

Abstract
Carbonic anhydrase XII (CA XII) is a transmembrane glycoprotein with an active extracellular CA domain that is overexpressed on cell surfaces of certain cancers. Its expression has been linked to tumor invasiveness. To characterize its catalytic properties, we purified recombinant secretory forms of wild-type and mutant CA XIIs. The catalytic properties of these enzymes in the hydration of CO2 were measured at steady state by stopped-flow spectrophotometry and at chemical equilibrium by the exchange of 18O between CO2 and water determined by mass spectrometry. The catalysis of CO2 hydration by soluble CA XII has a maximal value of kcat/Km at 34 μM−1⋅s−1, which is similar to those of the membrane-associated CA IV and to soluble CA I. The pH profiles of this catalysis and the catalyzed hydrolysis of 4-nitrophenylacetate indicate that the pKa of the zinc-bound water in CA XII is 7.1. His64 in CA XII acts as a proton shuttle residue, as evidenced by the reduced rate constant for proton transfer in the mutants containing the replacements His64 → Ala and His64 → Arg, as well as by the selective inhibition of the proton transfer step by cupric ions in wild-type CA XII. The catalytic rate of CO2 hydration by the soluble form of CA XII is identical with that of the membrane-bound enzyme. These observations suggest a role for CA XII in CO2/HCO3− homeostasis in cells in which it is normally expressed. They are also compatible with a role for CA XII in acidifying the microenvironment of cancer cells in which CA XII is overexpressed, providing a mechanism for CA XII to augment tumor invasiveness and suggesting CA XII as a potential target for chemotherapeutic agents.
Carbonic anhydrase XII (CA XII) is a recently identified mammalian CA that was initially discovered because its mRNA was overexpressed in renal cell cancers (1) and in a lung cancer cell line (2). Independently, its mRNA was discovered as one of the transcripts subject to regulation by the VHL tumor suppressor gene (3). This observation may explain the up-regulation of CA XII in renal cell cancers, because many renal cell carcinomas show loss of expression of the VHL tumor suppressor (4). That CA expression might be linked to tumor invasiveness was suggested by several authors (1, 5, 6) and supported by recent experiments demonstrating in an in vitro assay that acetazolamide inhibited invasiveness of several CA-expressing tumor cell lines (7).
The possible importance of CA XII in the spread of certain cancers led us to verify that CA XII is catalytically active, to characterize its kinetic properties, and to use site-directed mutagenesis to explore the role of the conserved proton shuttle residue His64 in catalysis (8). Detailed kinetic studies were carried out on secretory forms of wild-type and mutant CA XII isolated from media of cells overexpressing recombinant proteins, and also on the membrane form of wild-type CA XII present in membranes isolated from cells overexpressing the recombinant wild-type protein. These studies show that CA XII is an active isozyme whose catalytic properties resemble those of the membrane-associated isozyme CA IV. These studies make it likely that CA XII actually plays a role in CO2/bicarbonate homeostasis, as opposed to other CA-like proteins that are associated with cancer but use a conserved, although enzymatically inactive, CA domain for other physiological purposes such as ligand binding (9, 10).
Materials and Methods
Construction of Secretory and Mutant cDNAs for Human CA XII.
The full-length cDNA for CA XII in pBluescript (1) was used to generate the Q291X secretory form of CA XII by site-directed mutagenesis. The 260-residue CA domain in the CA XII precursor is preceded by a 29-residue signal sequence that is not present in the mature protein. A cytosine at nucleotide 871 (counting the A in the initiator ATG as +1) was mutated to a thymine (C → T), changing a glutamine (CAA) at position 291 to a stop codon (TAA). The resulting protein would no longer retain the transmembrane and C-terminal cytoplasmic domains. The His64 → Arg mutant (H64R) of Q291X CA XII was made by changing an adenine at nucleotide 281 to a guanine (A → G). The numbering of this conserved His residue is based on the His64 of the 260-residue conserved CA domain. The histidine 64 to alanine (H64A) mutant was made by changing a cytosine at nucleotide 280 to a guanine (C → G) and an adenine at nucleotide 281 to a cytosine (A → C). All mutants were made using a Chameleon double-stranded site-directed mutagenesis kit (Stratagene).
