

Abstract
The susceptibility to infections induced by Gram-negative bacteria is largely determined by innate immune responses to bacteria cell wall lipopolysaccharide (LPS). The stimulation of B cells by LPS enhances their antigen-presenting capacity and is accompanied by B cell proliferation and secretion of large quantities of LPS-neutralizing antibodies. Similar to macrophages and neutrophils, the LPS-induced activation of B cells is dependent on Toll-like receptor (TLR)4. Here, we demonstrate that the responses of B cells to LPS are also regulated by another TLR protein, RP105, which is predominantly expressed on mature B cells in mice and humans. The analysis of mice homozygous for the null mutation in the RP105 gene revealed impaired proliferative and humoral immune responses of RP105-deficient B cells to LPS. Using originally LPS-unresponsive Ba/F3 cells expressing exogenous TLR4 and RP105, we demonstrate the functional cooperation between TLR4 and RP105 in LPS-induced nuclear factor κB activation. These data suggest the existence of the TLR4–RP105 signaling module in the LPS-induced B cell activation.
Keywords: gene targeting, lymphocyte, activation, leucine-rich repeat, cell surface molecule
Introduction
A large body of evidence supports the important role of B cells in host responses against Gram-negative bacteria. These responses are largely triggered by bacteria cell wall LPS. Stimulation of B cells by LPS enhances their antigen-presenting capacity and is accompanied by B cell proliferation and secretion of large quantities of LPS-neutralizing Abs 1 2. In turn, LPS-neutralizing Abs are important for clearing LPS and Gram-negative bacteria out of the circulation 3. The responses of B cells to LPS are mediated by cell surface Toll-like receptor (TLR) 4, which belongs to the multimember family of mammalian proteins homologous to Toll protein of Drosophila melanogaster 4 5. The TLR proteins possess an extracellular leucine-rich repeat (LRR) domain, a single transmembrane domain, and a cytoplasmic domain homologous to the cytoplasmic portion of the IL-1 receptor 5. The significance of TLR4 in B cell activation by LPS was underscored by profoundly reduced in vitro and in vivo LPS responses of B cells harboring spontaneous or genetically engineered null mutations in the TLR4 gene 6 7 8. The LPS signaling via TLR4 is dependent on association of the LRR domain of TLR4 with the MD-2 protein produced by TLR4-expressing cells 9.
The essential role of TLR4 in B cell responses to LPS is contrasted by the relatively low expression levels of the TLR4–MD-2 complex on the surface of ex vivo–isolated mature B cells compared with peritoneal macrophages. Using the MTS510 mAb that in flow cytometry analyses recognizes TLR4 in association with MD-2 but not TLR4 alone, we found the expression levels of TLR4–MD-2 on B cells to be >10 times lower than on the peritoneal thioglycollate–elicited macrophages 10. The hardly detectable expression of TLR4–MD-2 on B cells suggests the possible cooperation between TLR4 and other TLR protein(s) in B cell signaling of LPS. In addition to TLR4, B cells express another member of the TLR family, the RP105 protein 11 12 13. In contrast to TLR4, which is expressed on immune cells of different types, the expression of RP105 is largely restricted to mature peripheral B cells 11. RP105 is associated with the MD-2 homologous protein MD-1 14 15. Robust B cell proliferation and upregulation of the cell surface–expressed MHC class II and costimulatory proteins B7.1 and B7.2 induced by Ab-mediated cross-linking of RP105 revealed this TLR as a potent regulator of B cell activation 11 16. To test whether RP105 controls B cell responses to LPS, as well as to address the possible role of RP105 in B cell development and activation, we have generated mice homozygous for the null mutation of the RP105 gene. Using mice deficient for RP105, we revealed an important role of RP105 in B cell activation by LPS in vitro and in vivo.
Materials and Methods
Generation of the RP105-deficient Mice.
The DNA clone containing the RP105 gene was obtained from Genome Systems, Inc., and the RP105 gene was subcloned into the pBluescript vector (Stratagene). For the construction of the RP105 gene-targeting vector, the 2.4-kB fragment of the exon 3 of the RP105 gene was replaced by the neomycin resistance (neor) gene under the control of the mouse phosphoglycerate kinase (PGK)-1 gene promoter. The HSV thymidine kinase gene driven by the PGK-1 promoter was introduced at the 3′ end of the targeting vector. The linearized DNA of the targeting vector was transfected into the E14.1 embryonic stem (ES) cells, followed by selection of the G418- and gancyclovir-resistant ES cell clones as described previously 17. The clones of double-resistant cells were collected and genotyped by Southern blot analysis using the probe located outside the targeting construct. Cells of three independent recombinant ES clones were used to produce chimeric mice, followed by the generation of heterozygous RP105+/− and homozygous RP105−/− mice. The RP105+/− mice were backcrossed four times to C57BL/6 mice and RP105−/− mice (F5) at 6–15 wk of age, and were used for most of the experiments. The wild-type littermates derived from the breeding of the RP105+/− mice were used as control. C3H/HeJ and C3H/HeN mice were obtained from Japan SLC, Inc. Mice were maintained in the animal facilities at Saga Medical School and Institute for Genetics.
Cell Staining and Flow Cytometry.
Single cell suspensions were prepared from different lymphoid organs and incubated at 106 cells per 20 μl on ice in staining buffer (PBS containing 0.5% BSA and 0.01% NaN3) with optimal amounts of FITC-, PE-, or biotin-conjugated Abs for 10 min. The following mAbs were purchased from BD PharMingen: S7 (anti-CD43), B3B4 (anti-CD23), and Ly1 (anti-CD5). The following Abs were prepared: RA3-6B2 (anti-B220), R33-24.12 (anti-IgM), 1.3-5 (anti-IgD), and Cfo-1 (anti-Thy1.2). Flow cytometric analysis was performed on a FACScan™ cytometer (Becton Dickinson).
