR product was analyzed on agarose gels. Before sequencing, 1.5 μl of second-round product was reamplified for 20 cycles (30 s at 95°C, 1 min at 63°C, and 2 min at 72°C) using appropriate 5′ primers and nested 3′ primers, DNA was isolated from preparative agarose gel using Spin-X columns (Costar Corp.). Cycle sequencing was performed using the Ready Reaction Dye Deoxy Terminator Cycle Sequencing Kit (Applied Biosystems) following the manufacturer's instructions and an ABI 373A sequencer (Applied Biosystems). Sequencing primers recognize sequences downstream of the respective rearranged Jκ genes (Table I).
Table I.
Igκ Locus–specific Oligonucleotides Used In PCR and Sequencing Reactions
| Primer | Specificity | |||
|---|---|---|---|---|
| A | PCR primers | |||
| κ light chain genes | ||||
| VK1 | GCG AAG CTT CCC TGA TCG CTT CAC AGG CAG TGG | |||
| VK2 | GCG AAG CTT CCC(AT)GC TCG CTT CAG TGG CAG TGG | |||
| VK3 | GCG AAG CTT CCC A(GT)(AC) CAG GTT CAG TGG CAG TGG | |||
| KG | GCG AAG CTT AAG CTT TCG CCT ACC CAC TGC TCT | 5′ of Jκ1 | ||
| KG1 | ACA GCC AGA CAG TGG AGT ACT ACC ACT GTG | immediately 5′ of Jκ1 | ||
| JK5E | GAT CCA ATC TCT TGG ATG GTG ACC | |||
| JK5A | GGG TCT AGA CAA CTG ATA ATG AGC CCT CTC CAT | |||
| B | Sequencing primers | |||
| κ light chain genes | ||||
| JK1 | AGA CAT AGA AGC CAC AGA CAT AG | |||
| JK2 | CTT AAC AAG GTT AGA CTT AGT GAA C | |||
| JK4 | TTC ACA CAA GTT ACC CAA ACA G | |||
| JK5 | GAA CTG ACT TTA ACT CCT AAC ATG |
Sequences are presented from 5′ to 3′. Nucleotides in brackets denote a nucleotide mix at this position. V, KG, and KGI oligonucleotides are 5′ primers, whereas J oligonucleotides are 3′ primers. The Vκ primers, recognizing all Vκ genes listed by Strohal et al. (52) in framework region 3, cross-react and thus cannot be assigned to specific Vκ families. The KG and KGI primers hybridize to a germline region upstream of Jκ1. All J primers are homologous to a region downstream of the respective J element. A shows the primers used for amplification, and B shows those used in sequencing reactions.
The primers used for amplification and sequencing of Ig heavy chain genes have been described by Ehlich et al. (23) and Löffert et al. (30). The VHH primer (30) was not used in the analyses of fraction C. KGI (Table I) was used only in the analyses of fraction B cells irrespective of κ protein staining.
Sequences were analyzed using DNAPLOT at www.genetik. uni-koeln.de/dnaplot/. The database used consists of mouse V gene sequences from an EMBL/GenBank/DDBJ nucleotide sequence database, a Kabat database (31), and the Vκ sequence list compiled by Kofler et al. (32).
Control Experiment to Confirm the Isolation of Single Cells by FACS®
We chose two mutant mouse strains in which a rearranged Ig heavy chain variable region gene was introduced by gene targeting into the heavy chain locus, replacing the JH elements (T15i mice, reference 28, and B1-8i mice, reference 33, containing a rearranged VH186.2 gene isolated from the hybridoma B1-8; reference 34). From each of the two mouse strains, which were homozygous for the introduced heavy chain, 4 × 105 κ light chain– positive splenic B cells were isolated by FACS® and subsequently pooled to yield a 1:1 mixture. Of this mixture single cells as well as two cells were sorted directly into microtubes containing PCR buffer. The inserted VHDHJH complexes of these splenic B cells were amplified in a semi-nested PCR approach analogous to the one described above. The B1-8 (T15i) gene was amplified by the 5′ primer VHA (VHT15) and the 3′ primers JH2E and JH2 (JH1E and JH1) in the first and second rounds of amplification, respectively. The primers used in this experiment have been described elsewhere (30).
Results
PCR Analysis of Single B Cell Precursors.
We extended our previously described single cell PCR system for the analysis of IgH genes (29) to simultaneously examine Igκ genes. For this purpose, seven Igκ gene–specific oligonucleotides were included to detect rearranged VκJκ complexes as well as Igκ loci in germline configuration (Table I).
To estimate the efficiency of the amplification of Igκ loci rearrangements, we used splenic, surface Igκ-positive B cells. 197 VκJκ joints were amplified from 210 single B cells (none, one, or two per cell). Assuming that ∼30% of all splenic B cells carry two VκJκ complexes (9, 16), this corresponds to a VκJκ rearrangement detection efficiency of ∼70%. To determine the detection efficiency of IgH gene joints, VHDHJH and DHJH gene rearrangements were amplified from 311 B cell precursors of the CD43+ fraction C (reference 23; excluding fraction C′ cells) in the presence of Igκ locus–specific oligonucleotides. Two IgH gene PCR products were obtained from 41% of cells. In the remaining cells, either one (51%) or no (7%) IgH gene PCR products were amplified. Thus, the efficiency of the amplification was sufficient to allow the simultaneous analysis of heavy and light chain loci.
When the interdependence of rearrangements of the various Ig loci is investigated by single cell analysis, it is essential to demonstrate that the amplification products are indeed derived from the same cell, and that the samples do not occasionally contain more than one cell. Therefore, a control experiment similar to the one described by Löffert et al. (30) was performed using two mutant mouse strains in which different heavy chain transgenes were inserted into the heavy chain locus, replacing the JH elements (T15i mice, reference 28, and B1-8i mice, reference 33). Cell suspension containing equal proportions of Igκ-positive splenic B cells from both strains was prepared. From this, either “one cell” or “two cell” samples were deposited into microtubes using the FACS®. Subsequently, the IgH transgenes were amplified from these cells, using appropriate PCR primers (30).