Construction of Mammalian Expression Vector.
The cDNA of the wild-type, full-length membrane form of CA XII and the secretory form (Q291X CA XII) and its H64A and H64R mutant derivatives were digested with EcoRI. The digested fragments were gel purified by Qiaex II kit (Qiagen, Chatsworth, CA) and subcloned into the mammalian expression vector pCXN at the EcoRI site (11).
Development of Stable Clones.
Wild-type, membrane-bound CA XII, Q291X CA XII, and H64R and H64A mutants of Q291X CA XII in pCXN were linearized by ScaI digestion and transfected into CHO-K1 cells by electroporation under conditions of 25 μF and 1,200 V in a 0.4-cm cuvette by using the Bio-Rad electroporation system. After 2–3 weeks with G418 (400 μg/ml), stable clones were picked and grown to confluency. Cell lysates and media were analyzed by Western blot using polyclonal CA XII antibody (1). The CA activity in cell lysates and media was determined by using the endpoint titration assay method of Maren (12, 13). Catalytic and Western blot assays showed that Q291X CA XII and both H64R and H64A mutants of Q291X CA XII stably expressed in CHO-K1 cells were secreted in the medium. The integral membrane form of wild-type CA XII was studied in membrane suspensions from CHO cells expressing wild-type CA XII.
Purification of Soluble HCA XII from Secretion Media.
The Q291X CA XII secretory form of wild-type and His64 mutant of Q291X CA XII were purified from secretion media by using a CA inhibitor affinity column as described (14) with the following modifications. Secretion medium was adjusted to a final concentration of 10 mM Hepes (pH 7.5) and applied to a p-aminomethylbenzenesulfonamide-agarose (Sigma) column. The affinity column was equilibrated with 10 mM Hepes (pH 7.5) buffer. Unbound proteins were removed by washing extensively with 10 mM Hepes (pH 7.5) buffer. The bound enzyme was eluted with 0.1 M sodium acetate (pH 5.6) and 0.5 M sodium perchlorate. Fractions containing enzyme were pooled and concentrated by Centricon-10 units, and concentrated enzyme preparations were dialyzed against 10 mM Tris⋅SO4 (pH 7.4) buffer. Based on SDS/PAGE, the affinity-purified enzyme was more than 90% pure. For antibody production, the affinity-purified enzyme was further purified over an S-300 sizing column (Amersham Pharmacia).
CA XII Antibody Production.
Purified Q291X CA XII (200 μg in 0.5 ml of phosphate buffer saline) was mixed with an equal volume of Freund's complete adjuvant before injection into a rabbit (14). Preimmune serum was collected before antigen injection. After 4 weeks, each rabbit was boosted by injection of 200 μg purified Q291X CA XII in incomplete Freund's adjuvant. Animals were bled 12 days after the second injection. The titer of the serum was checked by Western blot using purified Q291X CA XII enzyme and antibody was detected to dilutions of 1:10,000. Specificity of the antibody for CA XII was documented by Western blot (6).
Enzyme.
The concentration of active sites was determined by titration of the 18O exchange activity of the enzymes with the tight-binding inhibitor ethoxzolamide (Ki = 2 nM for wild-type CA XII).
Oxygen-18 Exchange.
Oxygen-18 exchange was used to measure catalysis by CA XII. The catalyzed and uncatalyzed rates of exchange of 18O from species of CO2 into water and the rates of exchange of 18O between 12C- and 13C-containing species of CO2 were measured at chemical equilibrium by mass spectrometry. Eqs. 1 and 2 demonstrate the catalytic pathway for the exchange of 18O from bicarbonate to water. In Eq. 2, BH is a buffer in solution and/or an amino acid side chain in the enzyme.
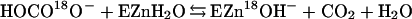 |
1 |
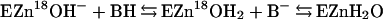 |
2 |
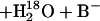 |
This method is capable of determining two rates in the catalytic pathway. The first is R1, the rate of interconversion of CO2 and HCO3− at chemical equilibrium.