Analysis of B Cell Proliferation In Vitro.
Splenic B cells were purified by negative selection with the anti-CD43 mAb S7 conjugated either to Dynabeads M-450 sheep anti–rat IgG (Dynal) or to CD43-coupled magnetic beads (Miltenyi Biotec) as described 18. B cell purity has been always >95%, as judged by flow cytometry analyses (data not shown). The wild-type and mutant B cells were incubated at 2 × 105 per well in 96-well flat-bottomed plates with goat anti–mouse IgM F(ab′)2 (Organon Teknika) in the presence or absence of recombinant mouse IL-4 (R&D Systems) or of rat anti–mouse CD40 mAb (LB429S; a gift from Dr. Nobuo Sakaguchi, Kumamoto University School of Medicine, Kumamoto, Japan). For the analysis of LPS responses, B cells were treated with LPS derived from Escherichia coli 0111:B4 (Sigma-Aldrich), E. coli 055:B5 (Sigma-Aldrich), or Salmonella minnesota Re595 (Sigma-Aldrich). Lipid A was either of bacterial (S. minnesota) or synthetic (a gift from Dr. Teruo Kirikae, Daiichi Pure Chemicals, Tokyo, Japan) origin.
Analysis of Lyn Phosphorylation.
Purified B cells (107 per group) were stimulated with lipid A (1 μg/ml) and harvested. Cells were solubilized in lysis buffer (1% Triton X-100, 150 mM NaCl, 50 mM Tris/HCl, pH7.5, 10 μg/ml soybean trypsin inhibitor, 1 mM PMSF, and 5 mM EDTA) supplemented with phosphatase inhibitors (10 mM NaF and 1 mM sodium vanadate). Lysates were applied to protein G Sepharose conjugated with anti-Lyn mAb Lyn8 (Wako Pure Chemicals). Precipitates (2.5 × 106 cells per lane) were subjected to SDS-PAGE (9% acrylamide) and electroblotting. Phosphotyrosine or Lyn was probed with antiphosphotyrosine mAb 4G10 (Upstate Biotechnology) or with Lyn8, respectively. Detection was conducted with goat anti–mouse IgG peroxidase and the SuperSignal chemiluminescence substrate (Pierce Chemical Co.).
Analysis of Humoral Immune Responses.
The littermate control and RP105-deficient mice were immunized intraperitoneally with 20 μg of TNP-LPS in PBS (three to five mice per group). The serum concentration of TNP-specific Abs at different time points was measured by ELISA. ELISA was performed by coating plastic plates with TNP-BSA (10 μg/ml), and serial serum dilutions were applied onto the plate. Bound Abs were revealed by goat Abs specific for IgM or IgG3 isotype (Caltag Labs.).
The analyses of serum concentrations of the Abs of different isotypes in mice immunized with T cell–independent and T cell–dependent antigens were done as described 18. In brief, wild-type, RP105+/−, and RP105−/− mice were immunized intraperitoneally with 5 μg of the T-independent antigen 4-hydroxy3-nitrophenylacetyl (NP)25-Ficoll in PBS or 50 μg of the alum-precipitated T-dependent antigen NP15–chicken γ-globulin (CG). The serum concentration of NP-specific Abs was measured by ELISA.
Analysis of the Nuclear Factor κB Activation in Ba/F3 Cells.
The IL-3–dependent Ba/F3 cells 19 carrying the luciferase gene under the control of the nuclear factor (NF)-κB responsive element 20 were used to study the role of TLR4 and RP105 in the LPS-induced NF-κB activation. The pEF-BOS expression vectors 21 encoding the RP105, TLR4, MD-1, or MD-2 proteins of human or mouse origin were constructed as described 9 10 12 14 15. These vectors, as well as the reporter gene, were transfected in the Ba/F3 cells as described previously 9 10. Ba/F3 cells expressing TLR4–MD-2, TLR4–RP105, or TLR4–RP105–MD-1 were established by sequential transfection of those expressing TLR4 and the reporter gene. Cell surface expression of human TLR4 and TLR4–MD-2 was shown previously 9. Expression of human RP105 and MD-1 was determined with the anti-RP105 mAb or the Ab against FLAG epitope (Sigma-Aldrich) that had been attached at the COOH terminus of MD-1, respectively. The expression levels of mouse RP105–MD-1 and TLR4–MD-2 were determined by FACS® using the mAbs RP404 and MTS510, which recognize RP105–MD-1 and TLR–MD2 complexes, respectively, but not RP105 or TLR4 alone (10; our unpublished results). Expression of mouse TLR4 alone was determined by immunoblot analysis with the Abs to the FLAG epitope that had been attached at the COOH terminus of TLR4 10. Transfected Ba/F3 cells were treated with lipid A at various concentrations for 4 h. The luciferase activity was measured in cell lysates as described 9.
Results and Discussion
To test whether RP105 controls B cell responses to LPS, as well as to address the possible role of RP105 in B cell development and activation, we have generated mice homozygous for the null mutation of the RP105 gene. The exon 3 of the RP105 gene, encoding 575 of 661 amino acid residues of RP105 protein, was replaced by the neor gene in E14.1 ES cells, which were used to generate R105−/− mice (Fig. 1, a and b). The targeted modification of the RP105 gene eliminates the region encoding most of the extracellular LRR domain, the transmembrane, and cytoplasmic domains. As expected, B cells derived from RP105−/− mice do not express RP105 on the cell surface (Fig. 1 c), and because of the replacement of almost an entire RP105 gene by neor are unlikely to express potentially functional truncated RP105 protein.
Figure 1.