127 “one cell” samples yielded indeed only one PCR product (Table II). In the case of the “two cell” samples, 50% of the tubes would be expected to contain two cells from the same mouse strain that would not be identified as “two cells” because both have given rise to identical PCR products. Two different PCR products were obtained in 53% of the “two cell” samples (Table II). The rare cases in which no PCR product was obtained (Table II) may be explained by a relatively poor amplification efficiency using this particular primer set, or, alternatively, these tubes may not have contained a cell. These results indicate that the direct deposition of cells by FACS® used in the experiments described below represents a reliable method for obtaining samples containing single cells.
Table II.
No. of PCR Products Obtained from Control Samples Containing Either “One” or “Two” Cells
| PCR products | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Cells per sample | No. of samples | B1-8 + T15 | B1-8 | T15 | no product | |||||
| 1 | 127 | 0 | 45 | 68 | 14 | |||||
| 2 | 90 | 48 | 19 | 19 | 4 | |||||
Either one or two cells of a 1:1 mixture of κ+ spenic B cells derived from T15i and B1-8i mice were deposited by FACS® into microtubes and their rearranged immunoglobulin genes were amplified by PCR. The numbers of different amplification products are shown for samples containing either one or two cells.
Igκ Gene Rearrangements in Early B Cell Precursors.
To investigate whether IgL gene rearrangements in B cell precursors can occur before μ chain expression, we had to look into the compartment of early B cell progenitors, where cells both with and without productively rearranged heavy chain genes are present. To classify different stages of B cell development in the bone marrow, we used the system developed by Hardy et al. (23), which divides B220+, surface Ig− cells into five cellular fractions according to their differential expression of CD43, heat stable antigen (HSA), and BP-1. For initial studies, we chose fraction C (excluding fraction C′; references 17, 23) of early B cell progenitors in which VκJκ rearrangements are six to seven times less frequent than in κ+ splenic B cells (17).
627 fraction C cells were examined. For 14 out of 50 cells bearing VκJκ rearrangements, the configurations of both IgH alleles were determined (Tables III and VI). Seven cells are potentially able to express μ chains because they harbor functional VHDHJH rearrangements. However, seven other cells contain an Igκ gene rearrangement in the absence of a functional VHDHJH complex. Two of these cells carry nonfunctional VHDHJH rearrangements at both IgH alleles, and four carry a nonproductive VHDHJH rearrangement together with a DHJH joint. Two nonproductive VHDHJH joints (in cells 298 and 717) comprise DH elements rearranged in reading frame 2 (in the nomenclature of Ichihara et al.; reference 35). Thus, these cells could have expressed a truncated heavy chain (Dμ protein; reference 36) before VHDHJH complex formation. The remaining cell harbors a rearranged Igκ allele and contains only DHJH complexes (cell 352). The DH elements in this cell are rearranged in reading frames other than reading frame 2.
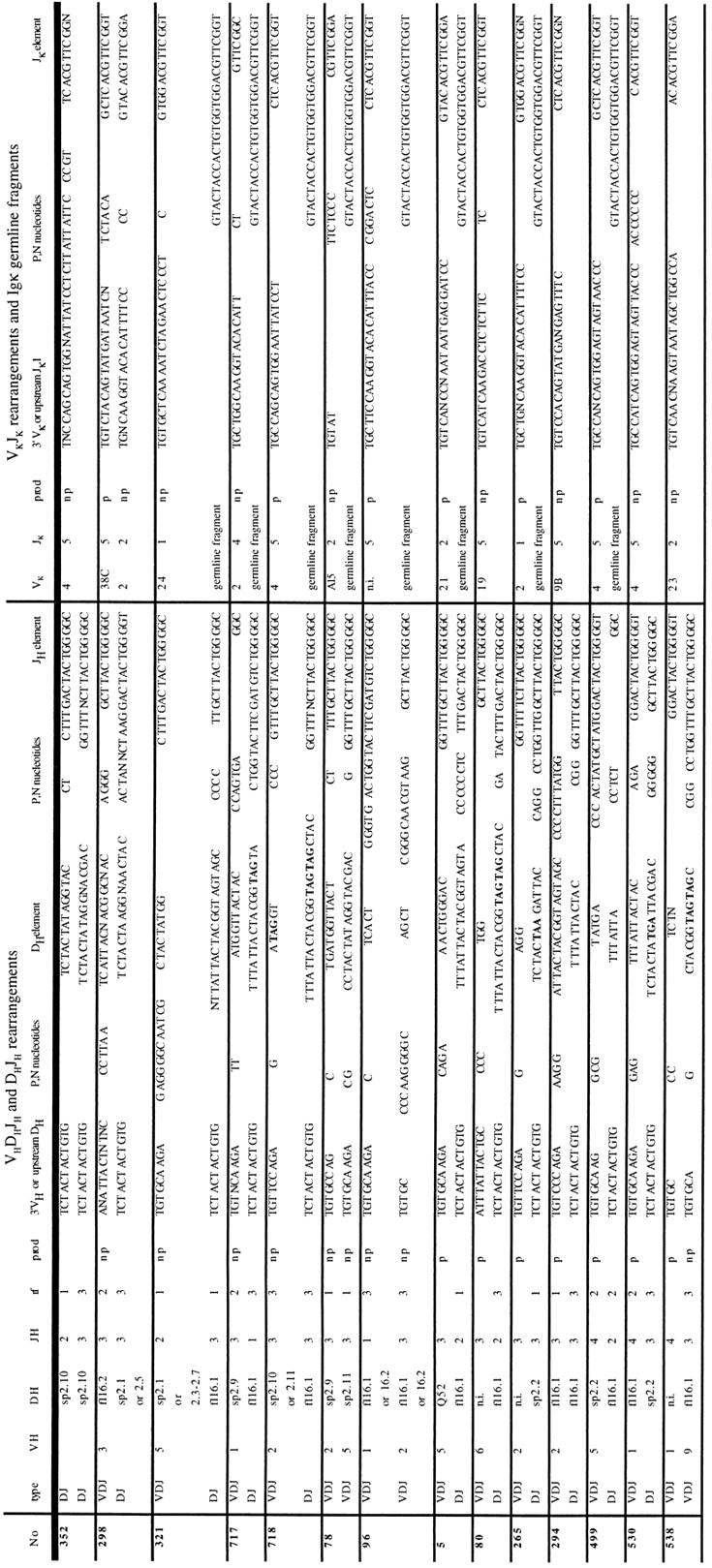
Table III.