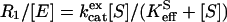 |
3 |
Here [E] is the total enzyme concentration, kcatex is a rate constant for maximal HCO3− to CO2 interconversion, [S] is the substrate concentration of HCO3− and/or CO2, and KeffS is an apparent substrate binding constant (15). This equation, when applied to the data for varying substrate concentration, or to measurement of R1 when [S] ≪ KeffS, can determine the values of kcatex/KeffS. In both theory and practice, kcatex/KeffCO2 is equivalent to kcat/Km for CO2 hydration as measured by steady-state methods (15, 16).
The second rate determined by this method is RH2O, the rate of release from the enzyme of water labeled with 18O, which is shown in Eq. 2. A proton donated from a donor group BH converts the zinc-bound hydroxide to zinc-bound water, which readily exchanges with unlabeled water. The 18O label is essentially infinitely diluted into the solvent H216O. The value of RH2O can be interpreted in terms of the rate constant from a predominant donor group to the zinc-bound hydroxide according to Eq. 4 (17), in which kB is the rate constant for proton transfer to the zinc-bound hydroxide, KB is the ionization constant for the donor group, and KE is the ionization constant of the zinc-bound water molecule.
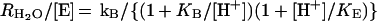 |
4 |
Measurements of the rate of distribution of 18O were made with an Extrel EMX-200 mass spectrometer and a membrane-inlet permeable to dissolved gases (16). Solutions contained 5 μM EDTA (except for measurement of Cu2+ inhibition) and the total ionic strength of solution was maintained at 0.2 M by the addition of Na2SO4. Unless otherwise indicated, experiments were carried out in the absence of buffers, which were not needed to maintain pH because these experiments were carried out at chemical equilibrium.
Steady-State Constants.
A stopped-flow spectrophotometer (model SX.18MV; Applied Photophysics, Surrey, U.K.) was used to measure initial velocities of the hydration of CO2 by recording the absorbance change of a pH indicator (18). Saturated CO2 solutions were made by bubbling CO2 into water at 25°C. Syringes with gas-tight seals were used to make CO2 dilutions from 17 mM to 1.4 mM. The pKa of the buffer indicator pairs and the observed wavelengths were as follows: Mes (pKa 6.1) and chlorophenol red (pKa 6.3, 574 nM), Mops (pKa 7.2) with p-nitrophenol (pKa 7.1, 400 nM), Hepes (pKa 7.5) with phenol red (pKa 7.5, 557 nM), Taps (pKa 8.4) with m-cresol purple (pKa 8.3, 578 nM), and Ches (pKa 9.3) and thymol blue (pKa 8.9, 590 nM). The buffer concentration was 25 mM unless otherwise indicated, and the total ionic strength for each buffer-indicator pair system was maintained at 0.2 M by the addition of the appropriate amount of Na2SO4. The mean of four to eight reaction traces of the first 5–10% of the reaction was used to determine initial rates. The uncatalyzed rates were subtracted, and the rate constants kcat and kcat/Km determined by nonlinear least-squares methods (Enzfitter; Biosoft, Cambridge, U.K.).
Rate of Ester Hydrolysis.
The catalysis of the hydrolysis of 4-nitrophenylacetate was measured by following the increase in absorbance at 348 nM, corresponding to the isosbestic point of nitrophenol and the nitrophenolate ion using the molar absorptivity 5.0 × 103 M−1⋅cm−1 (19). A Beckman Coulter DU650 spectrophotometer was used and initial velocities were determined under the conditions given in the legend to Fig. 3. The value of Km for catalysis was too large to measure because it exceeded the solubility of substrate. As a result, we were limited to observing catalytic rates that were first order in substrates from which we obtained kcat/Km.
Figure 3.

The kinetic constant kcat/Km (M−1⋅s−1) for the hydrolysis of 4-nitrophenylacetate catalyzed by human CA XII. Measurements were made at 25°C using solutions maintained at an ionic strength of 0.2 M using Na2SO4 and containing 5 μM EDTA and 25 mM of one of the buffers listed in the text. The solid line is a least-squares fit with the maximal of kcat/Km at 90 ± 2 and pKa 7.1 ± 0.1.
Results
Expression of Human CA XII Proteins in CHO Cells.