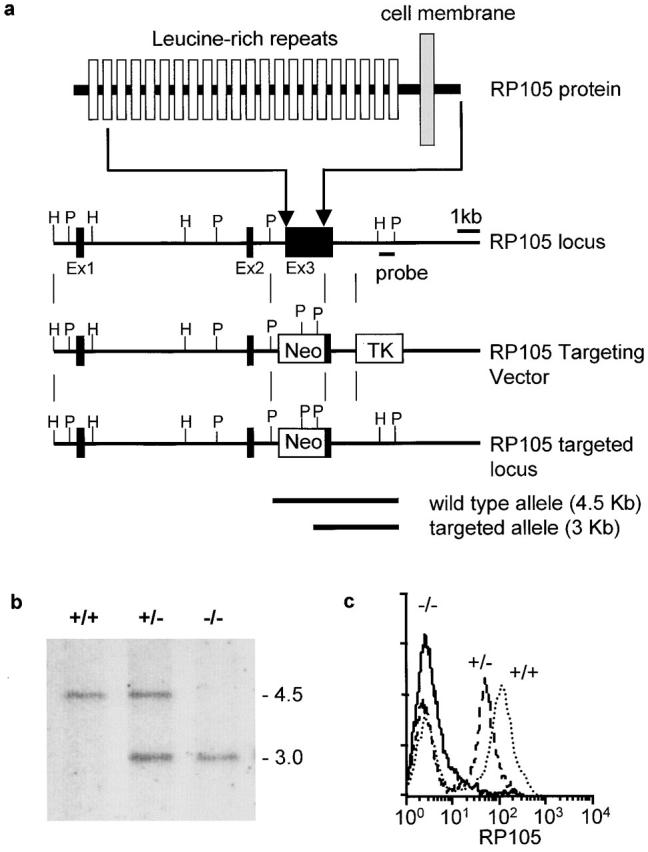
Targeted disruption of the murine RP105 gene by homologous recombination. (a) Schematic representation of the RP105 protein, RP105 genomic locus, targeting construct, and targeted RP105 locus. Positions of RP105 exons are shown as boxes. The indicated external probe recognizes 4.5 Kb and 3 Kb of the PstI-digested DNA of the wild-type and targeted RP105 genes, respectively. (b) Southern blot analysis of the RP105 mutation in mice. The tail DNA isolated from the wild-type C57BL/6, RP105+/−, and RP105−/− mice was digested with Pst1 and analyzed using the probe described above. Arrows indicate the position of the DNA fragments corresponding to the wild-type (4.5 Kb) or targeted (3 Kb) RP105 alleles. (c) Flow cytometry staining of RP105 was conducted as described in Materials and Methods. Spleen cells from the wild-type C57BL/6, RP105+/−, and RP105−/− mice were stained with an anti-RP105 mAb (RP/14) followed by goat anti–rat IgG-FITC.
The flow cytometric analyses and absolute numbers of the bone marrow cells, splenocytes, lymph nodes, and peritoneal cells derived from RP105-deficient mice did not reveal changes in sizes of developing and mature B cell subpopulations compared with the control wild-type mice (Fig. 2; Table ). These data exclude the significant role of RP105 in B cell development.
Figure 2.
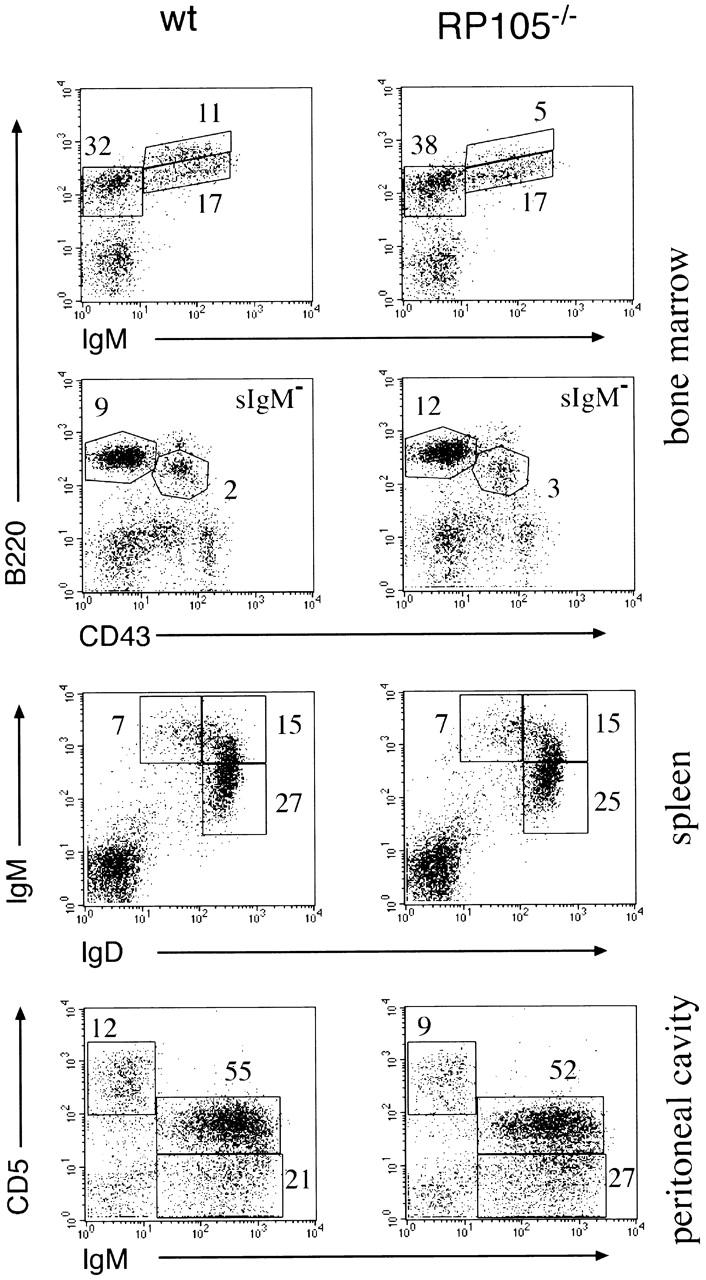
B lymphocyte populations in RP105-deficient mice. Flow cytometric analysis of bone marrow cells, splenocytes, thymocytes, and peritoneal cavity cells in 8-wk-old wild-type C57BL/6 and RP105-deficient mice. Numbers indicate the fraction (%) of gated cellular subpopulations within the lymphocyte population.