Junctional Region Sequences of Dh Jh, VhDh Jh, and Vκ Jκ Ig Gene Rearrangements in B Cell Progenitors from Fraction C
|
The two IgH or Igκ alleles of a cell are placed so that each line contains information about one allele. Heavy chain gene sequences are shown in the left part of the table and the corresponding light chain sequences in the right part. (P,N) Nucleotides not encoded in the germline, called either N nucleotides (53, 54) or P nucleotides (55). Sequences of the IgH locus are categorized based on their configuration in the column type as DJ or VDJ. Dh sequences were assigned to published Dh segments (56) if there was homology of at least four nucleotides. Numbers in column rf indicate the Dh element reading frame (35). Reading frame is not identified for DQ52 element, because of its inability to encode for Dμ protein. N.i. indicates that the respective D or V element could not be unambiguously assigned to some gene or gene family. Numbers in JH or Jκ columns indicate the J element used in the respective joint. For VhDhJh or Vκ Jκ rearrangements the V gene families used are indicated in the columns VH or Vκ. In the column prod: p, a productive VhDh Jh or VκJκ joint; np, a nonproductive VhDh Jh or VκJκ joint. Stop codons are shown in bold.
It has been suggested that cells incapable of expressing a pre-B cell receptor accumulate in fraction C (29). Thus, at least some of the cells carrying VκJκ joints observed in fraction C could represent dead-end cells that cannot mature further and may have persisted for a prolonged time in fraction C. Such prolonged persistence may increase the probability to rearrange Igκ genes. Therefore, we decided to also analyze fraction B, the earliest stage at which VHDHJH rearrangements are detected, for the presence of cells containing rearranged κ genes in the absence of productive VHDHJH complexes. According to our previous analysis, VκJκ rearrangements are 14 times less frequent in this cell population than in κ-positive splenic B cells (17).
To enrich for cells bearing Igκ rearrangements, we isolated cells that stained for κ chains intracellularly. 88 single fraction B cells positive for intracellular κ chains were analyzed. Igκ gene rearrangements were amplified (either one or two per cell) from 47 cells. For 15 of these we were able to determine the configuration of both heavy chain alleles. 8 out of 15 cells bearing VκJκ rearrangements contained a productive VHDHJH joint. Seven cells were found to harbor either DHJH joints on both heavy chain alleles (five cells) or a nonproductive VHDHJH joint on one allele and a DHJH joint on the other (two cells) (Tables IV and VI). Reading frame 2, which encodes Dμ protein, appeared on one or both alleles in all five cells that bear only DHJH rearrangements and in one of the two cells containing a DHJH joint together with a nonproductive VHDHJH. Only one VκJκ-bearing cell (cell 62, Tables IV and VI) that is unable to produce a (truncated) heavy chain was found in this experiment. It carried a nonproductive VHDHJH rearrangement (with the DH element in reading frame 3) on one allele and a DHJH joint in reading frame 1 on the other.
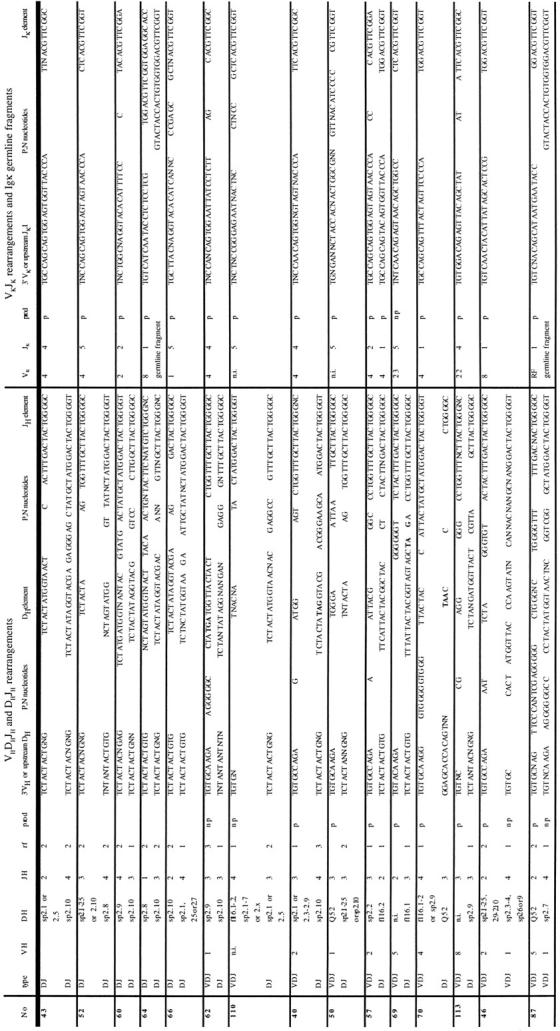
Table IV.
Sequences of Dh Jh, VhDh Jh, and Vκ Jκ Junctional Regions Ig Gene Rearrangements in Intracellular κ–expressing B Cell Progenitors from Fraction B
|
Designations are the same as in Table III.
However, in order to maximally enrich for κ chain producers we had isolated only cells that displayed high levels of κ protein. These cells might be already selected for μ chain or Dμ protein expression, considering that the stability of the κ protein could depend upon the presence of a (truncated) heavy chain in the cell. For this reason we decided to look again in cells from fraction B, this time not selected by intracellular staining for κ protein, but randomly selected by PCR for the presence of VκJκ rearrangements.
373 single cells sorted from fraction B were analyzed. In 32 cells we detected one or two rearrangements at the κ locus. In 11 of these cells we were also able to amplify and sequence rearrangements of both heavy chain alleles. In four cells no productive VHDHJH joint was present (Tables V and VI). One cell contained two nonproductive VHDHJH joints (one of which comprises a DH element in reading frame 2), and three cells carried DHJH rearrangements on both heavy chain alleles. None of these DHJH/DHJH cells harbored DH elements rearranged in reading frame 2.