The carbonic anhydrase assays on cell lysates and secretion medium were used to identify CHO clones expressing functionally active transmembrane and secretory forms of wild-type and mutant CA XII. The secretory forms of wild-type and His64 mutants were affinity purified, and their specific activities were initially compared by using the CO2 hydration endpoint titration assay of Maren (12). By this assay, the specific activity of wild-type enzyme was 250–262 units/mg. However, the replacement of His64 by Ala or Arg resulted in reduced specific activities of 55 and 60 eu/mg, respectively. When imidazole was present in the assay mixture, the specific activity of mutant enzymes was increased by 30%, suggesting a role for His64 as a proton acceptor during catalysis (8).
The recombinant CA XII proteins were characterized for their apparent molecular mass by using SDS/PAGE followed by Western blot. The results in Fig. 1 show a major polypeptide with an apparent molecular mass of 44 and a minor band of 43 kDa, for the full-length membrane form of CA XII (lane 1). The doublet of wild-type CA XII is caused by carbohydrate variation, which is similar to recombinant wild-type enzyme expressed in COS cells that was shown to be reduced to a single band of 39 kDa by treatment with PNGase F (1). The apparent molecular masses of the Q291X secretory forms of CA XII (hereafter referred to as soluble forms of CA XII) were 41 and 39 kDa. The broad bands reflect microheterogeneity in carbohydrates in the overexpressed soluble forms of the enzyme.
Figure 1.

Western blot analysis of membrane-associated and soluble forms of human CA XII. Cell extracts expressing wild-type CA XII or purified soluble Q291X CA XII and H64A and H64R derivatives of Q291X CA XII were analyzed by SDS/PAGE followed by Western blot using a polyclonal antibody against human CA XII. The apparent molecular masses of the major polypeptides are indicated in kDa.
Catalytic Properties of Human CA XII.
The pH profile for the rate constant kcat/Km for hydration of CO2 catalyzed by CA XII, determined by stopped-flow spectrophotometry, shows a very efficient enzyme with kcat/Km approaching diffusion control and with the apparent pKa of the zinc-bound water at 7.1 ± 0.1 (Fig. 2). The catalysis by CA XII of the hydrolysis of 4-nitrophenylacetate has a relatively small value of kcat/Km and an apparent pKa of 7.1 ± 0.1 (Fig. 3); this catalysis is inhibited by acetazolamide. This esterase function provides another estimate of the pKa of the zinc-bound water in CA XII.
Figure 2.

The kinetic constant kcat/Km (M−1⋅s−1) for the hydration of CO2 catalyzed by (wild-type) human CA XII. Data were measured by stopped-flow spectrophotometry at 25°C using 25 mM of the buffers listed in the text. The total ionic strength was maintained at 0.2 M by addition of Na2SO4. Solutions also contained 5 μM EDTA. The solid line is a least-squares fit to a single ionization with pKa = 7.1 ± 0.1 and with a maximum of (3.4 ± 0.3) × 107 M−1⋅s−1.
Oxygen-18 exchange catalyzed by three variants of soluble human CA XII: wild type, H64A, and H64R showed maximal values of R1/[E] that were identical within experimental uncertainty (0.11 ± 0.02 μs−1). The rate constant R1/[E] is described in Eq. 3 and contains rate constants for the steps of Eq. 1, the interconversion of CO2 and bicarbonate. The similar values of R1/[E] for these three variants indicate that the chemistry of this catalytic interconversion is not greatly altered by the mutations His64 → Ala and His64 → Arg. For wild type and H64A CA XII, the values of kcat/Km derived from 18O exchange using Eq. 3 had a more complex pH dependence than the values for kcat/Km determined by stopped-flow shown in Fig. 2. This finding suggests that the catalysis of 18O exchange at chemical equilibrium may be subject to other features in the active site such as nearby ionizations or the exchange properties of H218O at the zinc that do not affect the initial velocity.