Table 1.
Absolute Numbers of B Lineage Cells in Lymphoid Organs
| RP105+/+ | RP105−/− | |||
|---|---|---|---|---|
| Exp. 1 | Exp. 2 | Exp. 1 | Exp. 2 | |
| (×10−6) | (×10−6) | (×10−6) | (×10−6) | |
| Bone marrow | ||||
| Pro-B | 0.26 | 0.4 | 0.3 | 0.4 |
| Pre-B | 0.78 | 1.2 | 0.9 | 2.0 |
| Immature B | 0.58 | 0.8 | 0.6 | 1.0 |
| Recirculating B | 0.25 | 0.5 | 0.45 | 0.3 |
| Spleen | ||||
| B | 21.0 | 14.0 | 40.0 | 22.0 |
| T | 9.0 | 8.6 | 23.0 | 17.0 |
| Lymph nodes | ||||
| B | 1.6 | ND | 2.8 | ND |
| T | 9.0 | ND | 10.0 | ND |
| Peritoneal cavity | ||||
| CD5+IgM+ B | 1.9 | 2.1 | 0.6 | 0.5 |
| CD5−IgM− B | 0.8 | 0.7 | 0.4 | 0.3 |
Cells of lymphoid organs were counted, and absolute numbers of lymphoid cells were calculated from the percentage of each cell population. ND, not done; Exp., experiment.
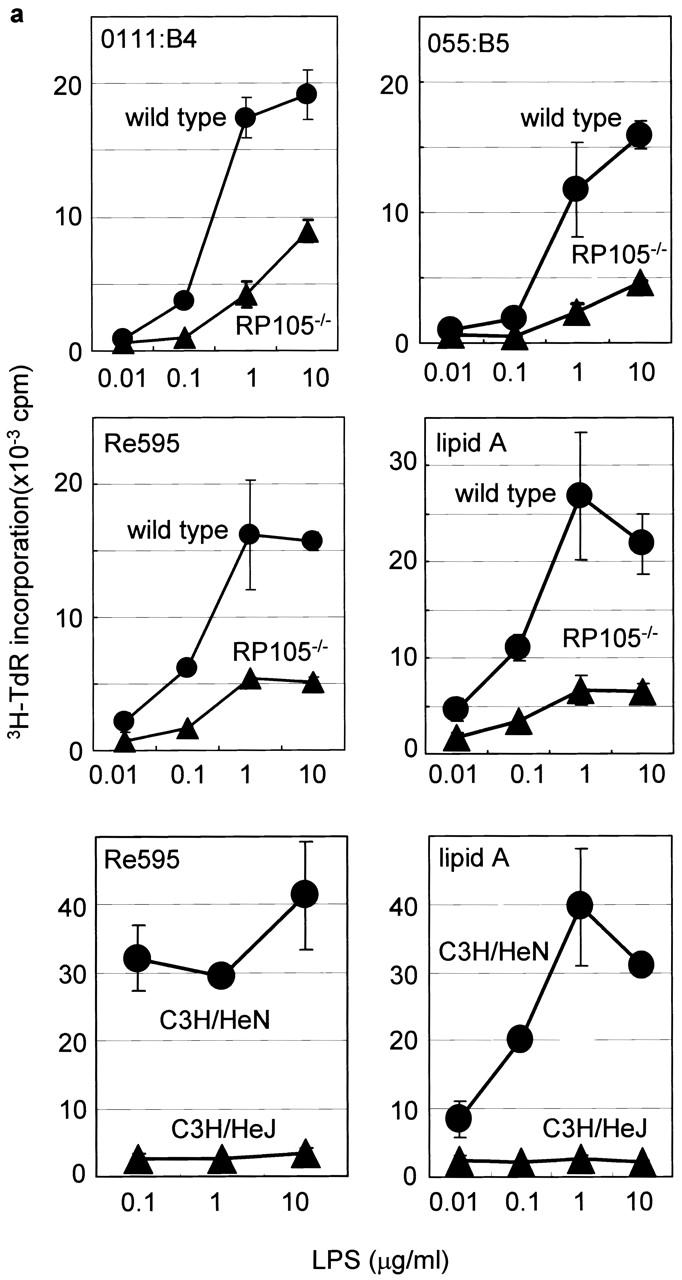
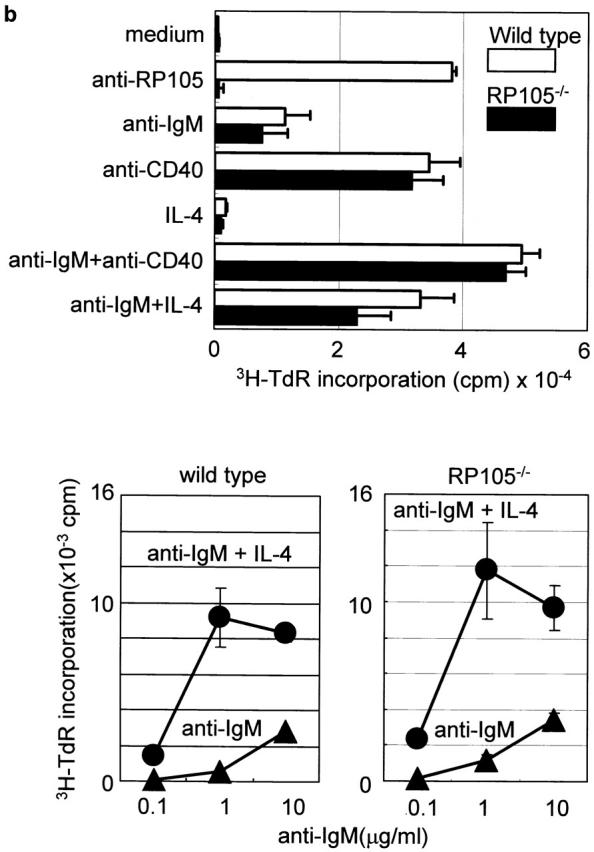
The responsiveness of RP105-deficient B cells to LPS was tested in vitro. Using the MTS510 mAb that recognizes TLR4 in association with MD-2, but not TLR4 alone, we found the expression levels of TLR4–MD2 on wild-type and RP105-defcient cells to be >10 times lower than on the surface of peritoneal thioglycollate–elicited macrophages 10. Furthermore, incubation of the wild-type and RP105-defcient B cells with LPS, anti-IgM, or anti-CD40 Abs did not upregulate surface expression of TLR4–MD-2 (data not shown). Incubation of the purified splenic RP105-deficient B cells with different amounts of LPS derived from three different bacteria strains (E. coli 0111: B4, E. coli 055: B5, and S. minnesota Re595) as well as with synthetic lipid A results in B cell proliferation at levels significantly lower than of LPS-triggered wild-type control B cells (Fig. 3 a). However, RP105-deficient B cells were not as impaired as TLR4-mutated C3H/HeJ B cells, which did not respond to LPS or lipid A (Fig. 3 a). Reduced proliferative response of RP105-deficient B cells was due to the reduced number of cells entering the cell cycle within the first 24 h after the stimulation with LPS, and could not be overcome by costimulation of B cells with IL-4 (data not shown). In contrast to impaired proliferation in response to LPS, the proliferative responses of RP105-deficient B cells and control wild-type B cells to the Abs to IgM, CD40, or CD38 and IL-4 were similar (Fig. 3 b; data not shown). These data indicate the specific role of RP105 in B cell responses to LPS.
Figure 3.
B cell activation in vitro. (a) Purified cells were stimulated with LPS from E. coli 0111:B4, E. coli 055:B5, S. minnesota Re595, and synthetic lipid A. After 3 d, cells were pulsed with 1 μCi/well 3H-TdR and harvested, and [3H]thymidine incorporation was counted as described (reference 9). Results were shown as mean values of cpm from triplicate wells with standard errors. (b) Top: purified B cells were stimulated with the following stimuli: the goat anti–mouse IgM F(ab′)2 Ab (5 μg/ml), the mAb to CD40 (5 μg/ml), and recombinant mouse IL-4 (5 