Igκ Chain Expression in Early B Cell Precursors.
Given the efficiency of VκJκ joint detection of ∼70% and the fact that single cell sorting procedure will leave up to 20% of the tubes empty, the overall frequency of cells bearing VκJκ rearrangements (either productive or nonproductive) in both fractions B and C is in the range of ∼11–16%.
To estimate the frequency of cells that are able to express κ chain at the early stages of B cell development, we stained fraction B cells for intracellular κ protein. We used wild-type mice and the 3-83κi mouse mutant in which a productive VκJκ gene segment encoding the VL region of antibody 3-83 (37) was inserted by gene targeting into its natural genomic localization so that its expression is controlled by the endogenous regulatory elements (21). Due to the fact that in wild-type mice two-thirds of the VκJκ rearrangements are out of frame, 3-83κi mice should show a threefold increase in the number of κ chain–expressing cells in fraction B. The result of this experiment is shown in Fig. 1: ∼7% of cells in fraction B in wild-type mice were found to express κ chains, whereas this value was 24% in the 3-83κi mutant, yielding almost exactly the expected 1:3 ratio.
These data are in agreement with the frequency of Vκ Jκ rearrangements in cells from fraction B estimated by PCR analyses. Together, these results suggest that Igκ gene rearrangement and expression follow each other rapidly.
Discussion
Reliability of the Assay System.
A control experiment in which either one or two cells were deflected into each reaction tube (Table II) confirmed that the method to isolate single cells by using the FACS® is highly reliable and that the PCR products obtained from one sample are indeed derived from a single cell. This is further supported by the fact that PCR amplification of one sample never generated more than four products (two from heavy chain loci and two from κ light chain loci; data not shown). There was also no indication for the presence of contaminating DNA molecules in the PCR, because rearranged Ig genes were never amplified from control samples containing embryonic stem cells and the sequences of all rearrangements were different. Therefore, it is unlikely that in the cases where rearranged κ genes were observed in the absence of productive VHDHJH complexes, the IgH gene rearrangements amplified were derived from a second cell present in the sample or from foreign DNA. The extent of a possible contamination in fractions B and C by CD43− pre-B or B cells due to inaccurate cell separation during FACS® sorting is discussed in the Materials and Methods section. However, the presence of such contaminating cells (all bearing productive VHDHJH joints) would result in an underestimation of the percentage of cells bearing VκJκ joints but no productive VHDHJH rearrangements in the early fractions of B cell progenitors.
Although most Ig genes present in the germline are recognized by the collection of the primers used, certain combinations of gene rearrangements in a cell could not be detected. In particular, all DH elements (except the DHQ52 element) are recognized by the same primer and the primers specific for Vκ genes are highly homologous in structure (Table I). Therefore, most of the DHJH joints using the same JH genes on both chromosomes or distinct VκJκ rearrangements with the same Jκ segment could not be resolved. For these reasons the number of cells with DHJH joints at both IgH loci and the number of cells bearing two VκJκ joints could be underestimated.
Igκ Gene Rearrangements Appear To Be Independent of Heavy Chain Expression in CD43+ B Cell Progenitors.
The question of whether expression of a productive VHDHJH rearrangement is a prerequisite for light chain gene rearrangement during B cell development or whether Igκ gene rearrangement can take place also in the absence of a membrane-bound μ chain has been discussed controversially. The analysis of Ig gene rearrangements of single B cell progenitors isolated ex vivo from wild-type mice addresses this issue directly.
Cells of the earliest B cell progenitor fractions in which VκJκ rearrangements are detectable, namely, cells of the CD43+ fractions B and C (17), were chosen for analysis. The results obtained are summarized in Table VI. Overall, 18 cells were found to carry Igκ rearrangements in the absence of a productive VHDHJH joint. However, six of these contained DHJH rearrangements in reading frame 2, and thus were able to express Dμ proteins. Like a μ chain, the Dμ protein could associate with the products of the λ5 and VpreB genes to form a pre-B cell receptor–like complex (7). It has been suggested that Dμ protein expression, similar to μ chain expression, provided a stimulatory signal for Igκ gene rearrangements (5, 38, 39). Among the other cells analyzed, three (cells 298, 717, and s147; Tables III, V, and VI) had nonproductive VHDHJH joints in which DH elements were rearranged in reading frame 2, and thus could have expressed a Dμ protein earlier. For these cells, as well as for the ones containing Igκ rearrangements and productive VHDHJH joints, we can neither deduce the order of rearrangements at heavy and light chain loci nor state their interdependence. However, the remaining nine cells have either nonproductive VHDHJH rearrangements (with the DH elements in reading frames 1 or 3) and/or DHJH joints in reading frames 1 or 3 (Table VI) and are thus unable to express μ or Dμ chains.
We cannot rule out the possibility that some of the heavy chain gene joints detected were formed by secondary rearrangement events; specifically, previously productive VHDHJH rearrangements could have been rendered nonproductive by VH gene replacement (40–42), and DHJH joints could also have been substituted by recombining upstream D and downstream J elements with possible changes to the reading frame (5, 40, 42, 43). However, it has been implied that expression of the recombination activating genes RAG1 and RAG2 is downregulated upon pre-B cell receptor expression, suggesting that recombination of heavy chain genes is terminated once a μ chain is expressed (44). Furthermore, Dμ protein expression has also been suggested to prevent further IgH gene rearrangements (20, 30). In line with this idea, recent data have shown that Dμ protein transgene expression leads to a partial block in VH to DHJH rearrangements (39). For these reasons, it is unlikely that a major fraction of cells carrying VκJκ joints but no DHJH rearrangement using reading frame 2 or no productive VHDHJH rearrangement had assembled their IgL genes while expressing Dμ or μ chains, respectively, and altered their IgH gene complexes during subsequent rearrangements.