The pH profile for kcat for hydration of CO2 catalyzed by CA XII can be fit to a single ionization with an apparent pKa at 8.0 ± 0.2 and a maximal value near 4 × 105 s−1 (Fig. 4). This aspect of the catalysis is nearly as fast as the most efficient of the carbonic anhydrase isozymes and its pH profile approximates the pKa of the proton acceptor group. The turnover number that characterizes catalysis by the carbonic anhydrases at steady state is limited in rate by proton transfer (20). The rate constant RH2O/[E] measured at chemical equilibrium is also limited by proton transfer (16) and measures the rate of release from the enzyme of H218O (Eq. 2). The bell-shaped value for wild type (Fig. 5) is characteristic of the efficient isozymes of CA and is consistent with a predominant proton shuttle residue as described by Eq. 4 with the values of constants given in the legend to Fig. 5. The values of RH2O/[E] at pH < 8 are much decreased for the mutants H64A and H64R CA XII (Fig. 5), indicating a role of His64 in catalysis. These data are consistent with His64 acting as a proton shuttle residue in CA XII as in CA II (8).
Figure 4.

The pH dependence of the steady-state turnover number kcat (s−1) for the hydration of CO2 catalyzed by human CA XII determined by stopped-flow spectrophotometry at 25°C. Solutions contained 5 μM EDTA and 25 mM of one of the buffers listed in Materials and Methods. The solid line is a least-squares fit with a maximal value of kcat at (4.0 ± 1.7) × 105 s−1 and an apparent pKa of 8.0 ± 0.2.
Figure 5.

The pH dependence of RH2O/[E] (s−1) catalyzed by human wild-type CA XII (●), H64A CA XII (▵), and H64R CA XII (■). Conditions were as described in Fig. 2. The solid line for wild-type CA XII is a least-squares fit of Eq. 4 to the data yielding (pKa)acceptor = 6.1 ± 0.1; (pKa)donor = 7.5 ± 0.1; and kB = (2.9 ± 0.2) × 105 s−1.
The values of RH2O/[E] are identical for solubilized CA XII and membrane-bound CA XII measured in particulate fractions (Fig. 6). The values of kcat/Km determined by 18O exchange using Eq. 3 were also identical for these two forms of CA XII (data not shown).
Figure 6.

The pH dependence of RH2O/[E] (s−1) catalyzed by human CA XII (●) and membrane-bound CA XII (□). Conditions were as described in Fig. 2.
Discussion
Four active membrane-associated carbonic anhydrases have been described to date. The first described was CA IV, a membrane-associated, glycosylphosphatidylinositol-anchored enzyme expressed on the plasma membrane of lung epithelial cells (21, 22) and endothelial cells of microcapillaries (23). It is a high-activity enzyme that contributes to the high rates of CO2/bicarbonate flux across membranes (14, 24, 25). The second membrane CA described was CA IX, a multidomain protein that is like CA XII in that it also is overexpressed in certain tumors (26–28) and in clear cell renal carcinomas (3). Whether the CA activity of CA IX is required for its transforming ability or the CA domain of CA IX protein might function in a ligand-binding role analogous to the inactive CA domain in RPTPβ and RPTPγ (29) remains to be established. A fourth membrane CA (CA XIV) was described recently. This membrane CA was cloned from mouse kidney by using a signal-sequence trap method. CA XIV is highly expressed in kidney membranes and is a high-activity enzyme, but has not been characterized extensively (30).
CA XII was the third membrane CA described. It was cloned and characterized initially from a renal cell carcinoma cDNA expression library (1) and found to be regulated by the VHL tumor suppressor gene (3). This observation may explain its overexpression in renal cancer cells, which frequently show loss of VHL expression (4). CA XII also was found to be expressed in normal human colon and colorectal tumors (6) and in endometrial epithelium (31). Recently, Parkkila et al. (7) reported that invasiveness of tumor cell lines expressing cytosolic CA II and CA XII was inhibited by the CA inhibitor acetazolamide. These results suggested that CA XII activity, alone or together with CA II, might play a role in tumor invasiveness.
We characterized the kinetic properties of purified, soluble forms of CA XII and showed that the membrane-associated form is equally active in catalysis. We also studied H64A and H64R mutants of the soluble CA XII to determine the role of the conserved His64 in the catalytic mechanism. Such studies provide a framework for interpreting the possible role of CA XII in acidifying the microenvironment of invasive tumors.