ng/ml). After 3 d, cells were pulsed with 1 μCi/well 3H-TdR, and B cell proliferation was measured as described above. (b) Bottom: purified B cells were stimulated with goat anti–mouse IgM F(ab′)2 Abs at various concentrations with or without IL-4, and proliferation was measured as described above. (c) Purified B cells were stimulated with lipid A (1 μg/ml) for 1 h (lanes 2 and 5) or 3 h (lanes 3 and 6). Cell lysates were immunoprecipitated with anti-Lyn mAb, and equally divided precipitates were subjected to SDS-PAGE (9% acrylamide) and electroblotting. Phosphotyrosine (top) or Lyn (bottom) was probed with antiphosphotyrosine or anti-Lyn mAb, respectively. Similar results were obtained from two independent experiments.

Lyn-deficient B cells are impaired in LPS-induced B cell proliferation, demonstrating an important role of Lyn in B cell signaling of LPS 22 23. Our earlier studies revealed the involvement of the Lyn–Btk–protein kinase C β signaling chain in B cell activation by RP105 16. It seems possible that RP105 has a role in LPS-induced tyrosine phosphorylation of Lyn. We examined tyrosine phosphorylation of Lyn after LPS stimulation in wild-type and RP105-deficient B cells (Fig. 3 c). A slight but appreciable increase in phosphorylation was observed 1 h after LPS addition in wild-type B cells (Fig. 3 c, lanes 1–3), whereas LPS-induced Lyn phosphorylation appeared to be weaker in RP105-deficient B cells (Fig. 3 c, lanes 4–6).
The positive role of RP105 in B cell activation by LPS was further confirmed by the analysis of humoral immune responses in RP105-deficient mice immunized with TNP-LPS. The titers of TNP-specific IgM and IgG3 were significantly lower in the sera of the immunized RP105-deficient mice compared with control mice (Fig. 4 a). However, immunization with T cell–independent antigen NP-Ficoll and T cell–dependent antigen NP-CG resulted in equally strong immune responses in both wild-type and RP105-deficient mice (Fig. 4b and Fig. c). Furthermore, similar titers of serum immunoglobulins of different isotypes in RP105-deficient and wild-type control mice (data not shown) suggest unaltered humoral responses of RP105-deficient B cells to environmental antigens present in the conventional animal facility.
Figure 4.
Ab responses to TNP-LPS, NP-Ficoll, and NP-CG in RP105-deficient mice. (a) Littermate control and RP105-deficient mice were immunized intraperitoneally with TNP-LPS (20 μg per mouse) and bled 1 or 2 wk later. The titers of TNP-specific IgM or IgG3 were measured. (b) NP-specific Abs in sera of wild-type (□), RP105+/− (⋄), and RP105−/− (○) mice immunized with T-independent antigen NP-Ficoll. The titers of NP-specific IgM, IgG3, and λ-bearing Abs were determined at days 0, 7, 14, and 33 after immunization. (c) NP-specific Abs in sera of wild-type (□), RP105+/− (⋄), and RP105−/− (○) mice immunized with T-dependent antigen NP-CG. The titers of NP-specific IgM, IgG1, and λ-bearing Abs were determined at days 0, 7, 14, 21, and 28 after immunization.