The data presented here are consistent with the earlier detection of VκJκ joints in B cell progenitors of mouse mutants unable to express μ chains (17–19) and support the view that Ig gene rearrangements in CD43+ B cell progenitors of the mouse follow the “stochastic” model.
If rearrangements of IgH and IgL loci indeed occur independently in CD43+ B cell progenitors, productive and nonproductive VHDHJH joints should distribute randomly in cells bearing VκJκ rearrangements. Although this is true insofar as the ratio of productive to nonproductive joints is similar in κ chain+ VκJκ rearrangement–containing and in total CD43+ cells (∼50%; Table VI and reference 29), it is also obvious that, overall, the CD43+ progenitor population is selected for productive VHDHJH joints, as their frequency would be only 24% in a random distribution (considering that one-third of the joints are in-frame and that ∼80% of the D elements in reading frame 3 contain stop codons). An over-representation of productive versus nonproductive VHDHJH joints in these early progenitors has been repeatedly observed in other experiments: 0.6 (reference 45), 0.6 (reference 30), and 0.8 (reference 46). How can this selection be explained? Two possibilities can be considered: either the bias is introduced by the expansion of pre-B cell receptor–expressing (and therefore μ+) CD43+ progenitors that have downregulated RAG-1 and -2 expression (44, 47), or the CD43-expressing progenitors that we have analyzed contain a subset of classical pre-B cells in which RAG-1 and -2 are re-induced to mediate gene rearrangement in IgL loci, but surface CD43 expression is (still) retained. The existence of such cells could explain the finding of Pelanda et al. (21) that in surface (s)Ig−, CD43+, HSA+ B cell precursors, the frequency of cells expressing κ light chains intracellularly is reduced approximately fourfold in the absence of the λ5 gene product. However, it is also possible that in the absence of λ5, κ and μ chain–expressing progenitors transit more rapidly into the compartment of sIgM+ B cells than in the wild-type. That CD43+ B cell precursors are in principle able to express sIg has been shown in mice containing productively rearranged heavy and light chain genes targeted into the corresponding Ig loci (Lam, K.-P., personal communication).
Given those complexities, we cannot exclude that some of the VκJκ rearrangements that we have found in the CD43+ B cell progenitors were induced upon pre-B cell receptor expression, although we consider this unlikely. However, the finding that about half of the CD43+ cells bearing VκJκ joints have yet to undergo IgH gene rearrangements for μ chain expression supports the concept originally developed from the analyses of mutant mice unable to express IgH chains (17), namely that in CD43+ B cell progenitors, rearrangements of heavy and light chain loci are initiated “stochastically”, with an approximately seven times higher frequency of rearrangements at the IgH than at the Igκ loci (see below).
The order of the rearrangements at the IgH and IgL loci determines the subsequent developmental route of the cell. If a μ chain is assembled first, a pre-B cell receptor will be expressed. The pre-B cell receptor gives a proliferative signal and directs the development of the cell to the CD43− pre-B cell compartment, where most IgL chain genes are rearranged (16, 48). However, if an IgL chain is expressed before or simultaneously with a μ chain, the cell is no longer dependent on the pre-B cell receptor to enter the B cell pool: as shown by Pelanda et al. (21), at least some conventional κ chains can substitute for the surrogate light chain and promote the development of progenitor B cells. Since we do not see any obvious bias towards some particular Vκ gene families among the κ chain sequences derived from CD43+ B cell progenitors, it seems that a large repertoire of VκJκ joints can be generated in this compartment.
The existence of a pre-B cell receptor–independent developmental pathway that may be evolutionary more ancient than the pre-B cell receptor–driven pathway (17) may allow the generation of B cells whose μ chains are incapable of pairing with the surrogate light chain and thus are bound to die unless rescued by a conventional IgL chain. For example, a fraction of VH81X-bearing heavy chains does not associate with the surrogate light chain (49), and thus these VH81X-expressing B cells must be generated via the pre-B cell receptor–independent pathway. ten Boekel et al. (49) found that ∼50% of heavy chains of early B cell progenitors using VH elements of the VHQ52 or VHJ558 families are unable to pair with the surrogate light chain. IgH chain–independent recombination of IgL chain genes thus might add antigen receptor specificities to the B cell repertoire that would not arise via the pre-B cell receptor– driven pathway.
Frequency of Igκ Gene Rearrangement and Expression in CD43+ B Cell Progenitors.
We found 50 out of 627 fraction C cells and 32 out of 373 fraction B cells harboring VκJκ rearrangements. (We disregard the data obtained from sorted κ chain expressing cells from fraction B, because this cell population was selected for high levels of κ chain expression; see Results.) Taking into account the detection efficiency of the assay (70%) and the proportions of cells bearing VκJκ joints in the absence of productive VHDHJH rearrangements (7 out of 14 in fraction C and 4 out of 11 in fraction B; Table VI), we estimate that 4–7% of cells in fractions B and C carry VκJκ joints in the absence of a productive VHDHJH joint, and a similar proportion of cells contains both VκJκ rearrangement(s) and a productive VHDHJH joint. Overall, the frequency of the cells carrying Igκ gene rearrangements is ∼15% of the total CD43+ B cell progenitor population in wild-type mice. This value correlates well with B cell production observed in λ5-deficient animals, which is reduced by ∼95% (17, 50) and is dependent on the generation of Ig light chains in the absence of pre-B cell receptor function. To obtain 5% of B cells generated in wild-type mice, Igκ genes must be rearranged in 15% of the B cell progenitors, assuming that one-third of the joints are in-frame and that the B cell receptor induces a similar extent of proliferation in the progenitor compartment as does the pre-B cell receptor.
The results reported here are in a good agreement with previous data based on quantitative PCR analyses, in which VκJκ rearrangements represented ∼7 and 15% in fractions B and C, respectively, taking the level of VκJκ rearrangements in splenic B cells as 100% (17). Our results do not contradict the experiments of ten Boekel et al. (16), who did not detect VκJκ rearrangements among 24 cells of early progenitor B cell phenotype (c-kit+, CD25−, B220+). Since this population includes fractions A, B, and C (according to Hardy's classification, reference 23), and no VκJκ rearrangements are detectable in fraction A (17), the frequency of cells bearing VκJκ joints in the population analyzed by ten Boekel and colleagues is expected to be lower than 1 in 24 in these cells.