The maximal value of kcat/Km describing the catalysis of CO2 hydration by human CA XII is similar to that of another membrane-bound isozyme, CA IV (24, 25), and also similar to CA I; moreover, these values are only slightly lower than those for the most efficient of the CA isozymes, CA II (Table 1). Of the rate constants describing this catalysis at steady state, kcat/Km is the most relevant to the physiologic function of CA XII. CA XII resembles CA IV also in its relatively weak hydrolysis of 4-nitrophenylacetate, a catalysis that is much smaller than that for CA II (Table 1). In terms of maximal velocity or kcat, CA XII most closely resembles CA I (Table 1). This places CA XII among the more efficient of the isozymes in the α class of carbonic anhydrases, in contrast with CA III, which is the least efficient.
Table 1.
Maximal pH-independent values of catalytic constants for wild-type human CA XII as compared with other prominent isozymes of CA
We also measured the catalysis by CA XII of the exchange of 18O between CO2 and water. Here RH2O/[E] describes the release of H218O from the enzyme and is limited in rate by proton transfer to the zinc-bound hydroxide (Eq. 2). The pH profile for RH2O/[E] is bell-shaped (Fig. 5), as observed for CA II (17), and by using Eq. 4, yields a value for the rate constant kB near 3 × 105 s−1 for intramolecular proton transfer (Fig. 5). The high-activity carbonic anhydrase isozymes are efficient in part because of the proton shuttle residue, His64, which transfers protons between the zinc-bound water or hydroxide and buffer in solution (20). His64 in CA XII is consistent with this role as shown in Fig. 5, in which values of RH2O/[E] are decreased by as much as 10-fold by the replacements His64 → Ala and His64 → Arg. This finding is very similar to observations of catalysis by H64A CA II (8). Also consistent with a shuttle role of His64 in CA XII is the selective inhibition of RH2O in wild-type CA XII by cupric ions with an inhibition constant of Ki = 1.9 ± 0.2 μM (data not shown); this inhibition has no effect on the rate of interconversion of CO2 and bicarbonate in the 18O exchange experiment. This result is also very similar to that observed with other isozymes in which His64 acts as a proton shuttle (24).
The pH dependence of kcat for hydration of CO2 is determined by the ionization of the proton acceptor His64 (8, 20); the data of Fig. 4 estimate this pKa near 8 for CA XII. This pKa is also evident in the interpretation according to Eq. 4 of RH2O in Fig. 5 in which the pKa 7.5 is identified as attributable to His64. The pKa near 6.1 determined in Fig. 5 is too low to be assigned only to the ionization of the zinc-bound water, which is estimated near pKa 7 in Figs. 2 and 3. Experimental uncertainties may account for this difference; however, it is possible that RH2O/[E] in this case does not reflect only the ionization of the zinc-bound water but is influenced by other ionizations near the active site or more complex features that determine the exchange of H218O at the zinc. This may be the same feature that added complexity to R1 described in Results.
The 18O exchange method is well suited to measuring catalysis by membrane-bound carbonic anhydrase in particulate fractions. The concentration of active sites is determined by titration with the tight-binding inhibitor ethoxzolamide (Ki = 2 nM). Such experiments show that catalysis by membrane-bound CA XII is identical to that of the solubilized, membrane-free preparation (Fig. 6). This result might be expected for a carbonic anhydrase attached to the membrane by an extension of or an addition to its C-terminal end. This C-terminal end is located on the backside of the enzyme with respect to the active site cavity in CA II (32) and CA IV (33). In any case, the identity in the kinetic properties of the native transmembrane CA XII and secretory form of CA XII indicate that the secretory form of CA XII could provide physiologically relevant data in drug screening assays aimed at identifying isozyme-specific CA inhibitors. Identification of an isozyme-specific inhibitor for CA XII might be a useful adjunct in chemotherapy for cancers that overexpress CA XII on their cell surfaces.
Acknowledgments
This work was supported by National Institutes of Health Grants GM25154 (to D.N.S.) and DK40163, GM34182, and GM53405 (to W.S.S.).