The involvement of RP105 in LPS signaling suggests the possible function of RP105 either as a TLR4-independent LPS receptor or as a coreceptor of TLR4. Considering unresponsiveness in C3H/HeJ mice (Fig. 3 a), RP105–MD-1 cannot substitute for the lack of TLR4. We showed previously that association of MD-2 greatly enhances LPS signaling via TLR4 9 10. However, the TLR4–MD-2 complex is hardly detectable on the B cell surface with the MTS510 mAb that specifically reacts with the TLR4–MD-2 complex 10. Moreover, the MTS510 mAb has little effect on LPS-dependent proliferation of B cells (our unpublished results). B cells may employ another molecule to regulate LPS signaling via TLR4. We hypothesized that the RP105–MD-1 complex, instead of MD-2, helps TLR4 to recognize and signal LPS in B cells. The functional cooperation between TLR4 and RP105 was tested in murine Ba/F3 cells that express neither TLR4 nor RP105 and do not respond to LPS 9. The Ba/F3 cells carrying the stably integrated exogenous TLR4, RP105, MD-1, or MD-2 genes alone or in different combinations were used. The expression levels of TLR4 or RP105 in the presence or absence of coexpressed RP105–MD-1 or TLR4, respectively, were similar as judged by immunoprecipitation or cell surface staining with the corresponding Abs (10; data not shown). The LPS-induced activation of NF-κB in these cells was tested by the analysis of the NF-κB–induced upregulation of the luciferase gene activity controlled by the NF-κB responsive element 20. The Ba/F3 cells expressing RP105–MD-1 did not respond to LPS derived from S. minnesota Re595 or lipid A (Fig. 5 a; data not shown). On the other hand, Ba/F3 cells expressing mouse TLR4 alone respond to LPS, albeit at levels significantly reduced compared with Ba/F3 cells expressing TLR4 and MD-2 (Fig. 5 a; reference 10). The RP105 influence on the LPS signaling via TLR4 was addressed by expression of RP105 with or without MD-1 and TLR4 in Ba/F3 cells. The LPS signaling via TLR4 alone was not affected by expression of RP105 alone, whereas it was significantly enhanced in the presence of RP105–MD-1 (Fig. 5 a). We also examined Ba/F3 cells expressing both TLR4–MD-2 and RP105–MD-1, the LPS responses of which were not different from Ba/F3 cells expressing TLR4–MD-2 (Fig. 5 b).
Figure 5.
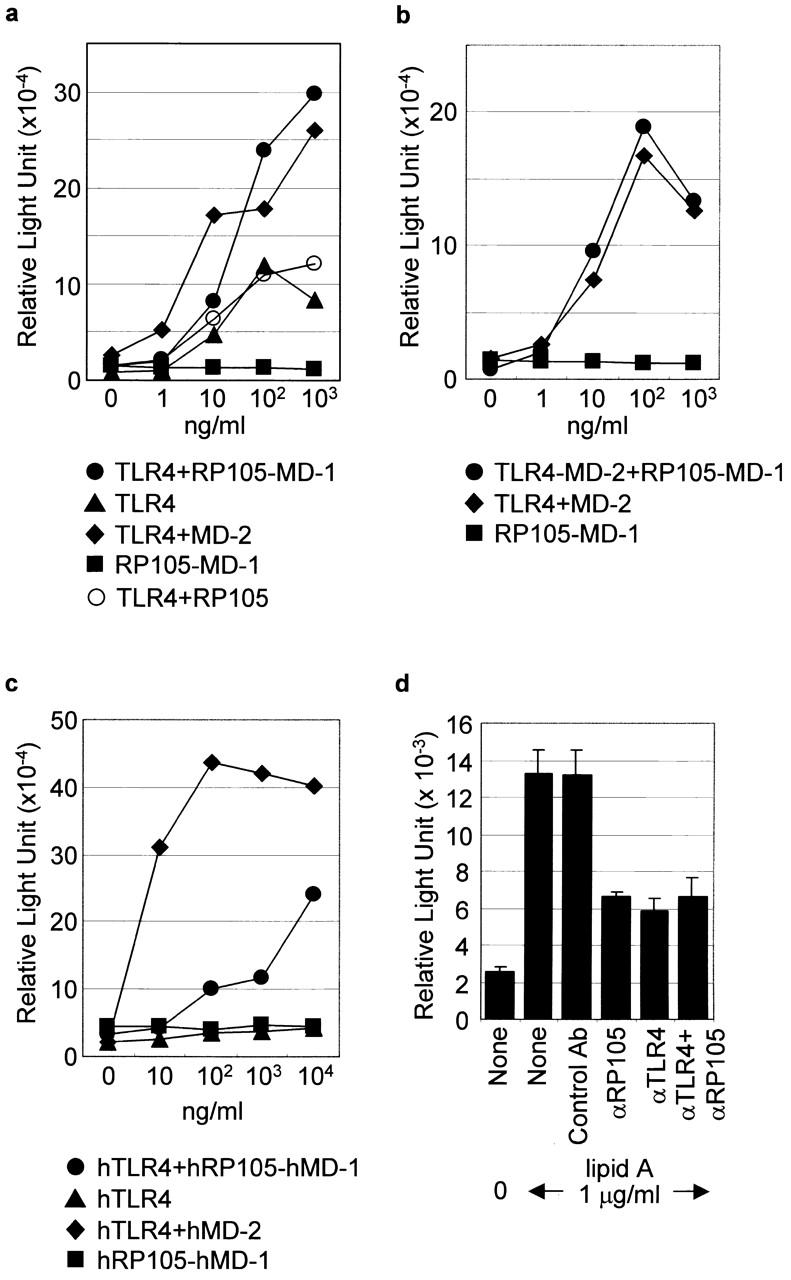
Functional association of TLR4 and RP105. (a and b) Ba/F3 cells expressing mouse TLR4, RP105, MD-2, or MD-1 as indicated were stimulated with lipid A (a) or LPS from S. minnesota Re595 (b) at different concentrations, and NF-κB activation was measured by luciferase activity in cell lysates as described (reference 9). (c) Ba/F3 cells expressing the human TLR4, RP105, MD-1, and MD-2 genes as indicated were stimulated with lipid A, and NF-κB activation was measured as described above. (d) The Ba/F3 cells expressing human TLR4 and RP105–MD-1 were stimulated with lipid A (1 μg/ml). Indicated mAbs (MHR73 for anti-RP105 and HTA125 for anti-TLR4, 20 μg/ml of each) were included during stimulation, and NF-κB activation was measured as described above. Results are represented as mean values from triplicate wells.