Immunoglobulin gene transcription and rearrangements are coordinately regulated during B cell development (for review see reference 51). It has been suggested that transcription of unrearranged genes is required for the initiation of the V(D)J joining process. We have observed that in 3-83κi mice carrying a productively rearranged VκJκ joint in the germline there are approximately three times more cells expressing κ light chains in fraction B compared with the wild-type situation (Fig. 1). This difference is quantitatively accounted for by the fact that two-thirds of the newly formed rearrangements in the wild-type cells are nonproductive. Therefore, this result suggests that the “opening” of the Igκ locus for transcription and for recombination occurs simultaneously and may thus be controlled by the same factor(s). Moreover, this result shows that at this early developmental stage wild-type cells rearranging Igκ genes express the recombinatorial products at the protein level.
Acknowledgments
We thank G. Zöbelein, C. Göttlinger and B. Hampel for their excellent technical help. We are grateful to Dr. L. Pao for critical reading of the manuscript.
This work was supported by the Deutsche Forschungsgemeinschaft through SFB 243, the Land Nordrhein-Westfalen, the EU Biotechnology (B104-CT96-0037), and the Human Frontier Science Program.
References
- 1.Alt F, Rosenberg N, Lewis S, Thomas E, Baltimore D. Organization and reorganization of immunoglobulin genes in A-MuLV-transformed cells: rearrangement of heavy but not light chains. Cell. 1981;27:381–390. doi: 10.1016/0092-8674(81)90421-9. [DOI] [PubMed] [Google Scholar]
- 2.Reth MG, Ammirati P, Jackson S, Alt FW. Regulated progression of a cultured pre-B-cell line to the B-cell stage. Nature. 1985;317:353–355. doi: 10.1038/317353a0. [DOI] [PubMed] [Google Scholar]
- 3.Coffman RL, Weissman IL. Immunoglobulin gene rearrangement during pre-B cell differentiation. J Mol Cell Immunol. 1983;1:31–38. [PubMed] [Google Scholar]
- 4.Karasuyama H, Kudo A, Melchers F. The proteins encoded by the VpreBand λ5 preB cell–specific genes can associate with each other and with μ heavy chain. J Exp Med. 1990;172:969–972. doi: 10.1084/jem.172.3.969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Reth MG, Petrac E, Wiese P, Lobel L, Alt FW. Activation of Vκgene rearrangement in pre-B cells follows the expression of membrane-bound immunoglobulin heavy chains. EMBO (Eur Mol Biol Organ) J. 1987;6:3299–3305. doi: 10.1002/j.1460-2075.1987.tb02649.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Iglesias A, Kopf M, Williams GS, Bühler B, Köhler G. Molecular requirements for the μ-induced light chain gene rearrangements in pre-B cells. EMBO (Eur Mol Biol Organ) J. 1991;10:2147–2156. doi: 10.1002/j.1460-2075.1991.tb07749.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Tsubata T, Tsubata R, Reth MG. Crosslinking of the cell surface immunoglobulin (μ-surrogate light chains complex) on pre-B cells induces activation of V gene rearrangements at the immunoglobulin κ locus. Int Immunol. 1992;4:637–641. doi: 10.1093/intimm/4.6.637. [DOI] [PubMed] [Google Scholar]
- 8.Schlissel MS, Morrow T. Ig heavy chain protein controls B cell development by regulating germ-line transcription and retargeting V(D)J recombination. J Immunol. 1994;153:1645–1657. [PubMed] [Google Scholar]
- 9.Coleclough C, Perry RP, Karjalainen K, Weigert M. Aberrrant rearrangements contribute significantly to the allelic exclusion of immunoglobulin gene expression. Nature. 1981;290:372–378. doi: 10.1038/290372a0. [DOI] [PubMed] [Google Scholar]
- 10.Cohn M, Langman RE. The protection: the unit of humoral immunity selected by evolution. Immunol Rev. 1990;115:11–147. doi: 10.1111/j.1600-065x.1990.tb00783.x. [DOI] [PubMed] [Google Scholar]
- 11.Schlissel MS, Baltimore D. Activation of immunoglobulin kappa gene rearrangement correlates with induction of kappa gene transcription. Cell. 1989;58:1001–1007. doi: 10.1016/0092-8674(89)90951-3. [DOI] [PubMed] [Google Scholar]
- 12.Blackwell TK, Malynn BA, Pollock RR, Ferrier P, Covey LR, Fulop GM, Phillips RA, Yancopoulos GD, Alt FW. Isolation of scid pre-B cells that rearrange kappa light chain genes: formation of normal signal and abnormal coding joins. EMBO (Eur Mol Biol Organ) J. 1989;8:735–742. doi: 10.1002/j.1460-2075.1989.tb03433.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Grawunder U, Haasner D, Melchers F, Rolink A. Rearrangement and expression of κ light chain genes can occur without μ heavy chain expression during differentiation of pre-B cells. Int Immunol. 1993;5:1609–1618. doi: 10.1093/intimm/5.12.1609. [DOI] [PubMed] [Google Scholar]