Abbreviation
- CA
carbonic anhydrase
References
- 1.Türeci O, Sahin U, Vollmar E, Siemer S, Göttert E, Seitz G, Parkkila A N, Shah G N, Grubb J H, Pfreundschuh M, et al. Proc Natl Acad Sci USA. 1998;95:7608–7613. doi: 10.1073/pnas.95.13.7608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Torczynski, R. M. & Bollon, A. P. (1996) U.S. Patent 5,589,579.
- 3.Ivanov S V, Kuzmin I, Wei M H, Pack S, Geil L, Johnson B E, Stanbridge E J, Lerman M I. Proc Natl Acad Sci USA. 1998;95:12596–12601. doi: 10.1073/pnas.95.21.12596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Latif F, Tory K, Gnarra J, Yao M, Duh F M, Orcutt M L, Stackhouse T, Kuzmin I, Modi W, Geil L, et al. Science. 1993;260:1317–1320. doi: 10.1126/science.8493574. [DOI] [PubMed] [Google Scholar]
- 5.Chegwidden W R, Spencer I M. Inflammopharmacology. 1995;3:231–239. [Google Scholar]
- 6.Kivelä A, Parkkila S, Saarnio J, Karttunen T J, Kivelä J, Parkkila A K, Waheed A, Sly W S, Grubb J H, Shah G, et al. Am J Pathol. 2000;156:577–584. doi: 10.1016/S0002-9440(10)64762-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Parkkila S, Rajaniemi H, Parkkila A K, Kivelä J, Waheed A, Pastoreková S, Pastorek J, Sly W S. Proc Natl Acad Sci USA. 2000;97:2220–2224. doi: 10.1073/pnas.040554897. . (First Published February 25, 2000; 10.1073/pnas.040554897) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Tu C K, Silverman D N, Forsman C, Jonsson B H, Lindskog S. Biochemistry. 1989;28:7913–7918. doi: 10.1021/bi00445a054. [DOI] [PubMed] [Google Scholar]
- 9.Barnea G, Silvennoinen O, Shaanan B, Honegger A M, Canoll P D, d'Eustachio P, Morse B, Levy J B, LaForgia S, Huebner K, et al. Mol Cell Biol. 1993;13:1497–1506. doi: 10.1128/mcb.13.3.1497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Wary K K, Lou Z W, Buchberg A M, Siracusa L D, Druck T, LaForgia S, Huebner K. Cancer Res. 1993;53:1498–1502. [PubMed] [Google Scholar]
- 11.Niwa H, Yamamura K, Miyazaki J. Gene. 1991;108:193–200. doi: 10.1016/0378-1119(91)90434-d. [DOI] [PubMed] [Google Scholar]
- 12.Maren T H. J Pharmacol Exp Ther. 1960;130:26–29. [PubMed] [Google Scholar]
- 13.Sundaram V, Rumbolo P, Grubb J H, Strisciuglio P, Sly W S. Am J Hum Genet. 1986;38:125–136. [PMC free article] [PubMed] [Google Scholar]
- 14.Zhu X L, Sly W S. J Biol Chem. 1990;265:8795–8801. [PubMed] [Google Scholar]
- 15.Simonsson I, Jonsson B-H, Lindskog S. Eur J Biochem. 1979;93:409–417. doi: 10.1111/j.1432-1033.1979.tb12837.x. [DOI] [PubMed] [Google Scholar]
- 16.Silverman D N. Methods Enzymol. 1982;87:732–752. doi: 10.1016/s0076-6879(82)87037-7. [DOI] [PubMed] [Google Scholar]
- 17.Silverman D N, Tu C K, Chen X, Tanhauser S M, Kresge A J, Laipis P J. Biochemistry. 1993;32:10757–10762. doi: 10.1021/bi00091a029. [DOI] [PubMed] [Google Scholar]
- 18.Khalifah R G. J Biol Chem. 1971;246:2561–2573. [PubMed] [Google Scholar]
- 19.Verpoorte J A, Mehta S, Edsall J T. J Biol Chem. 1967;242:4221–4229. [PubMed] [Google Scholar]