Roles of human RP105–MD-1 in the LPS signaling via human TLR4 were next addressed. Similar experiments were conducted with Ba/F3 cells expressing human TLR4 and RP105. The surface expression levels of TLR4 or RP105 in the presence or absence of coexpressed RP105–MD-1 or TLR-4, respectively, were similar (9; data not shown). In contrast to the Ba/F3 cells expressing murine TLR4, the Ba/F3 cells that express human TLR4 alone do not respond to lipid A (Fig. 5 c; reference 9). The reason for the observed species-specific differences in LPS signaling via murine and human TLR4 alone is unknown. Coexpression of RP105–MD-1 conferred LPS responsiveness less efficiently than that of MD-2 (Fig. 5 c). Treatment of Ba/F3 cells coexpressing human TLR4 and RP105–MD-1 with the mAb directed against human RP105 inhibited LPS-induced NF-κB activation as effectively as with anti-TLR4 mAb alone (Fig. 5 d). Treatment of Ba/F3 cells coexpressing human TLR4 and RP105–MD-1 with a combination of the anti-RP105 and anti-TLR4 mAbs did not result in further decrease in LPS-induced NF-κB activation (Fig. 5 d), thus supporting the cooperative rather than independent signaling capacities of TLR4 and RP105–MD-1.
RP105–MD-1 is thus able to enhance LPS signaling via TLR4 as MD-2 does. Either MD-2 or RP105–MD-1 is required for LPS recognition and signaling by TLR4. However, the TLR4–MD-2 complex is hardly detectable on the mouse B cell surface 10, where RP105–MD-1 is abundantly expressed. Taken together with the results with Ba/F3 transfectants, we would like to conclude that the RP105–MD-1 complex, rather than MD-2, regulates LPS signaling via TLR4 on the B cell surface. The structural basis for the RP105–MD-1 and TLR4 cooperation remains unclear. Immunoprecipitation of TLR4 or RP105 from lysates of splenic B cells or Ba/F3 cells coexpressing RP105–MD-1 and TLR4 has not resulted in coprecipitation of TLR4 or RP105, respectively (data not shown). Immunoprecipitation in the presence of LPS did not make any difference.
Finally, the important role of RP105 in B cell activation by LPS makes it a possible target for pharmacological intervention aimed at enhancement of humoral immune response against Gram-negative bacteria.
Acknowledgments
We thank Dr. Paul W. Kincade for his encouragement and support during the initial phase of the project. Drs. Yoshihiro Miura, Rintaro Shimazu, Kenji Fukudome, and Teruo Kirikae are also appreciated for their help and technical suggestions.
The study was supported in part by grants from Uehara Memorial Foundation and Monbusho (the Ministry of Education, Science, and Culture of Japan). This work was also supported by the Deutsche Forschungsgemeinschaft through SFB243 (to I. Su, K. Rajewsky, and A. Tarakhovsky) and by the Fonds der Chemie (A. Tarakhovsky). I. Mecklenbräuker is supported by a Graduiertenkolleg fellowship from the Deutsche Forschungsgemeinschaft.
Footnotes
H. Ogata and I. Su contributed equally to this work.
Abbreviations used in this paper: CG, chicken γ-globulin; ES, embryonic stem; LRR, leucine-rich repeat; neor, neomycin resistance; NF, nuclear factor; TLR, Toll-like receptor.