- 14.Kubagawa H, Cooper MD, Carroll AJ, Burrows PD. Light chain gene expression before heavy chain gene rearrangement in pre-B cells transformed by Epstein-Barr virus. Proc Natl Acad Sci USA. 1989;86:2356–2360. doi: 10.1073/pnas.86.7.2356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Benatar T, Tkalec L, Ratcliffe MJH. Stochastic rearrangement of immunoglobulin variable-region genes in chicken B-cell development. Proc Natl Acad Sci USA. 1992;89:7615–7619. doi: 10.1073/pnas.89.16.7615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.ten Boekel E, Melchers F, Rolink A. The status of Ig loci rearrangements in single cells from different stages of B cell development. Int Immunol. 1995;7:1013–1019. doi: 10.1093/intimm/7.6.1013. [DOI] [PubMed] [Google Scholar]
- 17.Ehlich A, Schaal S, Gu H, Kitamura D, Müller W, Rajewsky K. Immunoglobulin heavy and light chain genes rearrange independently at early stages of B cell development. Cell. 1993;72:695–704. doi: 10.1016/0092-8674(93)90398-a. [DOI] [PubMed] [Google Scholar]
- 18.Chen J, Trounstine M, Alt FW, Young F, Kurahara C, Loring JF, Huszar D. Immunoglobulin gene rearrangement in B cell deficient mice generated by targeted deletion of the JHlocus. Int Immunol. 1993;5:647–656. doi: 10.1093/intimm/5.6.647. [DOI] [PubMed] [Google Scholar]
- 19.Kitamura D, Roes J, Kühn R, Rajewsky K. A B cell deficient mouse by targeted disruption of the membrane exon of the immunoglobulin μ chain gene. Nature. 1991;350:423–426. doi: 10.1038/350423a0. [DOI] [PubMed] [Google Scholar]
- 20.Gu H, Zou Y, Rajewsky K. Independent control of Ig switch recombination at individual switch regions evidenced through cre-loxP mediated gene targeting. Cell. 1993;73:1155–1164. doi: 10.1016/0092-8674(93)90644-6. [DOI] [PubMed] [Google Scholar]
- 21.Pelanda R, Schaal S, Torres RM, Rajewsky K. A prematurely expressed Igκ transgene, but not a VκJκ gene segment targeted into the Igκ locus, can rescue B cell development in λ5-deficient mice. Immunity. 1996;5:229–239. doi: 10.1016/s1074-7613(00)80318-0. [DOI] [PubMed] [Google Scholar]
- 22.Hooper M, Hardy K, Handyside A, Hunter S, Monk M. HPRT-deficient (Lesch-Nyhan) mouse embryos derived from germline colonisation by cultured cells. Nature. 1987;326:292–295. doi: 10.1038/326292a0. [DOI] [PubMed] [Google Scholar]
- 23.Hardy RR, Carmack CE, Shinton SA, Kemp JD, Hayakawa K. Resolution and characterization of pro-B and pre-pro-B cell stages in normal mouse bone marrow. J Exp Med. 1991;173:1213–1225. doi: 10.1084/jem.173.5.1213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Miltenyi S, Müller W, Weichel W, Radbruch A. High gradient magnetic cell separation with MACS. Cytometry. 1990;11:231–238. doi: 10.1002/cyto.990110203. [DOI] [PubMed] [Google Scholar]
- 25.Springer T, Galfre G, Secher DS, Milstein C. Monoclonal xenogeneic antibodies to murine cell surface antigens: identification of novel leukocyte differentiation antigens. Eur J Immunol. 1978;8:539–551. doi: 10.1002/eji.1830080802. [DOI] [PubMed] [Google Scholar]
- 26.Gulley ML, Ogata LC, Thorson JA, Dailey MO, Kemp JD. Identification of a murine pan-T cell antigen which is also expressed during the terminal phases of B cell differentiation. J Immunol. 1988;140:3751–3757. [PubMed] [Google Scholar]
- 27.Grützmann, R. 1981. Vergleichende idiotypische Analyse von Rezeptoren mit Spezifität für Histokompatibilitätsantigene. Ph.D. thesis. University of Cologne, Cologne, Germany.
- 28.Taki S, Meiering M, Rajewsky K. Targeted insertion of a variable region gene into the immunoglobulin heavy chain locus. Science. 1993;262:1268–1271. doi: 10.1126/science.8235657. [DOI] [PubMed] [Google Scholar]
- 29.Ehlich A, Martin V, Müller W, Rajewsky K. Analysis of the B cell progenitor compartment at the level of single cells. Curr Biol. 1994;4:573–583. doi: 10.1016/s0960-9822(00)00129-9. [DOI] [PubMed] [Google Scholar]
- 30.Löffert D, Ehlich A, Müller W, Rajewsky K. Surrogate light chain expression is required to establish immunoglobulin heavy chain allelic exclusion during early B cell development. Immunity. 1996;4:133–144. doi: 10.1016/s1074-7613(00)80678-0. [DOI] [PubMed] [Google Scholar]
- 31.Kabat, E.A., T.T. Wu, H.M. Perra, K.S. Gottesman, and C. Foeller. 1991. Sequences of proteins of immunological interest. US Department of Health and Human Services, Washington DC. 2597 pp.