- 20.Lindskog S. Pharmacol Ther. 1997;74:1–20. doi: 10.1016/s0163-7258(96)00198-2. [DOI] [PubMed] [Google Scholar]
- 21.Waheed A, Zhu X L, Sly W S. J Biol Chem. 1992;267:3308–3311. [PubMed] [Google Scholar]
- 22.Fleming R E, Crouch E C, Ruzicka C A, Sly W S. Am J Physiol. 1993;265:L627–L635. doi: 10.1152/ajplung.1993.265.6.L627. [DOI] [PubMed] [Google Scholar]
- 23.Hageman G S, Zhu X L, Waheed A, Sly W S. Proc Natl Acad Sci USA. 1991;88:2716–2720. doi: 10.1073/pnas.88.7.2716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hurt J D, Tu C K, Laipis P J, Silverman D N. J Biol Chem. 1997;272:13512–13518. doi: 10.1074/jbc.272.21.13512. [DOI] [PubMed] [Google Scholar]
- 25.Baird T T, Waheed A, Okuyama T, Sly W S, Fierke C A. Biochemistry. 1997;36:2669–2678. doi: 10.1021/bi962663s. [DOI] [PubMed] [Google Scholar]
- 26.Závada J, Závadová Z, Pastoreková S, Ciampor F, Pastorek J, Zelník V. Int J Cancer. 1993;54:268–274. doi: 10.1002/ijc.2910540218. [DOI] [PubMed] [Google Scholar]
- 27.Saarnio J, Parkkila S, Parkkila A K, Waheed A, Casey M C, Zhou X Y, Pastoreková S, Pastorek J, Karttunen T, Haukipuro K, et al. J Histochem Cytochem. 1998;46:497–504. doi: 10.1177/002215549804600409. [DOI] [PubMed] [Google Scholar]
- 28.Liao S Y, Stanbridge E J. Cancer (Philadelphia) 2000;88:1108–1121. doi: 10.1002/(sici)1097-0142(20000301)88:5<1108::aid-cncr23>3.0.co;2-d. [DOI] [PubMed] [Google Scholar]
- 29.Peles E, Nativ M, Campbell P L, Sakurai T, Martinez R, Lev S, Clary D O, Schilling J, Barnea G, Plowman G D, et al. Cell. 1995;82:251–260. doi: 10.1016/0092-8674(95)90312-7. [DOI] [PubMed] [Google Scholar]
- 30.Mori K, Ogawa Y, Eibihara K, Tamura N, Tashiro K, Kuwahara T, Mukoyama M, Sugawara A, Ozaki S, Tanaka I, et al. J Biol Chem. 1999;274:15701–15705. doi: 10.1074/jbc.274.22.15701. [DOI] [PubMed] [Google Scholar]
- 31.Karhumaa P, Parkkila S, Türeci O, Waheed A, Grubb J H, Shah G, Parkkila A K, Kaunisto K, Tapanainen J, Sly W S, et al. Mol Hum Reprod. 2000;6:68–74. doi: 10.1093/molehr/6.1.68. [DOI] [PubMed] [Google Scholar]
- 32.Eriksson A E, Jones T A, Liljas A. Proteins Struct Funct Genet. 1988;4:274–282. doi: 10.1002/prot.340040406. [DOI] [PubMed] [Google Scholar]
- 33.Stams T, Chen Y, Boriack-Sjodin P A, Hurt J D, Liao J, May J A, Dean T, Laipis P J, Silverman D N, Christianson D W. Protein Sci. 1998;7:556–563. doi: 10.1002/pro.5560070303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Engstrand C, Jonsson B-H, Lindskog S. Eur J Biochem. 1995;229:696–702. doi: 10.1111/j.1432-1033.1995.tb20516.x. [DOI] [PubMed] [Google Scholar]
- 35.Krebs J F, Rana F, Dluhy R A, Fierke C A. Biochemistry. 1993;32:4496–4505. doi: 10.1021/bi00068a004. [DOI] [PubMed] [Google Scholar]
- 36.Jewell D A, Tu C, Paranawithana S R, Tanhauser S M, Lograsso P V, Laipis P J, Silverman D N. Biochemistry. 1991;30:1484–1490. doi: 10.1021/bi00220a006. [DOI] [PubMed] [Google Scholar]
- 37.Tu C K, Thomas H G, Wynns G C, Silverman D N. J Biol Chem. 1986;261:10100–10103. [PubMed] [Google Scholar]