References
- Kearney J.F., Lawton A.R. B lymphocyte differentiation induced by lipopolysaccharide. I. Generation of cells synthesizing four major immunoglobulin classes. J. Immunol. 1975;115:671–676. [PubMed] [Google Scholar]
- Oliver A.M., Martin F., Kearney J.F. IgMhighCD21high lymphocytes enriched in the splenic marginal zone generate effector cells more rapidly than the bulk of follicular B cells. J. Immunol. 1999;162:7198–7207. [PubMed] [Google Scholar]
- Reid R.R., Prodeus A.P., Khan W., Hsu T., Rosen F.S., Carroll M.C. Endotoxin shock in antibody-deficient mice. Unraveling the role of natural antibody and complement in the clearance of lipopolysaccharide. J. Immunol. 1997;159:970–975. [PubMed] [Google Scholar]
- Medzhitov R., Preston-Hurlburt P., Janeway C.A. A human homologue of the drosophila Toll protein signals activation of adaptive immunity. Nature. 1997;388:394–397. doi: 10.1038/41131. [DOI] [PubMed] [Google Scholar]
- Rock F.L., Hardiman G., Timans J.C., Kastelein R.A., Bazan J.F. A family of human receptors structurally related to Drosophila Toll. Proc. Natl. Acad. Sci. USA. 1998;95:588–593. doi: 10.1073/pnas.95.2.588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poltorak A., Xialong H., Sminova I., Liu M.-Y., Van Huffel C., Du X., Birdwell D., Alejos E., Silva M., Galanos C. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr micemutations in tlr4 gene. Science. 1998;282:2085–2088. doi: 10.1126/science.282.5396.2085. [DOI] [PubMed] [Google Scholar]
- Qureshi S.T., Lariviere L., Leveque G., Clemont S., Moore K.J., Gros P., Malo D. Endotoxin-tolerant mice have mutations in Toll-like receptor 4 (Tlr4) J. Exp. Med. 1999;189:615–625. doi: 10.1084/jem.189.4.615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoshino K., Takeuchi O., Kawai T., Sanjo H., Ogawa T., Takeda Y., Takeda K., Akira S. Toll-like receptor 4 (TLR4)-deficient mice are hyporesponsive to lipopolysaccharideevidence for TLR4 as the lps gene product. J. Immunol. 1999;162:3749–3752. [PubMed] [Google Scholar]
- Shimazu R., Akashi S., Ogata H., Nagai Y., Fukudome K., Miyake K., Kimoto M. MD-2, a molecule that confers lipopolysaccharide responsiveness on Toll-like receptor 4. J. Exp. Med. 1999;189:1777–1782. doi: 10.1084/jem.189.11.1777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akashi S., Shimazu R., Ogata H., Nagai Y., Takeda K., Kimoto M., Miyake K. Cell surface expression and LPS signaling via the Toll-like receptor 4-MD-2 complex on mouse peritoneal macrophages. J. Immunol. 2000;164:3471–3475. doi: 10.4049/jimmunol.164.7.3471. [DOI] [PubMed] [Google Scholar]
- Miyake K., Yamashita Y., Hitoshi Y., Takatsu K., Kimoto M. Murine B cell proliferation and protection from apoptosis with an antibody against a 105-kD moleculeunresponsiveness of X-linked immunodeficient B cells. J. Exp. Med. 1994;180:1217–1224. doi: 10.1084/jem.180.4.1217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyake K., Yamashita Y., Ogata M., Sudo T., Kimoto M. RP105, a novel B cell surface molecule implicated in B cell activation, is a member of the leucine-rich repeat protein family. J. Immunol. 1995;154:3333–3340. [PubMed] [Google Scholar]
- Miura Y., Miyake K., Yamashita Y., Shimazu R., Copeland N.G., Gilbert D.J., Jenkins N.A., Inazawa J., Abe T., Kimoto M. Molecular cloning of a human RP105 homologue and chromosomal localization of the mouse and human RP105 genes (Ly64 and LY64) Genomics. 1996;38:299–304. doi: 10.1006/geno.1996.0632. [DOI] [PubMed] [Google Scholar]
- Miyake K., Shimazu R., Kondo J., Niki T., Akashi S., Ogata H., Yamashita Y., Miura Y., Kimoto M. MD-1, a molecule that is physically associated with RP105 and positively regulates its expression. J. Immunol. 1998;161:1348–1353. [PubMed] [Google Scholar]
- Miura Y., Shimazu R., Miyake K., Akashi S., Ogata H., Yamashita Y., Narisawa Y., Kimoto M. RP105 is associated with MD-1 and transmits an activation signal in human B cells. Blood. 1998;92:2815–2822. [PubMed] [Google Scholar]
- Chan V.W.F., Mecklenbräuker I., Su I., Texido G., Leitges M., Carsetti R., Lowell C.A., Rajewsky K., Miyake K., Tarakhovsky A. The molecular mechanism of B cell activation by toll-like receptor protein RP105. J. Exp. Med. 1998;188:93–101. doi: 10.1084/jem.188.1.93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tarakhovsky A., Muller W., Rajewsky K. Lymphocyte populations and immune responses in CD5-deficient mice. Eur. J. Immunol. 1994;24:1678–1684. doi: 10.1002/eji.1830240733. [DOI] [PubMed] [Google Scholar]
- Texido G., Su I., Mecklenbräuker I., Saijo K., Malek S.N., Desiderio S., Rajewsky K., Tarakhovsky A. The B-cell-specific src-family kinase blk is dispensable for B-cell development and activation. Mol. Cell. Biol. 2000;20:1227–1233. doi: 10.1128/mcb.20.4.1227-1233.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palacios R., Steinmetz M. IL-3 dependent mouse clones that express B220 surface antigen, contain Ig genes in germ-line configuration, and generate B lymphocytes in vivo. Cell. 1985;41:727–734. doi: 10.1016/s0092-8674(85)80053-2. [DOI] [PubMed] [Google Scholar]
- Fujita T., Nolan G.P., Liou H.-C., Scott M.L., Baltimore D. The candidate proto-oncogene bcl-3 encodes a transcriptional coactivator that activates through NF-κB p50 homodimers. Genes Dev. 1993;7:1354–1363. doi: 10.1101/gad.7.7b.1354. [DOI] [PubMed] [Google Scholar]
- Mizushima S., Nagata S. pEF-BOS, a powerful mammalian expression vector. Nucleic Acids Res. 1990;18:5322. doi: 10.1093/nar/18.17.5322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishizumi H., Taniuchi I., Yamanashi Y., Kitamura D., Ilic D., Mori S., Watanabe T., Yamamoto T. Impaired proliferation of peripheral B cells and indication of autoimmune disease in lyn-deficient mice. Immunity. 1995;3:549–560. doi: 10.1016/1074-7613(95)90126-4. [DOI] [PubMed] [Google Scholar]
- Hibbs M.L., Tarlinton D.M., Armes J., Grail D., Hodgson G., Maglitto R., Stacker S.A., Dunn A.R. Multiple defects in the immune system of lyn-deficient mice, culminating in autoimmune disease. Cell. 1995;83:301–311. doi: 10.1016/0092-8674(95)90171-x. [DOI] [PubMed] [Google Scholar]