- 32.Kofler R, Duchosal MA, Dixon FJ. Complexity, polymorphism, and connectivity of mouse Vκ gene families. Immunogenetics. 1989;29:65–74. doi: 10.1007/BF00395853. [DOI] [PubMed] [Google Scholar]
- 33.Sonoda E, Pewzner-Jung Y, Schwers S, Taki S, Jung S, Eilat D, Rajewsky K. B cell development under the condition of allelic inclusion. Immunity. 1997;6:225–233. doi: 10.1016/s1074-7613(00)80325-8. [DOI] [PubMed] [Google Scholar]
- 34.Bothwell AL, Paskind M, Reth M, Imanishi-Kari T, Rajewsky K, Baltimore D. Heavy chain variable region contribution to the NPb family of antibodies: somatic mutation evident in a gamma-2a variable region. Cell. 1981;24:625–637. doi: 10.1016/0092-8674(81)90089-1. [DOI] [PubMed] [Google Scholar]
- 35.Ichihara Y, Hayachida H, Miyazawa S, Kurosawa Y. Only DFl16, DSP2, and DQ52 gene families exist in mouse immunoglobulin heavy chain diversity loci, of which DFL16 and DSP2 originate from the same primordial DHgene. Eur J Immunol. 1989;19:1849–1854. doi: 10.1002/eji.1830191014. [DOI] [PubMed] [Google Scholar]
- 36.Reth MG, Alt FW. Novel immunoglobulin heavy chains are produced from DJHgene segment rearrangements in lymphoid cells. Nature. 1984;312:418–423. doi: 10.1038/312418a0. [DOI] [PubMed] [Google Scholar]
- 37.Nemazee DA, Burki K. Clonal deletion of B lymphocytes in a transgenic mouse bearing anti-MHC class I antibody genes. Nature. 1989;337:562–566. doi: 10.1038/337562a0. [DOI] [PubMed] [Google Scholar]
- 38.Horne MC, Roth PE, DeFranco AL. Assembly of the truncated immunoglobulin heavy chain Dμ into antigen receptor-like complexes in pre-B cells but not in B cells. Immunity. 1996;4:145–158. doi: 10.1016/s1074-7613(00)80679-2. [DOI] [PubMed] [Google Scholar]
- 39.Tornberg UC, Bergqvist I, Haury M, Holmberg D. Regulation of B lymphocyte development by the truncated immunoglobulin heavy chain protein Dμ. J Exp Med. 1998;187:703–709. doi: 10.1084/jem.187.5.703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Reth MG, Gehrmann P, Petrac E, Wiese P. A novel VH to DJHjoining mechanism in heavy-chain-negative (null) pre-B cells results in heavy-chain production. Nature. 1986;322:840–842. doi: 10.1038/322840a0. [DOI] [PubMed] [Google Scholar]
- 41.Kleinfield R, Hardy RR, Tarlinton D, Dangl J, Herzenberg LA, Weigert M. Recombination between an expressed immunoglobulin heavy-chain gene and a germline variable gene segment in a Ly 1+ B-cell lymphoma. Nature. 1986;322:843–846. doi: 10.1038/322843a0. [DOI] [PubMed] [Google Scholar]
- 42.Maeda T, Sugiyama H, Tani Y, Kishimoto S. The DJH complex remains active in recombination to VHsegments after the loss of μ-chain expression in μ-positive pre-B cells. J Immunol. 1989;142:3652–3656. [PubMed] [Google Scholar]
- 43.Fitts MG, Mage RG. Secondary rearrangements and post-rearrangement selection contribute to restricted immunoglobulin DJHexpression in young rabbit bone marrow. Eur J Immunol. 1995;25:700–707. doi: 10.1002/eji.1830250311. [DOI] [PubMed] [Google Scholar]
- 44.Grawunder U, Leu TMJ, Schatz DG, Werner A, Rolink AG, Melchers F, Winkler TH. Down-regulation of RAG1 and RAG2 gene expression in preB cells after functional immunoglobulin heavy chain rearrangement. Immunity. 1995;3:601–608. doi: 10.1016/1074-7613(95)90131-0. [DOI] [PubMed] [Google Scholar]
- 45.Löffert D, Schaal S, Ehlich A, Hardy RR, Zou Y-R, Müller W, Rajewsky K. Early B-cell development in the mouse: insights from mutations introduced by gene targeting. Immunol Rev. 1994;137:135–153. doi: 10.1111/j.1600-065x.1994.tb00662.x. [DOI] [PubMed] [Google Scholar]
- 46.Fang W, Mueller DL, Pennel CA, Rivard JJ, Li Y-S, Hardy RR, Schlissel MS, Behrens TW. Frequent aberrant immunoglobulin gene rearrangements in pro-B cells revealed by a bcl-xL transgene. Immunity. 1996;4:291–299. doi: 10.1016/s1074-7613(00)80437-9. [DOI] [PubMed] [Google Scholar]
- 47.Decker DJ, Boyle NE, Koziol JA, Klinman NR. The expression of the IgH chain repertoire in developing bone marrow B lineage cells. J Immunol. 1991;146:350–361. [PubMed] [Google Scholar]
- 48.Li YS, Hayakawa K, Hardy RR. The regulated expression of B lineage associated genes during B cell differentiation in bone marrow and fetal liver. J Exp Med. 1993;178:951–960. doi: 10.1084/jem.178.3.951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.ten Boekel E, Melchers F, Rolink AG. Changes in the V(H) gene repertoire of developing precursor B lymphocytes in mouse bone marrow mediated by the pre-B cell receptor. Immunity. 1997;7:357–368. doi: 10.1016/s1074-7613(00)80357-x. [DOI] [PubMed] [Google Scholar]
- 50.Kitamura D, Kudo A, Schaal S, Müller W, Melchers F, Rajewsky K. A critical role of λ5 protein in B cell development. Cell. 1992;69:823–831. doi: 10.1016/0092-8674(92)90293-l. [DOI] [PubMed] [Google Scholar]
- 51.Sleckman BP, Gorman JR, Alt FW. Accessibility control of antigen-receptor variable-region gene assembly: role of cis-acting elements. Annu Rev Immunol. 1996;14:459–481. doi: 10.1146/annurev.immunol.14.1.459. [DOI] [PubMed] [Google Scholar]
- 52.Strohal R, Holmberg A, Kroemer G, Kofler R. Mouse Vκgene classification by nucleic acid sequence similarity. Immunogenetics. 1989;30:475–493. doi: 10.1007/BF02421180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Alt FW, Baltimore D. Joining of immunoglobulin heavy chain gene segments: Implication from a chromosome with evidence of three D-JHfusions. Proc Natl Acad Sci USA. 1982;79:4118–4122. doi: 10.1073/pnas.79.13.4118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Desiderio SV, Yancopoulos GD, Paskind M, Thomas E, Boss MA, Landau N, Alt FW, Baltimore D. Insertion of N regions into heavy-chain genes is correlated with expression of terminal deoxytransferase in B cells. Nature. 1984;311:752–755. doi: 10.1038/311752a0. [DOI] [PubMed] [Google Scholar]
- 55.Lafaille JJ, DeCloux A, Bonneville M, Takagaki Y, Tonegawa S. Junctional sequences of T cell receptor γδ genes: implications for γδ T cell lineages and for a novel intermediate of V(D)J joining. Cell. 1989;59:859–870. doi: 10.1016/0092-8674(89)90609-0. [DOI] [PubMed] [Google Scholar]
- 56.Chang Y, Paige CJ, Wu GE. Enumeration and characterisation of DJHstructures in mouse fetal liver. EMBO (Eur Mol Biol Organ) J. 1992;11:1891–1899. doi: 10.1002/j.1460-2075.1992.tb05241.x. [DOI] [PMC free article] [PubMed] [Google Scholar]